21
Strasburger’s Plant Sciences

Strasburger’s Plant Sciences

Strasburgeria robusta Guill.; Strasburgeriaceae named after the founder of this book, Eduard Strasburger
© Pete Lowry, Missouri Botanical Garden

Andreas Bresinsky, Christian Korner, Joachim W. Kadereit,Gunther Neuhaus and Uwe Sonnewald
Strasburger’s Plant Sciences
Including Prokaryotes and Fungi
With 1100 Figures and 63 Tables

Andreas BresinskyBotanical InstituteUniversity of RegensburgRegensburg, Germany
Christian KornerInstitute of BotanyUniversity of BaselBasel, Switzerland
Joachim W. KadereitInstitut fur Spezielle Botanik und Botanischer GartenJohannes Gutenberg-University MainzMainz, Germany
Gunther NeuhausCell BiologyUniversity of FreiburgFreiburg, Germany
Uwe SonnewaldDepartment of BiologyDivision of BiochemistryFriedrich-Alexander-University Erlangen-NurembergErlangen, Germany
Translation and CopyeditingAlison Davies, Stuart Evans (Chapters 1–4, 9, 10)David and Gudrun Lawlor, Stuart Evans (Chapters 5–8)Christian Korner, Stuart Evans (Chapter 11)Christian Korner, Lea Streule (Chapters 12–14)
Alison Davies, Garching, GermanyDavid and Gudrun Lawlor, Harpenden, UKStuart Evans, West Rainton, UKLea Streule, Basel, Switzerland
ISBN 978-3-642-15517-8 ISBN 978-3-642-15518-5 (eBook)ISBN 978-3-642-15519-2 (print and electronic bundle)DOI 10.1007/978-3-642-15518-5Springer Heidelberg New York Dordrecht London
This work is based on the 36th German language edition of Strasburger, Lehrbuch der Botanik, by Andreas Bresinsky, Christian Korner,Joachim Kadereit, Gunther Neuhaus, Uwe Sonnewald, published by Spektrum Akademischer Verlag, Heidelberg 2008.
Library of Congress Control Number: 2013944576
© Springer-Verlag Berlin Heidelberg 2013
This work is subject to copyright. All rights are reserved by the Publisher, whether the whole or part of the material is concerned, specifically therights of translation, reprinting, reuse of illustrations, recitation, broadcasting, reproduction on microfilms or in any other physical way, andtransmission or information storage and retrieval, electronic adaptation, computer software, or by similar or dissimilarmethodology now knownor hereafter developed. Exempted from this legal reservation are brief excerpts in connection with reviews or scholarly analysis or materialsupplied specifically for the purpose of being entered and executed on a computer system, for exclusive use by the purchaser of the work.Duplication of this publication or parts thereof is permitted only under the provisions of the Copyright Law of the Publisher’s location, in itscurrent version, and permission for use must always be obtained from Springer. Permissions for use may be obtained through RightsLink at theCopyright Clearance Center. Violations are liable to prosecution under the respective Copyright Law.The use of general descriptive names, registered names, trademarks, servicemarks, etc. in this publication does not imply, even in the absence ofa specific statement, that such names are exempt from the relevant protective laws and regulations and therefore free for general use.While the advice and information in this book are believed to be true and accurate at the date of publication, neither the authors nor the editorsnor the publisher can accept any legal responsibility for any errors or omissions that may be made. The publisher makes no warranty, express orimplied, with respect to the material contained herein.
Printed on acid-free paper
Springer is part of Springer ScienceþBusiness Media (www.springer.com)

Preface
Eduard Strasburger*February 1, 1844, Warsaw – {May 19, 1912, Bonn
Founder of the Lehrbuch der Botanik fur Hochschulen
(Botany Textbook for Universities)
(Photo by Dr. Wolfram Lobin/Uni Bonn)
The last English translation of Strasburger’s Lehrbuch der Botanik fur Hochschulen (Textbook of Botany for Universities)
was published in 1976 (30th Ed.). Since then, six new German editions have been published and were partially translated
into Italian, Spanish, Serbo-Croatian, Turkish, and Russian. Considering that plant sciences have developed and
expanded considerably since 1976, and that six more German editions have tried to keep pace with these changes,
a new English translation was long overdue. The present edition represents a balanced and comprehensive work on the
plant sciences, the book’s trademark and particular strength. The inclusion of bacteria, archaea, and the various lineages
referred to as fungi may not be justified from a phylogenetic perspective when dealing with plants, but is necessary
considering the important evolutionary and ecological interactions between plants and these organisms.
Strasburger’s Lehrbuch der Botanik fur Hochschulen has been available for almost 120 years now. Starting with its first
edition in 1894, the book has greatly influenced university teaching in Germany and neighboring countries, and its 36
editions also mirror the dynamic history of the plant sciences.The book was first founded by Eduard Strasburger and is
still published under his name. From the beginning, it was a multi-author effort, and Strasburger himself invited his
colleagues at the Botanical Institute of Bonn University as contributors to the first edition. Since that time more than 20
authors from a number of universities in three different countries contributed to the content. Although clearly all authors
of the first and of later editions shaped the book, Strasburger as its founder deserves special recognition. In his honor,
a New Caledonian tree, which is shown on page II, was named Strasburgeria.
Eduard Strasburger studied the natural sciences in Paris, Bonn, and Jena, receiving his doctorate in Jena before
completing his postdoctoral degree (‘‘Habilitation’’) in Warsaw in 1867. He was appointed professor of botany at the
University of Jena in 1869, at the age of 25, andmoved to BonnUniversity in 1881. Under his direction, the Botanical Institute
at Poppelsdorf Palace established itself as an international center of botany. In 1894, together with his colleagues F. Noll, H.
Schenck, andA.F.W. Schimper, he founded the Lehrbuch der Botanik fur Hochschulen, in the past often simply referred to as
the Bonner Lehrbuch. The Kleine Botanische Praktikum fur Anfanger (Short Botanical Practical for Beginners), which also
appeared in multiple editions, and the somewhat more extensive Das Botanische Praktikum (Botanical Practical) have
dominated microscopical laboratory work at universities for a long time. Strasburger’s research interests were primarily
in plant ontogeny and cytology. He discovered that the central processes underlying nuclear division (formation,
division, and movement of chromosomes) are the same in all eukaryotic organisms (1875), and he was the first to
observe that fertilization in flowering plants requires the fusion of the male sperm nucleus with the female egg nucleus.
From this he concluded that the cell nucleus must be the most important carrier of hereditary factors (1884).
The Authors
April 2013


Table of Contents
Preface . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . v
List of Topical Insights . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . ix
List of Boxes . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . xi
Volume 1
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1
Part I Structure . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11Gunther Neuhaus
1 Molecular Basics: The Building Blocks of Cells . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13
2 The Structure and Ultrastructure of the Cell . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 39
3 The Tissues of Vascular Plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 129
4 Morphology and Anatomy of Vascular Plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 161
Part II Physiology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 237Uwe Sonnewald
5 Physiology of Metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 239
6 Physiology of Development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 411
7 Physiology of Movement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 531
8 Allelophysiology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 569
Volume 2
Part III Evolution and Systematics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 607Joachim W. Kadereit . Andreas Bresinsky
9 Evolution . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 609
10 Systematics and Phylogeny . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 665

Part IV Ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1041Christian Korner
11 Basics of Plant Ecology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1043
12 Plant–Environment Interactions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1065
13 Ecology of Populations and Vegetation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1167
14 Vegetation of the Earth . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1217
Timeline . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1263
Sources . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1267
Index . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1273
viii Table of Contents

List of Topical Insights
Topical Insight 5.1: Galactolipids and Membrane Remodeling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 370
Christoph Benning
Topical Insight 5.2: Genetically Encoded Biosensors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 407
Wolf B. Frommer
Topical Insight 8.1: Host Targets of Bacterial Effectors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 598
Mary Beth Mudgett
Topical Insight 9.1: Homoploid Hybrid Speciation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 656
Loren Rieseberg
Topical Insight 10.1: Origin and Early Evolution of Flowers . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1014
Peter K. Endress . James A. Doyle
Topical Insight 12.1: What Plant Ecologists Can and Cannot Learn from a Satellite’s Eye . . . . . . . . . . 1074
Hamlyn G. Jones
Topical Insight 12.2: A World Without Fire . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1082
William Bond
Topical Insight 12.3: The Dynamic Pipeline: Coordination of Xylem Safety and Efficiency . . . . . . . . . 1093
Frederick C. Meinzer
Topical Insight 12.4: From Where Do Plants Take Their Water? . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1096
Todd E. Dawson
Topical Insight 12.5: Leaf Nitrogen: A Key to Photosynthetic Performance . . . . . . . . . . . . . . . . . . . . 1107
John R. Evans
Topical Insight 12.6: Plant Life in the P-Poor Part of the World . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1114
Hans Lambers
Topical Insight 12.7: Diversity of Traits: A Functional Link to Adaptation, Community Assembly,
and Ecosystem Structure and Function . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1131
Peter B. Reich
Topical Insight 12.8: Using Stable 13C Isotopes to Study Carbon and Water Relations . . . . . . . . . . . . 1137
Rolf Siegwolf
Topical Insight 13.1: Forest Structure and Gap Models . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1202
Hank H. Shugart


List of Boxes
Box 2.1: Cell Fractionation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 42
Box 2.2: The Nuclear Spindle . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 71
Box 3.1: Residual Meristems and Meristemoids . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 131
Box 4.1: Inflorescence Morphology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 183
Box 4.2: Types of Stele . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 195
Box 4.3: The Leaves of Carnivorous Plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 218
Box 4.4: Root Metamorphoses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 229
Box 5.1: Electrophysiology Methods . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 272
Box 5.2: Important Units in Photobiology . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 342
Box 6.1: Thale Cress: Arabidopsis thaliana (L.) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 418
Box 6.2: Conventions in Naming Genes, Proteins, and Phenotypes . . . . . . . . . . . . . . . . . . . . . . . . . 422
Box 6.3: Production of Transgenic Plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 423
Box 6.4: Application of Transgenic Plants . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 429
Box 6.5: Evolution of Plant Receptors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 528
Box 8.1: Cauliflower Mosaic Virus . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 589
Box 8.2: Biology of Crown Gall Tumors . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 593
Box 9.1: Recording and Analyzing Phenotypic and Genetic Variation . . . . . . . . . . . . . . . . . . . . . . . . 628
Box 9.2: Population Genetics . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 641
Box 10.1: The Origin of Life . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 676
Box 10.2: Phylogeny of Plants and Fungi . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 678
Box 10.3: From Unicellular Organisms to Multicellular Organisms . . . . . . . . . . . . . . . . . . . . . . . . . . . 695
Box 10.4: Occurence and Habit of Fungi (Including the Cellulose Fungi) . . . . . . . . . . . . . . . . . . . . . . 748
Box 10.5: Uses of Algae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 775
Box 10.6: Occurence and Diversity of Habits in Algae . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 779
Box 10.7: Occurence and Ecology of Mosses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 816
Box 10.8: Occurence and Ecology of Ferns and Fern Allies . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 874
Box 10.9: Seed Plants (Spermatophytina) . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 880
Box 10.10: Poales: The Evolution of Habitat Ecology and Pollination Biology . . . . . . . . . . . . . . . . . . . 949
Box 10.11: Chenopodiaceae: The Evolution of C4 Photosynthesis . . . . . . . . . . . . . . . . . . . . . . . . . . . . 961
Box 10.12: Asterales: Evolution of Secondary Pollen Presentation . . . . . . . . . . . . . . . . . . . . . . . . . . . 1007
Box 10.13: Mass Extinctions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1018
Box 11.1: Classification of Soils . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1061
Box 12.1: Effects of CO2 on Plant Growth . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1151
Box 13.1: Metapopulations: Consequences of Habitat Fragmentation for Survival of Species . . . . . 1173

Introduction
Botany: A Biological Science
Botany is the science of plants. The term was coined in the
first century by Dioscorides, who used it to mean
a (medicinal) herbal science. In fact, Greek botane means
‘‘grass,’’ as a common forage or economic plant. The
general Greek term for plant is phyton. These days it is
much more common to use the synonymous term ‘‘plant
science’’ than to use ‘‘botany.’’
Plants are primarily defined as those organisms whose
cells contain plastids as well as having true nuclei with
a nuclear membrane and several chromosomes. Plastids
may occur as chloroplasts, or organelles that may become
plastids under the right conditions. Chloroplasts are pho-
tosynthetic organelles that are able to convert light energy
into chemical energy and to fix carbon dioxide. Green
plants are photoautotrophic. Unlike other heterotrophic
(organotrophic) organisms, green plants are able to
survive without organic nutrition.
Fungi are also traditionally included in botany even
though they do not have any plastids. They are heterotro-
phic and behave saprotrophically (feed off dead organic
material), parasitically, or symbiotically (feed off living
organisms). Even though fungi are phylogenetically closer
to animals, they share some features with plants, e.g., they
possess vacuoles in their cell-wall-bound cells, they have a
sessile life style and they take up dissolved nutrients. Fungi
can also form practically obligate symbiotic relationships
with plants (mycorrhiza).
It can be rather problematic to differentiate between
animal and plant among the single-celled protists. Among
the flagellates, even between closely related species in the
same genus, there can be forms with and without plastids:
phytoflagellates and zooflagellates, respectively. The cells
of bacteria and archaea are generally smaller and funda-
mentally more simply organized than the cells of animals,
fungi, and plants (> Fig. 1). Bacteria and archaea do not
have a true cell nucleus and do not undergo cell multipli-
cation by nuclear or cellular division in the way that all
other organisms do, nor do their phototrophic forms
have plastids. The cells in these groups are distinguished
as prokaryotic cells from the eukaryotic cells of all
other organisms. Bacteria and archaea are thus prokary-
otes, whereas all other organisms (plants, fungi, animals;
all protists with a true cell nucleus) are eukaryotes.
There are no intermediate forms between the prokary-
otes and the eukaryotes in modern living organisms.
Even so, the oldest eukaryotes were derived from the
prokaryotes. The investigation of microscopically small
organisms, both prokaryotic as well as eukaryotic, is
a scientific discipline of its own – microbiology. This
includes viruses, viroids, and prions – subcellular systems
that hover at the boundary between the animate and
the inanimate.
Despite all the differences between prokaryotic cells
and eukaryotic cells, and the even more pronounced
differences between the various forms and functions of
the cells of higher animals and plants, there are many basic
commonalities. All organisms share similar molecules and
many fundamental systems essential to life. This also
applies for genes (hereditary factors). This basic unifor-
mity across all life forms indicates a shared phylogenetic
origin: all living organisms (probably) arose from a single
lineage (monophyletic origin).
What Is Life?
Every living system is defined by a particular series of
features. However, only all of these features together
allow the differentiation of an animate from an inanimate
organization. The classic signs of life include:
● Chemical composition. The dry mass of all organisms
is dominated by proteins, nucleic acids, polysaccha-
rides, and lipids. Additionally, there is a wealth of
heterogeneous organic molecules and ions. Organic
molecules, especially macromolecules, are only syn-
thesized by living organisms (biosynthesis with the
help of special catalysts, the enzymes).
● Systematically constructed complex structures. Life
is intrinsically linked to cellular organizational forms.
Even the simplest living organisms are characterized
by complex structures. This means the molecular and
supramolecular components are functionally linked
and dependent on each other. Only by functioning
together properly are they able to bring something to
life. None of the single components alone would be
able to fulfill this. The system is thus more than just
the sum of the parts, and life is always a product of
A. Bresinsky et al., Strasburger’s Plant Sciences, DOI 10.1007/978-3-642-15518-5,# Springer-Verlag Berlin Heidelberg 2013

a system. Below the complexity level of the cell there
is no independent life. The cells always contain
information-bearing structures, an array of various
enzymes, and are separated from their environment
by a selectively permeable membrane. It is not contra-
dictory to say that in most multicellular plants there
are plasmodesma (plasma canals in the cell walls)
between the tissue cells that are united into
a supercellular symplast.
● Nutrition. Organisms are rather ‘‘unlikely’’ construc-
tions in terms of energy and entropy. They are made
up of energy-rich, highly unstable molecules; their
high structural and functional organization represents
low entropy. The support of this labile condition is
only possible with the input of energy. Living systems
are therefore basically open systems; i.e., they take up
energy-rich photons or materials and release energy-
poor material (e.g., CO2, H2O). This metabolism is
intrinsically linked with energy exchange. The metab-
olism results in a constant energy imbalance (dynamic
balance with irreversible subprocesses: so-called flux
equilibrium). Metabolism and energy exchange allow
the energy-demanding construction of (macro)
molecules (anabolism) by linking it to an energy-
producing process such as the capture of solar energy
and/or the breaking up of energy-rich compounds
(catabolism). The low entropy capacity of the organ-
isms is sustained by the donation (dissipation) of
excessive entropy into the surrounding environment.
By using a dissipative structure, the organisms avoid
fatal chaotic events. Thus, life is not really a condition
but is rather a continuous process. Whereas the outer
form of organisms changes rather slowly, there is con-
tinuous turnover at the molecular level.
● Motion. Every actively living organism and every indi-
vidual cell shows signs of motion (motility). However,
many cells/organisms are able to switch to a latent
phase, forming seeds, spores, or cysts. During these
stages of life there are no obvious signs of motility and
almost all criteria for life are arrested.
● Stimulus perception and response.All organisms and
cells must be able to receive and respond to signals
from their environment. The diversity of mechanisms
evolved to do this is incredible.
● Development. Organisms are incapable of retaining
a particular structure indefinitely. No organism looks
the same throughout all its life phases. A newly formed
cell, arising from cell division, grows to the size of its
mother cell (growth). Multicellular organisms usually
start their individual development from just a single
cell (fertilized egg cell, a zygote; spore). Then they
grow by cell division until they reach their final size,
changing their shape in the process. Ontogeny, the
development to a sexually mature multicellular organ-
ism, is associated with morphological processes at the
cellular level that result in the differentiation of the
initially similar embryonic cells.
● Reproduction. The succession of generations is made
up of successive life or reproductive cycles. Life is
perpetuated in this way, in spite of the inability of
individuals to permanently retain a particular devel-
opmental phase and despite the inescapable fact that
all individuals must eventually die. Death is the last
stage in an individual’s development. Unlike ‘‘cata-
strophic death,’’ physiological death is often a result
of inner processes undergoing a program of self-
destruction. Conversely, organisms may only arise as
progeny of conspecific ancestors. Abiogenesis, or
. Fig. 1
Size comparison of prokaryotic cells and eukaryotic cells. (a) Bacterial cells (Escherichia coli). (b) Cells of an Elodea canadensis
leaf. Three plant cell characteristics can be seen: cell walls, chloroplasts, and central vacuoles. Both images are highly
magnified (�380)
2 Introduction

spontaneous generation, of a living system from inan-
imate material is, at least on today’s Earth, inconceiv-
able and has never been proven: omne vivum e vivo
(‘‘every life originates from another life’’). This rather
obvious standpoint is relatively new. Until the ground-
breaking work of L. Pasteur and H. Hoffmann around
1860, it was assumed that microorganisms, even fungi
and nematodes (worms) in fermenting and rotting
liquids, had arisen spontaneously.
● Replication. Reproduction is normally connected
with replication. This ensures the perpetuation of
a species in spite of the loss of individuals as a result
of changing environments. The replication rates are
often astounding in smaller organisms. Under optimal
conditions, bacterial cells can divide every 20 min.
This means that with unrestricted replication of
a single cell, its progeny would form a cell mass the
volume of Earth in less than 2 days. Larger organisms
tend to replicate more slowly, but the individuals are
better protected by a variety of different mechanisms.
● Inheritance. Ontogeny happens in much the same
way from generation to generation. The genetic infor-
mation is amplified and transmitted in the process. It
contains the program for the course of species-specific
ontogeny. The genetic information of all cellular
organisms – prokaryotes and eukaryotes – is saved
(stored) in the bases and nucleotide sequences of
deoxyribonucleic acid (DNA). These are linear or cir-
cular double-stranded macromolecules. Viruses can
store their genetic information in a single-stranded
DNA molecule and in ribonucleic acid (RNA; single
stranded or double stranded).
● Evolution. Copying (replication) and transmission of
the genetic information happens with great precision.
However, over many successive generations, changes
can occur that may be inherited (mutations). These
changes can be induced by environmental factors.
These can be partly a result of inherited switching on
(activation) and off (deactivation) of genes (epige-
netics). In the long term, quite big differences can
develop in a population that can differently affect
the reproductive ability of individuals. This natural
selection results in changes in the characteristics of
the members of a species and in the end can result
in the establishment of new species: evolution
and phylogeny.
A superior criterion for life in all organisms is their
reproductive ability. All remaining characteristics are
either critical to or a result of this central attribute.
The genetic information of all organisms contains the
developmental plan for complex molecular machinery,
whose prime function is its own reproduction. Life is (at
least on today’s Earth) only conceivable and verifiable as
a continuum. This knowledge is supported by the irre-
versibility of individual death and the extinction of
species. There is nothing comparable in inanimate nature.
Origin and Evolution of Life
The living organisms that exist today are the result of
a long evolutionary process. On the basis of radioactivity
and the composition of rock formations, the age of Earth
has been calculated as being about 4.6 billion years. The
study of the remains of organisms (fossils; paleontology)
in various old sediments has shown that other sorts of
plants and animals lived on Earth during earlier geological
epochs. The phylogenetic continuity can be seen in the
floras and faunas of past epochs of the living organisms:
the older they are the more different they are. Larger,
multicellular organisms first appeared toward the end of
the Precambrian (about 570 million years ago). Until then
single-celled organisms had dominated, and these were
mostly prokaryotes. There is evidence that extensive
colonies of cyanobacteria were already present in the
Archean (more than 3 billion years ago): the relevant
sediments in Australia and South Africa contain layered
stromatoliths over 30 cm in size. These are characteristic
biogenic sediments, which are still formed today in warm
waters, and were built by dense layers of phototrophic
cyanobacteria.
How could life have arisen? Answers to this funda-
mental question are sought by trying to recreate or simu-
late the primeval conditions that would have existed on
Earth at that time. A condition for the formation of a
simple self-replicating system was the presence of organic
(macro) molecules. In contrast to today, the conditions on
the still hot planet (Hadean eon) would have enabled
organic molecules to form abiogenically. The first atmo-
sphere contained water vapor as well as carbon dioxide,
nitrogen, and smaller fractions of reducing gasses, but
practically no free oxygen; therefore, there was no ozone
layer that could have filtered the energy-rich UV radiation
from the sun. These conditions would have enabled vari-
ous organic compounds to form. Abiogenic acetic acids
and energy-rich thioesters are even formed in watery mix-
tures of carbon monoxide, sulfuric acid, and metal
sulfides, like those thrown out by deep-sea thermal vents.
Certain places on primitive Earth would have become
enriched in such compounds as long as life did not exist
to digest them and no oxidation destroyed them.
Introduction 3

Even the simplest cells, such as those of the (recently
arisen) saprobiotic mycoplasmas (see below), are very
complex. Their origin from a chaotic mixture of molecular
building blocks via a single chance event is highly improb-
able. However, a likely scenario is that this happened in
a process of hypothetical intermediate steps (multistep
theory): if the necessary individual steps in this prebiotic
evolution were small enough, then the likelihood of them
having really happened over a vast timescale is sufficiently
large. Some molecules which could have arisen
abiogenetically show signs of enzymatic activity; i.e., they
function as biocatalysts. Certain RNA molecules
(ribozymes) can catalyze changes in themselves and,
together with heavy metal ions, can even initiate their
own propagation, albeit rather haphazardly (RNA
world). The decisive step toward independent life was
made when protein catalysts made the effective and pre-
cise replication of nucleic acids possible and the key to the
synthesis of these enzyme proteins was carried by the
nucleic acids. This double-step advance, which was prob-
ably a cumulative result of many small steps, formed a link
between proteins and nucleic acids that is absolutely fun-
damental for life in its current form. Thus, there was
a genetic code that could translate nucleotide sequences
from nucleic acids into protein sequences, and the sepa-
ration of gene (hereditary factor) from phene (a character
based on the hereditary information) was completed.
The first systems capable of self-replication, the hypo-
thetical progenotes and the subsequently evolved pro-
karyotes, were able to live organotrophically as long as
the abiotic formation of organic molecules continued.
However, increasing exploitation of resources to the
point where they became exhausted meant that
phototrophic forms became more prominent. Among
these were some forms that were able to split water to
release oxygen during photosynthesis. This slowly created
an oxidative atmosphere, allowing a much more effective
energy acquisition from organic molecules by cell respira-
tion. At the same time, an ozone layer was formed in the
stratosphere that absorbed the heavily mutagenic UV
radiation from the sun and enabled the colonization of
the ocean surfaces and the land. Fossil evidence from the
long Precambrian evolution is, not surprisingly, rather
rare and incomplete. However, sequence comparisons
from proteins and nucleic acids of related living organisms
can be used to reconstruct phylogeny. The more differen-
tiated the sequences, the earlier the last common ancestor
of the organisms must have lived. Evolutionary changes
have occurred at different rates in different parts of the
(partial) sequences. Therefore, only sequences (or partial
sequences) that change very slowly over time and are fairly
similar even between living, quite distantly related organ-
isms, are used in the reconstruction of early phylogeny.
The comparison of these highly conserved sequences
shows that the split between archaea and bacteria hap-
pened more than 3 billion years ago. Modern eukaryotic
cells have plastids and mitochondria, photosynthetic and
cell-respiration organelles, their own genetic code, and
synthesize some of their proteins themselves. These organ-
elles can only self-replicate and thus have a semiautono-
mous position in eukaryotic cells. They also have
numerous prokaryotic properties, such as the mode of
division, and details of their composition. Plastids seem
to be descendants of once-free-living bacteria, which
became integrated into the cells of primitive eukaryotes
as intracellular symbionts more than one billion years
ago and gradually developed into cell organelles
(Endosymbiont theory).
Remains of multicellular macroorganisms are first
found in sediments that are less than a billion years old.
These organisms are, without exception, eukaryotes.
Even their evolution, which can be increasingly better
reconstructed with molecular systematic techniques, has
been a result of the interaction between chance mutations
and directional selection (Darwinism). This is based on
the assumption that evolution is a result of the sum of
numerous small steps (gradualism). Even so, these have
been interspersed by major evolutionary transitions.
These do not differ from the small steps in terms of how
they arise, but differ rather in the gross effect of many
gradual evolutionary changes. They have been rarer events
than the other gradual evolutionary transitions but have
been more momentous. It seems that, repeatedly, repro-
ductive units that achieved independence at a certain
point in time have merged to form large, more complex
units. Thus, completely novel systems have emerged that
can form the basis of alternative, distinct lineages.
Limits of Life
The question for the limits of life has two components.
First, one can ask for the distributional limits of life, and
second for both lower and upper size limits of individuals.
The first aspect – an ecological component – is that,
despite a phenomenal range of adaptive strategies, general
conditions for life have quite narrow limits. They are
determined by maxima and minima of water content,
temperature, and light. The optimum for most organisms
is median temperatures (10–40�C) and high water
content. For this reason, it is possible to store food at
cool temperatures (fridge, freezer) or by drying
4 Introduction

(legumes, cereals, flour, bread, hay) or by pasteurization
(milk). In nature, the dry and cold regions are particularly
poorly colonized. Many organisms can survive tempera-
tures down to the freezing point of water by having latent
or dormant phases, but still die between 0�C and �10�C.Psychrophilic organisms (e.g., some snow algae) have
optimal growth temperatures between 1 and 2�C. Tem-
peratures over 100�C, which are rarely found on Earth’s
surface (hot springs, volcanoes), can support thermo-
philic organisms. Some archaea have temperature optima
around 100�C, possibly an adaptive relict from primeval
Earth. As phototrophic organisms are mainly responsible
for the production of organic material (biomass), life is
more-or-less restricted to the well-illuminated regions of
Earth’s surface and oceans. Earth is coated with
a comparatively thin biosphere that accounts for less
than 0.01% of its volume.
The largest life forms (both fossil and living) are found
among the vertebrates (dinosaurs, baleen whales) but also
larger and in greater numbers among the conifers and
deciduous trees as well as among clonal organisms such
as poplar (Populus), reed grass (Phragmites), bracken
(Pteridium), and fungi. The giants among the trees
(Sequoia, Cryptomeria, some Eucalyptus) are also the
heaviest life forms.
A more significant question for theoretical biology is
‘‘how small can a life form be?’’ ‘‘What is the lower limit of
complexity for self-replicating biosystems?’’ The smallest
cells are prokaryotic. They are found in mycoplasmas. The
diameter of these cell-wall-less prokaryotic cells is about
0.3 mm and their DNA can only code for about 500
different proteins. This is about the absolute minimum
possible for DNA replication, the realization of the genetic
information stored therein, the support of a heterotrophic
metabolism and energy exchange, and a simple cell struc-
ture (theoretically about 350 genes). In comparison, the
cells of a typical bacterium have a diameter of 2 mm and
contain over 3,000 different proteins; the diameter of most
eukaryotic cells lies between 10 and 100 mm, and the cells
can form over 30,000 different proteins. The complete
sequenced genome of themodel plantArabidopsis thaliana
has about 25,000 genes, 11,000 more than the fruit fly
Drosophila.
Viruses are much more simply organized and most of
them are even smaller. A virion (a viral particle) is not
a cell. Whereas, e.g., the simplest cell has both DNA
(information storage) and RNA (information retrieval),
a virion has neither DNA nor RNA. The nucleic acid is
often only associated with molecules of a single protein
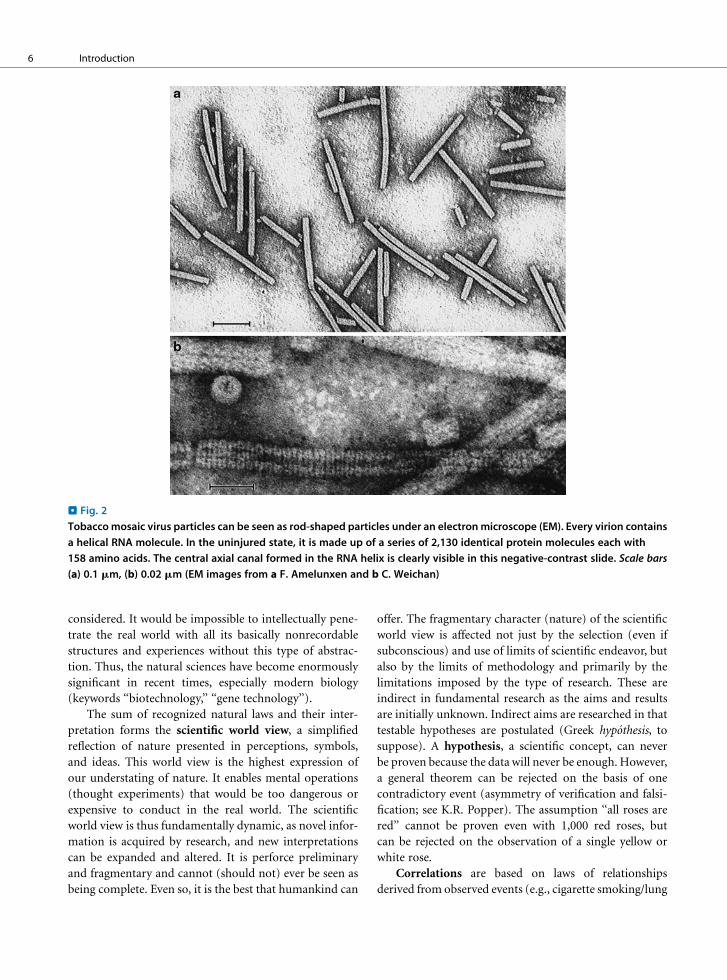
type such as in the tobacco mosaic virus (> Fig. 2), or it
may be surrounded by a protein sheath (capsid) made up
of a single protein or only a few different proteins. This
sort of capsid often has a crystalline symmetry.
Viruses or (bacterio)phages (viruses that attack pro-
karyotic cells) only partially fulfill the conditions neces-
sary for life. They have nometabolism or energy exchange,
no ability to replicate or synthesize proteins, and no ability
to reproduce independently. They can only reproduce by
using the metabolism and energy exchange of a living cell
– they are obligate parasites (‘‘borrowed life’’). The dis-
persal forms – virions – that exist outside living cells
represent lifeless organic systems.
The simplest organizational level is achieved by the
viroids. These are infectious nucleic acids (RNA) with no
associated proteins. The short, ring-shaped RNA mole-
cules do not code for any proteins. Some of the most
dangerous plant parasites known are viroids.
In spite of their particularly simple organization, neither
viruses nor viroids can be considered to be the most prim-
itive forms of life, as their reproduction depends on the
existence of living cells. Rather, they are genetic elements
that became independent of their support cells (vagabond
genes). In fact, there are segments of genetic information in
most (if not all?) eukaryotic cells and prokaryotic cells that
are inherited independently of the gene-carrying structures
(chromosomes, genophores), or at least that are able to
temporarily disassociate themselves from the structures.
This heterogeneous group includes the plasmids of many
bacteria and some eukaryotes, as well as the so-called inser-
tion sequences and transposons (jumping genes).
Biology as a Natural Science
Living nature is impressive because it supports a huge
diversity of life forms. Recording, describing, and system-
atically organizing all living and extinct organism types
(species) is the enormous, as yet unfinished, task of biol-
ogy, in particular, systematics. But biology is not just
restricted to the description of what is there, even more,
it strives to explain this diversity. Besides observation and
comparison, there is the experiment. An experiment is the
observation of a process under artificial predetermined or
controlled varied conditions. Data from experiments and
observations provide the raw material for constructing
hypotheses and theories, contributing toward the expla-
nation of causal relationships. (H. Poincare: ‘‘A heap of
facts is as much a science as a heap of stones make
a house.’’). By forming a repeatedly questioned theoretical
construct (see below), the discovery of correlating laws and
their final formulation into natural laws can incorporate
lots of observations into short, clear units that can then be
Introduction 5
considered. It would be impossible to intellectually pene-
trate the real world with all its basically nonrecordable
structures and experiences without this type of abstrac-
tion. Thus, the natural sciences have become enormously
significant in recent times, especially modern biology
(keywords ‘‘biotechnology,’’ ‘‘gene technology’’).
The sum of recognized natural laws and their inter-
pretation forms the scientific world view, a simplified
reflection of nature presented in perceptions, symbols,
and ideas. This world view is the highest expression of
our understating of nature. It enables mental operations
(thought experiments) that would be too dangerous or
expensive to conduct in the real world. The scientific
world view is thus fundamentally dynamic, as novel infor-
mation is acquired by research, and new interpretations
can be expanded and altered. It is perforce preliminary
and fragmentary and cannot (should not) ever be seen as
being complete. Even so, it is the best that humankind can
offer. The fragmentary character (nature) of the scientific
world view is affected not just by the selection (even if
subconscious) and use of limits of scientific endeavor, but
also by the limits of methodology and primarily by the
limitations imposed by the type of research. These are
indirect in fundamental research as the aims and results
are initially unknown. Indirect aims are researched in that
testable hypotheses are postulated (Greek hypothesis, to
suppose). A hypothesis, a scientific concept, can never
be proven because the data will never be enough. However,
a general theorem can be rejected on the basis of one
contradictory event (asymmetry of verification and falsi-
fication; see K.R. Popper). The assumption ‘‘all roses are
red’’ cannot be proven even with 1,000 red roses, but
can be rejected on the observation of a single yellow or
white rose.
Correlations are based on laws of relationships
derived from observed events (e.g., cigarette smoking/lung
. Fig. 2
Tobaccomosaic virus particles can be seen as rod-shaped particles under an electronmicroscope (EM). Every virion contains
a helical RNA molecule. In the uninjured state, it is made up of a series of 2,130 identical protein molecules each with
158 amino acids. The central axial canal formed in the RNA helix is clearly visible in this negative-contrast slide. Scale bars
(a) 0.1 mm, (b) 0.02 mm (EM images from a F. Amelunxen and b C. Weichan)
6 Introduction

cancer; but also the frequency of storks and human birth
rates in some regions). Correlations can mean there is
a causal relationship, but this does not have to be the
case. If two quantities B and C are correlated, then B can
cause C or the other way round; B and C could be caused
by a third, common, as yet unknown quantity A; they are
correlated but not causally, only coincidentally. Although
the lack of a correlation implies the lack of a causal rela-
tionship, a correlation is not evidence for one; therefore, it
cannot be used for the verification of an assumption.
The asymmetry of verification and falsification means
that forward steps in knowledge are achieved indirectly,
not directly, as inappropriate or inapplicable hypotheses
are rejected (method of trial and error). The aim, the
appropriate knowledge and explanatory reasons, can
only be achieved through disappointment and via detours
(Greek methodos means not only thorough research but
also detour).
With every failed falsification attempt, the probability
of finding the right hypothesis increases. When the hypoth-
esis can be applied to other areas independently of the
original research, it becomes more plausible. Comprehen-
sive hypotheses that, despite many attempts, remain
nonfalsifiable become theories. Theories are elements in
the scientific world view. A theory, e.g., the central biolog-
ical theory of descendancy or evolution, allowsmany events
to be explained and enables the formulation of numerous
testable postulates. From a theoretical scientific viewpoint,
a theory presents a disciplinary matrix or paradigm that
provides the intellectual framework for further experi-
mental work in an area of interest. Surprisingly, even
though specific observations and appropriate experiments
are made on the basis of hypotheses and theories, most
research is not inductive (based on experience and under-
standing) but deductive. It is not primarily targeted at
discovering the unknown or novel, but serves to check
and refine an existing paradigm. Of course, existing
‘‘tested’’ theories can be falsified. Then a new more com-
prehensive theory has to be developed. These scientific
revolutions (see L. Fleck; T.S. Kuhn) are only successful if
they can also explain why the previous theory appeared to
explain so much. Often it becomes apparent that the older
theory does in fact still hold true within certain limits. The
history of scientific biology has many examples of such
scientific revolutions, such as in the developments of cell
biology and genetics.
The teachings of the potential and limits of human
knowledge (as outlined above) form part of epistemology,
which is an important tenet in theoretical science as well as
in philosophy (see, e.g., I. Kant). Even so, the potential to
use knowledge from independent logic or mathematics to
form ideas about real life or nature remained a mystery for
a long time. (A. Einstein: ‘‘The incomprehensibility of the
world lies in its comprehensibility.’’).
Special Position of Biology
The uniqueness of life in nature gives biology a special
position among the sciences. Time and again it has been
questioned whether living systems and systems of abiotic
nature adhere to different laws, and special life forces
(vitalism) have often been postulated. However, to date
there is no known case where physical and chemical laws
have been disobeyed by living organisms. On the other
hand, organisms are incredibly complex systems, which
means that biological systems obey laws that would oth-
erwise not be observed. One speaks of emerging attri-
butes. An important consequence of living systems is that
biological materials cannot be logically or mathematically
penetrated in the same way as objects in physics and
chemistry. Biology is an exact science based on the recog-
nition of natural laws, but observation, description, and
comparison play a much greater role than in physics.
However, a complete derivation of all biological phenom-
ena from chemical and physical laws, in the sense of
a consistent reductionism, would be illusory.
The definition of life as a self-replicating system is
further supported by a fact that emphasizes the unique-
ness of the organism – biological teleonomy. Life forms
behave purposefully, they react expediently, and appear to
be constructed sensibly. Besides the question ‘‘why?’’
(causality), biology (and only biology among the sci-
ences) also justifiably asks ‘‘what for?’’ (finality). This
touches on the cyclical development of life; compare the
terms ‘‘developmental cycle,’’ ‘‘reproductive cycle,’’ and
‘‘generation cycle.’’ From any given starting point, these
cycles proceed along genetically predetermined develop-
mental lines until they return to a comparable starting
point (e.g., egg cells, spores). This results in semicyclical
events and chains of cause and effect. For example,
a particular developmental stage B can arise not only as
a result of the previous stage A but also via the subsequent
stages C, D, etc., also as a cause for the renewed occurrence
of stage A (even if it is chronologically out of sequence).
The final viewpoint bears as much weight as the causal
approach in biology. In inanimate nature, cyclical systems
(e.g., oscillations) do not have mechanisms whereby losses
are compensated for by energy gains, and they finally stop
altogether. Life forms, on the other hand, can replicate by
reproduction. Even in the research of evolution and the
origins of life, biology finds itself in an unusual position.
Introduction 7

While the highest priority is typically the search for natu-
ral laws – reflected in the regular repetition of structures or
processes – in fact here it is the singular, chance event that
is decisive. This is related to the reproduction and selec-
tion of organisms. Natural mutations are chance events
and not predictable. Such mutations can remain neutral
for a very long time until such time as the conditions
suddenly change, making the mutations have negative or
positive effects on the organism. If the mutation has
a favorable effect on the bearer, then according to evolu-
tion by natural selection in successive generations, the
mutation will become fixed. Life forms are, in this respect,
very effective enhancers: all their observable inheritable
traits are derived from improbable and thus rare chance
events (singularities) whose effects are retrospectively
enhanced by reproductive processes.
Animals and Plants
Since the historically based (rather than factually based)
tendency to specialize has been superseded, modern biol-
ogy is dominated by interdisciplinary cooperation.
Knowledge drawn from genetics, biophysics, and bio-
chemistry as well as physiology contributes to a broad
foundation for general biology. Even evolutionary and
developmental biology, as well as molecular and cell biol-
ogy, have grown beyond the boundaries of the classical
disciplines of botany and zoology. However, this connec-
tivity should not be allowed to disguise the fact that the
typical animal and typical plant (both terms used in the
colloquial sense) have numerous differences.
The typical animal is able to migrate. Its body is
compactly constructed, with all organs except those
required to interpret environmental signals being posi-
tioned inside. In order to see them, the animal’s body
has to be opened (‘‘anatomy’’ is derived from the Greek
wordmeaning ‘‘to separate and cut up’’). The large surface
areas necessary for breathing, nutrient resorption, and
excretion are folded inside the body cavity. The outer
surface area is reduced, and so the animal is a ‘‘closed’’
organism. The compact body structure enables the devel-
opment of central organs for circulation and excretion.
Even the nervous system, which enables rapid coordina-
tion, shows a tendency to become centralized over
evolutionary time. Most organs are formed in a limited
number and are at least rudimentarily present in the
embryo, growing proportionately with the growing
organism. Body symmetry is predominantly bilateral and
dorsiventral, as expected when the two perpendicularly
oriented vectors of gravity and motility are present.
Radially symmetrical forms (in the strictest sense) only
occur in sessile or aquatic species. The specialization of
tissues and organs is highly advanced. Even meristems are
specifically determined for the formation of particular
cells (stem cells of blood and immune systems, the skin,
the intestinal epithelium, etc.). The lifetime of even large
animals is limited. Regenerative potential is often quite
low in highly developed animals. Some highly differenti-
ated cells remain active throughout the animals’ lifetime
and are normally not regenerated in the adult phase (large
neurons, striated muscle fibers, cells of the optical lens).
A typical plant is usually ‘‘rooted’’ in one place for the
whole of its lifetime. The pollen, seeds, or spores of the
plant have, theoretically, limitless distribution possibili-
ties. The body area is maximized by unfolding and
branching. The plant is an ‘‘open’’ organism; perennial
plants grow with numerous shoot apices and grow more
in every vegetative period (for trees, annual growth from
all shoot tips, annual rings in the wood, etc.). Metabolic
waste products have to be removed by each cell individu-
ally; instead of centralization there is localized cellular
excretion. The body is mostly radially symmetrical.
There is an enormous regenerative potential; each shoot
apex can, in principle, grow a complete new plant. This
aspect is used extensively in horticulture and agriculture
for the vegetative propagation of plants by grafts, cuttings,
scions, bulbs, bulbils, etc. Furthermore, novel shoot apices
may arise in injury-related callus growths (tissues gener-
ated by chaotic cell proliferation). Thus, cell cultures (even
from single vegetative cells) can be used to successfully
generate whole plants, something that is not possible from
animal cells or tissues. It is not unusual to find plants that
live to 100 or even 1,000 years of age. Clones are immortal.
Thus, e.g., all apples of the same variety are perpetuated,
by grafts, from the same genetic clone as from the apple (of
that sort) that was first discovered, regardless of where it is
now cultivated.
Plants and animals also differ significantly in the struc-
ture and function of their cells. A general comparison
shows that plants cells are not only distinct as a result of
possessing plastids. They are not only phototrophic but
are also osmotrophic (only able to take up substances that
are dissolved), whereas animal cells are phagotrophic (able
to take up nutrients in particulate form). Flagellates
include mixotrophic species that are able to take up nutri-
ents in both ways (> Fig. 3). The plant cell has, in its fully
grown state, a central vacuole that makes up over 90% of
the cell volume, and a cell wall. The cell wall absorbs the
hydrostatic pressure of the vacuole (turgor) that would
otherwise cause the cell to burst. Turgor is a consequence
of osmosis; the molar total concentration of the cell sap in
8 Introduction

the vacuole is far greater than that of the imbibed water in
the cell walls. Animal tissue cells have neither large vacu-
oles nor tough cell walls that have a stabilizing function for
the individual cells. Their turgor is low because they are
surrounded by isotonic body and tissue fluids. Themass of
intercellular substances of the connective and supporting
tissues of animals fixes not cells but supercellular struc-
tures. During plant and fungal cell division, the first wall
primordium arises between the daughter cells via internal
secretion of wall substances. In contrast, typical animal cell
division occurs by pinching off daughter cells from the
mother cell (cleavage), and, whereas the cells of the plant
body are almost without exception anchored to their point
of origin, the cells of animals may migrate and be
translocated during development.
Fungal cells are – apart from not having plastids and
not exhibiting phototropism – more similar to typical
plant cells than animal cells. They are vacuolated,
osmotrophic cells with stable, nonrupturing cell walls
that generally do not cleave but divide by novel cell wall
formation (laid down by internal secretions).
Classification and Significance ofPlant Sciences
The investigation of the plant, fungal, and protist world –
in fact just like for the whole world of organisms – can be
considered from many different viewpoints. For example,
research areas could reflect the hierarchy of structures to
be investigated (> Table 1). Basic research aims to gain
understanding of the origin, diversity, and connectivity
between form and function. This places the research object
to the fore. Applied research is more concerned with
the use of plants, fungi, and microorganisms in human
and animal nutrition; for medicinal, toxic, and drug-
producing plants – the foundations of pharmacology;
plant breeding, genetic manipulation, and biotechnology;
for use in agriculture and forestry; for phytopathology,
pest and weed control; and for landscaping, nature and
animal conservation, and ecology (as defined by the mod-
ern media). Basic research provides the essential back-
ground knowledge for every type of applied research.
In this handbook, the description of the general struc-
tural basics is in Part I. This treats the areas from atomic
up to macroscopic dimensions: the overview ofmolecular
basics is followed by a discussion of the structure and fine
structure of the cell (cytology), followed by discussion of
plant tissues (histology) and then the outer structure as
seen with the naked eye (morphology).
The structures are presented in Part II according to the
general function in metabolic and energy exchange, change
of form (metamorphosis), and motility. The dynamics of
life processes are illustrated by this so-called plant physiol-
ogy. The discussion of the physiology of metabolism is
followed by discussion of developmental physiology and
then physiology of movement. An especially current topic
is allelophysiology – the diversity of physiological rela-
tionships that plants have with other organisms.
The division of this handbook into parts and chapters
should not obscure the fact that modern biology is distin-
guished by its interdisciplinary approach. Areas that were
. Fig. 3
Poterioochromonas malhamensis, a mixotrophic flagellate from the order Chrysomonadales (see also Fig. 10.83) with two
unequal anterior flagella and lobopodium (L) as well as a posterior anchorage appendage (�1,160). The cell on the left has
a nucleus (N) with a nucleolus, plastid (P), and storage vacuole (V). The cell on the right has a large digestive vacuole with
a half-digested algal cell in it (Interference contrast, microflash image from W. Herth)
Introduction 9

once separated have grown together and have given rise to
new, especially productive fields of research. This is what
happened when, e.g., descriptive cell research (cytology),
biochemistry, and molecular biology came together and
formed modern cell biology.
Part III firstly outlines evolution research and deals
with the natural laws and causes governing speciation and
its genetic background. Secondly, it is largely taken up
with botanical systematics. The study of relationships is
based on the results of many other disciplines, and sys-
tematics describes, names, and classifies more than
500,000 known plant species. The classification is based
on the reconstructed phylogeny of the plant kingdom.
Evidence from nucleic acids and proteins (molecular phy-
logeny) and from plant fossils (paleobotany) plays a domi-
nant role. The chapter on systematics contains information
from many different specialized areas that intensively deal
with individual organismal groups (microbiology, bacteri-
ology, mycology, etc.) as well as applied disciplines that
study the practical uses of plants for humans.
Plant ecology deals with the relationships that plants
and plant communities have with their biotic and abiotic
environment. Ecological botany aims to understand facts,
natural laws, and causes for the distribution and gregari-
ousness of plants in space and time.
Plant ecology highlights the significance of plant sci-
ences, especially in the modern world. In terms of energy,
all life on Earth is dependent on phototrophic organisms,
in particular, plants: they are the only relevant (in terms of
sheer numbers) producers and are found at the start of
almost all food chains and at the base of all food pyramids.
This has been true for at least a billion years. Thanks to
their enormous biodiversity, plants support the structure
and functioning of highly diverse ecosystems under a huge
range of conditions, from the Polar Regions to the Tropics.
This diversity and the function of individuals in the
biosphere are severely threatened by the no longer negli-
gible influence of approximately seven billion people.
Unsustainable land use and atmospheric changes mean
the threat to the biosphere has reached global proportions
(global change). Even humans belong to those organisms
whose existence as individuals and species entirely
depends on a stable environment. In these circumstances,
a climate protection scheme based on scientific principles
is more important than ever.
Plant science plays an important role in the develop-
ment of biological sciences. Many fundamental biological
principles have been developed on the basis of plant stud-
ies. These include the discovery of the cell and cell nucleus,
chromosomes, mitosis, meiosis, osmosis, and laws of
genetic inheritance. Even though the solutions to many
problems in modern biology have been found using
microorganisms and certain animals with particularly
appropriate systems, and many medically relevant ques-
tions regarding cancer, immune systems, thought, and
consciousness can only be answered using (higher) ani-
mals, botany nevertheless remains a key area of basic
biological research. This is evidenced by the enormous
advances made in modern plant sciences (see the model
plant Arabidopsis thaliana). As before, the applied plant
sciences are also immensely important. Plants and fungi
play a central role next to bacteria in biotechnology. It is
therefore not surprising that ‘‘green gene technology,’’ the
application of gene technology in agriculture, has rapidly
become increasingly significant. As everywhere in modern
biology, the deciphering of genomes (genomics) is being
continually updated as different protein complements
are discovered (proteonomics) and the metabolites of
different cells in the same organism are analyzed
(metabolonomics).
. Table 1
Biological fields of research and the complexity of the
objects studied
Structure Fields of research
Atoms Biophysics
Molecules Biochemistry
Semantic
macromolecules
Molecular biology
Genes, chromosomes Genetics
Cells Cell biology
Tissues Histology
Organs Anatomy, physiology
Organisms Morphology, developmental
physiology, systematics, phylogeny,
autecology
Populations Reproductive biology, evolutionary
biology
Communities Geobotany, community ecology
Ecosystems Biogeochemistry, ecosystem
biology
10 Introduction

















