2
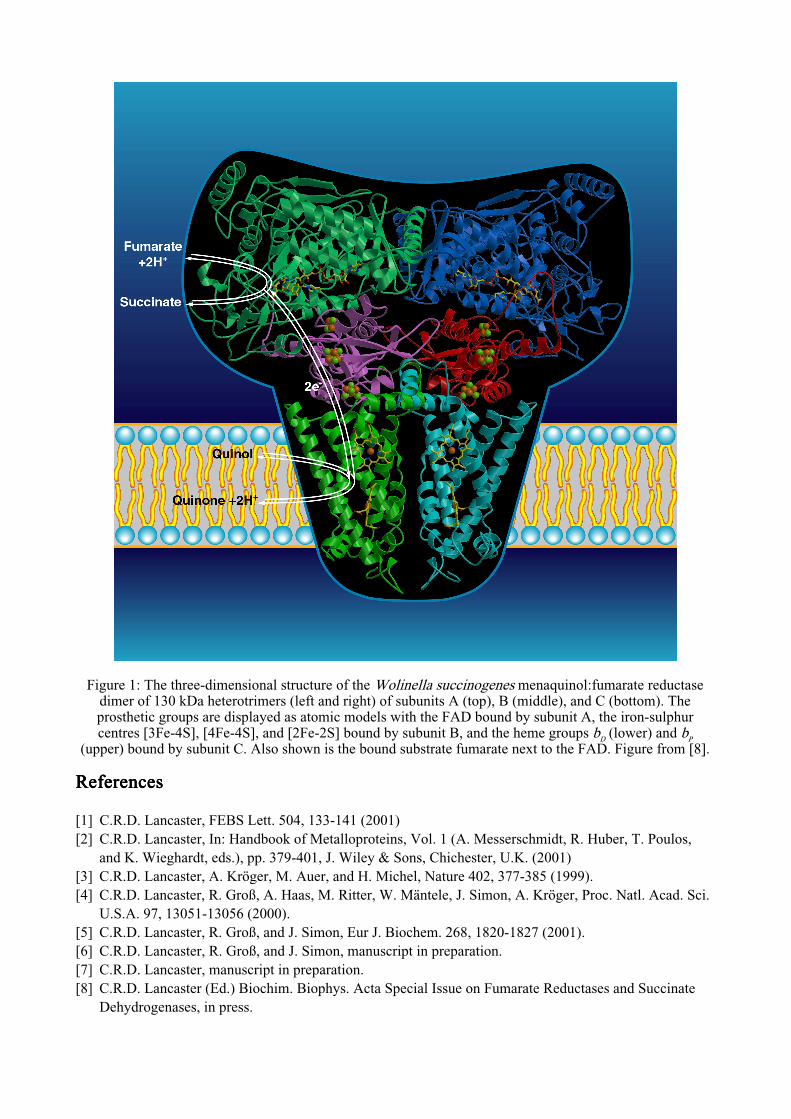
Structure of quinol:fumarate reductase from Wolinella succinogenes – Towards a mechanism of action C.R.D. Lancaster Max Planck Institute of Biophysics, Department of Molecular Membrane Biology, Heinrich-Hoffmann-Str. 7, D-60528 Frankfurt am Main, Germany Quinol:fumarate reductases (QFR) and succinate:quinone reductases (SQR) catalyse the reduction of fumarate to succinate with concomitant oxidation of hydroquinone (quinol) to quinone as well as the reverse reaction (see [1,2] for recent reviews). SQR (respiratory complex II) is involved in aerobic metabolism as part of the citric acid cycle and of the aerobic respiratory chain. QFR is involved in anaerobic respiration with fumarate as the terminal electron acceptor, and is part of the electron transport chain catalyzing the oxidation of various donor substrates (e.g. NADH, H 2 or formate) by fumarate. These reactions are coupled to ADP phosphorylation via an electrochemical proton gradient. The QFR complex from the anaerobic bacterium Wolinella succinogenes consists of two hydrophilic subunits, A and B, and one hydrophobic, membrane-integrated subunit C (Figure 1). The larger hydrophilic subunit A carries covalently bound FAD (8α-(N3)-histidyl flavin adenine dinucleotide) and subunit B contains three iron-sulphur centers, one each of the [2Fe-2S] 2+,1+ , [4Fe- 4S] 2+,1+ , and [3Fe-4S] 1+,0 type. Subunit C binds two haem b groups. At resolutions of up to 2.2 Å, the crystal structure of W. succinogenes QFR has been determined in three different crystal forms, all of the monoclinic space group P2 1 [3,4,5]. The unit cell of crystal form ‘‘A’’ is a = 85.1 Å, b = 188.8 Å, c = 117.8 Å, and β = 104.5º. Crystal form ‘‘B’’ has the unit cell dimensions a = 118.3 Å, b = 84.9 Å, c = 188.6 Å, β = 96.5º. There are two 130 kDa complexes in the asymmetric units of these unit cells. In the third crystal form, ‘‘C’’, which has the unit cell dimensions a = 80.9 Å, b = 289.6 Å, c = 153.3 Å, and β = 95.7º, there are four complexes in the asymmetric unit. All three crystal forms have contributed towards our understanding of the structure and mechanism of action of W. succinogenes QFR [3,4,5]. However, in spite of the recent progress, numerous question as to the precise mechanism of action remain. These are being approached by a combination of site-directed mutagenesis, substrate and inhibitor-binding studies, and functional and structural experiments, particularly with respect to the two active sites of the enzyme. During the beam time allocated to this proposal, six higher quality data sets for crystal form A and one for crystal form C could be collected (Table 1). Some of these data sets have provided the basis for two publications which are currently under preparation [6,7]. Table 1: Diffraction data collected at X11 (7 Table 1: Diffraction data collected at X11 (7 Table 1: Diffraction data collected at X11 (7 Table 1: Diffraction data collected at X11 (7-8 August & 22 8 August & 22 8 August & 22 8 August & 22-23 August 2001) 23 August 2001) 23 August 2001) 23 August 2001) resol. range measured unique complete I/σ(I) R sym * [Å] reflections reflections [%] (outer [%] crystal form A shell) 110Csd1 50.0-2.45 301,993 127,221 95.6 2.4 9.9 inhib3_100_b1 50.0-2.50 475,647 124,408 99.5 5.0 9.2 inhib7_96_b01 50.0-2.40 520,751 139,642 99.6 5.2 9.9 inhib5_96c1 20.0-2.23 581,079 173,348 99.1 2.5 8.4 subst95_5_01 15.0-2.80 218,851 80,848 91.7 2.0 10.0 110Ke1 15.0-2.50 454,721 123,439 99.7 3.0 9.4 crystal form C mutant_97d01 50.0 -- 3.20 282,879 102,803 88.9 2.9 8.1 Due to the optimum conditions for data collection provided at X11, only one crystal was required for each dataset listed above.





![Biochimica et Biophysica Acta - CORE · “non-classical” (type E) SQOR from Wolinella succinogenes is a QFR with no SQR activity [12]. The first three-dimensional structure of](https://static.documents.pub/doc/80x56/611715262f05f7769857d448/biochimica-et-biophysica-acta-core-aoenon-classicala-type-e-sqor-from-wolinella.jpg)











