STRUCTURE OF REGENERATED SYNAPTIC CONNECTIONS OF ALLOTRANSPLANTED PHASIC MOTONEURONS ON A TONlC MUSCLE IN CRAYFISH A thesis submitted in conformity with the requirements for the degree of Master of Science, Graduate Department of Zoology, University of Toronto O Copyright by Rahirn Hirji, 1999
Transcript
STRUCTURE OF REGENERATED SYNAPTIC CONNECTIONS OF ALLOTRANSPLANTED PHASIC MOTONEURONS ON A TONlC
MUSCLE IN CRAYFISH
A thesis submitted in conformity with the requirements for the degree of Master of Science,
Graduate Department of Zoology, University of Toronto
O Copyright by Rahirn Hirji, 1999
National Library l*I , cana- Bibliithèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques
395 Wellington Street 395. me WdCinm OnawaON K1AûN4 OttswaON K I A W canada Canada
The author has granted a non- exclusive licence aiiowing the National Library of Canada to reproduce, loan, distriiute or seii copies of this thesis in microfom, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fiom it may be printed or otherwise reproduced wîthout the author's permission.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.
STRUCTURE OF REGENERATED SYNAPTIC CONNECTIONS OF ALLOTRANSPLANTED PHASIC MOTONEURONS ON A TONlC MUSCLE IN
CRAYFISH
An abstract submitted in confonnity with the requirements for the degree of Master of Science in Zoology,
Graduate Department of Zoology, University of Toronto
Rahim Hi ji, 1999
ABSTRACT
Crustacean neuromuscular systems are highly defined into phasic and tonic
types exemplified by the deep and superficial flexor muscfes in the abdomen of the
crayfish Procambarus clarkii. The deep phasic muscle is used occasionally and briefly
for escape responses while the superficial tonic muscle is used continuously for routine
movements and maintaining posture. To assess the relative importance of the
motoneuron and the target muscle in detennining the synaptic phenotype of
regenerating neuromuscular connections, we attempted a mismatch experiment in
which the denervated tonic SFM received an allotransplanted phasic flexor newe. A
quantitative study of various structural parameters of these regenerated synapses
using serial section electron microscopy revealed that these synapses, on average,
possessed two dense bars that were often closely spaced. Mitochondria comprised a
relativeIy small (6%) volume of the tenninals- Collectively, these structural features
resemble those of native phasic synapses, thereby corroborating the physiological
evidence for the regeneration of phasic type synapses. Therefore, the motoneurons
and not their target muscle fibers are most influential in specifying the phenotype of
regenerated synapses.
ACKNOWLEDGMENTS
I would like to begin by expressing humble gratitude to my supervisor C.K. Govind,
who comrnand of knowledge, dedication and meticulous nature were integral in guiding
me to do my best. I would also like to thank Joanne Pearce for her patience and
willingness to help me whenever I was in need. Our mutual interest in stock markets
has helped fonn a bond that I hope will hold strong over time. 1 woold also like to thank
Asheer Sharman for his friendship, understanding and support in many aspects of my
life. Honourable mentions go out to Naz, Matt and Raymond. I would like to express
my deep gratitude to my family (Azim, Yasrnin, and Adil) for their ongoing motivational
support and for understanding my idiosyncrasies, and Fenulla for being there for me
and loving me for who i am; having al1 of you in my life makes me extremely fortunate.
TABLE OF CONTENTS
...................................................... I . Crustacean Neuromuscular Systems 1 A . Differentiation into excitatory and inhibitory axons ......................... 2
.................................... B . Differentiation into phasic and tonic axons 4
.................................... C . Synaptic differentiation within a tonic axon 8
................................... D . Structural correlates of transmitter release 10
............................................... II . Crustacean Neuromuscular Regeneration 12 A . Regeneration studies in the SFM ....................... ..... .............. 13 B . Neural allotransplantation in the SFM ......................................... 15
. . .................................................................... III . Experimental Objectives 17
MATERIALS AND METHODS ................................................................. 19
............................................................. I . Allotransplantation Procedure 19
II . Electron Microscopy ......................................................................... 20
...................................... III . Quantitative Analysis ...................... .. ... ... 21
IV . Volumetric Analysis ........... .... ..................................................... 23
................................................ I . Nature of Regenerated Nerve Terminais 51 ............................................................ A. Nerve terminal morphology 52
B . Mitochondrial volume .................................................................... 53 ............................................................... C . Dense bars per synapse 54
Structural-Functional Correlations ........................................................ Synapses .................................................................................. a . area ..................................................................................... b . number .................................................................................
Dense bars ................................................................................ a . length ........................ ... ...................................................... b . number ................................................................................
................................................................................ . c spacing
Other factors involved ..................................................................
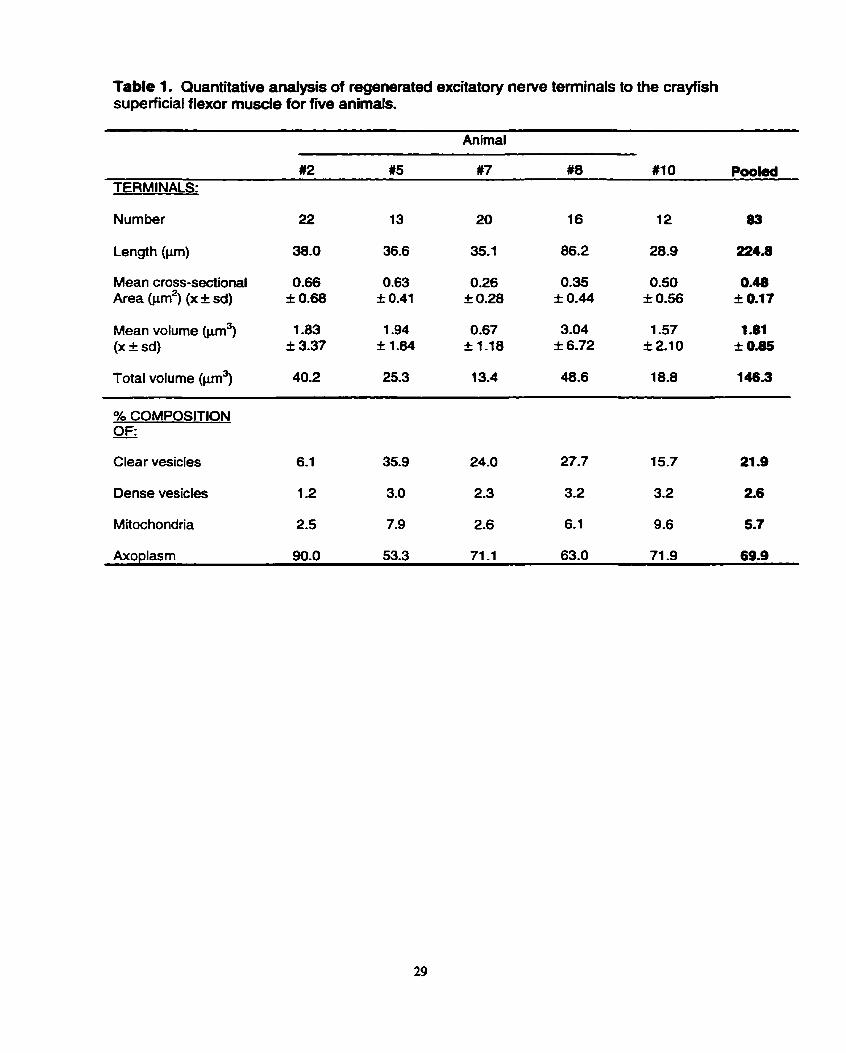
approximately one-fiih of terminal volume (21.9%) for clear vesicles. Dense core
vesicles, on the other hand, were much less prevalent; they occupied between 1.2%
and 3.2% of terminal volume in the five animals. Mitochondria also occupied markedly
l e s terminal volume than the cfear vesicles in each of the five animals; the range
across the five animals studied was 2.5% to 9.6% of terminal volume, with an overall
mean of 5.7% (Table 1). In most terminals across al1 five animals, the mitochondria
appeared simple and unbranched. Furthemore, some of the nerve terminals in each
of the five animals lacked mitochondria-
B. Synapses
Neuromuscular synapses, characterized by the close apposition of
presynaptic (newe terminal) and postsynaptic (muscle fiber) membranes, represent
the sites of information exchange between nerve and muscle. The space between
these two membranes is referred to as the synaptic cleft, which is filled with electron
dense material and is typically 15 nm wide (Fig. 2). Once terminal regions were
located, fully sectioned regenerated synapses were identified, and synaptic areas were
calculated (Table 2). Between 37 and 88 excitatory synapses were identified in each
of the five animals. This study also examined the number of synapses per prn of
terminal, revealing similar values among the five animals and a mean (across the five
anirnals) of 1.48 synapses per Pm of terminal. A comparison of the number of
synapses per unit terminal volume, however, revealed great variation among the five
animals. The range was from 1.34 to 5.22 with a mean of 2.52 synapses per pm3 of
newe terminal (Table 2).
Mean synaptic areas for excitatory synapses, which varied from 0.241 * 0.178 pmZ (animal #7) to 0.527 t 0.485 Pm2 (animal #5), were statistically analyzed to
Table 2. Quantitative analysis of regenerated excitatory synapses ta the crayfish superficial flexor muscle for f i e animais.
Animal
#2 R5 Ut? #8 #1 O Poolad SYNAPSES:
Number 54 57 70 88 37 306
Num ber/pm of terminal 1 -64 1-53 1.75 1.12 1.38 1.48
Mean area (pm2 ) 0.377 0.527 0.241 0.404 0.415 0.409 (X k sd) I 0.277 f 0,485 f 0.1 78 f 0.302 i 0246 I 0.111
Total area @rn2) 20.4 30.0 16.8 42.6 15.4 125.2
A r e a m of terminal (pm2) 0.54 0.82 0.48 0.49 0.53 0.57
~ r e a / ~ ~ m = of terminal m2) 0.51 1.19 1 -25 0.88 0.82 0.93
determine if the means were similar. The analysis revealed each of the animals had a
mean synaptic area that significantly differed from at least one other animal; the mean
synaptic area for animal #7 was significantly lower than each of the other four animals.
Overall, 3û6 regenerate excitatory synapses were measured to yield a mean synaptic
area of 0.409 I 0.1 11 pm2. There was considerable variance around this mean; in
fact, individual synaptic areas ranged from as low as 0.017 pm2 (animal #7) to as high
as 2.472 pm2 (animal W).
Due to the great variation in synaptic areas, we looked at the synaptic
area per pm of nenre terminal for each of the five animals. The values ranged from
0.48 to 0.82, with a mean synaptic area per prn of terminal of 0.57 (Table 2). Since
these regenerated terminais are quite thin, synaptic area was then normalized to
terminal volume. The resulting synaptic area per pm3 of terminal volume ranged from
0.51 to 1.25 with a mean of 0.93 for the five animals (Table 2). It is clear that, in
relation to terminal volume, synaptic area is fairly high.
C. Dense bars
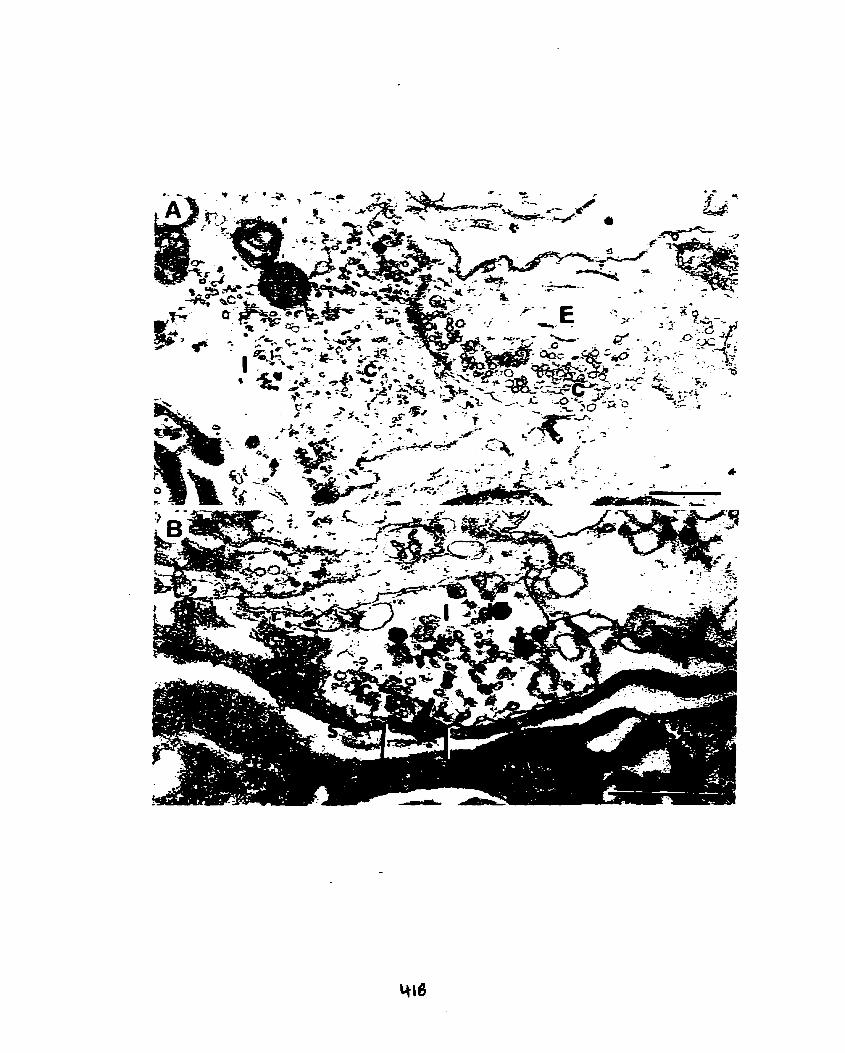
Dense bars are sites for the release of neurotransmitter from nerve
terminals and were obsewed in many of the synapses in al1 five animals (Figs. 2, 3).
Clusters of clear synaptic vesicles were often obsewed surrounding dense bars; we
have captured an omega shaped profile directly adjacent to a dense bar in a thin
section from animal #5 (Fig. 28) which is indicative of clear vesicle exocytosis (Pearce
et al., 1986). Many studies have emphasized the importance of dense bar properties,
especially dense bar number, length, and spacing between dense bars, in detemining
the initial release of neurotransmitter from a nerve terminal (Atwood and Wojtowicz,
1986; Atwood and Cooper, 1996). These parameters were examined in the present
Figure 3. A, B. Regenerate excitatory (E) terminais with spherical clear vesicles,
making synaptic contact (between bars) with muscle membrane in the region of
granular sarcoplasm (s). These synapses display several closely-spaced presynaptic
dense bars (arrowheads).
Scale bar: 0.25 Pm. Magnification: x83 300.
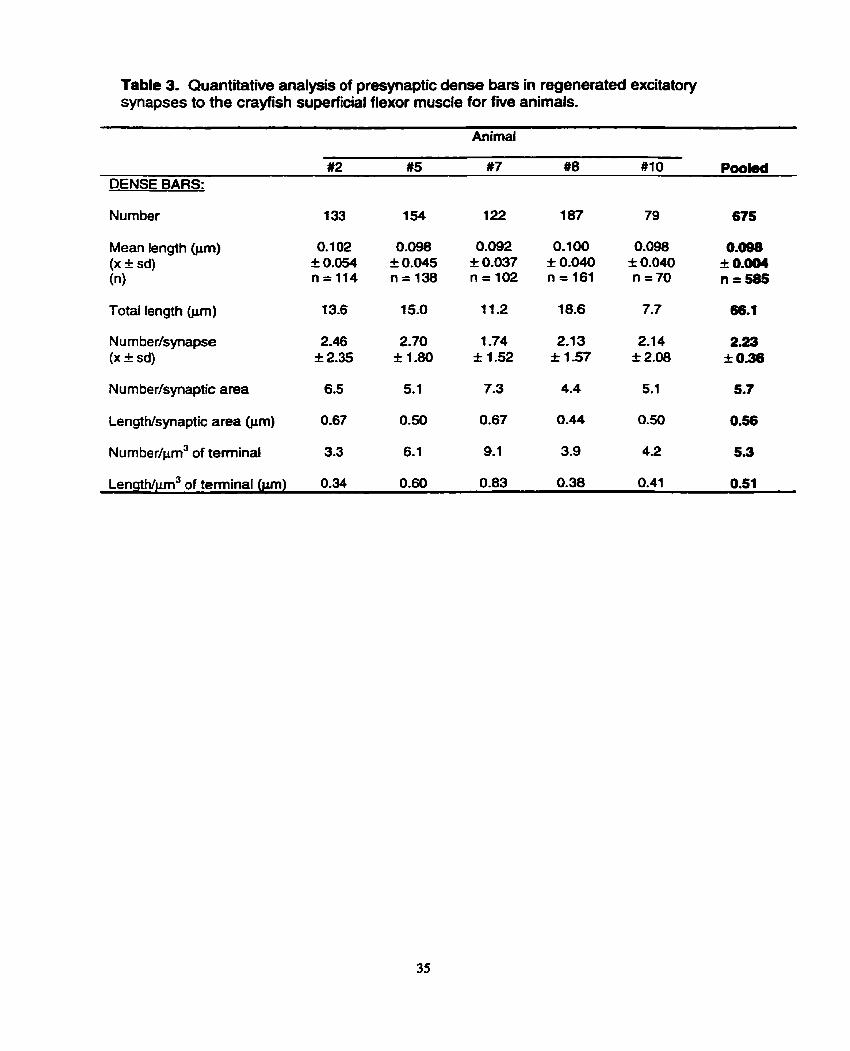
study through a quantitative analysis of dense bars idenüfied in al1 fully sectioned
synapses in the fnre animals (Table 3). Each of the f i e animals housed between 79
and 187 dense bars in total (across al1 synapses), which depicts a fairly wide variance
arnong animals possibly due to differences in the size and extent of the nerve teminal
region examined.
The mean dense bar length for al1 fully sectioned dense bars in the
regenerated excitatory nerve terminals was very close to 0.1 pm (Table 3) and found
not to Vary significantly across the five animals. This aflowed for an examination of the
nurnber of dense bars per synapse as a comparative measure for the f i e animals.
The mean across the f i e animals analyzed was 2.23 I 0.36 dense bars per synapse,
with animal #7 possessing a significantly lower mean (1 -74 I 1.52 dense bars per
synapse) than the other four animals (range of 2.1 3 I 1.57 to 2.70 t 1.80).
To compensate for differences in individual synaptic areas, the number
and length of dense bars per pm2 of synaptic area were also calculated (Table 3). For
the number of dense bats per pm2 of synaptic area, the range across the five animals
was from 4.4 (animal #8) to 7.3 (animal #7) with a mean of 5.7 across the five animals.
Dense bar length per unit synaptic area exhibited a narrower range, and varied from
0.44 prn (animal #8) to 0.67 Pm (animals #2, 5) with a mean of 0.56 Fm for the five
anirnals studied.
To take nerve terminal volume into account, the number of dense bats per
prn3 of terminal was also examined for each animal; the values ranged from 3.3
(animal #2) to 9.1 dense bars per unit volume (animal #7) (Table 3). Similariy, dense
bar length per pm3 of terminal was calculated to assess the amount of dense bar
material present per unit volume. The values ranged from 0.34 pm (animal #2) to 0.83
pm (animal #7), mirroring the pattern observed for number of dense bars per unit
Table 3. Quantitative analysis of presynaptic dense bars in regenerated excitatory synapses to the crayfish superficial flexor muscle for five animals.
Animal
#2 #5 #7 #8 #1 O P w k d DENSE BARS:
Number 1 33 1 54 1 22 187 79 675
Mean length hm) (X f sd) (n)
Total fength (pm) 13.6 15.0 11.2 18.6 7.7 66.1
Number/synaptic area 6.5 5.1 7.3 4.4 5.1 5.7
LengtNsynaptic area (pm) 0.67 0.50 0.67 0.44 0.50 0.56
~umbedprn~ of terminal 3.3 6.1 9.1 3.9 4.2 5.3
~ength/pm~ of terminal (jm) 0.34 0.60 0.83 0.38 0.41 0.51
volume. Interestingly. the number of dense bars per unit volume and the length of
dense bars per unit volume were significantly higher for animal #7. These findings can
be understood by realizing that the number of dense bars per synapse for this animal
was approximately 20% lower than the overall rnean, whereas the mean terminal
cross-sectional area was approximately 40% smaller than the average terminal.
Furthermore, since dense bar length is statistically similar across the five animals,
animal #7 also exhibits a significantly higher dense bar length per unit terminal volume
than the other four animals (Table 3).
A closer examination of the number of dense bars per synapse showed
wide variance (Table 4). ranging from synapses lacking a dense bar to a synapse in
animal #2 that housed 12 distinct dense ban over its 1.200 pm2 synaptic area.
Synapses with multiple dense bars convey a sense of complexity that is more readily
appreciated by viewing them in two-dimensional reconstruction. Selected examples of
such synapses seen in Figure 4 highlight this complexity and show that most possess
dense bars of variable length, although a few had primarily small dense bars. Figure 3
displays examples of synapses with four distinct dense bars on a single thin section.
Synapses with multiple dense bars are more potent releasers of neurotransmitter
molecules because they have more sites for vesicle docking and fusion, more calcium
channels, and a greater Iikelihood that calcium macrodomains will overlap (Cooper et
al., 1996). We therefore categorized synapses based on the number of dense bars
they possess (Table 4) and found that only a small percentage (between 5.3% and
22.9% with a mean of 14.2%) lacked a dense bar. Thus, the overwhelming majority of
synapses possessed active zones. Among these, a substantial number had a single
dense bar (between 24.3% and 44.3% with a mean of 30.4% of al1 synapses), and a
few synapses possessed two dense bars (between 15.7% and 21.6% with a mean of
Figure 4. Two-dimensional vie- of selected examples of regenerated excitatory
synapses showing the number and distribution of presynaptic dense bars. Each
synapse is defined by a series of horizontal lines representing thin sections. Each
dense bar is depicted as a filled circle situated on a horizontal line. When dense bars
traverse more than one thin section, the circles representing a single dense bar are
shown joined to each other and appear as a vertical bar.
Scale bars: 0.5 Pm.
Table 4. Quantitative analysis of dense bars in regenerated excitatory synapses to the crayfish superficial flexor muscle for fnre animals.
channel sensitivity to calcium may also exist in regenerated tenninals, resulting in a
disproportionately high release of transmitter (Msghina et al., 1999).
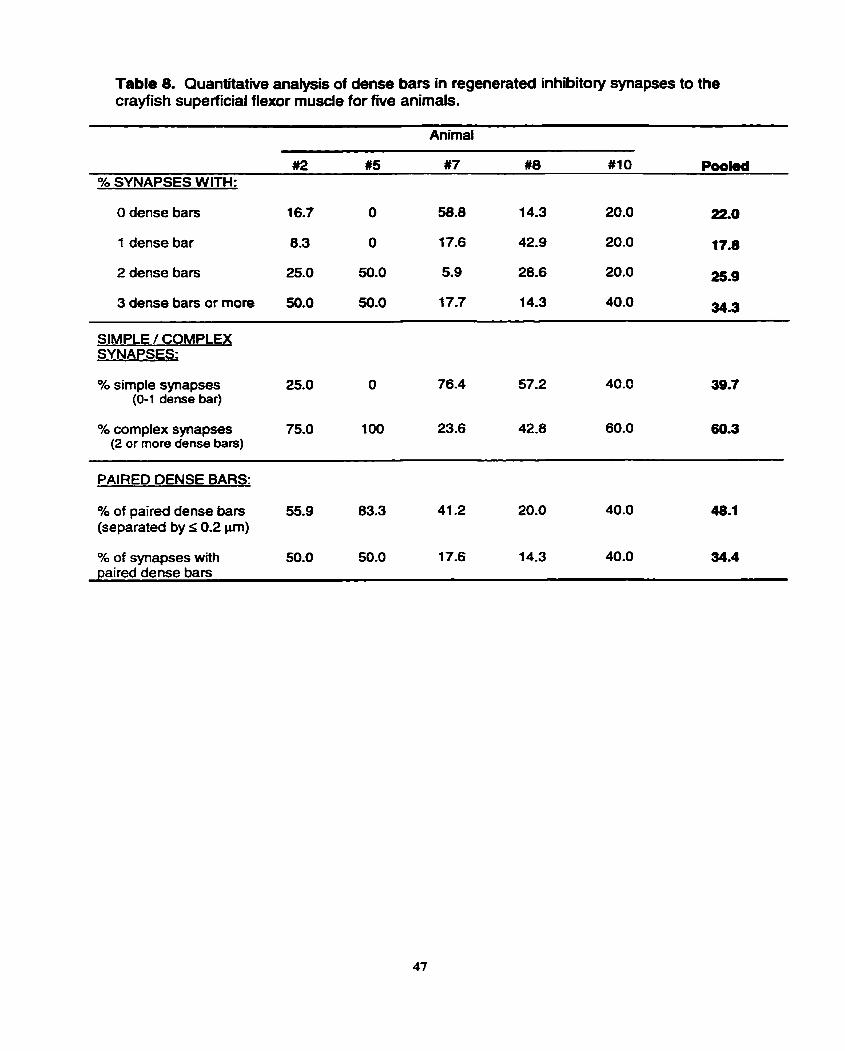
III. INHIBITORY INNERVATION
The major focus of these transplant studies was the excitatory innervation,
which was assessed in the first instance by the presence of EPSPs upon stimulation of
the regenerated nerve (Krause et al., 1998). Such electrophysiological detection of
innervation was not, however, obvious for an inhibitory neuron in that hyperpolarizing
postsynaptic potentials were not recorded. The use of hyperpolarizing potential as
indicators of inhibitory innervation is not conclusive since, depending on the muscle
fibers' equilibrium potential for chloride, inhibitory potentials can be depolarizing
(Atwood, 1976). However, the present structural study revealed clear-cut evidence for
inhibitory innervation in the form of nerve tenninals with elliptical clear synaptic
vesicles. Crustacean neuromuscular synapses fixed with aldehydes show distinctive
differences in the shape of their clear synaptic vesicles (Atwood and Wojtowicz, 1986);
excitatory vesicles are spherical and inhibitory ones are elliptical. The presence of
these inhibitory nerve terminals therefore provides data on synapse regeneration of a
distinctly different type of neuron. Hence, it may be instructive to compare these
regenerated inhibitory terminals with their excitatory counterparts.
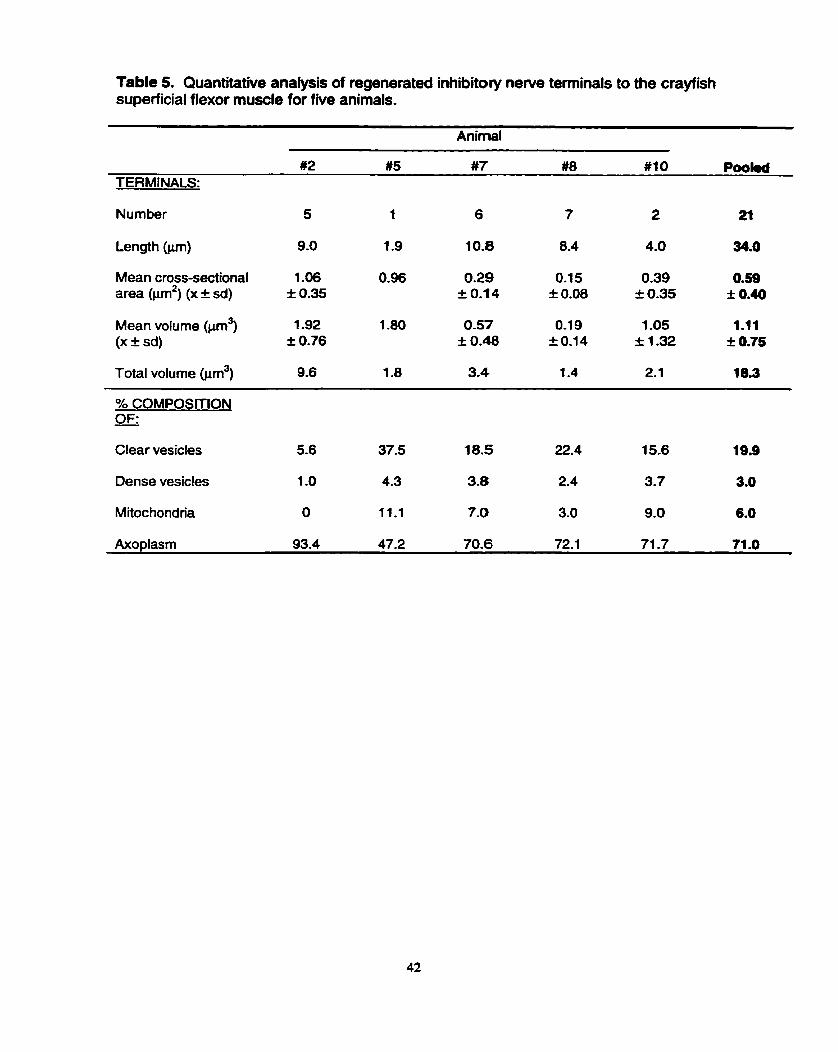
First, regenerate inhibitory terminals were found in each of the five
allotransplanted preparations, making them a consistent feature of the regenerated
innervation. Such consistency is noteworthy since the target muscle, given a choice of
six transplanted neurons with up to fwe excitors and a single inhibitor, accepted the
single inhibitor each time. This outcome most likely occurs because the muscle has
receptor sites for an inhibitory neuron in addition to those for excitatory axons.
Norrnally, fibers of the SFM receive a single inhibitory neuron and two to three
excitatory axons (Kennedy and Takeda, 1965b; Velez and Wyman, 1978a).
Second, the regenerate inhibitory terminals were fewer in number than
excitatory ones in each of the five preparations. This finding points to the possibility
that a single inhibitory neuron regenerated to the SFM, which is in keeping with its
previous history and with the fact that the transplanted phasic nerve would have
possessed a single inhibitory neuron. However, this conclusion is tentative and
requires physiological verif ication.
Third, the size of the regenerated inhibitory terminals, their mean
mitochondrial volume and number of dense bars per synapse were comparable to
those of their excitatory counterparts, suggesting that a phasic type of inhibitory axon,
analagous to the phasic excitatory axons, had regenerated. Such an idea is in
accordance with previous findings; transmitter release characteristics are similar
between excitatory and inhibitory axons to a given muscle fiber (Atwood and Bittner,
1971).
Fourth, inhibitory nerve terminals also make presynaptic contact with
excitatory newe terminals. A functional advantage of presynaptic inhibition is the
conservation of transmitter in excitatory nerve terminals (Bryan and Krasne, 1977).
Furthemore. removal of the inhibition produces a strong synaptic response, since
there is a large readily releasable pool of vesicles that can be drawn upon (Atwood and
Walcott, 1965). Atwood and Morin (1970) foond that axo-axonal synapses polarized
from the inhibitory to the excitatory newe terminal are encountered 10-20 times less
frequently than neuromuscular synapses in the crayfish claw opener muscle.
Physiological examination of these connections at low stimulation frequencies,
however, revealed that presynaptic inhibition is appmximately 10 times more effective
than the more prevalent postsyiiaptic inhibition. Thus, individual axo-axonal synapses
must be at least 100 times more effective than neuromuscular synapses (Atwood and
Morin, 1970). The greater potency of am-axanal contacts may result from the fact that
they are contacting small excitatory nerve terminais instead of much larger muscle
fibers. Therefore, the inhibitory transmitter released upon low frequency stimulation
would have a greater impact on the small excitatory newe terminal membrane
potential than that of a large muscle fiber (Atwood and Morin, 1970). The number of
axo-axonal contacts representative of presynaptic inhibition were seen infrequently in
the current study, yet it is important ta note that these types of contacts, in addition to
al1 of the factors discussed previously, are also capable of controlling the quantal
output from excitatory neuromuscular junctions.
Overall, the presence of regenerated inhibitory innervation shows
robustness of the allotransplanted neural tissue, and alço shows that the muscle target
welcomes, or perhaps even demands, innervation by two different types of axons. In
other words, the muscle has the receptors for both the excitatory and inhibitory axons
and, as a resuit, both types regenerate. Quite possibly, the number of receptor sites
may Iimit the density, and hence the nurnber, of axons regenerating to the SFM.
SUMMARY
1. Crayfish have robust regenerative powers as shown by the fact that after
transecting the superficial flexor newe to the SFM in the abdomen, connecthMy
maps were re-established within 8 weeks (Ely and Velez, 1982). Synaptic
regenerative abilities were further explored through studies involving the
allotransplantation of the superficial flexor newe from a donor animal to a
previously denewated host animal (Krause and Velez, 1995). These studies
showed that the transplanted motoneurons re-established synaptic connectivi
patterns similar to the native patterns.
2. A mismatch experiment involving an allotranspfantation of the phasic flexor nerve
to the previously denewated tonic SFM was attempted (Krause et al., 1998) to
determine if the regenerated connections were tonic in nature (indicating an
instructive retrograde influence), or phasic in nature (indicating that the
regenerating motoneuron follows an autonomous program when forming synaptic
connections). The regenerating newe in these mismatch experiments showed
axons with numerous sprouts indicative of phasic axons; tonic axons have fewer
sprouts. Electrop hysiolog ical studies on experimental animals revealed 3-fold
larger initial EPSPs at low frequencies of stimulation in the regenerated
neurornuscular system when compared to the native SFM system. The EPSP
data strongly suggest that the newly formed connections possess phasic
properties, at least at low frequencies of stimulation.
3. My study examined the ultrastructural properties of the regenerated connections in
these mismatch experiments using thin serial section electron microscopy.
Altogether, fwe animals which had been previously examined with
electrophysiology (Krause et al., 1998) were selected for study. Qualitative
features of regenerated synaptic terminals were typical crustacean-like; nerve
terminals were populated with mostly dear vesicles, a few dense core vesicles and
rnitochondria and possessed well defined synapses, many with one or more dense
bars (active zones). Quantitative analysis of these structural parameters allowed
me to correlate them with the large EPSP and also provided cues as to whether
they were of the phasic type.
4. Previous studies have repeatedly pointed to three primary structural features which
distinguish phasic and tonic terminals: terminal morphology, mitochondrial
volume, and the number of dense bars per synapse. Tonic regenerate terminals
were approximately fie-fold larger than the phasic regenerate terminals examined
in the current study. Phasic regenerate terminals also had fewer mitochondria;
only 5.7% of terminal volume was occupied by mitochondria, compared to a
relatively high 19.2% for tonic regenerate terminals. An examination of the
number and length of dense bars per unit synaptic area as well as per unit volume
revealed No- to four-fold differences between phasic and tonic regenerate
connections. The above mentioned features strongly suggest that the regenerated
newe terminals examined in the current study are phasic in nature.
5. To explore structural-functional correlations in the regenerated system, synapses
and dense bars were examined. Although the phasic regenerate connections had
a smaller mean synaptic area, they had more synapses per unit volume compared
to tonic regenerate connections. An examination of dense bar number and
distribution reveafs that many phasic regenerate synapses were of the complex
variety, and many have 3 or more dense bars. Furthemore, phasic regenerate
synapses had over three-fold more of their dense bars capable of interacting with
an adjacent dense bar when compared to tonic regenerate connections. A
cornparison of the number of synapses possessing an interacting pair of dense
bars revealed a similar trend. These findings collectively support the physiological
finding that the regenerated connections are capable of higher initial transmitter
release than the lower output, tonic connections.
6. Although physiokgical evidence for inhibitory innervation in the form of lPSPs was
not obtained, electron microscopic examination of innervation sites revealed the
presence of inhibitory nerve terrninals. Ultrastructural analysis of the inhibitory
connections revealed key similarities with excitatory connections in newe terminal
morphology, mitochondrial volume and dense bar number per synapse. These
findings suggest that synaptic features of inhibitory innervation are similar to that of
excitatory innervation. lnhibitory nerve terminals also make pre-synaptic contact
with excitatory newe terminals via presynaptic dense bars located on the inhibitory
membrane. Such axo-axonal contacts may represent pre-synaptic inhibition,
which may reduce excitatory impulse activity and help conserve transmitter.
7. In sum, the electrophysiological findings of Krause et al. (1998) of large EPSPs in
regenerated systems were supported by the current ultrastructural analysis.
Although not specifically tested in the mismatch condition, it is quite possible that
connection specificity is maintained by the presence of target derived molecular
cues, which guide regenerated connections to previous sites of innervation. These
cues, however, may have fiffle or no impact on the nature of the regenerated
connections. Thus, an allotransplanted rnotoneuron may use its surroundings to
arrive at the appropriate locations within its novel surroundings, but utilires its
autonomous program to fom synaptic connections.
REFERENCES
Arcaro KF, Lnenicka GA (1 995) lntnnsic differences in axonal growth from crayfish fast and slow motoneurons. Dev Bi01 168:272-283.
Atwood HL (1 976) Organization and synaptic physiology of crustacean neuromuscular systems. Prog Neurobro17:291-391.
Atwood HL (1 982) Synapses and neurotransmitters. In T h e Biology of Crustaceam (H.L. Atwood, and D.C. Sandeman, eds.), pp. 105-150. Academic Press, New York.
Atwood HL, Bittner GD (1971) Matching of excitatory and inhibitory inputs to crustacean muscle f ibers. J Neurophysiol34: 1 57-1 70.
Atwood HL, Cooper RL (1 996) Synaptic diversity and differentiation: crustacean neuromuscular junctions. Invert Neurosci 1 :291-307.
Atwood HL, Govind CK (1990) ActMty-dependent and age-dependent recruitment and regulation of synapses in identified crustacean neurons. J Grp Bi01 153:105- 127.
Atwood HL, Kwan 1 (1976) Synaptic development in the crayfish opener muscle. J Neurobiol7:289-312.
Atwood HL, Lnenicka GA (1986) Structure and function in synapses: emerging correlations. TINS 9:248-250.
Atwood HL, Marin L (1 983) Ultrastructure of synapses with different transmitter- releasing characteristics on motor axon terminais of a crab, Hyas areneas. Cell Ess Res 231 :103-115.
Atwood HL, Morin WA (1 970) Neuromuscuhr and axoaxonal synapses of the crayfish opener muscle. J Ultrastmct Res. 32:351-369.
Atwood HL, Walcott B (1 965) Recording of electrical activity and movement from legs of walking crabs. Can J Zoo1 43:657-665.
Atwood HL, Wojtowicz JM (1 986) Short-terni and long-term plasticity and physiological differentiation of crustacean motor synapses- Int Rev Neurobiol28:275-362.
Atwood HL, Lang F, Morin WA (1972) Synaptic vesicles: selective depletion in crayfish excitatory and inhibitory axons. Science 176:1353-1355.
Atwood HL, Lnenicka GA, Marin L (1985) Morphological responses to conditioning stimulation in a phasic motor axon of crayfish (Procarnbams clarkir). J Physiol (Lond) 365:26P.
Atwood HL, Karunanithi S, Georgiou J, Charlton MP (1997) Strength of synaptic transmission at neuromuscular junctions of crustaceans and insects in relation to calcium entry. lnvert Neurosci 3:81-87.
Augustine GJ, Chariton MP (1 986) Calcium dependence of presynaptic calcium current and post-synaptic response at the squid giant synapse. J Physiol (Lond) 381 :6 1 9-64.
Bittner GD (1 968) Oierentiation of newe tenninals in the crayfish opener muscle and its f unctional significance. J Gen Physiol 51 :731-758.
Bradacs H, Cooper RL, Msghina M, Atwood HL (1 997) Differential physiology and morpho10 y of phasic and tonic motor axons in a crayfish limb extensor muscle. J Ekp Bio 7 200:677-691.
Bryan JS, Krasne FB (1 97ï) Presynaptic inhibition: The mechanism of protection from habituation of the crayfish lateral giant fibre response. J Physiol (Lond) 271 ~369-390.
Chad JE, Eckert R (1 984) Calcium domains associated with individual channels can account for anomalous voltage relations of Ca-dependent responses. Biophys J 45:993-999.
Clernent JF, Taylor AK, Velez SJ (1 983) Effect of a limited target area on regeneration of specif ic neuromuscular connections in the crayfiçh. J Neurophys 49:2l6- 226.
Cooper RL, Hampson DR, Atwood HL (1995a) S aptotagmin-like expression in the motor newe terminais of crayfish. Brarir 4" es 703:214-216.
Cooper RL, Marin L, Atwood HL (1 995b) Synaptic differentiation of a single motor neuron: Conjoint definition of transmitter release, presynaptic calcium signds, and ultrastructure. J Neurosci 1 5:4209-4222.
Cooper RL, Stewart BA, Wojtowicz JM, Wang S, Atwood HL (1995~) Quantal measurement and analysis methoûs compared for crayfish and Drosophila neurom uscular junctions, and rat hippocam pus. J Neurosci Methods. 6 1 :66-79.
Cooper RL, Winslow JL, Govind CK, Atwood HL (1 996) Synaptic structural complexity as a factor enhancing probability of calcium-mediated transmitter release. J Neurophysiol75:2451-2466.
Coulthard R (1 998) The roles of rnotoneurons and their muscle targets in synaptogenesis during regeneration of a foreign transplant. MSc. thesis, University of Toronto, pp. 1-82.
Couteaux R, Pecot-Dechavassine M (1 970) Vesicles synaptiques et poches au niveau des "zones activew de la fonction neuromusculaire. Compt Rend Acad Sci Ser D 271 :2346-2349.
Dahm LM, Landmesser LT (1991) The regulation of synaptogenesis dunng normal development and following activity blockade. Dev Bi01 130:621-644.
Delaney KR, Zucker RS, Tank DW (1989) Calcium in motor nerve terminals associated with posttetanic potentiation. J Neurosci 9:3558-3567.
DeRosa RA, Govind CK (1978) Transmitter output increases in an identifiable lobster motoneurone with growth of its muscle fibers. Nature (Lond) 273:676- 678.
Dudel J, Kuffler SW (1 961 ) Presynaptic inhibition at the crayfish neuromuscular junctions. J Physiol (Lond) 1 55543-562.
Ely P, Velez SJ (1982) Regeneration of specific neuromuscular connections in the crayfish. 1. Pattern of connections and synaptic strength. J Neurophysiol 47~656-665.
Evoy WH, Beranek R (1 972) P hannacoiogical localization of excitatory and in hibito s aptic regions in crayfish slow abdominal flexor muscie fibres. Comp en $am 3:li&l86.
z Evoy WH, Kennedy D, Wilson DM (1 967) Discharge patterns of neurones supplying
tonic abdominal flexor muscles in the crayfish- J Exp Bi01 46:393-411.
Fatt P, Katz B (1 953) The effect of inhibitory nerve impulses on a crustacean muscle f iber. J Physiol (Lond) 1 21 :374-388.
Florey E, Cahiil MA (1982) The innewation pattern of crustacean skeletal muscle. Cell Tissue Res 224527-541.
Gilfary HL, Kennedy D (1 969) Pattem generation in a crustacean motoneuron. J Neurophysiol32:595-606.
Goda Y, Sudhof TC (1 997) Calcium regulation of neurotransmitter release: reliably unreliable? Current Opinion Ce11 Bi0 951 3-51 8.
Goelet P, Castellucci VJ, Schacher S, Kandel ER (1986) The long and short of long- term memory - a rnolecular framework. Nature 322:419-422.
Goransson LG, Hunt WP, Velez SJ (1 98) Regeneration studies on a crayfish neuromuscular system II. Effect of changing the nerve entry point into the muscle field on the gradient of innervation. J Neurobiol 1 9:let -1 52.
Govind CK, Atwood HL, Lang F (1973) Synaptic differentiation in a regenerating crab- f im b muscle. Proc Nat/ Acad Sci USA 70:822-826.
Govind CK, Pearce J, Wojtowicz JM. Atwood HL (1 994) "Strong" and "Wear synaptic differentiation in the crayfish opener muscle: structural correlates. Synapse 1 6~45-58.
Harrington CC (1 993) An investigation of structural relationships of the crayfish neuromuscular junctions. M.Sc. thesis, University of Toronto, pp. 1-92.
Heuser JE, Reese TS (1 979) S naptic vesicle exocytosis captured by quick f reezing . F In 'The Neurosciences ourth Study Program," F.O. Schmitt and F.G. Worden, Eds., MIT Press, Cambridge, MA, pp. 573-600.
Heuser JE, Reese TS, Dennis MJ, Jan Y, Jan L, Evans L (1979) Synaptic vesicle exocytosis captured by quick f reezing and correlated with quanta! transmitter release. J Ce11 Bi01 81 :275-300.
Hildebrand JG, Townsel JG, Kravitz €A (1 974) Distribution of acetylcholine, choline, choline acetyltransferase and acetylcholinesterase in regions and single identified axons of the lobster nervous system. J Neurmhem 23:951-963.
Hill RH, Govind CK (1 981 ) Comparison of phasic and tonic synaptic terminals in lobster muscle. Ce11 Tissue Res 221 :303-310.
Hirokawa N, Sobue K, Kanda K, Haraba A, Yorifugi H (1989) The cytoskeletal architecture of the presynaptic terminal and molecular structure of synapsin 1. J Ce11 Bi01 lOû:111-126.
Hong SJ, Lnenicka GA (1993) Long-terni changes in the neuromuscular synapses of a crayfish motoneuron produced by calcium influx. Brain Res 605:121-127.
Hong SJ, Lnenicka GA (1 995) ActMty-dependent reduction in voltage-dependent calcium cuvent in a crayfis h rnotoneuron. J Neurosci 1 535394547.
Hoy RR (1 969) Degeneration and regeneration in abdominal flexor motor neurons in the crayfish. J EXp Zoo1 1 72:219-232.
Hubbard JI, Llinas R, Quastel DMJ (1 969) Electrophysiological Analysis of Synaptic Transmission. Edward Arnold: London.
Hunt W P, Velez SJ (1982) Regeneration of specific neuromuscular connections in the crayfish II. Effect of changes in the target area. J Neurophys 47:666-676.
Hunt WP, Velez SJ (1989a) Regeneration of an identifiable motoneuron in the crayfish 1. Pattems of reconnection and synaptic strength established in normal and altered target areas. J Neurobiol20:703-717.
Hunt WP. Velez SJ (1989b) Regeneration of an identifiable motoneuron in the crayfish II. Patterns of reconnection and synaptic strength established in presence of an extra nerve. J Neurobiol20:718-730.
l ravan i J (1 965) Mem branedepolarisation der Muskelfasern des Offnerm uskels des Flusskrebses auf Nervenreiz und Kaliumapplikation. Expenentia 21 :609-610.
Jahromi SS, Atwood HL (1 974) Three-dimensional ultrastructure of the crayfish neuromuscular apparatus. J Ce11 Bi01 63:599-613.
Jessel TM, Kandel ER (1993) Synaptic transmission: a biochemical and self modifiable forrn of cell-cell communication. Ce1172 / Neuron 10 (suppl):l -30.
Katz B (1966) Nerve, Muscle and Synapse. New York: McGraw-Hill.
Kennedy D, Bittner GD (1 974) Ultrastructural correlates of motor axon nerve regeneration in crayfish. CeIl Tissue Res 148:97-110.
Kennedy O, Takeda K (1965a) Reflex control of abdominal flexor muscles in the crayfish. 1. The twitch system. J Exp Bi01 43321 1-227.
Kennedy D, Takeda K (1965b) Reflex control of abdominal flexor muscles in the crayfish. II. The tonic system. J Exp Bi01 43:229-246.
King MJR, Atwood HL, Govind CK (1 996) Structural features of crayfish phasic and tonic neuromuscular terminais. J Comp New 372:618-626.
Krause KM, Velez SJ (1 995) Regeneration of neuromuscular connections in crayfish allotransplanted neurons. J Neurobiol27:154-1 71.
Krause KM, Pearce J, Govind CK (1998) Regeneration of phasic motor axons on a crayfish tonic muscle: neuron specifies synapses. J Neurophysiol80:994-997.
Krause KM, Pearce J, Velez SJ, Govind CK (1996) Structure of allotransplanted
ganglia and regenerated neuromuscular connections in crayfish. J Neurobid 30:439-453.
Kuffler SW, Yoshikami D (1975) The number of transmitter mofecules in a quantum: an estimate from iontophoretic application of acetylcholine at the neuromuscular synapse. J Physiol (Lond) 251 :465-482.
Linder TM (1 974) The accumulative properties of facilitation at crayfish neuromuscular synapses. J Physiol (Lond) 238223,234.
Llinas R (1 977) In Approaches to the Cell Biology of Neurons, W.M. Cowan and J.A. Ferendelli, Eds. (Society for Neuroscience, Behtesda, MD) vo1.2, pp. 139-1 60.
Uinas R, Sugimori M, Silver RB (1992) Microdomains of high calcium concentration in a presynaptic terminal. Science 256:677-679.
Lnenicka GA, Atwood HL (1 985a) Agedependent long-term adaptation of crayfish phasic motor axon synapses to altered activity. J Neurosci 5:459-467.
Lnenicka GA, Atwood HL (1 985b) Long-terni facilitation and long-temi adaptation at synapses of a crayfish p hasic motoneuron. J Neurobiol 1 6:97-110.
Lnenicka GA, Marin L, Atwood HL (1 986) Morphological transformation of synaptic tenninals of a phasic motoneuron by long-term tonic stimulation. J Neurosci 62252-2258-
Lnenicka GA, Hong SJ, Combatti M, LePage S (1 991) Activity-dependent development of synaptic varicosities at crayfish motor teminals. J Neurosci 1 1 :1040-1048.
Manilow R, Otmakhov N, Blum KL, Lisman $ (1994) Visualking hippocampal synaptic function b optical detection of Ca ' entry through the Nmethyl-0-aspartate channel. 6 rocNatlAcadSciUSA91:8170-8174-
Meiss DE, Govind CK (1 979) Regional differentiation of neuromuscular synapses in a lobster receptor muscle. J Ekp Bio! 79:99-114.
Msghina M, Charlton MP, Atwood HL (1 995) Differentiation of transmitter release properties and calcium transients in phasic and tonic motor nerve endings of crustacea. Soc Neurosci Abstr 21 : 1 38.1 5.
Msghina M, Govind CK, Atwood HL (1 998) Synaptic structure and transmitter release in crustacean phasic and tonic motor neurons. J Neurosci l8:lW4-l382.
Msghina M, Millar AG, Charlton MP, Govind CK, Atwood HL (1 999) Calcium entry related to active zones and differences in transmitter release at phasic acd tonic synapses. Ce11 Mol Neurosci In press.
Nguyen PV, Atwood HL (1990) Expression of long-terni adaptation of synaptic transmission requires a critical period of protein synthesis. J Neurosci 10:lOgg- 1109.
Nguyen PV, Atwood HL (1994) Altered impulse activity modifies s naptic physiology and mitochondria in crayfish phasic motor neurons. J europhysiol72:2944- 2955.
hl'
Nordlander RH, Sin er M (1972) Uectron rnicroscopy of severed motor fibers in the crayfish. Z 1 elttotsch 1 26:157-181.
Otsuka M, lversen LL, Hall ZW, Kra* EA (1966) Release of amma-aminobutyric acid frorn inhibitory nerves in lobster. Proc Nat1 Acad 8 ci USA 56:1110-1115.
Pamas 1, Atwood HL (1 966) Phasic and tonic neuromuscular s stems in the abdominal
723. 6 flexor muscles of the crayfish and rock lobster. Comp iochem Physiol18:701-
Propst JW, Ko CP (1 987) Correlations between active zone ultrastructure and synaptic function studied with freeze-fracture of physiologically identified frog neuromuscular junctions. J Neurasci 7:3654-3664.
Pumplin DW, Reese TS, Llinas R (1981) Are the resynaptic membrane particles the calcium channels? Proc Nat1 Acad Sci U 8 A 78:7210-7213.
Quigley PA, Cooper RL, Govind CK, Atwood HL (1 996) Recruitrnent of active synapses at the cra ish neuromuscular junction visualized with the fluorescent dye FM 1 -43. Soc C eurosci Abstr 22:309.9.
Renger JJ, Atwood HL, Wu CF (1997) Single-bouton rewrding from the larval neurornuscular junction of Drosqphila CAMP cascade mutants. 38m Annual Drosophila Research Conference.
Rheuben MB (1 985) Quantitative comparison of the structural features of slow and fast neuromuscular junctions in Manduca. J Neurosci 31704-1716.
Roberts WM, Jacobs RA, Hudspeth AJ (1990) Colocalization of ion channels involved in frequency selectivity and synaptic transmission at presynaptic active zones of hair cells. J Neurosci 10:3664-3684.
Robitaille R, Adler EM, Charlton MP (1990) Strategic location of calcium channels at transmitter release sites of frog neuromuscular synapses. Neuron 5773-779.
Selverston Al, Remler MP (1 972) Neural geometry and activation of crayfish fast flexor motoneurons. J Neurophysiol35:797-814.
Simon SM, Llinas RR (1 985) Compartmentalization of the submembrane calcium activity during calcium influx and its significance in transmitter release. Biophys J 48:485-498.
Smith SJ, Augustine GJ, Charlton MP (1985) Transmission at voltage-clarnped giant s napse of the squid: evidence for cooperativity of presynaptic calcium action. &oc Nat1 Acad Sci USA 82622-625.
Smith SJ, Buchanan J, Osses LR, Chariton MP, Augustine GJ (1993) The spatial distribution of calcium signals in squid presynaptic terminals. J Physiol (Lond) 472 1573-593.
Stewart BA, Atwood HL (1 992) Synaptic plasticity in a regenerated crayfish phasic motoneuron. J Neurobiol23:881-889.
Stewart BA, Schuster CM, Goodman CS, Atwood HL (1996) Homeostasis of synaptic transmission in D m h i l a with genetically altered newe terminal morphology. J Neurosci 1 630774886.
Takeuchi A (1 976) Studies of inhibitory effects of GABA in invertebrate nervous systems. In =GABA in Newous S stem Functionw (E. Roberts, T.N. Chase, and D.B. Tower, eds.), pp. 255-267. A aven, New York.
Takeuchi A, Takeuchi N (1965) Lacalized action of gamma-butyric acid on crayfish muscle. J Physiol (Lond) 1 77:225-238.
Uchizono K (1 967) lnhibitory s apses on the stretch receptor neurone of the crayf is h. Nature (Lon J" ) 207:642-643.
Velez SJ, Wyman RJ (1 978a) Synaptic connecüvity in a crayfish neuromuscular system. 1. Gradient of innervation and synaptic strength. J Neurophysiol 41 :75-84.
Velez SJ, Wyman RJ (1 978b) Synaptic connectivity in a crayfish neuromuscular system. II. Neive-muscle matching and neive branching patterns. J Neurophysiol41:85-96.
Walrond JP, Govind CK, Huestis SE (1 993) Two structural adaptations for regulating transmitter release at lobster neuromuscular synapses. J Neurosci 13:4831- 4845.
Walrond JP, Reese TS (1985) Strucutre of axon terrninals and active zones at synapses on lizard twitch and tonic muscle fibers. J Neurosci !%Il 1 8-1 1 31.
Wiens TJ (1 982) Srnall systems of neurons: Control of rhythmic and reflex activity. In The Biology of Crustacean (H.L. Atwood and D.C. Sandeman, eds.), Vol. 4, pp. 1 93-240. Academic Press, New York.
Wilson DM, Davis WJ (1965) Newe impulse pattems and reflex control in the rnotor system of the crayfish cîaw. J &p Bi01 43:193-210.
Wojtowicz JM, Mann L, Atwood HL (1994) Activity-induced changes in synaptic release sites at the crayfish neuromuscular junction. J Neurosci 14:3688-3703.
Wojtowicz JM, Smith BR, Atwood HL (1 991) Activity-dependent recruitment of silent synapses. Ann NY Acad Sci 627:169-179.
Zhong Y, Wu CF (1 991) Altered synaptic plasticity in Drosophila memory mutants with a defective cyclic AMP cascade. Science 251 : 1 98-201.
Zucker RS (1 973) Changes in the statistics of transmitter release during facititation. J Physiol (Lond) 229:787-810.
Zucker RS, Delaney KR, Mulkey R, Tank DW (1991) Presynaptic calcium in transmitter release and posttetanic potentiation. Ann NY Acad Sci 635:lgl- 207.
Zucker RS, Fogelson AL (1 986) Relationship between transmitter release and presynaptic calcium influx when calcium enters through discrete channels. Proc Nat1 Acad Sci USA 83:3032-3036.
Quantitative analysis of regenerated excitatory newe terminais to the crayfish superficial flexor muscle upon allotransplantation of a tonic superficial flexor nerve. Surgeries perfomed by Kristen Krause, and data collected by Joanne Pearce and Rahim Hirji.
TONIC REGENERATE
TERMINALS:
Mean cross-sectional 2.67 area @m2) (x I sd) I 2.28
Oh COMPOSITION OF:
CIear vesicles 22.1
Dense vesicles 1.7
Axoplasm 57.2
SYNAPSES:
Number
Mean area ( p 2 ) (X + sd) ~urnber/~m= of terminal
DENSE BARS:
Number
Mean length (pm) (X f sd) (n)
Nurnber/synapse (X t sd)
~urnber f~ rn~ of terminal
~ e n ~ t h l p m ~ of terminal (P)
PAIRED DENSE BARS:
% of paired dense bars 6.0 (separated by I 0.2 pm)








































































































![Multiscale modeling, stochastic and asymptotic approaches ... › articles › proc › pdf › 2014 › 04 › proc144703.pdfislands with many synaptic recurrent connections [29,51].](https://static.documents.pub/doc/80x56/5f04b3557e708231d40f45ae/multiscale-modeling-stochastic-and-asymptotic-approaches-a-articles-a-proc.jpg)


