

STRUCTURE OF REGENERATED SYNAPTIC CONNECTIONS OF ALLOTRANSPLANTED PHASIC MOTONEURONS ON A TONlC
MUSCLE IN CRAYFISH
A thesis submitted in conformity with the requirements for the degree of Master of Science,
Graduate Department of Zoology, University of Toronto
O Copyright by Rahirn Hirji, 1999

National Library l*I , cana- Bibliithèque nationale du Canada
Acquisitions and Acquisitions et Bibliographie Services services bibliographiques
395 Wellington Street 395. me WdCinm OnawaON K1AûN4 OttswaON K I A W canada Canada
The author has granted a non- exclusive licence aiiowing the National Library of Canada to reproduce, loan, distriiute or seii copies of this thesis in microfom, paper or electronic formats.
The author retains ownership of the copyright in this thesis. Neither the thesis nor substantial extracts fiom it may be printed or otherwise reproduced wîthout the author's permission.
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque nationale du Canada de reproduire, prêter, distribuer ou vendre des copies de cette thèse sous la forme de microfiche/film, de reproduction sur papier ou sur format électronique.
L'auteur conserve la propriété du droit d'auteur qui protège cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.

STRUCTURE OF REGENERATED SYNAPTIC CONNECTIONS OF ALLOTRANSPLANTED PHASIC MOTONEURONS ON A TONlC MUSCLE IN
CRAYFISH
An abstract submitted in confonnity with the requirements for the degree of Master of Science in Zoology,
Graduate Department of Zoology, University of Toronto
Rahim Hi ji, 1999
ABSTRACT
Crustacean neuromuscular systems are highly defined into phasic and tonic
types exemplified by the deep and superficial flexor muscfes in the abdomen of the
crayfish Procambarus clarkii. The deep phasic muscle is used occasionally and briefly
for escape responses while the superficial tonic muscle is used continuously for routine
movements and maintaining posture. To assess the relative importance of the
motoneuron and the target muscle in detennining the synaptic phenotype of
regenerating neuromuscular connections, we attempted a mismatch experiment in
which the denervated tonic SFM received an allotransplanted phasic flexor newe. A
quantitative study of various structural parameters of these regenerated synapses
using serial section electron microscopy revealed that these synapses, on average,
possessed two dense bars that were often closely spaced. Mitochondria comprised a
relativeIy small (6%) volume of the tenninals- Collectively, these structural features
resemble those of native phasic synapses, thereby corroborating the physiological
evidence for the regeneration of phasic type synapses. Therefore, the motoneurons
and not their target muscle fibers are most influential in specifying the phenotype of
regenerated synapses.

ACKNOWLEDGMENTS
I would like to begin by expressing humble gratitude to my supervisor C.K. Govind,
who comrnand of knowledge, dedication and meticulous nature were integral in guiding
me to do my best. I would also like to thank Joanne Pearce for her patience and
willingness to help me whenever I was in need. Our mutual interest in stock markets
has helped fonn a bond that I hope will hold strong over time. 1 woold also like to thank
Asheer Sharman for his friendship, understanding and support in many aspects of my
life. Honourable mentions go out to Naz, Matt and Raymond. I would like to express
my deep gratitude to my family (Azim, Yasrnin, and Adil) for their ongoing motivational
support and for understanding my idiosyncrasies, and Fenulla for being there for me
and loving me for who i am; having al1 of you in my life makes me extremely fortunate.

TABLE OF CONTENTS
...................................................... I . Crustacean Neuromuscular Systems 1 A . Differentiation into excitatory and inhibitory axons ......................... 2
.................................... B . Differentiation into phasic and tonic axons 4
.................................... C . Synaptic differentiation within a tonic axon 8
................................... D . Structural correlates of transmitter release 10
............................................... II . Crustacean Neuromuscular Regeneration 12 A . Regeneration studies in the SFM ....................... ..... .............. 13 B . Neural allotransplantation in the SFM ......................................... 15
. . .................................................................... III . Experimental Objectives 17
MATERIALS AND METHODS ................................................................. 19
............................................................. I . Allotransplantation Procedure 19
II . Electron Microscopy ......................................................................... 20
...................................... III . Quantitative Analysis ...................... .. ... ... 21
IV . Volumetric Analysis ........... .... ..................................................... 23
RESULTS ........................................................................................... 25
I . Excitatory Innewation ................. ... .................................................. 25 A . Nerve terminais ........................................................................... 25 B . Synapses .................................................................................. 30 C . Dense bars ................................................................................. 32
............................................... II . Inhibitory Innervation . ...... ........... .... 40 A . Nerve terminais ............. .... ...................................................... 40 B . Synapses ................................................................................... 43 C . Dense bars ............................................................................... 43 D . Presynaptic inhibition ................................................................... 49
.............................................................. DISCUSSION ....................... .. 51
................................................ I . Nature of Regenerated Nerve Terminais 51 ............................................................ A. Nerve terminal morphology 52
B . Mitochondrial volume .................................................................... 53 ............................................................... C . Dense bars per synapse 54

Structural-Functional Correlations ........................................................ Synapses .................................................................................. a . area ..................................................................................... b . number .................................................................................
Dense bars ................................................................................ a . length ........................ ... ...................................................... b . number ................................................................................
................................................................................ . c spacing
Other factors involved ..................................................................
lnhibitory Innervation ........................................................................
REFERENCES ........................................................................................ 75
APPENDIX ................................................ ...... ...................................... 82

Nerve allotransplantation has ernerged as a useful method for exploring how
motoneurons are capable of fonning specific connections with target cells in the crayfish
Procambarus clarkii (Krause and Velez, 1995). More recently, allotransplantation has
provided an opportunity to test whether the motoneuron or the target muscle detemines
the nature of the regenerating synaptic connections (Coulthard, 1998; Krause et al.,
1998). The current study examines regenerated neuromuscular terminals that fomied
after allotransplantation of a foreign phasic flexor nenre to the tonic superficial flexor
muscle (SFM) in the crayfish abdomen.
One of the primary aims of this study is to determine if ultrastructural
features can be correlated with the large initial EPSPs recorded by Krause et al. (1998)
from the regenerated connections. Another objective is to determine if the regenerated
terminals are phasic in nature, as suggested by the electrophysiology. Through the
exploration of these objectives, I will be able to detennine if the motoneuron or the target
muscle specifies the nature of the regenerating synaptic contacts. I will first provide the
relevant background material related to the crustacean neuromuscular system, paying
particular attention to the types of innervation present, how they are differentiated, and
their function in their respective systems. Next, crustacean neurornuscular regeneration
in the SFM systern will be explored.
1. CRUSTACEAN NEUROMUSCULAR SYSTEMS
One of the most important reasons for the popularity of the crustacean
neuromuscular system in experimental research is that it serves as a good model of the
vertebrate CNS (Atwood, 1976) since the two share key similarities. First, both a n
receive innervation from inhibitory and excitatory neurons, and these two types of

neurons can interact with one another. Second, the cnistacean muscle fiber, like the
vertebrate CNS, has numerous synapses owing to multiteminal innervation. Partiy
because of their heuristic value and partly because of their intrinsic interest, crustacean
neuromuscular systems have received much sc~ t iny by researchers over the past 50
years; the resulting information has been reviewed primarily by Atwood and his
collaborators (Atwood, 1976, 1982; Atwood and Govind, 1990; Atwood and Wojtowicz.
1986; Atwood and Cooper, 1996).
One striking aspect of the cnistacean neuromuscular system is its highly
differentiated nature as reflected in the following properties; (i) there are both excitatory
and inhibitory axons innnewating a muscle, (ii) there are two broad categories of
excitatory and inhibitory axons, phasic and tonic, and (iii) there are differences in
transmitter output capabilities in populations of synapses of single excitatory tonic
axons. l will briefly address each of these aspects as background for studies on the
regeneration of crustacean neuromuscular systems.
A. Differentiation into excitatory and inhibitory axons
Motoneurons can be classified as either excitatory or inhibitory, depending
on the response they elicit in the muscle fibers they innervate (Katz, 1966). While
excitatory motoneurons release the transmitter L-glutamate and bring about contractions
of the muscle fiber by depolarking its membrane potential, inhibitory neurons prevent
contraction by holding the membrane potential at resting level or by hyperpolarizing it
(Atwood, 1976). Stimulation of an inhibitory axon triggers the release of the transmitter
gamma-aminobutyric acid (GABA), which interacts with postsynaptic receptors and
increases chloride conductance. As a result, the membrane potential of the muscle fiber
moves toward the equilibnum potential of chloride (Fatt and Katz, 1953). and

counteracts the depolarizing effect of the action potentials in contracting muscle fibers,
thereby inhibiting muscle contraction (Parnas and Atwood, 1966).
Inhibitory neurons make synaptic contact with the muscle fiber membrane
and bring about postsynaptic inhibition. In addition, some inhibitory neurons make
synaptic contact with the excitatory axon and bRng about presynaptic inhibition (Atwood,
1976). Such peripheral presynaptic inhibition is represented by polarized axo-axonal
contacts from an inhibitory nerve terminal onto an excitatory newe terminal or axonal
region and has been obsewed in many crustacean neuromuscular systems (Atwood
and Morin, 1970; Atwood, 1982). The inhibitory transmitter GABA, which is responsible
for postsynaptic inhibition by inducing an increase in chloride conductance, is also
responsible for presynaptic inhibition through a similar process (Takeuchi, 1976). As a
result, fewer quanta of L-glutamate are released per excitatory impulse (Dudel and
Kuffler, 1961), which decreases the extent of depolarization of the muscle membrane,
and yields a smaller muscle contraction. Presynaptic inhibition can lessen transmitter
release by hyperpolarizing or depolarizhg the excitatory nerve terminal, reducing the
size of the excitatory action potential, or hindering impulse propagation in the excitatory
axon (Atwood and Wojtowicz, 1986).
lnhibitory innervation is present in most crustacean muscles (Atwood and
Wojtowicz, 1986) and appears to share some key similanties with the prevailing
excitatory innervation. The differences in transmitter release and facilitation
characteristics of an inhibitory axon innervating different muscle fibers, as well as
different inhibitory axons innervating the same muscle fiber, minor the differences
observed with excitatory innervation (Atwood, 1976). Atwood and Bittner (1971) have
suggested that the physiological characteristics of excitatory and inhibitory synapses are
similar for a given muscle fiber, supporting the idea that the factors responsible for the

differentiation of excitatory synapses are also responsible for sirnilar differentiation in
in hibitory synapses (Atwood, 1 976).
Both excitatory and inhibitory innervation are found in crustacean
neuromuscular systems. Excitatory axons are known to act postsynaptically, whereas
inhibitory axons can act both pre- and post-synaptically. These two types of
rnotoneurons can be further categorized into phasic and tonic varieties, depending on a
number of characteristics
B. Differentiation into phasic and tonic axons
Excitatory rnotoneurons supplying many crustacean limb muscles were
classified as either Tasr or uslow", depending upon the type of muscle contraction they
induced when the motoneuron was stimulated (Atwood, 1976). Research on the fast
flexor and fast extensor neurons of crayfish and lobster abdominal muscles (Kennedy
and Takeda, 1 965 a,b; Parnas and Atwood, 1966; Selverston and Remler, 1972) helped
develop a phasic / tonic dichotomy for excitatory motoneurons. Phasic and tonic
motoneurons Vary in a number of properties, including baseline activity, size,
morphology, response to stimulation, and plasticity. Phasic motoneurons (such as the
ones supplying the fast abdominal extensor and flexor muscles in crayfish and lobsters)
are filiform in morphology and are considered to be relatively silent, since they fire quite
inf requently. W hen they do fire, however, a significant arnount of transmitter is
reieased; this tendency accounts for the rapid and strong muscle contraction (referred to
as a twitch contraction) obsewed in fibers innenrated by phasic axons. Thus, phasic
axons are useful when vigorous, imrnediate activity is necessary (Atwood and
Wojtowicz, 1986; Atwood and Govind, 1990; Atwood and Cooper, 1996). However, this

levei of contraction cannot be maintained due to rapid fatigue at the neuromuscular
synapses of these axons during continuous stimulation (Atwood and Wojtowicz, 1986).
Tonic axons (such as the ones supplying the slow abdominal extensor and
flexor muscles), on the other hand, are varicose in morphology and are considered fairîy
active since they usually fire continuously at a low frequency of stimulation (Atwood and
W ojtowicz, 1 986). Initially, tonic axons release less transmitter but their synapses
exhibit greater fatigue resistance than their phasic counterparts. In addition, tonic axons
are capable of facilitating transmitter retease at higher firing frequencies. Tonic axons
typically control routine movements and posture (Kennedy and Takeda, 1965a.b;
Atwood and Wojtowicz, 1986). Subtle changes in tonic axon firing frsquency are
capable of slightly modifying movements. The axons supplying the slow superficial
flexor muscle of the crayfish abdomen are tonic axons which have been frequently
studied (Kennedy and Takeda, 1965 a,b; Evoy et al., 1967).
Recent studies have tried to correlate the physiological characteristics of
phasic and tonic neurons with their respective morphological characteristics (Msghina et
al., 1998, 1999). The physiological differences can be accounted for by either
presynaptic or postsynaptic disparities. Since it is believed that presynaptic factors are
the most important physiological determinants, researchers have focused primarily on
the presynaptic region. Examinations of the crayfish claw closer muscle have shown
phasic axons to be thin and non-varicose, and tonic axons to be thick and varicose
(Lnenicka et al., 1986, 1991). Furaiemore, it was shown that phasic terminals house
uniformly distributed synapses, whereas tonic teminals house synapses which are
found primarily on the varicosities (Hill and Govind, 1981; Lnenicka et al., 1986; Florey
and Cahill, 1982). A surprising finding in these studies was that tonic terminals have
more synapses than phasic terminals. This is contrary to what one would expect given

the physiological characteristics of the N o types of neurons. Therefore, there must be
some other presynaptic factor contributing to the difference. One difference identified
by investigators comparing phasic and tonic tenninals of the crayfish Iimb extensor
muscle is the amount of dense bar material found at the synapses (King et al., 1996).
Although fewer in number, phasic synapses were shown to have more dense bar
material than the tonic synapses; the phasic synapses were found to house longer
dense bars and a greater number of dense bars.
Not al1 excitatory motoneurons can be easily categonzed as either phasic or
tonic; some axons exhibit properties of both categories. For instance, the largest axon
that innervates the slow flexor muscles in the crayfish abdomen fires in bursts and
exhibits very little background activity, both of which are features characteristic of phasic
axons (Kennedy and Takeda, 1965b; Gillary and Kennedy, 1969). However, there is
considerable facilitation and little neuromuscular fatigue seen in this axon; these are
features characteristic of tonic axons. Clearly, this motoneuron possesses intenediate
properties, as do many other crustacean motoneurons (Atwood, 1976; Atwood and
Wojtowicz, 1986).
Inhibitory motoneurons, like their excitatory counterparts, can also be
classified on a phasic 1 tonic spectrum; inhibitory axons to the crayfish fast abdominal
muscles appear phasic (like their excitatory counterparts), while the inhibitory axon to
the crayfish opener muscle is considered tonic (Wilson and Davis, 1965), and the
inhibitory axon to the crayfish slow flexor muscles has intennediate properties (Kennedy
and Takeda, 1965 a,b; Evoy et al., 1967). Further research is required, however, to
determine whether inhibitory neuromuscular junctions exhibiting phasic properties
experience rapid fatigue similar to their excitatory counterparts.

Long terni adaptation studies show that sustained increases in electrical
activity of a phasic motoneuron alter its physiology and rnorphology to that of a tonic
motoneuron. There are a number of proteins that may play a role in the conversion of
phasic motoneurons; these same proteins may also be responsible for the obsewed
inherent differences between phasic and tonic nerve terminals. For example, both the
initial reduction in EPSP amplitude and the increase in fatigue resistance could anse
from a decrease in the number of active calcium channels in crayfish nerve terminals
(Atwood and Cooper, 1996). Studies in the moth, Manâuca, and in the lizard, Anolris,
have demonstrated that the number of putative calcium channels at active zones is
lower in tonic than in phasic synaptic connections (Rheuben, 198-5; Walrond and Reese,
1985). Other proteins that rnay be altered include voltage sensitive sodium and
potassium channefs, vesicle associated proteins (Le. VAMPS, syntaxins, synaptotagmin,
and NSF), second rnessengers such as calcium calmodulin-dependent kinases, and
proteins associated with mitochondria (e.g. cytochrome oxidase proteins) (Atwood and
Cooper, 1996). Obtaining knowledge regarding the cellular mechanisms responsible for
regulating the transcription of mRNAs that are translated into proteins would enhance
Our understanding of the rnolecular differences between types of motoneurons (e.g.
phasic and tonic ones) with differing patterns of activity and the changes that result
during activity-dependent transformation (Atwood and Cooper, 1 996). Hence, molecular
differences may be partially responsible for quanta! output disparities between native
phasic and tonic synaptic connections.
Thus, although both excitatory and inhibitory axons can be classified as
either phasic or tonic, some motoneurons have intermediate properties. Long term
adaptation studies suggest that inherent molecular differences may help account for the
differences obsewed between phasic and tonic motoneurons. lnsig hts into the

correlates of synaptic output can be gained by exarnining muscle systems innervated by
a single tonic axon.
C. Synaptic differentiation within a tonic axon
It is a cornmon feature of a single tonic axon to fom neuromuscular
synapses that rekase dierent amounts of transmitter. The best examples of such
synaptic differentiation are the single excitatory axons to the crayfish opener muscle
(reviewed by Atwood and Wojtowicz, 1986) and to the lobster accessory flexor muscle
(reviewed by Govind and Walrond, 1989). The summary below is based fargely on
these studies.
Although individual crustacean muscle fibers are innewated by only a single
excitatory motoneuron, the synaptic potentials produced by neuronal stimulation Vary
both in their amplitude and in their ability to facilitate. In the limb accessory flexor
muscle of the American tobster, for instance. synapses generate larger junctional
potentials on distal muscle fibers than on proximal fibers. The former are referred to as
high-output synapses and exhibit little faciiitation, whereas the latter are low-output and
show considerable facilitation (DeRosa and Govind, 1978; Meiss and Govind, 1979).
Similarly, the excitatory motoneuron in the crayfish limb opener muscle foms regionally
differentiated high- and low-output synapses; high-output synapses are found on
proximal fibers while low-output synapses are found on more central ones (Atwood and
Bittner, 1 971 ; Bittner, 1 968; Linder, 1974). When exposed to low stimulation
frequencies, the larger initial EPSPs recorded in the proximal bundles are 8-10 times
larger than the ones recorded from the central fibers (Iravani, 1965; Cooper et al.,
1995b). Subsequent analysis has revealed that approximately 1/4 of this EPSP
disparity can be attnbuted to a higher input resistance in the proximal fibers. The

remainder of the EPSP disparity is likely due to diïerences in the probability of
transmitter release at synapses, including differences in calcium current and synaptic
cornplexity (Cooper et al., 1995b). The observed disparities between proximal and
central synapses in terms of junctional potential amplitudes and quantal content
positively correlate with discrepancies in dense bar number and length within the
synapses (Govind et a1.,1994). Zucker et al. (1991) showed that a single pulse elicits a
greater calcium flux for synapses housing long, or multiple dense bars as opposed to
small, simple synapses. Although the average dense bar is twice as long in proximal
than central synapses, quantal content is three to five times greater at the same low
frequency. This discrepancy indicates a non-linear relationship between dense bar
length and quantal output, which is in agreement with previous findings at other
crustacean synapses (Atwood and Marin, 1983). As a result, proximal muscle fibers in
the crayfish exhibit manifold larger EPSPs than their central counterparts.
The varicosities made by the excitatory motoneuron innervating the crayfish
limb opener muscle c m be visualized in living preparations with the aid of the dye 4-Di-
2-Asp (Magrassi et al., 1987). Once identified, the varicosities can be individually
contacted by a macropatch electrode for the purpose of recording synaptic currents and
labelling the varicosity with electron dense fluorescent beads. These varicosities can
then be serially sectioned, reconstructed and analyzed with the aid of a transmission
electron microscope (Wojtowicz et al., 1994). A cornparison of the innervation pattern of
different regions of the crayfish opener muscle indicates that on high-output proximal
fibers there are fewer varicosities and l e s terminal branching than on low-output central
fibers (Harrington, 1993; Govind et a1.J 994). Furthemore, quantal release is higher in
proximal terminals, which intuitively appears to disagree with the structural differences
between proximal and central terminals. Thus, it appears that transmission at the

neuromuscular juncüon is predominantly influenced by specific nerve terminal properties
rather than quantitaave differences at the macroscopic level (Govind et al., 1994). Such
an idea supports the use of electron microscopy for conducting ultrastructural analyses
through serial reconstruction in pursuit of the structural basis of transmitter release
differences.
The differentiation of a single tonic motoneuron innewating a group of
muscle fibers suggests that presynaptic properties are primarily responsible for
differential transmitter output and the resulting differential initial EPSP amplitudes at low
stimulation frequencies. Differences in the probability of release at synapses have been
explored primarily through an examination of calcium's involvement in the process of
synaptic transmission.
D. Structural correlates of transmitter release
The idea that calcium is integral to the process of synaptic transmission is
widely accepted. Depolarization induces calcium entry through voltage sensitive
calcium channels in nerve terminais, precipitating a release of neurotransmitter
molecules from spherical synaptic vesicfes (approximately 46-70 nm in diameter)
containing quanta (packets of 10' transmitter molecules) (Atwood. 1982). However. the
direct mechanism by which calcium ions induce a release of vesicles from the
presynaptic terminal has yet to be elucidated and is therefore the focus of much
research. It is clear that an influx of calcium sharply increases transmitter release
probability from the basal state, and that the probability of release is determined by the
mechanisms responsible for coupling the fusion apparatus to calcium ions (Goda and
Sudhof, 1997). In fact, it has been demonstrated that an increase in calcium
concentration can result in a geometric increase in the amount of transmitter released

up to the fourth power (Smith et al., 1985; Augustine and Chariton, 1986; Zucker and
Fogelson, 1 986).
Transmission electron micrographs depict an electron dense material,
known as a dense bar, with clustered synaptic vesicles on either side representing a
putative site of transmitter release, or active zone (Atwood, 1982; Heuser and Reese,
1979). A dense bar may contain cytoplasmic portions of various proteins responsible for
the movement of vesicles toward calcium channefs and release zones contained within
the synapse (Hirokawa et a1.,1989). Quanta are released from the presynaptic terminal
into the synaptic cleft upon fusion of the vesicle with the presynaptic plasmalemma at an
active zone (Katz, 1966; Kuffler and Yoshikami, 1975; Couteaux and Pecot-
Dechavassine, 1 970; Heuser and Reese, 1 979; Heuser et al., 1 979). Furthemore, the
fusion sites of synaptic vesicles are depicted as small circular depressions when
examined in freeze-fracture view (Pearce et al., 1986). The close proximity between the
large intramembranous particles and exocytotic vesicles, coupled with the fact that the
delay between calcium influx and transmitter release is brief, implicated the large
particles as being calcium channels (Pumplin et al., 1981). Given the brief delay, the
idea that more distant structures are directly involved in the release process can be
discounted. Furthermore, a very short latency (approximately 200 ps) between calcium
influx and post-synaptic response has been demonstrated in squid giant synapse
voltage clamp studies (Uinas, 1977). Such a short delay suggests that only vesicles
situated near the quantal emission domains are released when an action potential
occurs (Llinas, 1977). Robitaille et al. (1 990) used specific ligands to show that calcium
channel bands at active zones of the frog neuromuscular junction have locations
corresponding to the intramembranous particles. The active zone is not composed

solely of calcium channels; calcium-activated potassium channels are also present
(Roberts et al., 1990).
There are two key components involved in the calcium dependent control of
transrnitter release probability: calcium channels wtiich allow calcium to enter the
terminal, and calcium sensors present in the fusion machinery that are capable of
regulating exocytosis (Goda & Sudhof, 1997). During repetitive stimulation, calcium
sensitive indicators have further supported the idea that calcium channels are localized
at presynaptic active zones in the squid (Llinas et ai., 1992; Smith et al., 1993).
Recently, the calcium binding protein synaptotagmin, thought to be a putative calcium
sensor for evoked release, has been identified in crayfish motor neurons (Cooper et al.,
1 995a).
In summary, the location, size and number of the large intramembranous
active zone particles support the hypothesis that calcium channels couple an action
potential with transrnitter release (Pumplin et aI.,1 981 ; Simon and Llinas, 1985; Walrond
and Reese, 1985). As a result, the "structure-function hypothesisw has been proposed; it
postulates that the probability of transmitter release from a release site is influenced by
the number of calcium channels located in close proximity to the site (Walrond and
Reese, 1985; Atwood and Lnenicka, 1986). Such a hypothesis raises the possibility that
long term changes in transmitter release could be made via controlling the number
andfor distribution of dense bars present in the nerve terminal (Govind and Walrond,
1 989).
II. CRUSTCEAN NEUROMUSCULAR REGENERATlON
There are a few isolated studies that focus on the neuromuscular synaptic
regeneration obsented when limbs are removed and new ones form in their place. One

such study of the regenerating limb buds of crabs revealed that initial synaptic contacts
were poorly facilitating while later ones were highly facilitating for an identified tonic
motoneuron (Govind et al., 1973). A more recent study in regenerating crayfish clawed
Iimbs showed that the synaptic pfasticity of a phasic motoneuron did not recapitulate
features of its primary development (Stewart and Atwood, 1992). While these two
studies were performed using limb muscles, most studies of neuromuscular
regeneration have been conducted on a body muscle viz the superficial flexor muscle
(SFM) in the abdomen of crayfish. The ability of motoneurons to regenerate in this
system has allowed Velez and his callaborators to explore how axons are able to form
synaptic contacts with the target muscle (Ely and Velez, 1982; Hunt and Velez, 1982,
1 989a, b).
The crayfish abdomen has two very different neuromuscular systems; the
phasic flexor muscle is innervated by the phasic flexor nerve and is active during flight
responses, whereas the tonic superficial flexor nerve innervates the SFM and is
responsible for fine routine movements. The tonic SFM, with its well-known anatomy
and limited number of cells, offers advantages for examining synaptic regeneration.
Over the past two decades, Velez and his collaborators have conducted a series of
elegant studies on the regeneration of neuromuscular synapses on this muscle.
A Regeneration studies in the SFM
Regeneration studies may help us understand synaptogenesis and
neuromuscular interactions. They may aid in detemining whether functional synaptic
properties are a result of an autonomous program of the motoneuron, or are influenced
by retrograde influence from target muscle. There are studies supporting each of these
views. In the crayfish leg extensor muscle, the fact that both phasic and tonic murons

innervate the same target muscle fibers supports the notion that specific neuromuscular
properties are determined presynaptically (King et al., 1996). In the crayfish leg opener
muscle, however, the occurrence of high- and low-output endings of a single tonic
motoneuron on different muscle fibers suggests a target-specific influence on specific
neuromuscular properties (Cooper et al., 1995b).
It is well known that crayfish have excellent powers of regeneration, both of
entire body parts such as limbs as well as of tissues such as nerves. The latter in
particular has allowed the study of the regeneration of synaptic connections in the
superficial flexor muscle (SFM) of the abdomen by Velez and his collaborators. The
SFM consists of approximately 40 tonic muscle fibers arranged in a thin sheet (Velez
and Wyman, 1978a,b). It is considered a "slow" muscle, because of its long sarcomere
length, and graded tension throug h surnmation. The muscle receives polyneural
innervation from six axons in total; five of them provide excitatory innervation, white the
other one provides inhibitory innervation (Kennedy and Takeda, l96Sb). It is important
to note that not al1 excitatory axons have the same properties; some are more tonic than
others, and many of them fire spontaneously. Each SFM muscle fiber is innervated by
at least three axons; two of them are excitatory, and the other one is inhibitory (Kennedy
and Takeda, 1965b; Velez and Wyman, 1978a). Excitatory axons likely use the
transmitter L-glutamate (Evoy and Beranek, 1972; Hildebrand et al., 1974), whereas
inhibitory axons use GABA (Otsuka et al., 1966; Uchizono, 1967; Takeuchi and
Takeuchi, 1965).
The muscle fibers of the SFM are innervated in a position dependent
manner; each motoneuron consistently innewates certain fibers, and EPSP size varies
depending on the fiber's location (Velez and Wyman, 1978a,b). When the native
superficial flexor nerve was transected, connectivity maps were re-established within 8

weeks (Ely and Velez, 1982) via axon sprouts, or satellite axons, growing from the
proximal nerve stump (Krause et al., 1996). Such branching of motoneurons has also
been obsenred in vertebrates upon curare application to reduce neuromuscular a c t ~ t y
(Dahm and Landmesser, 1991). It is likely that since the muscle is not being stimulated
by the motoneurons innervating it, activity dependent retrograde signals induce axon
sprouting (reviewed by Jessel and Kandel, 1993). Thus, in the case of a regenerating
nerve and a denervated muscle, it is quite possible that muscle fibers signal axon
sprouting and reinnervation of denewated targets. Successful regeneration, however,
only occurred approxirnately half of the time (Ely and Velez, 1982). If the newe is cut
very close to the ventral cord, the nerve cannot regenerate because the distance that
the regenerating nerve must travel is too great. Regeneration is first obsewed at 3
weeks post-transection, at which point the transected distal nerve has degenerated and
is incapable of forming viable synaptic connections.
The specificity of regenerated synaptic connections was maintained even
after various experimental manipulations were performed, including a reduction in the
size of the muscle field (Clement et al., 1983) and a change in the nerve's entry location
into the muscle (Goransson et al., 1988). These studies strongly suggest the presence
of cues on the muscle surface that guide the formation of specific synaptic connections
by the regenerating axons. To further test the possibility of guidance by the target
muscle of the regenerating axons, transplanting the SFM nerve into an unusual location
was attempted (Krause and Velez, 1995).
B. Neural allotransplantation in the SFM
The SFM system is ideal for transplantation studies due to the small number
of muscle fibers (40) involved, the well-âocumented innervation of these fibers, and the

regeneration abilities of the rnotor axons to these fibers. A technique for
allotransplanting a SFM newe with its associated ganglia from one animal to the
denervated SFM of another was developed to test whether neurons are able to detect
their position within the target muscle during synaptogenesis, thereby producing a
position-dependent connectivity pattern (Krause and Velez, 1995). The regeneration of
neuromuscular contacts to a previously denewated muscle indicates that the
motoneurons in the transplanted ganglia survive, even though these ganglia are from
another crayfish and are placed in a foreign location without any physical connection to
the host animal's circulatory or nervous system (Krause et al., 1996). The transplanted
ganglia showed structural integrity; they had nucleated cell bodies, blood vessels and
lacunae (indicative of vascularization), healthy neuropil containing dendritic profiles with
synaptic contacts, as well as clear and dense vesicles.
Efectron micrographs of the transplanted newe reveal differences with the
contralateral intact newe (Krause et al., 1996). Specifically, the transplanted nerve is
larger than its intact counterpart, but has five axons over the lateral SFM like its intact
counterpart. Each of these f i e axons has several axon sprouts, rather than appearing
as single axon profiles (as in the intact condition). Axon sprouting is known to occur
from the cut central stump of the motoneuron to the crayfish opener muscle where these
satellite branches may either fuse with the distal stump (Kennedy and Bittner, 1974) or
grow out to the target muscle (Nordlander and Singer, 1972) in order to reconnect the
nerve to the muscle. Interestingly, although the donor nerve was transplanted on the
ventral surface of the SFM, it regenerated on the dorsal surface in a path resembling the
original host SFM nerve, and had twice the diameter when examined 8-10 weeks post-
transplantation (Krause et al., 1 996).

Viable neuromuscular connections were first detected 2 weeks after the
operation; innervation was observed across the entire muscle approximately 4 weeks
later (Krause and Velez, 1995; Krause et al., 1996). Not al1 muscle fibers, however,
were inneniated; the transplanted newe formed synapses on approximately 60% of the
muscle fibers with an innervation pattern very similar to the intact one. Thus, complete
innewation of target muscle is not required for specificity of reconnection following newe
allotransplantation (Krause and Velez, 1995). The specificity of connection of
allotransplants could imply that axons use targetderived positional information to create
similar connectivity maps (Krause and Velez, 1995). Since the regenerate terrninals
were often adjacent to the original degenerating nerve teminals, it is possible that the
new terminals follow the trail of the degenerating terminals to maintain specificity
(Krause et al., 1996).
III. EXPERIMENTAL OBJECTIVES
Recently, Krause et al (1998) used the allotransplantation technique to
mismatch the regenerating nenie to its target muscle. Specifically, they allotransplanted
the phasic branch of the third newe root to a denervated SFM. The transplanted ganglia
survived for 8-10 weeks and showed intact lwking cell bodies and neuropil. The phasic
nerve regenerated ont0 the dorsal surface of the SFM and the axons showed a large
number of sprouts, between 10-50 for individual axons. The large number of sprouts
was also seen for phasic axons in culture (Arcaro and Lnenicka, 1995) and contrasted
sharply with the much fewer sprouts that tonic axons showed in allotranspfanted and
culture conditions. Hence a phasic nerve appeared to have successfully regenerated
ont0 the tonic SFM muscle.

Electrophysiological examination of the neuromuscular synapses formed by
these regenerated phasic axons displayed some very large (25 mV) EPSPs, which are
reminiscent of phasic synapses. Therefore, phasic type synapses appeared to have
regenerated ont0 a tonic muscle, raising the possibility that the neuron rather than its
target muscle regulates the type of synapses that regenerate.
My primary objective was to examine the fine structure of these regenerated
connections via thin serial section electron microscopy which would provide a
quantitative description of the regenerated synapses and their dense bar active zones.
Such a description would allow me to correlate structure with physiology and assess
whether the observed ultrastructural features were capable of accounting for the very
farge initial EPSPs.
A description of the fine structure of these regenerated neuromuscular
connections would also allow me to compare them with other known phasic and tonic
neuromuscular connections in crayfish. This com parison would hopef ully address the
debate regarding whether the motoneuron or the target muscle is most influential in
determining the type of synaptic connections made between a mismatched foreign
phasic nerve and native tonic muscle. If regenerated synapses are deemed to be phasic
in nature, it can be suggested that the motoneuron is most influential in deterrnining the
type of synaptic connections made between a foreign phasic nerve and a native tonic
muscle.

MATERIALS AND METHODS
1. ALLOTRANSPàANïAWON PROCEDURE
The allotransplantation procecure outlined below was perfomed by Kristen
Krause using 15 donor and 15 host crayfish. Adult crayfish, Procambarus clarkri; were
purchased from a Louisiana supply company and held in aerated freshwater tanks in
individual compartments at 22 OC. The host SFM in the third abdominal segment was
denewated by severing the superficial flexor newe of the third root proximal to the
medial edge of the muscle (Clement et al., 1983). Since the newe was cut close to its
point of emergence from the CNS, the regeneration of this nerve was effectively
prohibited (Hoy, 1969). After a penod of 48 hours, these denervated animals were
allotransplanted with a donor flexor newe using a minimally invasive allotransplantation
procedure developed by Krause and Velez (1995). After donors were sacrificed, the
nerve cord was exposed by the removal of the deep flexor and extensor muscles. The
third ganglion and its third root with a long segment of the deep branch which supplies
the phasic muscles, were transplanted along with the fourth ganglion and the connecting
nerve cord to reinnervate the SFM by a phasic nerve.
The transplant obtained from the donor animal was introduced into the
abdominal cavity of the host animal through a hole forrned by removing the left
swimmeret of the third abdominal segment (Krause and Velez, 1995). In order to
accomplish the delicate task of positioning the phasic nerve on the ventral surface of the
host SFM, a tiny needle hole was made in the ventral cuticle irnmediately underneath
the SFM, A human hair was threaded through this hole and made to emerge from the
swimmeret opening and was attached to the deep phasic newe with surgical thread.
The hair was then pulled back out of the hole so that the transplanted newe came to lie

against the SFM, since such an arrangement is necessary for neurornuscular
regeneration to proceed.
A total of 15 crayfish underwent the allotransplantation procedure; five of
these host animals died, and the remaining 10 animals suwived (Krause et al., 1998).
These were examined electrophysiologically 8-1 0 weeks later and eight of these animals
exhibited characteristics of reinnervation, in the form of EPSPs, in response to
stimulation of the regenerate nerve. Five of these animals were used for the present
electron microscopie investigation of regenerated neuromuscular synaptic connections.
il. ELECTRON MICROSCOPY
The following is an outline of the procedure followed by Joanne Pearce to fix
the neuromuscular tissue once the allotranplanted newe regenerated. The SFM tissue
was prepared for electron microscopy using procedures standard to our laboratory
(Govind et al., 1994). The SFM preparations were initially fixed in situ for one hour at
room temperature by superfusing in a prirnary fixative containing 2.5% glutaraldehyde,
0.5% fonnaldehyde, 1 mM calcium chloride, and 0.1M sodium cacodylate buffer (pH
7.4). Next, the ganglion, nerve, and muscle were removed and fixed for an additional
hour in an identical fixative. The tissue was then washed for one hour in a O.lm sodium
cacodylate buffer containing 4 % sucrose and 1 mM CaCI2. The tissue was postfixed for
one hour in 2% OsOI in a O.lm cacodylate buffer, rinsed briefly in buffer before being
dehydrated using a graded ethanol series. Following dehydration, the tissue was placed
for 30 minutes in propylene oxide and left immersed in a 50% propylene oxide - 50°h
Epon-Araldite mixture overnight to allow the resin to infiltrate gradually. The following
day, the tissue samples were placed in moulds of fresh Epon-Araldite. After remaining

at room temperature for eight hours, the moulds were placed in a 60 OC oven for 48
hours for curing.
After the tissue was fixed and embedded in resin, I began my project by
cutting serial thin sections of the tissue using a diamond knife mounted on a Reichert
OMU 2 ultramicrotome. The thickness of each serial section was estimated from its
interference color (the wlor of the reflection of the section as it floated on water).
Successive sections adhered to one another forming ribbons of sections that were
transferred ont0 single slot gnds and placed ont0 Fornivar-coated slotted gridstands.
After a drying period, the gnds holding the serial sections were stained with uranyl
acetate for 30 minutes, and then lead citrate for two minutes. Sections were then
examined for nerve terminal regions using a Zeiss 9s electron microscope. Areas of
interest were photographed in serial (final magnification approximately ~27,000) and
analyzed both quantitatively and qualitatively.
Ill. QUANTITATIVE ANALYSIS
The total number of excitatory and inhibitory nerve terminais was
determined from the serially thin sectioned samples prepared for each of the f i e
animals. The length of an individual nerve terminal was deterrnined by summing the
estimated thicknesses of the sections on which the nerve terminal was present The
total excitatory and inhibitory newe terminal length was calculated by summing the
terminal lengths of every terminal in each of the five animals. The synaptic area was
determined by summing the product of section thickness and synapse length (measured
using calipers pre-set at 2mm) for each of the sections on which the synapse was
present. Only fully sectioned (complete) synapses were used to calculate mean
synaptic area and total synaptic area. If a section was missing for a synapse. the

synaptic length for that section was estimated as the average of the preceding and
following section synaptic lengths.
The nurnber of dense bars per synapse was calculated for complete
synapses; these synapses were then categorized into those containing 0, 1, 2, 3 or
more dense bars to get a better idea of dense bar distribution. The number of dense
bars per synaptic area and per unit terminal volume was also calculated. The length of
dense bars cut in cross-section was deterrnined by summing the thicknesses of the
sections on which the dense bar was present; if a section was missing, then the dense
bar length was deemed incompfete and was omitted from calculations of the total dense
bar length and the mean dense bar length. For dense bars that were cut longitudinally,
length was determined by measuring the extent of the dense bar and multiplying that
value by the magnification factor of the electron micrograph. Dense bar length per
synaptic area and per unit volume was also calculated for each of the five animafs.
Synapses with two or more dense bars were examined for adjacent dense
bars. If adjacent dense bars were obsetved in a single thin section, the distance
between the midpoints of the dense bars was measured and then multiplied by the
magnification factor to obtain a separation distance in Pm. For adjacent dense bars
appearing on different thin sections, the separation distance, c, was determined using
the Pythagorean theorern, a2 + b2 = c2 , where a is the average distance between the
two sections and b is the distance separating the two dense bars had they appeared on
the same section. The number of dense bars that were separated from an adjacent
dense bar by less than or equal to 0.2 Pm was noted.
Two-dimensional reconstructions were produced for some excitatory and
inhibitory synapses to illustrate the number and spacing of dense bars as a visual
indicator of synaptic complexity. In these reconstructions a series of straight parallet

Iines were used to represent a synapse, with each individual Iine representing the
synaptic length on one s e d l section. Dense bars were denoted on these No-
dimensional reconstructions by filled circles in the appropnate locations; dense bars
spanning more than one section had their filled circles connected with a fine.
Statistical tests were used to compare synapse size, dense bar length and
other parameters among means of the five animals. Specifically, a one-way ANOVA
was perfomed to compare inter-animal differences in synaptic area, dense bar length
and dense bar number per synapse. Significance was tested using an alpha level of 5
percent (p c.05). Significant differences in any of these parameters were further
explored using the student's t-test.
IV. VOLUMETRIC ANALYSIS
The percent composition of clear vesicles, dense core vesicles,
mitochondria and axoplasm was deterrnined with the aid of a dot gnd acetate sheet.
The sheet was superimposed on representative terminals found in each of the five
animals. Specifically, the total number of dots landing on each of the terminal
constituents was divided by the total nurnber of dots landing on that particular terminal
to yield a percentage composition for each of the constituents (Le. clear vesicles, dense
core vesicles, mitochondria). This was done for terminals in every fifth section of a
serially sectioned sample.
The mean cross-sectional area of regenerate terminals was calculated by
multiplying the average number of dots per terminal (detemined by adding up the total
number of dots contained within the terminal of interest for al1 sections analyzed, and
dividing this total by the number of sections analyzed) by the squared distance between
the dots on the gridsheet. The volume of the excitatory and inhibitov terminals was

determined by simply multiplying the mean cross-sectional area of each terminal by the
overall length of the terminal (calculated by adding up the individual thicknesses of the
sections in which the terminals were found). Total volume for the regenerate terminals
was obtained by summing the individual terminal volumes calculated using the above
method.

RESULTS
The present experiments on the regeneration of neuromuscular synapses
on the SFM following allotransplantation of a phasic nerve were initiated by Krause et al.
(1 998) who showed that the transplanted newe functionally innewated the muscle
fibers, since stimulation of the regenerated nerve produced EPSPs in eight out of ten
transplant preparations. The muscle fibers in six of seven preparations were innervated
by two or three excitatory axons, many of which generated large (up to 25 mV) EPSPs,
strongly suggesting that the regenerated newe terminals formed phasic-like synaptic
contacts with the tonic SFM (Krause et al., 1 998). Interestingly, electrophysiological
evidence for inhibitory innervation, in the fonn of IPSPs, was not obtained.
My aim was to examine these regenerated neuromuscular terminals in order
to characterize the structural features of their synapses and active zones. Hence,
samples from five different anirnals on which electrophysiological studies had been
performed were examined with thin serial section electron microscopy. The findings are
presented separately for excitatory and inhibitory innervation below.
1. EXCITATORY INNERVATION
A Nerve terminal8
Tissue blocks containing groups of muscle fibers were suweyed in 2-5 pm
increments for the presence of nerve terminal regions. In these survey sections, a useful
indicator of innervation was the presence of muscle granular sarcoplasrn that often
heralded nerve terminals and was usually located around the periphery of the muscle
fiber (Fig. 1). Some of these innervation sites were cut in serials of 1 W-300 sections for
quantitative analysis. A total of between 12-22 excitatory nerve terminals

Figure 1. A. Selected site of innervation in the crayfish SFM by allotransplanted
regenerating axons showing several profiles of excitatory (E) terminals that are densely
populated by spherical clear vesicles. These terminals make synaptic contacts (arrows)
with muscle granular sarcoplasm (s), and at some synapses a presynaptic dense bar is
seen.
B. Another innervation site showing several excitatory (E) terminals in contact with
muscle granular sarcoplasm (s) and containing spherical clear synaptic vesicles. This
innervation site also shows profiles of excitatory axons (EA) recognized by spherical
clear vesides and of an inhibitory axon (IA) recognized by elliptical clear vesicles. The
inhibitory axon gives rise to an inhibitory (1) terminal.
Scale bars: 1 Pm, Magnification: A x24 400; B x17 700.


were identified for each of the five animals. Examination of both the survey and serial
sections revealed that the areas of innewation usually consisted of three to five distinct
excitatory nerve tenninals on any given section (Fig. 1). The terminals were easily
recognized by populations of small, clear synaptic vesicles, a few larger dense core
vesicles, and mitochondrîa. The teminals also displayed characteristic synaptic contacts
with the muscle membrane in the fom of parallel, densely stained pre- and post-
synaptic membranes (Fig. 2). At many of these synapses, distinct active zones for
neurotransmitter release were seen in the fom of presynaptic dense bodies surrounded
by a cluster of clear vesicles.
The majority of the newe terrninals at an innervation site were excitatory
judging from the spherîcal nature of lheir synaptic vesicles (Atwood and Wojtowicz,
1986) (Figs. 1, 2). The few inhibitory nerve terminals identified among the five animals
had elliptically-shaped clear synaptic vesicles indicating inhibitory innervation; these
terminals are described later in this thesis.
The regenerate excitatory nerve terminals, in general, were fairly thin (Fig.
l ) , with mean cross-sectional areas as low as 0.26 I 0.28 pm2 (animal W 7 ) (Table 1).
These terminals were characterized by relatively more clear vesicles than dense
vesicles and mitochondria (Figs. 1, 2). To substantiate these qualitative observations, a
volumetric analysis was perforrned to detemine the percentage of newe terminal
volume that was occupied by the various compositional elements (Le. clear vesicles,
dense core vesicles, and mitochondria). Such an analysis revealed consistently low
percentage values for dense core vesicles and mitochondria relative to the clear
vesicles (Table 1 ). Specifically, clear vesicles occupied between 1 5.7% to 35.9% of
excitatory terminal volume in four of the five animals; a single animal showed an
unusually low volume of 6.1 %. However, the mean across al1 animals was

Figure 2. A. A regenerate excitatory (E) nerve terminal populated with mainly spherical
clear (c) synaptic vesicles, a few dense core (d) vesicles and rnitochondria (m). The
terminal shows a distinct synaptic contact (between bars) made with the muscle
membrane in the region of granular sarcoplasm (s). The synapse possesses a
presynaptic dense bar (arrowhead).
B. Profiles of two excitatory (E) nerve tenninals recognized by the spherical clear
synaptic vesicles and rnaking synaptic contact (between bars) with muscle membrane
(s). The synapse of the upper terminal has a presynaptic dense bar (arrowhead) next to
which is an omega-shaped synaptic profile depicting vesicle exocytosis. The capturing
of such an event confirrns that the regenerated synaptic connections are viable.
Scale bars: 0.5 Pm. Magnification: A x41 900; B x66 300.

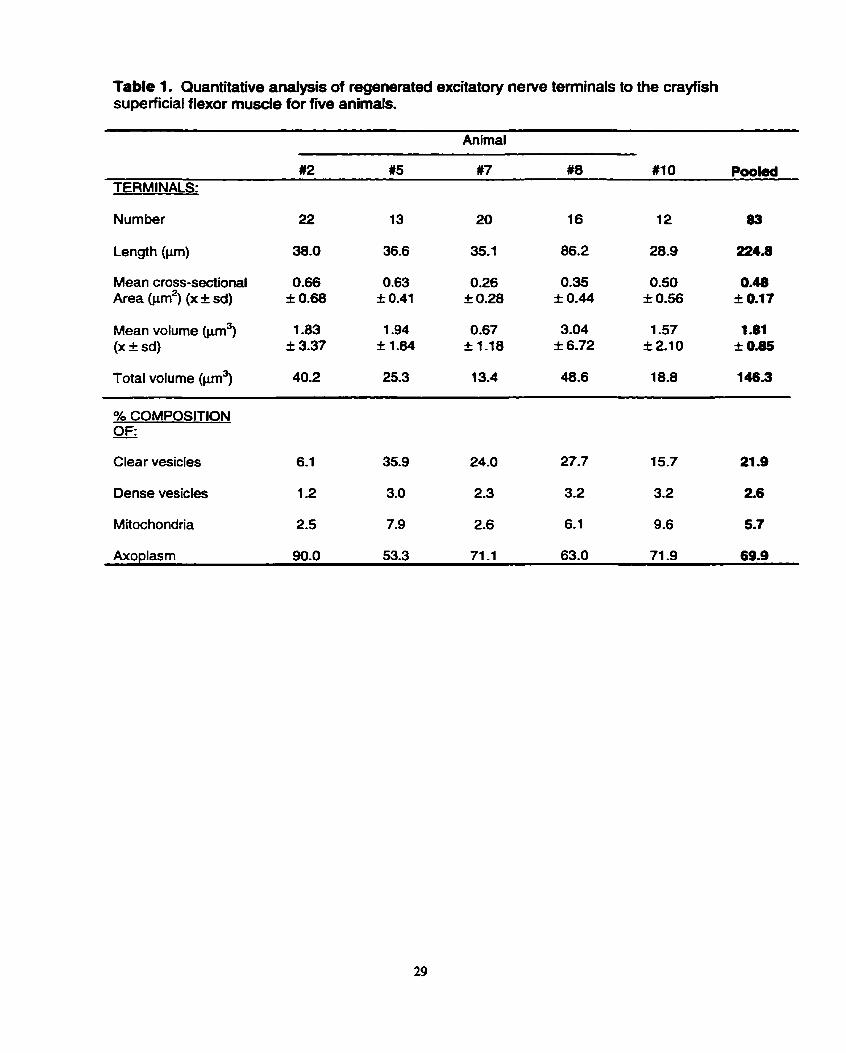
Table 1. Quantitative analysis of regenerated excitatory nerve tenninals to the crayfish superficial flexor muscle for fwe animals.
Animal
#2 t 5 #7 #8 #IO PooM TERMINALS:
Number 22 13 20 16 12 83
Mean cross-sectionai 0.66 0.63 0.26 0.35 0.50 0.48 Area @m2) (x f sd) f 0.68 f 0.41 I 0.28 + 0.44 f 0.56 k 0.17
Mean volume (p) 1.83 1.94 0.67 3.04 1 -57 1.81 (X f sd) f 3.37 S 1.84 I l -18 i 6.72 I 2.1 0 + 0.85
Total volume @m3) 40.2 25.3 13.4 48.6 18.8 146.3
% COMPOSITION OF: - Clear vesicles 6.1 35.9 24.0 27.7 15.7 21.9
Dense vesicles 1 -2 3.0 2.3 3.2 3.2 2.6
Mitochondria 2.5 7.9 2.6 6.1 9.6 5.7
Axoplasm 90.0 53.3 71 -1 63.0 71 -9 69.9

approximately one-fiih of terminal volume (21.9%) for clear vesicles. Dense core
vesicles, on the other hand, were much less prevalent; they occupied between 1.2%
and 3.2% of terminal volume in the five animals. Mitochondria also occupied markedly
l e s terminal volume than the cfear vesicles in each of the five animals; the range
across the five animals studied was 2.5% to 9.6% of terminal volume, with an overall
mean of 5.7% (Table 1). In most terminals across al1 five animals, the mitochondria
appeared simple and unbranched. Furthemore, some of the nerve terminals in each
of the five animals lacked mitochondria-
B. Synapses
Neuromuscular synapses, characterized by the close apposition of
presynaptic (newe terminal) and postsynaptic (muscle fiber) membranes, represent
the sites of information exchange between nerve and muscle. The space between
these two membranes is referred to as the synaptic cleft, which is filled with electron
dense material and is typically 15 nm wide (Fig. 2). Once terminal regions were
located, fully sectioned regenerated synapses were identified, and synaptic areas were
calculated (Table 2). Between 37 and 88 excitatory synapses were identified in each
of the five animals. This study also examined the number of synapses per prn of
terminal, revealing similar values among the five animals and a mean (across the five
anirnals) of 1.48 synapses per Pm of terminal. A comparison of the number of
synapses per unit terminal volume, however, revealed great variation among the five
animals. The range was from 1.34 to 5.22 with a mean of 2.52 synapses per pm3 of
newe terminal (Table 2).
Mean synaptic areas for excitatory synapses, which varied from 0.241 * 0.178 pmZ (animal #7) to 0.527 t 0.485 Pm2 (animal #5), were statistically analyzed to

Table 2. Quantitative analysis of regenerated excitatory synapses ta the crayfish superficial flexor muscle for f i e animais.
Animal
#2 R5 Ut? #8 #1 O Poolad SYNAPSES:
Number 54 57 70 88 37 306
Num ber/pm of terminal 1 -64 1-53 1.75 1.12 1.38 1.48
~urnber /~ rn~ of terminal 1 3 4 2.25 5.22 1.81 1.97 2.52
Mean area (pm2 ) 0.377 0.527 0.241 0.404 0.415 0.409 (X k sd) I 0.277 f 0,485 f 0.1 78 f 0.302 i 0246 I 0.111
Total area @rn2) 20.4 30.0 16.8 42.6 15.4 125.2
A r e a m of terminal (pm2) 0.54 0.82 0.48 0.49 0.53 0.57
~ r e a / ~ ~ m = of terminal m2) 0.51 1.19 1 -25 0.88 0.82 0.93

determine if the means were similar. The analysis revealed each of the animals had a
mean synaptic area that significantly differed from at least one other animal; the mean
synaptic area for animal #7 was significantly lower than each of the other four animals.
Overall, 3û6 regenerate excitatory synapses were measured to yield a mean synaptic
area of 0.409 I 0.1 11 pm2. There was considerable variance around this mean; in
fact, individual synaptic areas ranged from as low as 0.017 pm2 (animal #7) to as high
as 2.472 pm2 (animal W).
Due to the great variation in synaptic areas, we looked at the synaptic
area per pm of nenre terminal for each of the five animals. The values ranged from
0.48 to 0.82, with a mean synaptic area per prn of terminal of 0.57 (Table 2). Since
these regenerated terminais are quite thin, synaptic area was then normalized to
terminal volume. The resulting synaptic area per pm3 of terminal volume ranged from
0.51 to 1.25 with a mean of 0.93 for the five animals (Table 2). It is clear that, in
relation to terminal volume, synaptic area is fairly high.
C. Dense bars
Dense bars are sites for the release of neurotransmitter from nerve
terminals and were obsewed in many of the synapses in al1 five animals (Figs. 2, 3).
Clusters of clear synaptic vesicles were often obsewed surrounding dense bars; we
have captured an omega shaped profile directly adjacent to a dense bar in a thin
section from animal #5 (Fig. 28) which is indicative of clear vesicle exocytosis (Pearce
et al., 1986). Many studies have emphasized the importance of dense bar properties,
especially dense bar number, length, and spacing between dense bars, in detemining
the initial release of neurotransmitter from a nerve terminal (Atwood and Wojtowicz,
1986; Atwood and Cooper, 1996). These parameters were examined in the present

Figure 3. A, B. Regenerate excitatory (E) terminais with spherical clear vesicles,
making synaptic contact (between bars) with muscle membrane in the region of
granular sarcoplasm (s). These synapses display several closely-spaced presynaptic
dense bars (arrowheads).
Scale bar: 0.25 Pm. Magnification: x83 300.


study through a quantitative analysis of dense bars idenüfied in al1 fully sectioned
synapses in the fnre animals (Table 3). Each of the f i e animals housed between 79
and 187 dense bars in total (across al1 synapses), which depicts a fairly wide variance
arnong animals possibly due to differences in the size and extent of the nerve teminal
region examined.
The mean dense bar length for al1 fully sectioned dense bars in the
regenerated excitatory nerve terminals was very close to 0.1 pm (Table 3) and found
not to Vary significantly across the five animals. This aflowed for an examination of the
nurnber of dense bars per synapse as a comparative measure for the f i e animals.
The mean across the f i e animals analyzed was 2.23 I 0.36 dense bars per synapse,
with animal #7 possessing a significantly lower mean (1 -74 I 1.52 dense bars per
synapse) than the other four animals (range of 2.1 3 I 1.57 to 2.70 t 1.80).
To compensate for differences in individual synaptic areas, the number
and length of dense bars per pm2 of synaptic area were also calculated (Table 3). For
the number of dense bats per pm2 of synaptic area, the range across the five animals
was from 4.4 (animal #8) to 7.3 (animal #7) with a mean of 5.7 across the five animals.
Dense bar length per unit synaptic area exhibited a narrower range, and varied from
0.44 prn (animal #8) to 0.67 Pm (animals #2, 5) with a mean of 0.56 Fm for the five
anirnals studied.
To take nerve terminal volume into account, the number of dense bats per
prn3 of terminal was also examined for each animal; the values ranged from 3.3
(animal #2) to 9.1 dense bars per unit volume (animal #7) (Table 3). Similariy, dense
bar length per pm3 of terminal was calculated to assess the amount of dense bar
material present per unit volume. The values ranged from 0.34 pm (animal #2) to 0.83
pm (animal #7), mirroring the pattern observed for number of dense bars per unit
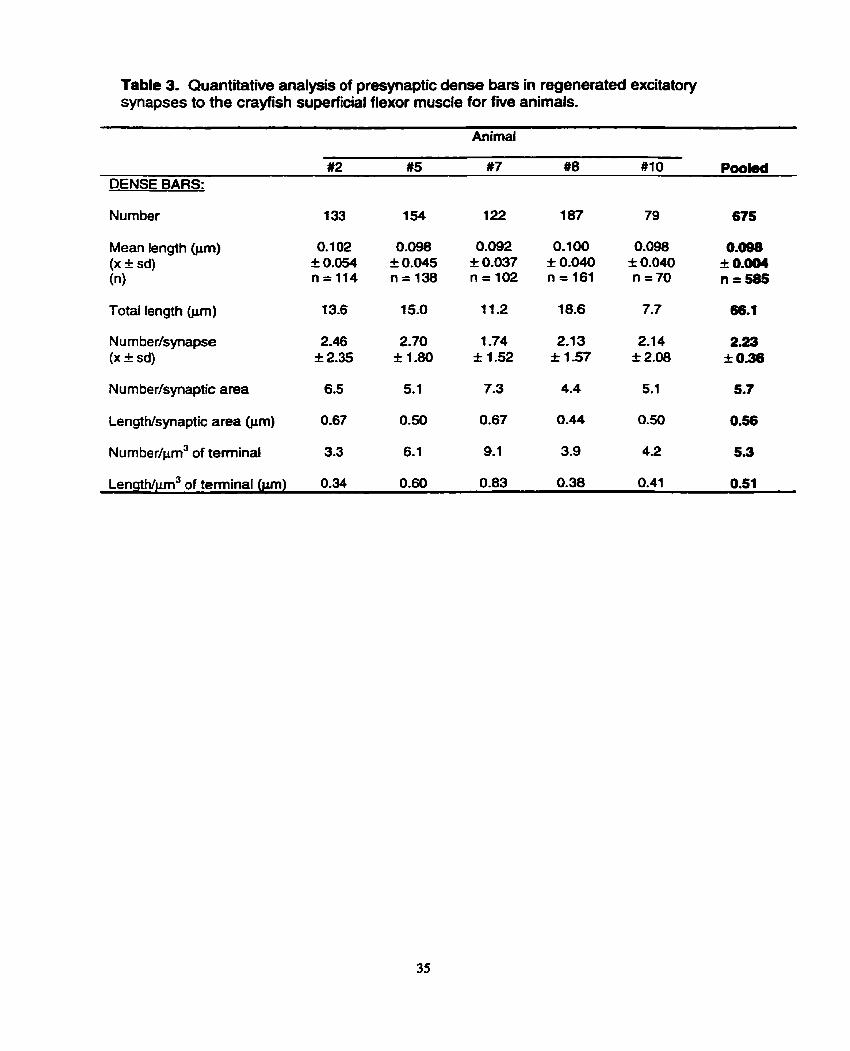
Table 3. Quantitative analysis of presynaptic dense bars in regenerated excitatory synapses to the crayfish superficial flexor muscle for five animals.
Animal
#2 #5 #7 #8 #1 O P w k d DENSE BARS:
Number 1 33 1 54 1 22 187 79 675
Mean length hm) (X f sd) (n)
Total fength (pm) 13.6 15.0 11.2 18.6 7.7 66.1
Number/synaptic area 6.5 5.1 7.3 4.4 5.1 5.7
LengtNsynaptic area (pm) 0.67 0.50 0.67 0.44 0.50 0.56
~umbedprn~ of terminal 3.3 6.1 9.1 3.9 4.2 5.3
~ength/pm~ of terminal (jm) 0.34 0.60 0.83 0.38 0.41 0.51

volume. Interestingly. the number of dense bars per unit volume and the length of
dense bars per unit volume were significantly higher for animal #7. These findings can
be understood by realizing that the number of dense bars per synapse for this animal
was approximately 20% lower than the overall rnean, whereas the mean terminal
cross-sectional area was approximately 40% smaller than the average terminal.
Furthermore, since dense bar length is statistically similar across the five animals,
animal #7 also exhibits a significantly higher dense bar length per unit terminal volume
than the other four animals (Table 3).
A closer examination of the number of dense bars per synapse showed
wide variance (Table 4). ranging from synapses lacking a dense bar to a synapse in
animal #2 that housed 12 distinct dense ban over its 1.200 pm2 synaptic area.
Synapses with multiple dense bars convey a sense of complexity that is more readily
appreciated by viewing them in two-dimensional reconstruction. Selected examples of
such synapses seen in Figure 4 highlight this complexity and show that most possess
dense bars of variable length, although a few had primarily small dense bars. Figure 3
displays examples of synapses with four distinct dense bars on a single thin section.
Synapses with multiple dense bars are more potent releasers of neurotransmitter
molecules because they have more sites for vesicle docking and fusion, more calcium
channels, and a greater Iikelihood that calcium macrodomains will overlap (Cooper et
al., 1996). We therefore categorized synapses based on the number of dense bars
they possess (Table 4) and found that only a small percentage (between 5.3% and
22.9% with a mean of 14.2%) lacked a dense bar. Thus, the overwhelming majority of
synapses possessed active zones. Among these, a substantial number had a single
dense bar (between 24.3% and 44.3% with a mean of 30.4% of al1 synapses), and a
few synapses possessed two dense bars (between 15.7% and 21.6% with a mean of

Figure 4. Two-dimensional vie- of selected examples of regenerated excitatory
synapses showing the number and distribution of presynaptic dense bars. Each
synapse is defined by a series of horizontal lines representing thin sections. Each
dense bar is depicted as a filled circle situated on a horizontal line. When dense bars
traverse more than one thin section, the circles representing a single dense bar are
shown joined to each other and appear as a vertical bar.
Scale bars: 0.5 Pm.


Table 4. Quantitative analysis of dense bars in regenerated excitatory synapses to the crayfish superficial flexor muscle for fnre animals.
Animal
#2 #5 #7 #8 #l O Poolsd O/O SYNAPSES W ITH:
O dense bars 16.7 5.3 22.9 4.5 21.6 14.2
1 dense bar 25.9 26.3 31 -4 44.3 24.3 30.4
2 dense bars 20.4 15.8 15.7 17.0 21 -6 18.1
3 dense bars or more 37.0 526 30.0 34.1 32.4 37.2
SIMPLE / COMPLEX SYNAPSES:
% simple synapses 42.6 31.6 54.3 48.8 45.9 44.6 (0-1 dense bar)
% complex synapses 57.4 68.4 45.7 51 -2 54.1 55.4 (2 or more dense bars)
PAIRED DENSE BARS:
O h of paired dense bars 39.8 39.6 37.7 14.4 29.1 32.1 (separated by I 0.2 pm)
% of synapses with 40.7 38.6 25.7 t 4.8 21 -6 28.3 paired dense bars

18.1 %). There was a higher percentage of synapses with three or more dense bars
(37.2%) than synapses with zero, one or two dense bars. Therefore, a large
proportion of synapses possessed multiple dense bars and this can be appreciated if
the synapses are categorized as simple and complex; simple synapses are those with
0-1 dense bar while complex ones are those with 2 or more dense bars (Cooper et al.,
1995b). As Table 4 reveals, there are more complex synapses than simple ones in
regenerated excitatory temiinals in four of the five animals, with animal #7 being the
exception. There was a range from 45.7% complex synapses for animal #7 to a high
of 68.4% of these synapses for animal #S.
Figure 4 also illustrates that dense bars can be veiy closely spaced and
that such close spacing may facilitate transmitter release through the overlapping of
adjacent calcium macrodomains. Previous studies have demonstrated that 0.2 Pm is
the maximum distance for dense bar interaction to occur (Cooper et al., 1996).
Therefore, adjacent dense bars separated by l e s than or equal to 0.2 pm can be
considered an interacting pair. The dense bar spacing analysis revealed that the
percentage of al1 dense bars capable of interacting with an adjacent dense bar varied
from 14.4% (animal #8) to 39.8% (animal #2), with a rnean of 32.1% (Table 4).
Wojtowicz et al. (1994) found that only complex synapses were active at low
frequencies of stimulation; synapses with closely spaced pairs of dense bars are likely
recruited first at such frequencies. Thus, the percentage of synapses with at least one
closely spaced pair of dense bars was calculated and found to range from 14.8%
(animal #8) to 40.7% (animal #2), with a mean of 28.3% (Table 4). Thus, more than
one-fourth of al1 synapses identified in the current study have a closely spaced pair of
dense bars that likely interact to enhance transmitter release.

Il. INHIBKORY INNERVATION
A Nerve terminal*
Regenerated inhibitory nerve teminals were identified on the basis of the
shape of their clear synaptic vesicles; research performed by Atwood and Morin
(1 970), and Atwood et al. (1 972) found that inhibitory clear vesicles appear elliptical
upon aldehyde and osmium fixation, whereas excitatory clear vesicles appear
spherical (Figs. 1A, 5A). In my study, inhibitory nerve tenninals were not as prevalent
as their excitatory counterparts. Each area of innervation had between zero and two
inhibitory newe terminal profiles on any given section, whereas the same sections had
between three and six excitatory nerve terminal profiles. Furthemore, the five anirnals
studied yielded a total of 21 inhibitory terminals, but yielded 83 excitatory terminals, an
approximately 4-fold difference. ln hibitory newe terminals were found primarily at the
periphery of muscle fibers, and were often located adjacent to excitatory newe
terminals (Fig. 1 A, 5A).
There was a wide variance in mean cross-sectional area of regenerated
inhibitory newe terminals from 0.15 I 0.08 pm2 (animal #8) to 1 .O6 I 0.35 prn2 (animal
#2) (Table 5). The regenerate inhibitory nerve terminals, similar to their excitatory
counterparts, appeared to have more of their volume occupied by clear vesicles than
dense vesicles or mitochondria. The volumetric composition of these terminals was
analyzed and revealed that mi le 15.6% to 37.5% of inhibitory terminal volume in four
of the five animals was occupied by clear vesicles (mean of 19.9%), only 1 .O% to 4.3%
was occupied by dense vesicles (Table 5). The volume of terminal occupied by
mitochondria exhibited a much wider range (from 0% to 11.1% arnong the five
animals), but the mean was a relatively low 6.0% of terminal volume. The

Figure 5. A. Regenerate excitatory (E) and inhibitory (1) nerve terminals can be
distinguished on the basis of the shape of their clear vesicles (c); vesicles contained
within excitatory terminals are spherical in appearance, whereas inhibitory vesicles are
elliptical, or irregularly shaped.
B. A regenerate inhibitory terminal defined by elliptical or irregularly shaped clear
synaptic vesicles (c) shows a synaptic contact (between bars) made with the muscle
membrane in the region of granular sarcoplasm (s)- A presynaptic dense bar
(arrowhead) is seen at this synapse. d = dense core vesicle.
Scale bars: A 0.5 Pm. Magnification: A x38 500; B x54 200.
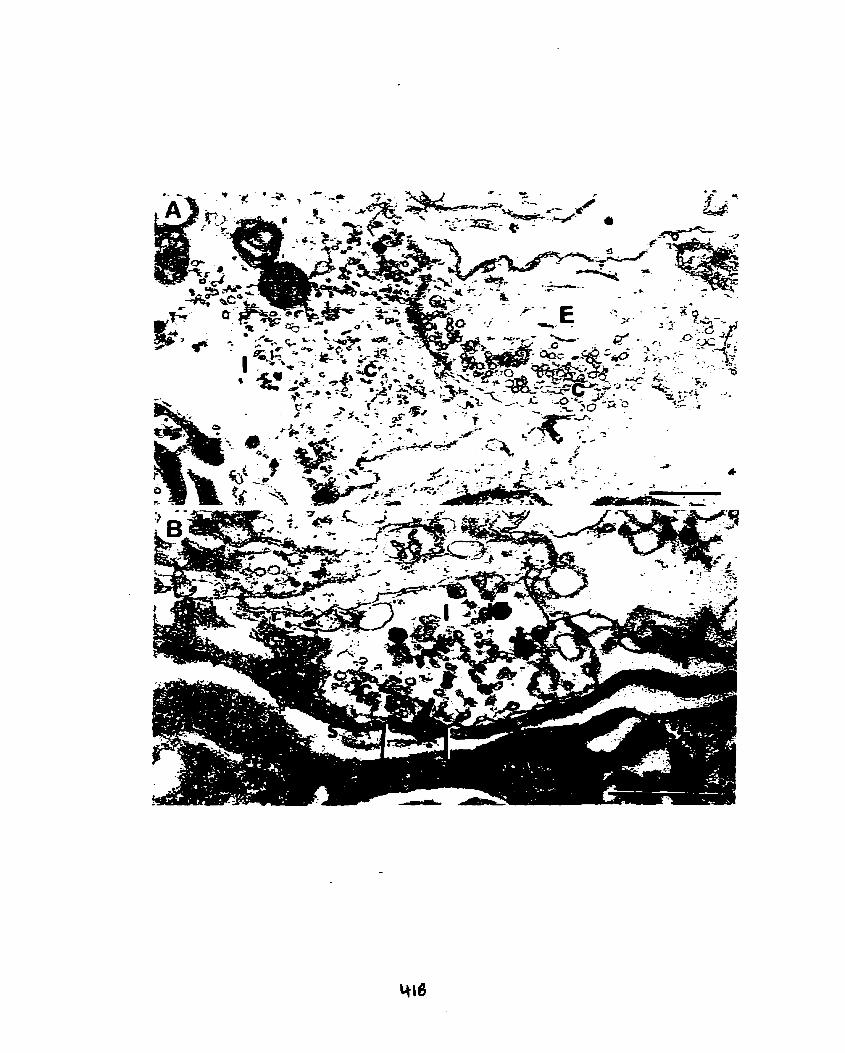
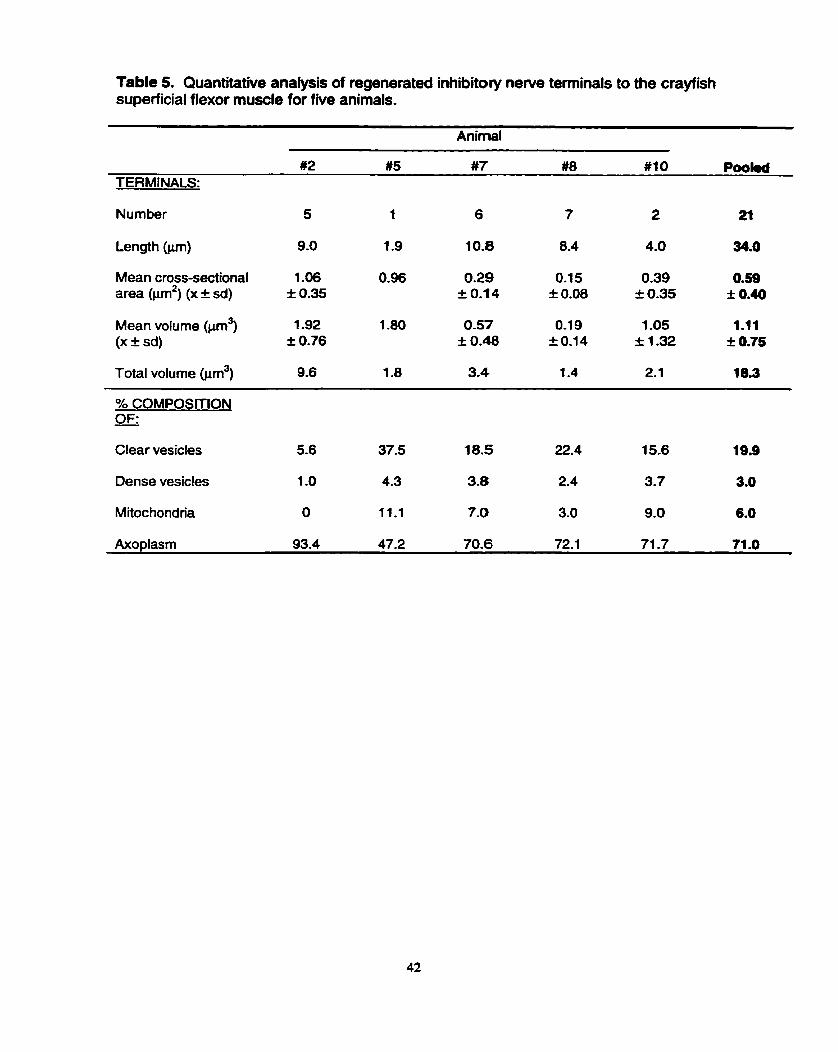
Table 5. Quantitative analysis of regenerated inhibitory nerve tenninals to the crayfish superficial flexor muscle for five animals.
m #5 #7 #8 #IO Pookd TERMINALS:
Number 5 1 6 7 2 21
Mean cross-sectional 1.06 0.96 0.29 0.15 0.39 0.59 area @un2) (x * sd) f 0.35 i: 0.1 4 f 0.08 i 0.35 ; 0.40
Mean volume m3) 1.92 1 .a0 0.57 0.1 9 1 .O5 1.11 (X f sd) f 0.76 f 0.48 f 0.14 1 1 .32 i 0.75
Total volume 9.6 1.8 3.4 1.4 2.1 18.3
% COMPOSITION OF: - Clear vesicles 5.6 37.5 18.5 22.4 15.6 19.9
Dense vesicles 1 .O 4.3 3.8 2.4 3.7 3.0
Mitochondna O 11.1 7.0 3.0 9.0 6.0
Axo~lasm 93.4 47.2 70.6 72.1 71 -7 71 .O

mitochondria, found in most, but not all, of the nerve terrninals, were simple and
unbranched in most cases.
B. Synapses
Regenerated inhibitory nerve teminals also made synaptic contacts with
muscle fibers; their synapses, however, appeared to have a narrower synaptic cleft
and be less densely stained than excitatory synapses (Fig. SB). Between 2 and 17
inhibitory synapses were identïfÏed for each animal to yield a total of 43 synapses
(Table 6). The mean synaptic areas, which varied from 0.21 1 I 0.146 pm2 (animal #8)
to 0.668 i 0.360 pm2 (animal #5), were found to be statistically different only when
comparing animal #5 to animal #8 (Table 6). The variance is exemplified by the fact
that individual synaptic areas ranged from as low as 0.059 pm2 (animal #IO) to as high
as 2.290 pm2 (animal #2). The synaptic area per terminal length and volume was also
calculated and showed wide variation among the five animals.
C. Dense bars
Dense bars with vesicles clustered around them were frequently identified
in inhibitory synapses (Fig. 56). Dense bar quantification yielded between 6 and 34
dense bars in each of the five animals (Table 7). An analysis of dense bar length was
then performed; the mean dense bar length was 0.097 I 0.015 pm, but there was a
signifiant difference between animal #2 and #IO. Specifically, animal #IO had the
lowest mean dense bar length (0.075 I 0.014 pm), whereas animal #2 had the highest
mean among the five animals (0.1 12 0.040 ym). Since the number and size of
inhibitory teminals varied widely between animals, the number of dense bars per
synapse was calcufated for each synapse in each of the five animals. The mean

Table 6. Quantitative analysis of regenerated inhibitory synapses to the crayfish superficial flexor muscle for f i e animals.
Animal
#2 #5 #7 #8 #IO Pwkd SYNAPSES:
Number 12 2 17 7 5 43
Numberfpm of terminal 1.34 1 .O7 1 -71 1 .O1 1.24 1 .n
~ u r n b e r / v ~ of terminal 1 -25 1.11 5.00 5.00 2.38 2.95
Mean area (lun2 ) 0.573 0.660 0.290 0.21 1 0.547 0.414 (X k sd) 10.639 i 0.360 * 0.259 k 0.146 i 0.626 f: 0.470
Total area ()un2 ) 6.9 1.3 4.9 1.5 2.7 17.3
Aredpm of terminal @m2) 0.75 0.72 0.48 0.1 8 0.53 0.53
~ r e d ~ r n ~ of terminal @m2) 0.72 0.74 1.44 1 .O6 1 -30 1 .O5

Table 7. Quantitative analysis of presynaptic dense bars in regenerated inhibitory synapses to the crayfish superficial flexor muscle for fwe animals.
Animal
#2 #5 #7 #8 #1 O Poolad DENSE BARS:
Number 34 6 17 10 10 TI
Mean length (pm) (X f sd) (n)
Total length (pm) 3.8 0-6 1.5 1.1 0.8 7.8
Numberlsynapse (X i sd)
Numbedsynaptic area 4.9 4.6 3.5 6.7 3.7 4.7
LengtNsynaptic area (pm) 0.55 0.46 0.31 0.73 0.30 0.47
~ u m b e r / ~ r n ~ of terminal 3.5 3.3 5.0 7.1 4.8 4.7
~ e n g t h / ~ m ~ of terminal (pm) 0.40 0.34 0.44 0.76 0.36 0.46

number of dense bars per synapse for each of the five animals was statistically similar,
and ranged from 1.00 I 1.58 (animal #7) to 3.00 * 1.41 (animal #5), with an overall
mean of 2.05 0.86 dense bars per synapse across al1 animals.
To standardize for the wide variation in synaptic area, both the number
and length of dense bars per pm3 of newe terminal were calculated (Table 7). The
range for the number per unit volume was from 3.3 (animal #5) to 7.1 (animal #8).
Although animal #5 had the highest mean number of dense bars pet synapse, it had
the lowest number of dense bars per pm3 of terminal. Dense bar length per unit
volume ranged from 0.34 pm (animal #5) to 0.76 pm (animal #8) with a mean of 0.46
pm and followed an inter-animal trend similar to that of the nurnber of dense bars per
unit terminal volume.
The inhibitory innervation was examined more closely in terrns of the
number of dense bars per synapse (Table 8). It is clear frorn Table 8 that there was
tremendous variability, presumably because of a fairly small sample size, in several of
the animals (e.g. 6 dense bars in animal #5 and 10 each in animals #8 and #IO). The
cumulative value from al1 five anirnals shows that approximately one-fifth of the
synapses lack a dense bar, one-fifth have a single dense bar, and almost one-quarter
have two dense bars. Regenerated synapses possessing three or more dense bars
are the most numerous, comprising more than one-third of al1 synapses. Reclassifying
synapses as simple (with 0-1 dense bar) or cornplex (with 2 or more dense bars)
shows that 39.7% of the synapses are simple and 60.3% are complex. A two-
dimensional reconstruction of some selected complex synapses provides a visual
insight into their complexity (Fig. 6).
Close spacing (I 0.2 pm) between dense bars may facilitate transmitter
release via overlapping of adjacent calcium macrodomains. An analysis of such paired
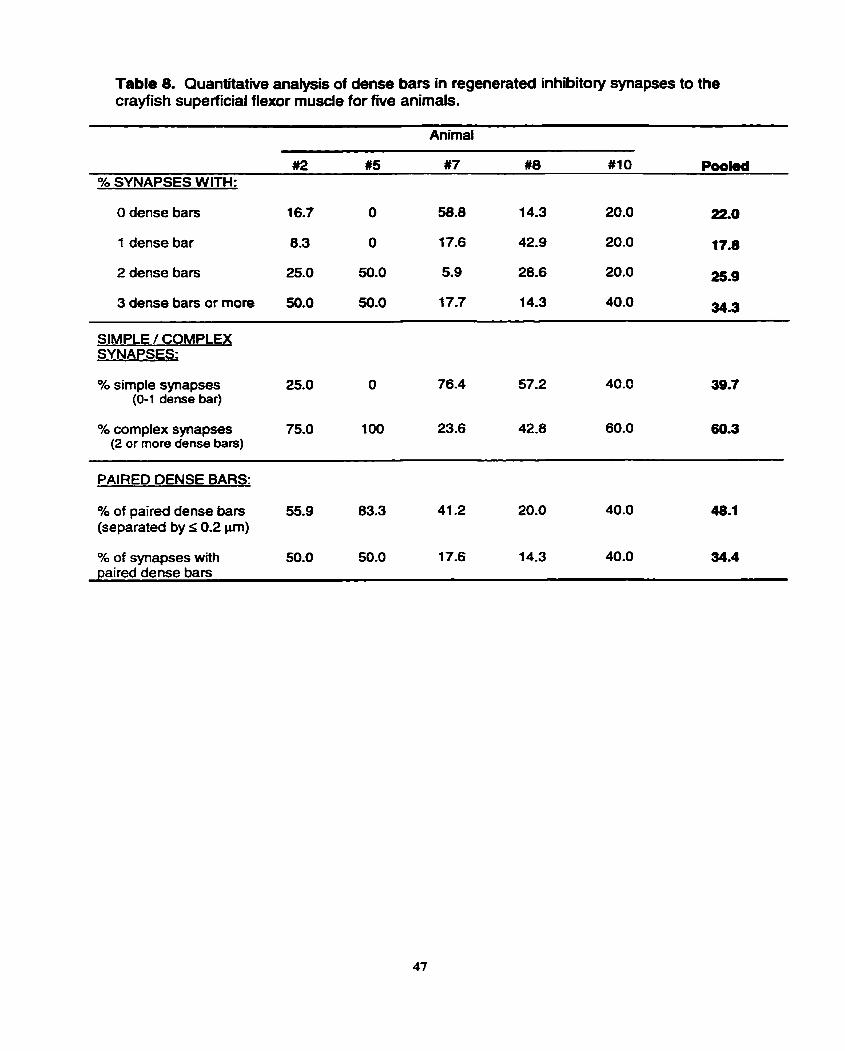
Table 8. Quantitative analysis of dense bars in regenerated inhibitory synapses to the crayfish superficial flexor muscle for fwe animals.
Animal
m #5 #7 #8 #1 O Poolsd O h SYNAPSES WiTH:
O dense bars 16.7 O 58.8 14.3 20.0 22.0
1 dense bar 8.3 O 17.6 42.9 20.0 17.8
2 dense bars 25.0 50.0 5.9 28.6 20.0 25.9
3 dense bars or more 50.0 50-0 17.7 1 4-3 40.0 34.3
SIMPLE / COMPLU( SYNAPSES:
% simple synapses 25.0 O 76.4 57.2 40.0 39.7 (0-1 dense bar)
% complex synapses 75.0 100 23.6 42.8 60.0 60.3 (2 or more dense ban)
PAIRED DENSE BARS:
O h of paired dense bars 55.9 83.3 41.2 20.0 40.0 48.1 (separated by I 0.2 prn)
O h of synapses with 50.0 50.0 17.6 14.3 40.0 34.4 paired dense bars

Figure 6. Two-dimensional views of selected examples of regenerated inhibitory
synapses showing the number and distribution of presynaptic dense bars. Each
synapse is defined by a series of horizontal lines representing thin sections. Each
dense bar is depicted as a filled circle situated on a horizontal Iine. When dense bars
traverse more than one thin section, the circles representing a single dense bar are
shown joined to each other and appear as a vertical bar.
Scale bars: 0.5 Pm.


dense bars was undertaken for the inhibitory synapses (Table 8). The percentage of
the paired intaracting dense bars varied widely from 20.0% in animal #8 to 83.3% in
animal #5 with a mean of 48.1% for the five animais. When the interacting paired
dense bars were considerd in the context of synapses, my analysis revealed that a
substantial percentage of synapses possessed interacting pairs; the range was from
14.3% to 50.0%. with a mean of 34.4%.
D. Presynaptic inhibition
Contacts between adjacent excitatory and inhibitory axons were identified
in two of the five anirnals studied (Fig. 7). The intemembrane space between the
excitatory and inhibitory axons did not display a pronounced density; this finding is in
agreement with previous characterizations of axo-axonal contacts (Atwood and Morin,
1970). At these axo-axonal contacts, the dense bar, with elliptical clear vesicles
clustered around it, was situated on the inhibitory axon membrane. Hence, the
synaptic contact is from the inhibitory axon to the excitatory axon (Fig. 7). Such a
contact is representative of presynaptic inhibition (Atwood and Wojtowicz, 1986).

Figure 7. A, B. Two consecutive serial micrographs showing an excitatory (E) newe
terminal (with spherical ckar vesicles) abutting an inhibitory (1) nerve terminal (with
elliptical clear vesicles). A putative synaptic contact frorn the inhibitory to the excitatory
terminal is indicated by the dense bar (arrow) situated on the inhibitory side, although
synaptic membranes are not clearfy defined. Such a contact would suggest
presynaptic inhibition.
Scale bars: 0.25 Pm. Magnification: x56 300.


DISCUSSION
The present study of regenerated nerve terminals from an allotransplanted
phasic newe on the tonic SFM in crayfish was undertaken with the aim of
characterizing the fine structure of the newe terminals, as well as their synapses and
active zones. Electrophysiology of these regenerated newe terminals was performed
by Krause et al. (1998), who showed that the transplanted nerve functionally
innewated the muscle fibers, since stimulation of the regenerated newe produced
EPSPs. Their study also showed that the two to three excitatory axons innervating
individual muscle fibers gave rise to some very large (25 mV) EPSPs reminiscent of
phasic motor axons in the deep abdominal flexor muscles in crayfish (Kennedy and
Takeda, 1965a). This view was supported with the finding that, at 1 Hz stimulation, the
mean size cf the regenerated EPSP was 8.01 * 7.08 mV (n=102) when the
allotransplanted nerve was the phasic branch, compared to a mean EPSP of 2.3 I
1.41 mV (n=103) when the allotransplanted nerve was the tonic branch (Krause et al.,
1998). The native tonic innervation of the SFM displayed a mean EPSP size of 2.2 I
0.18 mV (Krause and Velez, 1995). Therefore, Krause et al. (1998) suggested that
phasic type synapses regenerated, at least in ternis of the initial release of transmitter,
when a phasic nerve was allotransplanted to the tonic SFM muscle. In view of these
physiological findings, the present ultrastructural analysis of the regenerated
innervation was undertaken primarily for carroboration.
There is a large body of evidence describing features of phasic and tonic
motor nerve terminals in several different crustaceans, notably crabs, crayfish and
lobster (Atwood and Wojtowicz, 1986; Atwood and Cooper, 1996). My study involves
the tonic superficial flexor muscle system, which is innervated by only tonic

rnotoneurons in the native condition, and only phasic motoneurons in the current
experimental condition. Since there are no electron microscopic data on the phasic
flexor nerve innervating the deep flexor muscle, I used the extensor muscle of the
crayfish limb, which is innervated by both a phasic and a tonic excitatory motoneuron,
as a basis for comparison with my results. Collectively, studies in the extensor muscle
point to at least three structural features that distinguish phasic from tonic tenninals:
nerve terminal rnorphology, mitochondrial volume, and the number of dense bars per
synapse. Each of these variables is examined below and compared using other
studies to detemine if the regenerated terminals have a phasic phenotype.
A. Newe terminal morphology
In crayfish limb muscles that receive both phasic and tonic motoneurons
such as the closer (Lnenicka et al., 1986, 1991) and extensor (King et al,, 1996;
Bradacs et al., 1997) it has been well established that the phasic newe terminals are
thin and filiform in appearance and the tonic terminafs as more stout and varicose. As
a result the tonic axon had a volume four times that of the phasic axon for an
equivalent length of terminal in the crayfish extensor muscle (King et al., 1996). In the
present study between 12 and 22 regenerate terminals were examined for each of the
five animals, and the serial sectioning revealed that they were non-varicose and fairly
thin with a mean cross-sectional area of approximately 0.48 t 0.17 ~ r n ~ . In contrast, in
an ongoing electron rnicroscopic study of regenerated newe terminals to the SFM
arising from a tonic allotransplanted nerve, the terminals are distinctly varicose and
fairly thick with a mean cross-sectional area of 2.67 + 2.28 pm2 (see appendix). On the
basis of these measurements, the tonic regenerate terminals have an almost five-fold
greater cross-sectional area and volume than the phasic regenerate terminals. These

differences in terminal morphology in regenerated phasic and tonic teminals parallel
those of previous studies invokring phasic and tonic terminals and strongly suggest
that the regenerated teminals in the cunent study are phasic.
B. Mitochondrial volume
The most striking difference between phasic and tonic terminals is in the
volume of mitochondria with a 3-5 fold diierence between the two in favor of the tonic.
For instance, King et al. (1 996), using serial section electron microscopy, documented
an approximately three-fold greater mitochondrial volume in the tonic teminals (1 6%)
compared to their phasic (5%) counterparts in the crayfish limb extensor muscle.
Bradacs et al. (1997) found a similar difference in mitochondria using light and
confocal microscopy in the same muscle; the tonic terminals showed 16%
mitochondrial volume and the phasic ones 5%. In the crayfish claw doser muscle,
Lnenicka et al. (1 986, 1991 ) recorded an approximately four-fold higher mitochondrial
volume in tonic terminals compared to the phasic ones. The comparatively high
rnitochondrial content presumably allows tonic motoneurons to stay active for
prolonged periods of time, and the low mitochondrial content of the phasic
motoneurons limits them to fire in brief bursts (Kennedy and Takeda, 1965a.b).
Furthemiore, maintaining an adequate supply of transmitter and sustaining the
process of vesicle recycling requires energy (Atwood, 1976); tonic terminals appear to
be more adapted to meet these energy demands because of the relatively high
prevalence of mitochondria,

In the current study, we examined mitochondrial content, and discovered
that only 5.7% of the regenerated newe terminal volume was attributed to
mitochondrial volume. This value is surprisingly similar to that recorded for the phasic
terminals in the Iimb extensor musde (see above), suggesting that the regenerate
terminals were phasic in nature. In fact, many of these regenerate terminals did not
have any mitochondria over the length that they were sectioned and analyzed.
Furthemore, the mitochondria contained within regenerated terminals were simple and
unbranched, which is in accordance with observations made by Lnenicka et al. (1986)
in phasic terminals to the closer muscle of the crayfish claw. It is interesting to note
that physiological conditioning of the phasic axon in the closer muscle of juvenile
crayfish will alter the phenotype towards a tonic condition including increasing
mitochondrial volume but not to the level of the native tonic terrninal (Lnenicka et al.,
1986). It therefore appears that differences in mitochondrial volume between phasic
and tonic axons represent intrïnsic differences which, via extrinsic influences, may be
altered within Iimits but are unlikely to cross over.
Bearing this in mind, we may anticipate that regenerating nerve terminais
on the SFM from an allotransplanted tonic newe would show a much higher
mitochondrial volume than the phasic regenerate terminals. Preliminary data show this
to be the case as regenerate tonic terminals display 19.2% mitochondrial volume (see
appendix). Thus, the mitochondrial volume of the regenerate terminals in my study
clearfy identifies them as phasic in nature.
C. Dense bars per synapse
Another difference that emerged between phasic and tonic nerve terminals
in the crayfish Iimb extensor muscle examined with serial section electron microscopy

was that phasic synapses possessed nearfy two dense bars per synapse white tonic
ones possessed one (King et al., 1996). In a subsequent study for a small number of
physiologically identified nerve terminals, the number of dense bars per synapse was
found to be similar for phasic and tonic terminals at approximately one per synapse
(Msghina et al., 1998). In the most recent study of such physiologically identified
terminals, the number and length of dense bars were computed for nerve terminal
volume to compensate for variation in synapse size (Msghina et al., 1999). The
number per unit volume of newe terminal was 2.13 for phasic terminals and 0.76 for
tonic terrninals; similady, the dense bar length per unit volume of terminal was 0.26 prn
for the phasic and 0.08 pm for the tonic terrninals. Both parameters display an
approximately three-fold higher amount of active zone materiai per unit volume for
phasic terminals than for their tonic counterparts. Dense bar data from King et al.
(1 996), also involving the extensor muscle, revealed a similar three-fold difference in
the above mentioned parameters.
In my study of regenerating nerve terminals from an allotransplanted
phasic nenre on the SFM, the average number of dense bars per synapse was 2.2 and
this was for a reasonably consistent synapse size among the five animals. However, to
compensate for variability in synapse size, I also calculated the dense bar data based
on terminal volume. The mean number of dense bars per unit terminal volume for
phasic regenerate terminals was 5.3, and the mean dense bar length per unit terminal
volume was 0.51 Pm. In an ongoing study in our lab of regenerated synapses from an
allotransplanted tonic newe to the SFM, we found a more than four-fold difference in
favour of the phasic regenerate synapses compared to the tonic ones when
considering both dense bar number (5.3 for phasic vs. 1.3 for tonic) and length (0.51
pm for phasic vs. 0.12 pm for tonic) per unit volume (unpublished observations).

Furthemore, a cornparison between there two types of regenerate synapses for the
number of dense bars per pm2 of synaptic area (5.4 for phasic vs. 2.3 for tonic) or
dense bar length per pm2 of synaptic area (0.53 pm for phasic vs. 0.19 pm for tonic)
reveals a two- to three-fold difference in favour of phasic regenerate connections.
Clearly, the dense bar characteristics of the regenerated terminals in the current study
help identify these terminals as phasic.
II. STRUCTURAL - FUNCTIONAL CORRELAllONS
My electron microscopie examination of regenerated connections yielded
quantitative data on various structural parameters including terminals, synapses and
their active zones which are Iikely responsible for the initial large release of transmitter
upon low frequency stimulation. In this section, I will attempt to correlate structural
features to synaptic output, because of a large body of evidence in support of such a
correlation. The findings of researchers regarding ultrastructural differences has given
rise to the ustructure-function hypothesis", which States that the probability of a
quantum of transmitter being released (binomial parameter p) is correlated to the
arnount of dense bar matenal present (Walrond and Reese, 1985; Atwood and
Lnenicka, 1 986). The structure-function hypothesis has gained support from many
types of studies. For example, studies of differential transmitter release arising from a
single tonic neuron to the crayfish leg opener muscle have revealed high-output
synapses as containing more dense bar material as compared with their low-output
counterparts (Govind et al., 1994; Cooper et al., 1995b). Further support of the
hypothesis comes from studies involving the vertebrate newous system. At the frog
neuromuscular junctions, the total length of the dense bars correlates with the number
of quanta released by a single nerve impulse (Propst and Ko, 1987). Furthemore, at

lizard neuromuscular junctions, the amount of transrnitter released correlates with the
number of intramembranous particles located at the dense bar (Walrond and Reese,
1 985).
This study examines many of the structural parameters of regenerated
synapses and their active zones to determine their degree of correlation to the large
initial EPSP.
A Synapses
a) area
Since synapses represent the sites of contact between nerve and muscle,
it is intuitive to expect higher output connections to have greater synaptic areas than
their lower output counterparts. However, studies by King et al. (1 996) and Msghina et
al. (1998), comparing the phasic and tonic motoneurons innervating the extensor
muscle of the crayfish leg, found that phasic synapses (higher output), were
significantly smaller than tonic synapses (lower output). This finding is interesting,
since the phasic synapses release 100 to 1000 times more transmitter than their tonic
counterparts (Msghina et al., 1998,1999). Hence, synapse size appears to be
inversely correlated with transmitter output since small phasic synapses release more
transmitter than large tonic synapses, at least for low frequencies of stimulation. My
study found that the regenerated synapses exhibited a mean size of 0.409 t 0.1 11
p2, which is much smaller than the mean sire of 1 .O3 I 1.24 pm2 for tonic regenerate
synapses (see appendix). Thus, the inverse relationship between synapse size and
transmitter output may be present for regenerated synapses as well as for intact ones.

b) number
Previously, it was believed that a greater number of synapses were
required to achieve greater quantal output. This idea stems partially from the fact that
the quantal content at indiiidual phasic synapses is not very high; the closer and
extensor muscles of the crab Pachygapsus yielded quantal values between 0.5 and 3
per phasic synapse upon 1 Hz stimulation (Hubbard et al., 1969). Since phasic EPSPs
are relatively large, the density of innervation and the total number of synapses
activated at low frequencies must be fairly high (Atwood, 1976). Specifically, it was
found that at low frequencies of stimulation, synapses with two or more dense bars
(complex synapses) were activated and subsequentfy released transmitter to yield an
EPSP (Zucker, 1 973; Schikorski and Stevens, 1 997; Msghina et al., 1 998). This idea
has been supported by optical studies using the fluorescent dye FMI-43 to examine
active synapses (Msghina et al., 1995; Quigley et al., 1996).
Furthemore, although Cooper et al. (1995b) found a four-fold greater
quantal output at proximal synapses to the crayfish leg opener muscle when compared
to their central counterparts, the difference was attributed to a higher probability of
release, not to a larger number of proximal synapses. Since numerous studies found
that the number of synapses per pm of terminal was fower in the higher output
systems than in the lower output ones (King et al., 1996; Bradacs et al., 1997; Msghina
et al., 1998). it was concluded that synapse number is not a factor when accounting for
the initial EPSP disparity.
My study also found that the number of synapses per pm of regenerated
terminal was low (comparable to high output connections studied previously).
However, King et al. (1996) found an almost two-fold difference in favour of phasic
connections when comparing the number of synapses per pm3 of terminal, suggesting

that high-output connections make more synaptic contacts per unit volume. The
results of the current study revealed many more (approximately four-fold difference)
synapses per pm3 of terminal in the phasic regenerate connections when compared to
the tonic regenerate ones (see appendix). Thus, even though the quantal output per
individual synapse may be low, the high number of synapses per unit terminal volume
(indicative of high density innervation) coupled with the relatively high percentage of
complex synapses could partially explain the high initial EPSPs recorded. In other
words, both a larger number of synapses and a greater probability of release per
synapse may have a physiological impact.
B. Dense bars
a) length
Much research has focussed on the importance of dense bars to the
process of transmitter release. Dense bar length is a parameter often used to explain
differential transmitter release in high- and low-output connections. Since calcium
channel density has been shown to be constant at both lobster (Walrond et al., 1993)
and crayfish (Govind et al., 1994) synapses, a longer dense bar would be indicative of
a larger number of calcium channels, and would facilitate a greater influx of calcium for
a single impulse (Cooper et al., 1996). Given equal probability of channel opening
during an impulse, the ensuing overlap of adjacent calcium microdomains will
exponentially increase transmitter release (Augustine and Charlton, 1986) and
contribute to EPSP amplitude at low stimulation frequencies.
Studies on proximal / central connections in the leg opener muscle
(Govind et al., 1994), as well as phasic and tonic synapses to the leg extensor muscle
(King et al., 1996) in crayfish have revealed that connections producing higher initial

EPSPs have longer mean dense bar lengths. In the current study, however, the mean
dense bar length was fairly close to 0.1 Pm, a value which is almost identical to that
found in other low-output, or tonic, systems (Cooper et al., 199513; King et al., 1996;
Coulthard, 1998). However, a few very long dense bars (i.e. seven that were 0.25 pm
or longer in length and traversed four serial sections) were identified in three of the f i e
anirnals; dense bars of this length are not found in connections yielding mean initial
EPSPs smaller than the mean regenerate initial EPSPs recorded for this study.
Nonetheless, Our findings do not appear to support the idea that longer dense bars are
required ta enhance initial EPSP amplitude. Similarly, Cooper et al. (1995b) did not
find differences in dense bar lengths between tenninals on proximal (high-output) and
central (low-output) muscle fibers. Msghina et al. (1998), studying the extensor
muscle in the crayfish leg, also found that tonic and phasic dense bar lengths did not
differ significantly.
b) nurnber
Unlike dense bar length data, findings related to the number of dense bars
per synapse found in high- and low-output connections in the crayfish opener muscle
(Govind et al., 1 994; Cooper et al., 1 995b), as weH as in phasic and tonic connections
in the crayfish extensor muscle (King et al., 1996; Msghina et al., 1999) are vety
consistent. These studies have invariably shown that a higher nurnber of dense bars
per synapse is correlated with greater quantal output.
High- and low-output synapses, as mentioned earlier, differ in their
synaptic strength. Studies have shown that central varicosities in the crayfish leg
opener muscle c m have the largest number of synapses while concurrently generating
the lowest quantal output following a single impulse- Further examination uncovers a

relatively low percentage of cornplex synapses (2 or more dense bars) and a higher
percentage of synapses wittiout any dense bars when compared to the higher output,
proximal varicosities (Cooper et al., 1995b). In fact, almost half of the proximal
synapses are of the wmplex variety, whereas the ovennrhelming majority of central
synapses are tenned simple, possessing only a single dense bar, or no dense bar at
al1 (Govind et al., 1994). Similady, my study found that more than half of the
regenerated synapses identified were complex. These findings further support the
importance of complex synapses in enhancing quantal release, resulting in a higher
EPSP recorded in the muscle fibers at low frequencies of stimulation.
Previous studies looked at the percentage of completely sectioned
synapses possessing 3 or more dense bars in both high- and low-output synapses
(Cooper et al., 1995b; King et al., t 996). Cooper et al. (1995b) found a large disparity
in proximal versus central synapses containing 3 or more dense bars (28% and 0.6%.
respectively). Similariy, King et al. (1996) found that 17.3% of phasic synapses,
compared to 2.1% of their tonic counterparts, possessed 3 or more dense bars.
These studies point to the relative complexity of the synapses as a contributor to the
amplitude of initial EPSPs. Larger EPSPs are correlated with a higher percentage of
complex synapses, especially those with 3 or more dense bars. The current study
identified fairly high levels of complex synapses containing 3 or more dense bars
(37.2%), which would suggest that the regenerated synapses were in fact capable of
releasing relatively large amounts of transmitter.
Complex synapses have a higher probability of transmission and likely
admit more calcium per impulse, thereby allowing the nerve ending with more complex
synapses to release more transmitter at low frequencies of simulation (Cooper et al.,
1996). This structural hypothesis has been explored in Orosophila by examining both

the structure and function of the RP3 neuron, which innervates the ventral abdominal
longitudinal muscle 6. In one study, a hypomorphic mutant which depresses
expression of Fasll reduces the number of varicosities at the neutomuscular junction
was examined (Stewart et al., 1996). Synaptic transmission was maintained at a
normal level by what appears to be a compensatory mechanism; individual synapses
were larger and had more dense bars per synapse in the mutant, thereby enhancing
quanta! output. A study conducted by Renger et al. (1997) found that the rutabaga
(rut') mutation, which suppresses the level of cyclic AMP in the nervous systern,
exhibits lower than normal synaptic transmission (Zhong and Wu, 1991), possibly by
hindering the vesicle docking process (Atwood et al., 1997). Again, individual
synapses of this mutant are larger than normal and possess more dense bars. It
appears as if the synaptic structure adapts to synaptic transmission deficiencies by
producing larger synapses of a more complex varisty.
Zucker (1991) illustrated that a single pulse elicits a greatet calcium influx
for synapses housing long, or multiple dense bars, as opposed to small, simple
synapses. Cooper et al. (1995b) further explored the involvement of calcium in
differential transmitter release in high- and low-output connections and found greater
calcium buildup in high-output terminals at low frequencies of stimulation- Higher
calcium entry would be expected to result in a greater mean quanta1 content (Cooper
et al., 1995b). Recent work on rnammalian central synapses has provided evidence
for differences in calcium entry as being responsible for differing probabilities of
transmitter release at individual boutons (Manilow et al., 1994).
Further support of calcium's involvement comes from long terrn adaptation
studies, which show that sustained increases in electrical activity of a phasic terminal
alter its physiology and morphology to those of tonic terminals (Atwood et al., 1985;

Lnenicka et al., 1986, 1991). Calcium influx in the soma is necessary for long tem
adaptation to occur in phasic motoneurons in crayfish (Hong and Lnenicka, 1993).
Nguyen and Atwood (1990) showed that an increase in electrïcal activity caused a
decrease in calcium cunents found in the soma; this change was dependent on protein
synthesis. In other words, certain key proteins rnay be up- or dom-regulated to
suppress calcium cunents upon enhanced electrical activity (Hong and Lnenicka,
1995). A similar result rnay occur at nerve terrninals. Therefore, it seems plausible
that native tonic and phasic neurons have differences in calcium currents, which rnay
partially account for the differences in quantal output. These studies support the idea
that complex synapses, with their relatively high calcium influx upon low frequency
stimulation, are capable of releasing large amounts of transmitter. Thus, a relatively
high proportion of complex synapses, as observed in the current study, would
presumably produce a greater quantal output than systems with a lower percentage of
complex synapses.
c) spacing
A number of mechanisms rnay be responsible for the enhanced
transmitter release probability obsewed in synapses containing multiple dense bars.
For instance, dense bars rnay serve to attract and concentrate vesicles at the active
zone. Alternatively, calcium channels contained within closely spaced dense bars rnay
foster overlapping "calcium domains" (Chad and Eckert, 1984; Simon and Llinas,
1985) upon stimulation induced calcium influx, resulting in higher transmitter release.
Thus, a synapse's structural complexity can impact transmitter release since the
distance between dense bars affects the intraterminal calcium concentration; widely
spaced dense bars would be less likely to interact, and would thereby invoke a lower

calcium concentration, than closely spaced ones (Wojtowicz et a1.,1991, 1994; Cooper
et al., 1995b).
Calcium microdomains are present immediately adjacent to calcium
channels; the intemal calcium concentration falls off rapidly as the distance from the
channel increases (Chad and Eckert, 1984; Simon and Llinas, 1985; Zucker and
Fogelson, 1986). The lateral spread of transient calcium domains around an individual
calcium channel is approximately 100-200nm (Simon and Llinas, 1 985; Zucker and
Fogelson, 1986). In synapses possessing more than one dense bar, there is a range
of separation distances between adjacent dense bars; this distance can be as low as
50 nm (Cooper et al., 1996). When dense bars are separated by 200 nrn (0.2 pm) or
less, the intracellular calcium concentration is greatest at the centre of each dense bar,
and at the midpoint between them. As a result, the calcium concentration will be
enhanced at the edges of the dense bar (the location of synaptic vesicle docking and
release), thereby increasing the transient probability of transmitter release. The extent
of the spatial enhancement of transmitter release relies on the number of calcium
channels involved, and on the concentration of calcium required to induce vesicular
release (Cooper et a1.,1996). Although the mean paired dense bar separation did not
differ significantly between tonic regenerate connections (see appendix) and
regenerate connections from the cunent study, phasic regenerate connections had
over five-fold more (32.1% vs. 6.0%) of their dense bars capable of interacting with an
adjacent dense bar. Such a disparity presumably helps explain tha relatively high
quanta1 output at the phasic regenerate synapses examined in the current study.
Since maximum intracellular calcium concentration is greater at the
midpoint between dense bars and at the centre of each one, quanta1 release may have
been further enhanced in synapses with three closely spaced dense bars. The

interaction between these three dense bars would presumably result in a greater influx
of calcium at low frequencies of stimulation, allowing for enhanced quantal release.
A study by Cooper et al. (1996) indicates that ~y,:zpses with two or more
closely spaced dense bars (complex synapses) have a greater transmitter release
probability than those with zero or one dense bar (simple synapses). Wojtowicz et al.
(1994) showed that complex synapses might correlate to the quantal parameter n at
low stimulation frequencies, since the value for n is consistent with the number of
closely spaced pairs of dense bars. At higher frequencies, the ensuing buildup of
intraterminal calcium (Delaney et al., 1989) may enhance dense bar interactions
through the spread of free calcium and binding with immobile, and possibly mobile,
calcium buffers (Cooper et a1.,1996). Further enhancement of calcium concentration
would serve to facilitate the interaction of more widely spaced dense bars, giving the
nerve an additional mechanism by which frequency could alter synaptic output (Cooper
et al., 1996). In my study, therefore, the number of synapses with one or more closely
spaced pairs of dense bars may represent the number of synapses that are active at
low stimulation frequencies. Approximately one-fourth (28.3%) of al1 phasic
regenerate synapses from the cuvent study are of this variety, compared to only one-
twelfth (8.5%) of al1 tonic regenerate synapses (see appendix), or approximately a
three-fold difference in favour of the higher-output, phasic regenerate connections.
This finding supports the initial EPSP disparity identified in the physiological studies
involving these allotransplant preparations.
The importance of calcium entry at an adjacent dense bar is dependent on
the concentration of calcium necessary to induce transmitter release. Apparently, a
relatively small increase in intracellular calcium concentration is capable of enhancing
transmitter release, even though that increment in calcium concentration could not

directly evoke much release (Delaney et al., 1989). Therefore, it is likefy that a small
additional amount of calcium entering the terminal through an adjacent dense bar
could enhance the probability of transmitter release at the synapse (Cooper et al.,
1996). Thus, in the cunent study, the higher prevalence of interacting dense bars in
regenerate synapses suggests a higher transmitter release than in control synapses,
which did not have as many interacting dense bars.
In this study, I have demonstrated that the differences in quantal output
reside primarily in the probability of synaptic release, rather than in the number of
synapses or the number of active zones. Factors contributing to a higher probability of
release include a greater number of dense bars per synapse and a higher prevalence
of "cornplex" synapses with closely spaced dense bar pairs. In neuromuscular
systems innervated by a single excitatory tonic motor neuron, such as the crayfish leg
opener muscle, such factors are believed to be sufficient in accounting for quantal
output and EPSP disparities between high- and low-output fibers. In systems
innervated by distinct phasic and tonic excitatory motor neurons, however, the 100 to
1000 fold disparity in quantal content at low stimulation frequencies can not be
accounted for by dense bar properties or synapse complexity. Since quantal content
measurements have not been made in the regenerated synapses of a flexor newe
onto a previously denewated tonic SFM, it is difficult to conclude whether the structural
features are capable of fully accounting for the large EPSPs recorded.
C) Other factors involved
The structural features discussed above may be sufficient in accounting
for the large initial transmitter output. However, it is quite possible that other
differences beyond the scope of the cuvent study exist, and these additional factors

may contribute to the large initial EPSP amplitudes. One possibility is the density of
postsynaptic glutamate receptors; a relatively high density at regenerated synaptic
contacts rnay allow more of the released transmitter to bind postsynaptically, thereby
producing a greater EPSP. Another possibility relates to differences in calcium
buffering, which rnay increase the amount of calcium available in the regenerated
teminals. Given the importance of calcium in synaptic transmission, diierences in
buffenng capacities may have a significant impact on quantal output, and hence on
EPSP amplitude. A further possibility involves calcium extrusion rates. Msghina et al.
(1999) found that the calcium decay rate is 2-3 times higher for tonic terminals when
cornpared to their phasic counterparts; as a result, calcium accumulation is
suppressed. A relatively low calcium extrusion rate in regenerated tenninals would
enhance calcium accumulation, thereby increasing quantal output. Elevated calcium
channel sensitivity to calcium may also exist in regenerated tenninals, resulting in a
disproportionately high release of transmitter (Msghina et al., 1999).
III. INHIBITORY INNERVATION
The major focus of these transplant studies was the excitatory innervation,
which was assessed in the first instance by the presence of EPSPs upon stimulation of
the regenerated nerve (Krause et al., 1998). Such electrophysiological detection of
innervation was not, however, obvious for an inhibitory neuron in that hyperpolarizing
postsynaptic potentials were not recorded. The use of hyperpolarizing potential as
indicators of inhibitory innervation is not conclusive since, depending on the muscle
fibers' equilibrium potential for chloride, inhibitory potentials can be depolarizing
(Atwood, 1976). However, the present structural study revealed clear-cut evidence for
inhibitory innervation in the form of nerve tenninals with elliptical clear synaptic

vesicles. Crustacean neuromuscular synapses fixed with aldehydes show distinctive
differences in the shape of their clear synaptic vesicles (Atwood and Wojtowicz, 1986);
excitatory vesicles are spherical and inhibitory ones are elliptical. The presence of
these inhibitory nerve terminals therefore provides data on synapse regeneration of a
distinctly different type of neuron. Hence, it may be instructive to compare these
regenerated inhibitory terminals with their excitatory counterparts.
First, regenerate inhibitory terminals were found in each of the five
allotransplanted preparations, making them a consistent feature of the regenerated
innervation. Such consistency is noteworthy since the target muscle, given a choice of
six transplanted neurons with up to fwe excitors and a single inhibitor, accepted the
single inhibitor each time. This outcome most likely occurs because the muscle has
receptor sites for an inhibitory neuron in addition to those for excitatory axons.
Norrnally, fibers of the SFM receive a single inhibitory neuron and two to three
excitatory axons (Kennedy and Takeda, 1965b; Velez and Wyman, 1978a).
Second, the regenerate inhibitory terminals were fewer in number than
excitatory ones in each of the five preparations. This finding points to the possibility
that a single inhibitory neuron regenerated to the SFM, which is in keeping with its
previous history and with the fact that the transplanted phasic nerve would have
possessed a single inhibitory neuron. However, this conclusion is tentative and
requires physiological verif ication.
Third, the size of the regenerated inhibitory terminals, their mean
mitochondrial volume and number of dense bars per synapse were comparable to
those of their excitatory counterparts, suggesting that a phasic type of inhibitory axon,
analagous to the phasic excitatory axons, had regenerated. Such an idea is in
accordance with previous findings; transmitter release characteristics are similar

between excitatory and inhibitory axons to a given muscle fiber (Atwood and Bittner,
1971).
Fourth, inhibitory nerve terminals also make presynaptic contact with
excitatory newe terminals. A functional advantage of presynaptic inhibition is the
conservation of transmitter in excitatory nerve terminals (Bryan and Krasne, 1977).
Furthemore. removal of the inhibition produces a strong synaptic response, since
there is a large readily releasable pool of vesicles that can be drawn upon (Atwood and
Walcott, 1965). Atwood and Morin (1970) foond that axo-axonal synapses polarized
from the inhibitory to the excitatory newe terminal are encountered 10-20 times less
frequently than neuromuscular synapses in the crayfish claw opener muscle.
Physiological examination of these connections at low stimulation frequencies,
however, revealed that presynaptic inhibition is appmximately 10 times more effective
than the more prevalent postsyiiaptic inhibition. Thus, individual axo-axonal synapses
must be at least 100 times more effective than neuromuscular synapses (Atwood and
Morin, 1970). The greater potency of am-axanal contacts may result from the fact that
they are contacting small excitatory nerve terminais instead of much larger muscle
fibers. Therefore, the inhibitory transmitter released upon low frequency stimulation
would have a greater impact on the small excitatory newe terminal membrane
potential than that of a large muscle fiber (Atwood and Morin, 1970). The number of
axo-axonal contacts representative of presynaptic inhibition were seen infrequently in
the current study, yet it is important ta note that these types of contacts, in addition to
al1 of the factors discussed previously, are also capable of controlling the quantal
output from excitatory neuromuscular junctions.
Overall, the presence of regenerated inhibitory innervation shows
robustness of the allotransplanted neural tissue, and alço shows that the muscle target

welcomes, or perhaps even demands, innervation by two different types of axons. In
other words, the muscle has the receptors for both the excitatory and inhibitory axons
and, as a resuit, both types regenerate. Quite possibly, the number of receptor sites
may Iimit the density, and hence the nurnber, of axons regenerating to the SFM.

SUMMARY
1. Crayfish have robust regenerative powers as shown by the fact that after
transecting the superficial flexor newe to the SFM in the abdomen, connecthMy
maps were re-established within 8 weeks (Ely and Velez, 1982). Synaptic
regenerative abilities were further explored through studies involving the
allotransplantation of the superficial flexor newe from a donor animal to a
previously denewated host animal (Krause and Velez, 1995). These studies
showed that the transplanted motoneurons re-established synaptic connectivi
patterns similar to the native patterns.
2. A mismatch experiment involving an allotranspfantation of the phasic flexor nerve
to the previously denewated tonic SFM was attempted (Krause et al., 1998) to
determine if the regenerated connections were tonic in nature (indicating an
instructive retrograde influence), or phasic in nature (indicating that the
regenerating motoneuron follows an autonomous program when forming synaptic
connections). The regenerating newe in these mismatch experiments showed
axons with numerous sprouts indicative of phasic axons; tonic axons have fewer
sprouts. Electrop hysiolog ical studies on experimental animals revealed 3-fold
larger initial EPSPs at low frequencies of stimulation in the regenerated
neurornuscular system when compared to the native SFM system. The EPSP
data strongly suggest that the newly formed connections possess phasic
properties, at least at low frequencies of stimulation.
3. My study examined the ultrastructural properties of the regenerated connections in
these mismatch experiments using thin serial section electron microscopy.

Altogether, fwe animals which had been previously examined with
electrophysiology (Krause et al., 1998) were selected for study. Qualitative
features of regenerated synaptic terminals were typical crustacean-like; nerve
terminals were populated with mostly dear vesicles, a few dense core vesicles and
rnitochondria and possessed well defined synapses, many with one or more dense
bars (active zones). Quantitative analysis of these structural parameters allowed
me to correlate them with the large EPSP and also provided cues as to whether
they were of the phasic type.
4. Previous studies have repeatedly pointed to three primary structural features which
distinguish phasic and tonic terminals: terminal morphology, mitochondrial
volume, and the number of dense bars per synapse. Tonic regenerate terminals
were approximately fie-fold larger than the phasic regenerate terminals examined
in the current study. Phasic regenerate terminals also had fewer mitochondria;
only 5.7% of terminal volume was occupied by mitochondria, compared to a
relatively high 19.2% for tonic regenerate terminals. An examination of the
number and length of dense bars per unit synaptic area as well as per unit volume
revealed No- to four-fold differences between phasic and tonic regenerate
connections. The above mentioned features strongly suggest that the regenerated
newe terminals examined in the current study are phasic in nature.
5. To explore structural-functional correlations in the regenerated system, synapses
and dense bars were examined. Although the phasic regenerate connections had
a smaller mean synaptic area, they had more synapses per unit volume compared
to tonic regenerate connections. An examination of dense bar number and

distribution reveafs that many phasic regenerate synapses were of the complex
variety, and many have 3 or more dense bars. Furthemore, phasic regenerate
synapses had over three-fold more of their dense bars capable of interacting with
an adjacent dense bar when compared to tonic regenerate connections. A
cornparison of the number of synapses possessing an interacting pair of dense
bars revealed a similar trend. These findings collectively support the physiological
finding that the regenerated connections are capable of higher initial transmitter
release than the lower output, tonic connections.
6. Although physiokgical evidence for inhibitory innervation in the form of lPSPs was
not obtained, electron microscopic examination of innervation sites revealed the
presence of inhibitory nerve terrninals. Ultrastructural analysis of the inhibitory
connections revealed key similarities with excitatory connections in newe terminal
morphology, mitochondrial volume and dense bar number per synapse. These
findings suggest that synaptic features of inhibitory innervation are similar to that of
excitatory innervation. lnhibitory nerve terminals also make pre-synaptic contact
with excitatory newe terminals via presynaptic dense bars located on the inhibitory
membrane. Such axo-axonal contacts may represent pre-synaptic inhibition,
which may reduce excitatory impulse activity and help conserve transmitter.
7. In sum, the electrophysiological findings of Krause et al. (1998) of large EPSPs in
regenerated systems were supported by the current ultrastructural analysis.
Although not specifically tested in the mismatch condition, it is quite possible that
connection specificity is maintained by the presence of target derived molecular
cues, which guide regenerated connections to previous sites of innervation. These

cues, however, may have fiffle or no impact on the nature of the regenerated
connections. Thus, an allotransplanted rnotoneuron may use its surroundings to
arrive at the appropriate locations within its novel surroundings, but utilires its
autonomous program to fom synaptic connections.

REFERENCES
Arcaro KF, Lnenicka GA (1 995) lntnnsic differences in axonal growth from crayfish fast and slow motoneurons. Dev Bi01 168:272-283.
Atwood HL (1 976) Organization and synaptic physiology of crustacean neuromuscular systems. Prog Neurobro17:291-391.
Atwood HL (1 982) Synapses and neurotransmitters. In T h e Biology of Crustaceam (H.L. Atwood, and D.C. Sandeman, eds.), pp. 105-150. Academic Press, New York.
Atwood HL, Bittner GD (1971) Matching of excitatory and inhibitory inputs to crustacean muscle f ibers. J Neurophysiol34: 1 57-1 70.
Atwood HL, Cooper RL (1 996) Synaptic diversity and differentiation: crustacean neuromuscular junctions. Invert Neurosci 1 :291-307.
Atwood HL, Govind CK (1990) ActMty-dependent and age-dependent recruitment and regulation of synapses in identified crustacean neurons. J Grp Bi01 153:105- 127.
Atwood HL, Kwan 1 (1976) Synaptic development in the crayfish opener muscle. J Neurobiol7:289-312.
Atwood HL, Lnenicka GA (1986) Structure and function in synapses: emerging correlations. TINS 9:248-250.
Atwood HL, Marin L (1 983) Ultrastructure of synapses with different transmitter- releasing characteristics on motor axon terminais of a crab, Hyas areneas. Cell Ess Res 231 :103-115.
Atwood HL, Morin WA (1 970) Neuromuscuhr and axoaxonal synapses of the crayfish opener muscle. J Ultrastmct Res. 32:351-369.
Atwood HL, Nguyen PV (1 995) Neural adaptation in crayfish. Amer Zoo1 35:28-36.
Atwood HL, Walcott B (1 965) Recording of electrical activity and movement from legs of walking crabs. Can J Zoo1 43:657-665.
Atwood HL, Wojtowicz JM (1 986) Short-terni and long-term plasticity and physiological differentiation of crustacean motor synapses- Int Rev Neurobiol28:275-362.
Atwood HL, Lang F, Morin WA (1972) Synaptic vesicles: selective depletion in crayfish excitatory and inhibitory axons. Science 176:1353-1355.
Atwood HL, Lnenicka GA, Marin L (1985) Morphological responses to conditioning stimulation in a phasic motor axon of crayfish (Procarnbams clarkir). J Physiol (Lond) 365:26P.
Atwood HL, Karunanithi S, Georgiou J, Charlton MP (1997) Strength of synaptic transmission at neuromuscular junctions of crustaceans and insects in relation to calcium entry. lnvert Neurosci 3:81-87.
Augustine GJ, Chariton MP (1 986) Calcium dependence of presynaptic calcium current and post-synaptic response at the squid giant synapse. J Physiol (Lond) 381 :6 1 9-64.

Bittner GD (1 968) Oierentiation of newe tenninals in the crayfish opener muscle and its f unctional significance. J Gen Physiol 51 :731-758.
Bradacs H, Cooper RL, Msghina M, Atwood HL (1 997) Differential physiology and morpho10 y of phasic and tonic motor axons in a crayfish limb extensor muscle. J Ekp Bio 7 200:677-691.
Bryan JS, Krasne FB (1 97ï) Presynaptic inhibition: The mechanism of protection from habituation of the crayfish lateral giant fibre response. J Physiol (Lond) 271 ~369-390.
Chad JE, Eckert R (1 984) Calcium domains associated with individual channels can account for anomalous voltage relations of Ca-dependent responses. Biophys J 45:993-999.
Clernent JF, Taylor AK, Velez SJ (1 983) Effect of a limited target area on regeneration of specif ic neuromuscular connections in the crayfiçh. J Neurophys 49:2l6- 226.
Cooper RL, Hampson DR, Atwood HL (1995a) S aptotagmin-like expression in the motor newe terminais of crayfish. Brarir 4" es 703:214-216.
Cooper RL, Marin L, Atwood HL (1 995b) Synaptic differentiation of a single motor neuron: Conjoint definition of transmitter release, presynaptic calcium signds, and ultrastructure. J Neurosci 1 5:4209-4222.
Cooper RL, Stewart BA, Wojtowicz JM, Wang S, Atwood HL (1995~) Quantal measurement and analysis methoûs compared for crayfish and Drosophila neurom uscular junctions, and rat hippocam pus. J Neurosci Methods. 6 1 :66-79.
Cooper RL, Winslow JL, Govind CK, Atwood HL (1 996) Synaptic structural complexity as a factor enhancing probability of calcium-mediated transmitter release. J Neurophysiol75:2451-2466.
Coulthard R (1 998) The roles of rnotoneurons and their muscle targets in synaptogenesis during regeneration of a foreign transplant. MSc. thesis, University of Toronto, pp. 1-82.
Couteaux R, Pecot-Dechavassine M (1 970) Vesicles synaptiques et poches au niveau des "zones activew de la fonction neuromusculaire. Compt Rend Acad Sci Ser D 271 :2346-2349.
Dahm LM, Landmesser LT (1991) The regulation of synaptogenesis dunng normal development and following activity blockade. Dev Bi01 130:621-644.
Delaney KR, Zucker RS, Tank DW (1989) Calcium in motor nerve terminals associated with posttetanic potentiation. J Neurosci 9:3558-3567.
DeRosa RA, Govind CK (1978) Transmitter output increases in an identifiable lobster motoneurone with growth of its muscle fibers. Nature (Lond) 273:676- 678.
Dudel J, Kuffler SW (1 961 ) Presynaptic inhibition at the crayfish neuromuscular junctions. J Physiol (Lond) 1 55543-562.

Ely P, Velez SJ (1982) Regeneration of specific neuromuscular connections in the crayfish. 1. Pattern of connections and synaptic strength. J Neurophysiol 47~656-665.
Evoy WH, Beranek R (1 972) P hannacoiogical localization of excitatory and in hibito s aptic regions in crayfish slow abdominal flexor muscie fibres. Comp en $am 3:li&l86.
z Evoy WH, Kennedy D, Wilson DM (1 967) Discharge patterns of neurones supplying
tonic abdominal flexor muscles in the crayfish- J Exp Bi01 46:393-411.
Fatt P, Katz B (1 953) The effect of inhibitory nerve impulses on a crustacean muscle f iber. J Physiol (Lond) 1 21 :374-388.
Florey E, Cahiil MA (1982) The innewation pattern of crustacean skeletal muscle. Cell Tissue Res 224527-541.
Gilfary HL, Kennedy D (1 969) Pattem generation in a crustacean motoneuron. J Neurophysiol32:595-606.
Goda Y, Sudhof TC (1 997) Calcium regulation of neurotransmitter release: reliably unreliable? Current Opinion Ce11 Bi0 951 3-51 8.
Goelet P, Castellucci VJ, Schacher S, Kandel ER (1986) The long and short of long- term memory - a rnolecular framework. Nature 322:419-422.
Goransson LG, Hunt WP, Velez SJ (1 98) Regeneration studies on a crayfish neuromuscular system II. Effect of changing the nerve entry point into the muscle field on the gradient of innervation. J Neurobiol 1 9:let -1 52.
Govind CK, Walrond JP (1 989) Structural plasticity at crustacean neuromuscular synapses. J Neurobiol20:409-421.
Govind CK, Atwood HL, Lang F (1973) Synaptic differentiation in a regenerating crab- f im b muscle. Proc Nat/ Acad Sci USA 70:822-826.
Govind CK, Pearce J, Wojtowicz JM. Atwood HL (1 994) "Strong" and "Wear synaptic differentiation in the crayfish opener muscle: structural correlates. Synapse 1 6~45-58.
Harrington CC (1 993) An investigation of structural relationships of the crayfish neuromuscular junctions. M.Sc. thesis, University of Toronto, pp. 1-92.
Heuser JE, Reese TS (1 979) S naptic vesicle exocytosis captured by quick f reezing . F In 'The Neurosciences ourth Study Program," F.O. Schmitt and F.G. Worden, Eds., MIT Press, Cambridge, MA, pp. 573-600.
Heuser JE, Reese TS, Dennis MJ, Jan Y, Jan L, Evans L (1979) Synaptic vesicle exocytosis captured by quick f reezing and correlated with quanta! transmitter release. J Ce11 Bi01 81 :275-300.
Hildebrand JG, Townsel JG, Kravitz €A (1 974) Distribution of acetylcholine, choline, choline acetyltransferase and acetylcholinesterase in regions and single identified axons of the lobster nervous system. J Neurmhem 23:951-963.
Hill RH, Govind CK (1 981 ) Comparison of phasic and tonic synaptic terminals in lobster muscle. Ce11 Tissue Res 221 :303-310.

Hirokawa N, Sobue K, Kanda K, Haraba A, Yorifugi H (1989) The cytoskeletal architecture of the presynaptic terminal and molecular structure of synapsin 1. J Ce11 Bi01 lOû:111-126.
Hong SJ, Lnenicka GA (1993) Long-terni changes in the neuromuscular synapses of a crayfish motoneuron produced by calcium influx. Brain Res 605:121-127.
Hong SJ, Lnenicka GA (1 995) ActMty-dependent reduction in voltage-dependent calcium cuvent in a crayfis h rnotoneuron. J Neurosci 1 535394547.
Hoy RR (1 969) Degeneration and regeneration in abdominal flexor motor neurons in the crayfish. J EXp Zoo1 1 72:219-232.
Hubbard JI, Llinas R, Quastel DMJ (1 969) Electrophysiological Analysis of Synaptic Transmission. Edward Arnold: London.
Hunt W P, Velez SJ (1982) Regeneration of specific neuromuscular connections in the crayfish II. Effect of changes in the target area. J Neurophys 47:666-676.
Hunt WP, Velez SJ (1989a) Regeneration of an identifiable motoneuron in the crayfish 1. Pattems of reconnection and synaptic strength established in normal and altered target areas. J Neurobiol20:703-717.
Hunt WP. Velez SJ (1989b) Regeneration of an identifiable motoneuron in the crayfish II. Patterns of reconnection and synaptic strength established in presence of an extra nerve. J Neurobiol20:718-730.
l ravan i J (1 965) Mem branedepolarisation der Muskelfasern des Offnerm uskels des Flusskrebses auf Nervenreiz und Kaliumapplikation. Expenentia 21 :609-610.
Jahromi SS, Atwood HL (1 974) Three-dimensional ultrastructure of the crayfish neuromuscular apparatus. J Ce11 Bi01 63:599-613.
Jessel TM, Kandel ER (1993) Synaptic transmission: a biochemical and self modifiable forrn of cell-cell communication. Ce1172 / Neuron 10 (suppl):l -30.
Katz B (1966) Nerve, Muscle and Synapse. New York: McGraw-Hill.
Kennedy D, Bittner GD (1 974) Ultrastructural correlates of motor axon nerve regeneration in crayfish. CeIl Tissue Res 148:97-110.
Kennedy O, Takeda K (1965a) Reflex control of abdominal flexor muscles in the crayfish. 1. The twitch system. J Exp Bi01 43321 1-227.
Kennedy D, Takeda K (1965b) Reflex control of abdominal flexor muscles in the crayfish. II. The tonic system. J Exp Bi01 43:229-246.
King MJR, Atwood HL, Govind CK (1 996) Structural features of crayfish phasic and tonic neuromuscular terminais. J Comp New 372:618-626.
Krause KM, Velez SJ (1 995) Regeneration of neuromuscular connections in crayfish allotransplanted neurons. J Neurobiol27:154-1 71.
Krause KM, Pearce J, Govind CK (1998) Regeneration of phasic motor axons on a crayfish tonic muscle: neuron specifies synapses. J Neurophysiol80:994-997.
Krause KM, Pearce J, Velez SJ, Govind CK (1996) Structure of allotransplanted

ganglia and regenerated neuromuscular connections in crayfish. J Neurobid 30:439-453.
Kuffler SW, Yoshikami D (1975) The number of transmitter mofecules in a quantum: an estimate from iontophoretic application of acetylcholine at the neuromuscular synapse. J Physiol (Lond) 251 :465-482.
Linder TM (1 974) The accumulative properties of facilitation at crayfish neuromuscular synapses. J Physiol (Lond) 238223,234.
Llinas R (1 977) In Approaches to the Cell Biology of Neurons, W.M. Cowan and J.A. Ferendelli, Eds. (Society for Neuroscience, Behtesda, MD) vo1.2, pp. 139-1 60.
Uinas R, Sugimori M, Silver RB (1992) Microdomains of high calcium concentration in a presynaptic terminal. Science 256:677-679.
Lnenicka GA, Atwood HL (1 985a) Agedependent long-term adaptation of crayfish phasic motor axon synapses to altered activity. J Neurosci 5:459-467.
Lnenicka GA, Atwood HL (1 985b) Long-terni facilitation and long-temi adaptation at synapses of a crayfish p hasic motoneuron. J Neurobiol 1 6:97-110.
Lnenicka GA, Marin L, Atwood HL (1 986) Morphological transformation of synaptic tenninals of a phasic motoneuron by long-term tonic stimulation. J Neurosci 62252-2258-
Lnenicka GA, Hong SJ, Combatti M, LePage S (1 991) Activity-dependent development of synaptic varicosities at crayfish motor teminals. J Neurosci 1 1 :1040-1048.
Magrassi L, Puwes D, Lichtman JW (1987) Fluorescent probes that stain living nerve terminals. J Neurosci 7: 1 207-1 21 4.
Manilow R, Otmakhov N, Blum KL, Lisman $ (1994) Visualking hippocampal synaptic function b optical detection of Ca ' entry through the Nmethyl-0-aspartate channel. 6 rocNatlAcadSciUSA91:8170-8174-
Meiss DE, Govind CK (1 979) Regional differentiation of neuromuscular synapses in a lobster receptor muscle. J Ekp Bio! 79:99-114.
Msghina M, Charlton MP, Atwood HL (1 995) Differentiation of transmitter release properties and calcium transients in phasic and tonic motor nerve endings of crustacea. Soc Neurosci Abstr 21 : 1 38.1 5.
Msghina M, Govind CK, Atwood HL (1 998) Synaptic structure and transmitter release in crustacean phasic and tonic motor neurons. J Neurosci l8:lW4-l382.
Msghina M, Millar AG, Charlton MP, Govind CK, Atwood HL (1 999) Calcium entry related to active zones and differences in transmitter release at phasic acd tonic synapses. Ce11 Mol Neurosci In press.
Nguyen PV, Atwood HL (1990) Expression of long-terni adaptation of synaptic transmission requires a critical period of protein synthesis. J Neurosci 10:lOgg- 1109.
Nguyen PV, Atwood HL (1994) Altered impulse activity modifies s naptic physiology and mitochondria in crayfish phasic motor neurons. J europhysiol72:2944- 2955.
hl'

Nordlander RH, Sin er M (1972) Uectron rnicroscopy of severed motor fibers in the crayfish. Z 1 elttotsch 1 26:157-181.
Otsuka M, lversen LL, Hall ZW, Kra* EA (1966) Release of amma-aminobutyric acid frorn inhibitory nerves in lobster. Proc Nat1 Acad 8 ci USA 56:1110-1115.
Pamas 1, Atwood HL (1 966) Phasic and tonic neuromuscular s stems in the abdominal
723. 6 flexor muscles of the crayfish and rock lobster. Comp iochem Physiol18:701-
Pearce J, Govind CK, Shivers RR (1 986) lntramembranous organization of lobster excitatory neuromuscular synapses. J Neurocytol 1 5:241-252.
Propst JW, Ko CP (1 987) Correlations between active zone ultrastructure and synaptic function studied with freeze-fracture of physiologically identified frog neuromuscular junctions. J Neurasci 7:3654-3664.
Pumplin DW, Reese TS, Llinas R (1981) Are the resynaptic membrane particles the calcium channels? Proc Nat1 Acad Sci U 8 A 78:7210-7213.
Quigley PA, Cooper RL, Govind CK, Atwood HL (1 996) Recruitrnent of active synapses at the cra ish neuromuscular junction visualized with the fluorescent dye FM 1 -43. Soc C eurosci Abstr 22:309.9.
Renger JJ, Atwood HL, Wu CF (1997) Single-bouton rewrding from the larval neurornuscular junction of Drosqphila CAMP cascade mutants. 38m Annual Drosophila Research Conference.
Rheuben MB (1 985) Quantitative comparison of the structural features of slow and fast neuromuscular junctions in Manduca. J Neurosci 31704-1716.
Roberts WM, Jacobs RA, Hudspeth AJ (1990) Colocalization of ion channels involved in frequency selectivity and synaptic transmission at presynaptic active zones of hair cells. J Neurosci 10:3664-3684.
Robitaille R, Adler EM, Charlton MP (1990) Strategic location of calcium channels at transmitter release sites of frog neuromuscular synapses. Neuron 5773-779.
Schikorski T, Stevens CF (1 997) Quantitative ultrastructural analysis of hippocampal excitatory synapses. J Neurosci t7:5858-5867.
Selverston Al, Remler MP (1 972) Neural geometry and activation of crayfish fast flexor motoneurons. J Neurophysiol35:797-814.
Simon SM, Llinas RR (1 985) Compartmentalization of the submembrane calcium activity during calcium influx and its significance in transmitter release. Biophys J 48:485-498.
Smith SJ, Augustine GJ, Charlton MP (1985) Transmission at voltage-clarnped giant s napse of the squid: evidence for cooperativity of presynaptic calcium action. &oc Nat1 Acad Sci USA 82622-625.
Smith SJ, Buchanan J, Osses LR, Chariton MP, Augustine GJ (1993) The spatial distribution of calcium signals in squid presynaptic terminals. J Physiol (Lond) 472 1573-593.
Stewart BA, Atwood HL (1 992) Synaptic plasticity in a regenerated crayfish phasic motoneuron. J Neurobiol23:881-889.

Stewart BA, Schuster CM, Goodman CS, Atwood HL (1996) Homeostasis of synaptic transmission in D m h i l a with genetically altered newe terminal morphology. J Neurosci 1 630774886.
Takeuchi A (1 976) Studies of inhibitory effects of GABA in invertebrate nervous systems. In =GABA in Newous S stem Functionw (E. Roberts, T.N. Chase, and D.B. Tower, eds.), pp. 255-267. A aven, New York.
Takeuchi A, Takeuchi N (1965) Lacalized action of gamma-butyric acid on crayfish muscle. J Physiol (Lond) 1 77:225-238.
Uchizono K (1 967) lnhibitory s apses on the stretch receptor neurone of the crayf is h. Nature (Lon J" ) 207:642-643.
Velez SJ, Wyman RJ (1 978a) Synaptic connecüvity in a crayfish neuromuscular system. 1. Gradient of innervation and synaptic strength. J Neurophysiol 41 :75-84.
Velez SJ, Wyman RJ (1 978b) Synaptic connectivity in a crayfish neuromuscular system. II. Neive-muscle matching and neive branching patterns. J Neurophysiol41:85-96.
Walrond JP, Govind CK, Huestis SE (1 993) Two structural adaptations for regulating transmitter release at lobster neuromuscular synapses. J Neurosci 13:4831- 4845.
Walrond JP, Reese TS (1985) Strucutre of axon terrninals and active zones at synapses on lizard twitch and tonic muscle fibers. J Neurosci !%Il 1 8-1 1 31.
Wiens TJ (1 982) Srnall systems of neurons: Control of rhythmic and reflex activity. In The Biology of Crustacean (H.L. Atwood and D.C. Sandeman, eds.), Vol. 4, pp. 1 93-240. Academic Press, New York.
Wilson DM, Davis WJ (1965) Newe impulse pattems and reflex control in the rnotor system of the crayfish cîaw. J &p Bi01 43:193-210.
Wojtowicz JM, Mann L, Atwood HL (1994) Activity-induced changes in synaptic release sites at the crayfish neuromuscular junction. J Neurosci 14:3688-3703.
Wojtowicz JM, Smith BR, Atwood HL (1 991) Activity-dependent recruitment of silent synapses. Ann NY Acad Sci 627:169-179.
Zhong Y, Wu CF (1 991) Altered synaptic plasticity in Drosophila memory mutants with a defective cyclic AMP cascade. Science 251 : 1 98-201.
Zucker RS (1 973) Changes in the statistics of transmitter release during facititation. J Physiol (Lond) 229:787-810.
Zucker RS, Delaney KR, Mulkey R, Tank DW (1991) Presynaptic calcium in transmitter release and posttetanic potentiation. Ann NY Acad Sci 635:lgl- 207.
Zucker RS, Fogelson AL (1 986) Relationship between transmitter release and presynaptic calcium influx when calcium enters through discrete channels. Proc Nat1 Acad Sci USA 83:3032-3036.

Quantitative analysis of regenerated excitatory newe terminais to the crayfish superficial flexor muscle upon allotransplantation of a tonic superficial flexor nerve. Surgeries perfomed by Kristen Krause, and data collected by Joanne Pearce and Rahim Hirji.
TONIC REGENERATE
TERMINALS:
Mean cross-sectional 2.67 area @m2) (x I sd) I 2.28
Oh COMPOSITION OF:
CIear vesicles 22.1
Dense vesicles 1.7
Axoplasm 57.2
SYNAPSES:
Number
Mean area ( p 2 ) (X + sd) ~urnber/~m= of terminal
DENSE BARS:
Number
Mean length (pm) (X f sd) (n)
Nurnber/synapse (X t sd)
~urnber f~ rn~ of terminal
~ e n ~ t h l p m ~ of terminal (P)
PAIRED DENSE BARS:
% of paired dense bars 6.0 (separated by I 0.2 pm)
% of synapses with 8.5 paired dense bars







