J. Mol. Biol. (1969) 39, 335-349 ies on the Nuclear 4 to 7 s RNA of the Novikoff Hepat YOICHI MORIYAMA, JAMES L. HODNETT, ARCHIE W. PRESTAYKOAND HARRIS BUSCH Department of Pharmacology Baylor Urkuersity College of Medicine Houston, Texas 77025, U.S.A. (Received 14 September 1968) Nuclei isolated from Novikoff ascites hepatoma cells by the citric acid procedure were found to be a useful source of nuclear 4 to 7 s RNA, i.e., approximately Q mg were obtained per 100 g wet weight of cells. The 4 to 7 s RNA was separated into four main peaks by chromatography on Sephadex GIOO columns and 11 or more components by polyacrylamide gel electrophoresis. Two uridylic acid-rich RNA fractions (approximately 5 to 7 s), which are found only in nuclei, were purified by preparative polyacrylamide gel electrophoresis; one was obtained in an electrophoretically homogeneous state and had a base composition of 22% adenine, 26% uracil, 33% guanine and 19% cytosine. The labeling patterns with either [14C]sodium formate or [methyV4C]methionine as a precursor were different for the uridylic acid-rich nuclear RNA than for the 4 s and 5 s RNA. Evidence is presented that the uridylic acid-rich RNA of peak II is not a degradation product of high molecular weight RNA. 1. Introduction Although the presence of low molecular weight RNA in nuclei and nucleoli of mam- malian cells ha,s been known for some time (Muramatsu & Busch, 1965; Okamura & Busch, 1965; Steele, Okamura & Busch, 1965), it is only recently that studies have been made on the numbers and types of RNA present. In studies from this laboratoq (Nakamura, Prestayko & Busch, 1968), nucleoli of Novikoff hepatoma cells were shown to contain transfer RNA, 5 s RNA and an RNA fraction rich in uridylic acid. In ra,t liver nuclei, Peacock & Dingman (1967) demonstrated the presence of low molecular weight RNA species by polyacrylamide gel electrophoresis. In addition, Larsen, Galibert, Lelong & Boiron (1967) reported that nuclei of KR cells contained an $ s RNA that was absent from the cytoplasm. Pene, Knight & Darnell (1968) have reported recently another type of nuclear and nucleolar low molecular weight, RNA that is extracted by the hot phenol-sodium dodecyl sulfate procedure. In an effort to obtain, sufficient RNA to carry out structural studies such as those that have been made on transfer RNA (Holley et al., 1965a,b) and on 5 s -RNA (Rrownlee, Sanger & Barrell, 1968; Forget & Weissman, 1967), it seemed essential to use richer sources than nucleolar 5 to 7 s RNA since the yield of this RNA was very low. In the present study, Novikoff hepatoma ascites cells were used because the yield of nuclear RNB is approximately nine times that of nuclei of liver cells and the yield of nuclear 4 to 7 s RNA was six times that of normal liver cells. Sufficient quantities of 4 to 7 s RNA were obtained by zonal ultracentrifugation to 335
Transcript
J. Mol. Biol. (1969) 39, 335-349
ies on the Nuclear 4 to 7 s RNA of the Novikoff Hepat
YOICHI MORIYAMA, JAMES L. HODNETT,
ARCHIE W. PRESTAYKO AND HARRIS BUSCH
Department of Pharmacology
Baylor Urkuersity College of Medicine Houston, Texas 77025, U.S.A.
(Received 14 September 1968)
Nuclei isolated from Novikoff ascites hepatoma cells by the citric acid procedure were found to be a useful source of nuclear 4 to 7 s RNA, i.e., approximately Q mg were obtained per 100 g wet weight of cells. The 4 to 7 s RNA was separated into four main peaks by chromatography on Sephadex GIOO columns and 11 or more components by polyacrylamide gel electrophoresis. Two uridylic acid-rich RNA fractions (approximately 5 to 7 s), which are found only in nuclei, were purified by preparative polyacrylamide gel electrophoresis; one was obtained in an electrophoretically homogeneous state and had a base composition of 22% adenine, 26% uracil, 33% guanine and 19% cytosine. The labeling patterns with either [14C]sodium formate or [methyV4C]methionine as a precursor were different for the uridylic acid-rich nuclear RNA than for the 4 s and 5 s RNA. Evidence is presented that the uridylic acid-rich RNA of peak II is not a degradation product of high molecular weight RNA.
1. Introduction Although the presence of low molecular weight RNA in nuclei and nucleoli of mam- malian cells ha,s been known for some time (Muramatsu & Busch, 1965; Okamura & Busch, 1965; Steele, Okamura & Busch, 1965), it is only recently that studies have been made on the numbers and types of RNA present. In studies from this laboratoq (Nakamura, Prestayko & Busch, 1968), nucleoli of Novikoff hepatoma cells were shown to contain transfer RNA, 5 s RNA and an RNA fraction rich in uridylic acid. In ra,t liver nuclei, Peacock & Dingman (1967) demonstrated the presence of low molecular weight RNA species by polyacrylamide gel electrophoresis. In addition, Larsen, Galibert, Lelong & Boiron (1967) reported that nuclei of KR cells contained an $ s RNA that was absent from the cytoplasm. Pene, Knight & Darnell (1968) have reported recently another type of nuclear and nucleolar low molecular weight, RNA that is extracted by the hot phenol-sodium dodecyl sulfate procedure.
In an effort to obtain, sufficient RNA to carry out structural studies such as those that have been made on transfer RNA (Holley et al., 1965a,b) and on 5 s -RNA (Rrownlee, Sanger & Barrell, 1968; Forget & Weissman, 1967), it seemed essential to use richer sources than nucleolar 5 to 7 s RNA since the yield of this RNA was very low. In the present study, Novikoff hepatoma ascites cells were used because the yield of nuclear RNB is approximately nine times that of nuclei of liver cells and the yield of nuclear 4 to 7 s RNA was six times that of normal liver cells. Sufficient quantities of 4 to 7 s RNA were obtained by zonal ultracentrifugation to
335
336 Y. MORIYANA E!Z’ dL.
permit fractionation of the RNA by exclusion chromatography and preparative gel electrophoresis ; analyses of nucleot,ide compositions were made on electrophoretically distinct fractions of this RNA. Studies were also made on labeling of the low mole- cular weight nuclear RNA fractions.
2. Materials and Methods (a) Animals and tumor cells
Male albino rats, weighing 200 to 250 g, were obtained from Cheek Jones Company (Houston) and were fed ad l&turn on Purina laboratory chow. The Novikoff ascites hepatoma cells were transplanted 6 to 7 days before the experiments. For studies on methylation, 50~~ of L-[methyZ-14C]methionine (specific activity, 46.0 me/m-mole, Schwartz Bioresearch, Inc.) was injected intraperitoneally into each of 2 rats at 30 min, 3 hr and 24 hr before killing. For 14C-labeling studies of nuclear RNA, 100~~ of [“*Cl- sodium formate (specific activity, 42.8 me/m-mole, Radiochemical Centre, Amersham, England) was injected 1 hr before killing. The tumor-bearing rats were killed by cervical dislocation and tumor cells were collected from the peritoneal fluid by centrifugation.
(b) Isolation of nuclei
The citric acid procedure used for isolation of nuclei from tumor cells was similar to that described previously (Higashi, Adams & Busch, 1966; Higashi, Narayanan, Adams & Busch, 1966). The cells were suspended in cold 5% citric acid (I:9 w/v) and were homogenized in a continuous homogenizer (5 x 10m3-in. pestle clearance) (Busch & Desjardins, 1965). After 10 to 12 passes, the homogenate was centrifuged at 600 g for 10 min. The precipitate was resuspended in 0.25 If-sucrose containing 1.5% citric acid and the suspension was layered over 2 vol. of 0.88 M-sucrose containing 1.5% citric acid; the nuclei were sedimented by centrifugation at 900 g for 10 min. In the small-scale experiments, the tumor cells were homogenized in a tightly fitting Teflon pestle glass homogenizer (2 to 3 x lo-s-in. pestle clearance).
(c) Extraction of RNA
RNA was extracted from nuclear preparation by the dodecyl S04+hot phenol pro- cedure (Muramatsu, Hodnett & Busch, 1966). The nuclear pellet was homogenized for 1 min in 0.3% dodecyl SO4 solution containing 0.14 M-sodium chloride and 0.05 M-sodium acetate buffer pH 5.1 (2 ml. of dodecyl SO4 solution was used for 1 g of original tissue). An equal volume of 90% phenol containing 0.1% S-hydroxy-quinoline was then added, and the mixture was homogenized for another minute. The homogenate was shaken at, 65°C for 10 min and then at room temperature for 15 min. After centrifugation at 5000 g for 30 min the aqueous layer was separated and back-extracted with phenol solution. 2 vol. of ethanol containing 2% potassium acetate was added to the aqueous phase and the solution was kept at -20°C overnight to precipitate the RNA. The RNA was dis- solved in about 5 ml. of water and purified by reprecipitation with ethanol containing 2% potassium acetate.
(d) Isolation of 4 to 7 s RNA
Isolation of 4 to 7 s RNA from total nuclear RNA was carried out by zonal ultra- centrifugation (Higashi, Adams & Busch, 1966). A linear gradient of 5 to 45% sucrose solution containing 0.1 M-sodium chloride, 0.001 M-EDTA, 0.01 M-sodium acetate buffer pH 5.1 (total volume 1200 ml.) was made in the zonal ultracentrifuge rotor (Beckman B-IV). Approximately 100 to 120 mg of nuclear RNA dissolved in 20 ml. of 3% sucrose was added through the core. After centrifugation at 35,000 rev./min for 15 hr at 2 to 4’C. the gradient was pumped out and fractionated with an ISCO automatic fractionator; the absorbancy at 254 rnp was determined (Fig. 1). The 4 to 7 s RNA fractions were pooled and the RNA was precipitated overnight with 2 vol. of ethanol containing 20/, potassium acetate at -20°C. For small-scale analyses, a %-ml. 5 to 40% linear sucrose
t Abbreviation used: dodecyl SO4, sodium dodecyl sulfate.
RNA. (a) Pattern for 120 mg of R.NA from a rate zonal centrifugation in the zonal ultracentrifuge;
the pooled shadowed portion was the 4 to 7 s RNA fraction subjected to later fractionation; (b) analytical sucrose density-gradient centrifugation profile for 0.2 mg of the nuclear RNA used in (a); (c) analytical scale profile of the 4 to 7 s RNA obtained from (a).
density-gradient was made in the same buffer used for ultracentrifugation and centrifuged at 25,000 rev./min for 16 hr in a Spinco SW25 rotor. After centrifugation, the gradients were fractionated with an ISCO automatic fractionator (Steele et al., 1965).
(e) Preparation of 14C-labeled ribosowuzl RNA
Each Novikoff hepatoma-bearing rat was injected intraperitoneally with 100 PC of [l*C]sodium formate (specific activity, 42.8 me/m-mole, obtained from the Radiochemical Centre, Amersham, England) 24 hr before killing. The ribosomes were isolated from tumor cells of 5 rats by the method described by Takanami (1961). RNA was extracted from ribosomes with dodecyl SO,-hot phenol. The total ribosomal RNA was separated and purified twice by centrifugat,ion on a small-scale sucrose density-gradient.
For determination of radioactivity, the gradients were fractionated into l-ml. fractions with an ISCO fractionator. Each fraction was hydrolyzed in 0.5 N-perchloric acid at 70°C for 30 min and counted in 10 ml. of fluor-containing solution (Muramatsu et ul., 1966) using a Packard Tricarb scintillation counter.
(f) Column chromatography of 4 to 7 S RNA on Sephadex Cl00 A sample of 15 to 20 mg of 4 to 7 s RNA was applied on a 2.5 em x 80 cm column of
Sephadex Cl00 equilibrated at 4°C with 0.05 M-ammonium acetate buffer (pH 5.1); the flow rate was 2.3 to 2.5 ml./cm2/hr. Fractions of 3.7 ml. were collected and assayed for optical density at 260 rnp. The fractions of each peak were pooled separately and the RNA was precipitated overnight at -20°C by addition of 2 vol. of ethanol containing 2% potassium acetate. Rechromatography of the precipitated RNA from each peak was
carried out under the same conditions described above (Fig. 2). The purified RR’A fractions of each peak were precipitated with ethanol containing 2% potassium acetate.
(g) Anal@ of nucleotide corr~position
The RNA was hydrolyzed in 0.3 N-potassium hydroxide for IS hr at 37°C (Davidson & Mmellie, 1952). The hydrolysatc was adjusted to pH 3 to 4 with 0.5 s-perchloric acid in the cold and centrifuged to remove potassium perchlorate. The supernatant solution
338
3 o!
Iv
I
I!\ ii
J t.p
2.0 1
it
'I !!
\ !! 1
:I!
IO t r I J i 1
. i -1
/a-+-L.+ 40 60 80
Fraction number
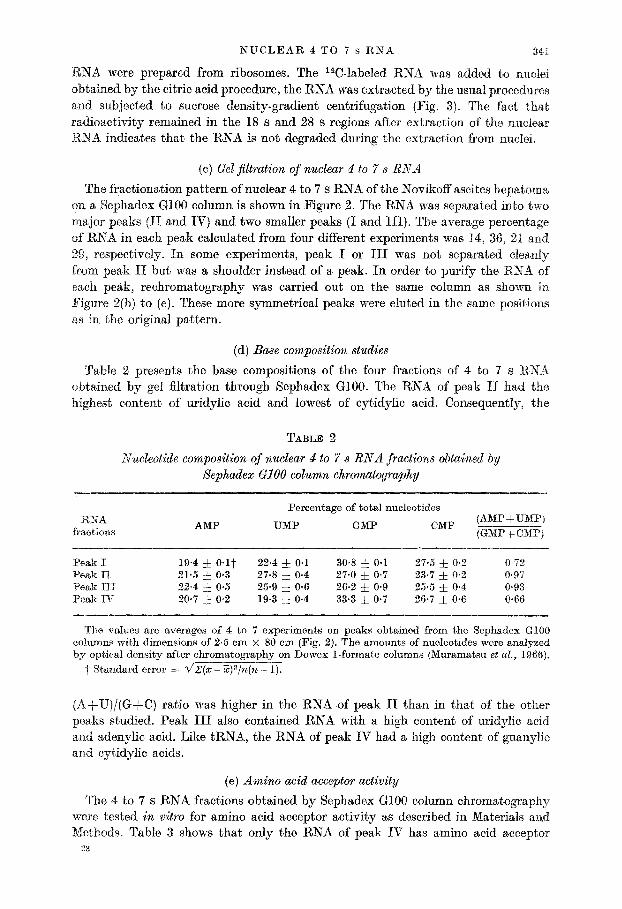
FIG. 2. Sephadex GlOO column chromatography of Novikoff ascites hepatoma nuclear 4 to 7 B RSA.
(a) Whole nuclear 4 to 7 s RNA; (b) to (e) rechromatography of RNA from peaks I to IV obtained from (a).
was placed on a Dowex l-formate column (0.5 cm x 15 cm) ; the nucleotides were elut,ed with a linear 0 to 4 x-formic acid gradient (Muramatsu et al., 1966). The fractions in each peak were pooled and after desiccation the mononueleotides were dissolved in 0.1 N-
hydrochloric acid; the optical density of individua.1 nucleotides was determined at the wavelength of maximum absorbancy for each nucleotide.
(h) AW&LO acid acceptor activity
The capacity of RNA to accept amino acids was studied as described previously (Nakamura et al., 1968). The RNA was incubated at 37°C for 30 min with a 14C-labeled amino acid mixture and aminoaeyl RNA synthetase prepared from E. co% (Nakamura et al., 1968). After the reaotion was stopped by chilling and addition of trichloroacetie acid, the acid-insoluble fraction was precipitated and the radioactivity was determined in a Packard Tricarb scintillation counter.
(i) Polpcrylamide gel elect~ophoressis
The purification of acrylamide and N,N’-methylenebisacrylamide (Eastman Organic Chemicals) was carried out as described by Locning (1967). Polyacrylamide gels (8%) were prepared in a buffer containing 0.04 If-Tris-acetic acid (pH ‘i.S), 0.02 M-sodium acetate and 0.002 M-EDTA (Loening, 1967). 10 ~1. of N,N,N’N’ tctramethylenediamine
NUCLEAR 4 TO 7 s RNA 3 3 iI
and O*GG ml. of lOo/o ammonium persulfate were added to 25 ml. of gel solution and tile mixture was allowed to polymerize in 0.5 em x 7.5 cm glass tubes, After a pre-rnr, a,t. cl mA per tube for 30 min at 0 to 4°C (Peacock & Dingman, 1967), 0.1 mg ofRil\TA samples in lo% sucrose solution were layered on the gels and eleetrophoresis was performed at 6 ma per tube for 90 min. The gels were transferred and stained overnight in a solution containing 1% acridine orange, 15% acetic acid and 2% lanthanum acetate (Gould, 1966). Destaining was carried out in 5% acetic acid. The gels were scanned for absorbanoy with a Joyce-Loebl Chromoscan Mk. II.
After scanning, the gels containing radioactive nucleic acids were sliced using a Canalco gel slicer with stainless steel blades set & in. apart. Each slice was transferred into a vial and dissolved in 30% hydrogen peroxide for 15 hr at 37°C before counting. The samples were hydrolyzed and radioa,ctivity was determined in a Packard Tricarh scintillation counter.
(j) Preparative polyacrylarnide gel electrophoresis
A Canalco prep-disc electrophoresis apparatus was used for the separation and isolat,ion of various components of 4 to ‘7 s nuclear RNA. A ll-mg sample of RNA (in 1 ml. of IO!/, sucrose) was layered on a cylinder of 8% acrylamide gel, 4 cm in height, which was polymerized as described for the analytical gel electrophoresis procedure in a P/D 320 upper column. The flush buffer and the upper electrode reservoir buffer were 0.04 af-Tris
aeetato pH 7.2 at room temperature, 0.02 M-sodium acetate and 0.002 M-EDT+&; the lower oleetrode buffer was 0.1 M-Tris acetate, pH 7.2,0.05 M-sodium acetate and 0.002 x:- EDTA. Electrophoresis was carried out at 30 mA (constant current) for 5 to 7 hr. The flow rate of flush buffer was 1 ml./min and fractions were collected at 3-min int,ervals. Elution profiles were determined by measuring the optical density of each fraction at 260 ml-L. Two drops of 5 N-perchloric acid were added to I-ml. fractions and hydrolyzed a,G 70% for 20 min. After cooling, 10 ml. of scintillation fluor was added and the samples were counted in a Packard Tricarb liquid-scintillation counter.
3. Ressllts
(a) Purity of nuclei and yield of muclear RNA
As described previously (Busch, 1967; Higashi, Narayanan, Adams & Busch, 196§), the nuclei of Novikoff hepatoma obtained by the citric acid procedure were virtually free of perinuclear cytoplasmic contamination. The yield of nuclear RNA is sho~u in Table 1. The nuclei of Novikoff hepatoma cells contain two to three times as
much RNA as those of liver cells and they compose as much as 30 to 5Oo/o of the cell volume as compared with 3 to 7% for nuclei of liver cells (Busch, 1967 ; Higashi, Xarayanan, Adams & Busch, 1966). Accordingly, a much greater quantity of nuclear
TABLE 1
Yield of RNA from nuclei of Novikoff hepatonwt isolated by the citric acid procedure
Cell fraction Yield
Cells (g/rat) Nuclei (g/l00 g tumor) Nuclear total RNA (mg/lOO g tumor) Nuclear 4 to 7 s RNA (mg/lOO g tumor)
9.4 & 0.17t 23.4 i: 1.4 64~8 & 2.6
9.0 & 0.28
t Standard error = z/X (Z - Z)“/%(n- 1). The values are averages of 6 experiments in which 15 rats were usually used. The values for
nriplei are net weights.
340 Y. MORIYAMA ET AL
RNA was isolated from equivalent weights of Novikoff hepatoma cells than from liver, i.e. the yield was 648 mg/kg as compared with 73 mg]kg for rat liver (Hodnett & Busch, 1968).
(b) Sucrose density-gradient centrifugation
The sucrose density-gradient centrifugation profile of Novikoff ascites hepatoma nuclear RNA is shown in Figure l(b). The RNA was separated by zonal ultracentri- fugation into four peaks with approximate sedimentation coefficients of 4 to 7 s, 18 s, 28 s and 45 s (Fig. l(a) ). The 4 to 7 s RNA peak was well separated from the other peaks, both by zonal centrifugation and in the analytical density gradient eentrifugation (Fig. l(b) ). The result of recentrifugation of the separated 4 to 7 s RNA on a small scale sucrose gradient is shown in Figure l(c). A single ‘symmetrical peak was found and no rapidly sedimenting RNA was present. The yield of 4 to 7 s RNA was 90 & 2.8 mgjkg Novikoff hepatoma (Table 1). In studies on rat liver the corresponding yield was 14 mg/kg (Hodnett & Busch, 1968).
In order to determine whether the nuclear 4 to 7 s RNA was a breakdown product from ribosomal RNA during extraction of RNA, purified 14C-labeled 18 s and 28 s
(a) ’ I I I I 18s
fr - 1000
O%- Ii : :
- 500
(b)
Fraction number Bottom
FIG. 3. Sucrose density-gradient sedimentation profiles of Novikoff ascites hepatoma nuclear RNA extracted from nuclear preparations (1 g) to which 14C-labeled purified ribosomal 18 s or 28 s RNA (0.4 mg) was added. The RNA was extracted by the dodecyl SOI-hot phenol procedure.
(a) r4C-labeled ribosomal 18 s RNA alone; (b) RNA from nuclei to which 18 s RNA (a) had been added; (c) ‘*C-labeled ribosomal 28 s RNA; (d) RNA from nuclei to which labeled 28 s RNA (c) had been added. (-) Optical density; (--@--a--) radioactivity.
KUCLEAR 4 TO 7 s RNA 341
RNA were prepared from ribosomes. The 14C-labeled RNA was added to nuclei obtained by t,he citric acid procedure, the RNA was extracted by the usual procedures and subjected to sucrose density-gradient centrifugation (Fig. 3). The fact, that radioactivity remained in the 18 s and 28 s regions after extraction of the nuclear RNA indicates that the RKA is not degraded during Dhe extraction from nuclei.
(c) Gel filtration of nuclear 4 to 7 s RNA
The fractionation pattern of nuclear 4 to 7 s RNA of the Novikoff ascites hepatoma. on a Sephadex GlOO column is shown in Figure 2. The RNA was separated into two major peaks (II and IV) and two smaller peaks (I and III). The average percentage of RNA in each peak calculated from four different experiments was 14, 36, 21 and 29, respectively. In some experiments, peak I or III was not separated clea,nly from peak II but was a shoulder instead of a peak. In order to purify the RNA of each peak, rechromatography was carried out on the same column as shown in Figure 2(b) to (e). These more symmetrical peaks were eluted in the same positions a,s in the original pattern.
(d) Base composition studies
Table 2 presents the base compositions of the four fractions of 4 to 7 s RXA obtained by gel filtration through Sephadex GlOO. The RNA of peak II had the highest content of uridylic acid and lowest of cytidylic acid. Consequently, the
TABLE 2
Nucleotide cornposition of nuclear 4 to 7 s RNA fractions obtained by Sephadex Cl00 column chromatography
RNA fractions AMP
Percentage of total nucleotides
UMP GMP CMP (AMPjUNP) (GMP + CMP)
Peak I 19.4 f O*l? 22.4 i 0.1 30.8 & 0.1 27.5 -& 0.2 0~72 Peak II 21.5 * 0.3 27.8 & 0.4 27.0 & 0.7 23.7 * 0.2 0.97 Peak III 22.4 & 0.5 25.9 f 0.6 26.2 f 0.9 25.5 * 0.4 0.93 Peak IV 20.7 & 0~2 19.3 * 0.4 33.3 * 0.7 26.7 f 0.6 0.66
The values are averages of 4 to 7 experiments on peaks obtained from the Sephadex GlOO columns with dimensions of 2.5 cm x 80 cm (Pig. 2). The amounts of nucleotides were analyzed by optical density after chromatography on Dowex 1-formate columns (Muramatsu et al., 1966).
t Standard error = dZ(x - Z)a/n(n - 1).
(A+U)/(G+-C) ratio was higher in the RNA of peak II than in that of the other peaks studied. Peak III also contained RNA with a high content of uridylic acid and adenyhc acid. Like tRNA: the RNA of peak IV had a high content of guanylic and cytidylic acids.
(e) Amino acid acceptor activity
The 4 to 7 s RNA fractions obtained by Sephadex GlOO column chromat’ography were tested in vitro for amino acid acceptor activity as described in Materials and Nethods. Table 3 shows that only the RNA of peak IV has amino acid acceptor
23
342 Y. MORIYAMA EI’ AL.
TABLE 3
Amino acid acceptor activity of nuclear 4 to 7 s RNA fractions obtained by Sephadex al00 column chromatography
RNA Specific activity
Amount of RNA Distribution
(ots/min/mg RNA) in e$h)peak of activity cl (%I
Peak I 6420 14 0.8 Peak II 9560 36 3.4 Peak III 8580 21 1.8 Peak IV 327,000 29 94.0
The specific activity in each peak was calculated as cts/min incorporated into acid-insoluble fractions/mg RNA. The distributions of activities were determined from the percentage of total RNA found in each peak.
01 2345
Distance moved (cm)
FIG. 4. Densitometric tracings on a Chromoscan Mk. II of the gel patterns shown in Plate I The Roman numerals indicate the peaks from which the RNA was obtained (Fig. 2). The Arabic numbers are arranged from the fastest moving band numbered 1 to the slowest moving band numbered 11.
. .
I 2 3 4 5
PLATE I
I 1 2
PLATE II
PLATE I. Polyacrylamide gel electrophoresis patterns of 4 t,o 7 s RNA fractions obt.ained b> Sephadex GIOO column chromatography (Fig. 2). Slot 1: total 4 to 7 s RSA. Slots 2 to 5: pea,ks I to IV, respectively.
PLLATE II. Polyacrglamide gel electrophoresis patterns of 4 to 7 s RNA from whole Sovikoff ascires cells (slot 1) and nuclei (slot 2).
“,$&& I ‘“,! hi
4, ‘,
:
.,-I ,i , .I,,( ‘% . .m :
I 2 3 4
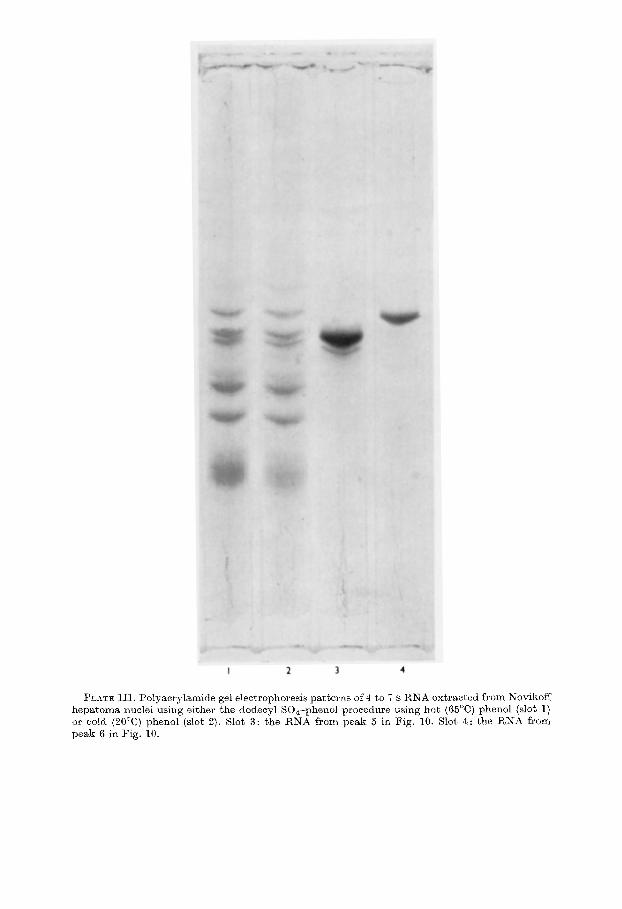
PLATE III. Polyaorylamide gel electrophoresis patterns of 4 to i s RNA extracted from Novikoff hepatoma nuclei using either t,he dodecyl SO&-phenol procedure using hot (66“C) phenol (slot I) or cold (20°C) phenol (slot 2). Slot 3: the RNA from peak 5 in Fig. 10. Slot 4: the RNA from peak 6 in Fig. 10.
NUCLEAR 4 TO 7 s RSA 343
activity, i.e. peak 1V contained more than 90% of the total acceptor activity of the rmclear 4 to 7 s RNA.
(f) Polyacrylamide gel electrophoresis
Plate I shows the polyacrylamide gel electrophoresis patterns of whole 4 to 7 s RNA and the RNA fractions obtained by Sephadex GlOO column chromatography. Densitometric traces of these gels stained with acridine orange are shown in Figure 4. in the total 4 to 7 s RNA, 11 bands were observed as seen in the upper graph of Figure 4 and slot 1 of Plate I. In peak 1: which was the most heterogeneous, bands numbered 8 to 11 were concentrated. Peak II was a mixture of bands 4 to 7 ; band 6 was the most prominent band in this peak. Two faster moving bands, 3 and 4, were found in peak III. The dense, broad, fastest moving band t,hat corresponds t,o tRKA (Richards, Co11 & Gratzer, 1965) was found in peak IV.
0
I I I I Whole ceil 4-7s RNA 2
4’ /
u
4 3
Distance moved km)
FIG. 5. Densitometrio tracings of the gel patterns shown in Plate II. t‘pper diagram: whole cell 4 to 7 s RNA; lower diagram: nuclear 4 to 7 s RNA. The whole
cell RNA is enriched in peaks 1, 2 and 4; peaks 5, 6 and 7 are enriched in the nuclear 4 to 7 s BXA as are peaks 3 and 4.
Plate XI shows the polyacrylamide gel electrophoresis patterns obtained from nuclear and whole cell 4 to 7 s RNA. The corresponding densitometric patterns are shown in Figure 5. Two dense bands 1 and 4’ which correspond to tRNA and 5 s
R,BiA were prominent, in whole cell 4 to 7 s RNA. Bands 5 to 7, the main fracti0n.s of Sephadex peak II, were found in extracts of the whole cell as well as in those of nuclei.
(8) Studies on methylation of nuclear 4 to 7 s RNA
Figure 6 shows the density-gradient sedimentation profile of nuclear RNA labeled with [~~e&.J-14C]methionine. The radioactivity distribution profiles indicate that labeling rate of 4 to 7 s RNA is somewhat slower than that of 45 s and 28 s RNA
344 Y. MORIYAMA ET AL.
"l(b) 3hr
(cl 24hr 28 5
Top Fraction number
Bottom
E'IG. ti. Sucrose density-gradient sedimentation profiles of Sovikoff ascites hepat,oma nuclear RNA labeled with L-[methyZ-14C]methionine.
Rats wwe killed at’ 30 min (a), 3 hr (b) and 24 hr (c) after injection of L-[methyZ-14C]methiorrinc. (---) Optical density; (--e--e--) radioactivity.
(Fig. 6(a) and (b) ). The incorporation of 14C into 4 to 5 s RNA continued up to 24 hours after injection of [mn,ethyE-14C]methionine ; the labeling was decreased in the high molecular weight RKA at this time. The turnover rate of the methyl groups was fastest in the 45 s RXA.
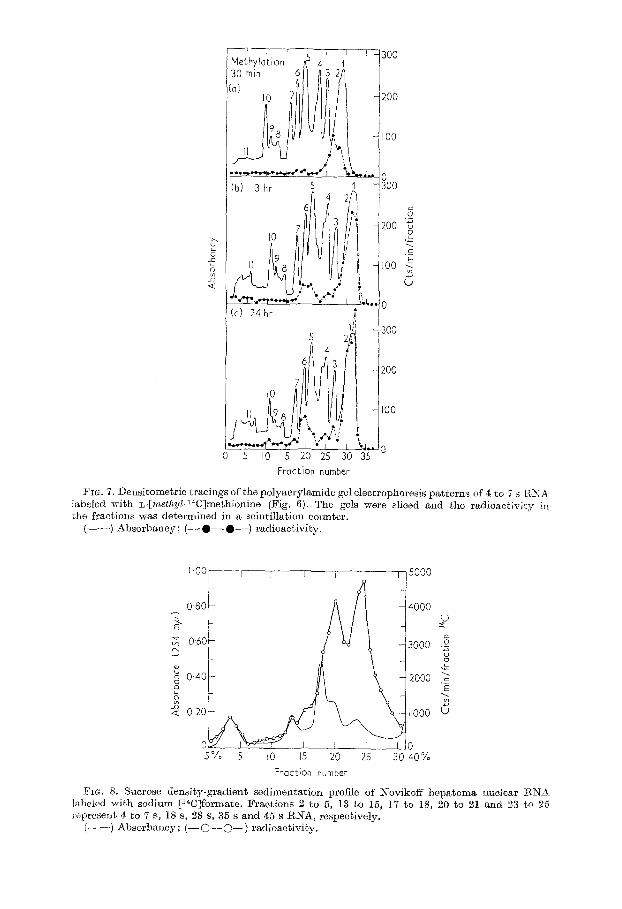
The corresponding Chromoscans from polyacrylamide gel eloctrophoresis of the 4 to 7 s RNA labeled with [ntethyl-14C]methionine are shown in Figure 7. At 30 minutes after injection of the tracer, 14C was found only in t’he tRNA region of Chromoscan peaks 1 and 2. At three hours, aradioactivepeak appearedin Chromoscan peaks 5 and 6. After 24 hours, the radioactivity distribution pattern was similar to that at t,hree hours except that more of the radioa,cbiviby was localized to Chromoscan peak 6.
(h) Studies 012: “C-labeled nuclear 4 to ri s RNA
Figure 8 shows a sucrose density-gradient centrifugat,ion profile of Novikoff hep- atoma nuclear RNA labeled with [14C]sodium formate for one hour. The marked incorporation of 14C into 35 s and 45 s nuclear RNA was similar to that reported previously with other tracers (Steele et al., 1965). The specific activity in cts/min/mg of each fraction was approximately 100,000 for 4 to 7 s and 1s S, 80,000 for 28 S,
320,000 for 35 s: 660,000 for 45 s, respectively. The 4 to 7 s RNA of Figure 8 was collected and analyzed by polyacrylamide
preparatory gel electrophoresis (Fig. 9). Six peaks were found which correspond to
Cc) 24 hr
Fraction number
FIG. 7. Densitometric tracings of the polgacrylamide gel electrophoresis patterns of 4 to 7 s I?,XA labeled with L-[met~yZ-14C]methionine (Fig. 6). The gels were sliced and the radioactivity in the fractions was determined in a scintillation counter.
(---) Absorbancy; (--O--e--) radioactivity.
E 2 0.40
b 2 020
0 5% 5 IO 15 20
- 2000 .j
Ai ’ IO00 5
25 30 cdO%
Fraction number
FIG. 8. Sucrose density-gradient sedimentation profile of Kovikoff hepatoma nuclear ENX labeled with sodium [14CJformate. Fractions 2 to 13 to 5, 15, 17 to 18, 20 to 21 and 23 to 26 represent 4 to 7 S, 18 s, 28 s, 35 s and 45 s RNA, respectively.
(-----) Absorbancy; (-O--s--) radioactivity.
346 Y. IIIORIYAMA ET AL.
Fraction number
FIG. 9. Electrophoretic pattern on a preparative polyacrylamide gel of Novikoff hepatoma nuclear 4 to 7 s RNA labeled wit,h sodium [14C]formate. The 4 to 7 s RNA was obtained from the nuclear RNA of Fig. 8. Marked differences are apparent in the specific activities for the individual RNA peaks.
(-e-o--) Absorbancy; (--O--O--) radioactivity.
Chromoscan peaks 1 to 7 on analytical gel electrophoresis (Fig. 4). The specific activity of first peak (tRNA) was about 30,000 to 60,000. A higher specific activity of 100,000 to 140,000 was found in peaks 2 and 3 which correspond to Chromoscan peaks 3 and 4 from the analytical gel. The next three peaks corresponding to Chromo- scan peaks 5 to 7, or the uridylic acid-rich fraction of Sephadex peak II, contained RNA with a low specific activity, namely, 14,000 to 16,000 cts/min/mg.
(i) PurQication and base composition of uridylic acid-rich RNA
The RNA of Sephadex peak II which contained uridylic acid-rich RNA was fractionated by preparative polyacrylamide gel electrophoresis. As shown in Figure 10, five peaks were found which correspond to bands 3 to 7 of the analytical gel electrophoresis. The RNA’s of the shadowed areas of two major peaks 5 and 6 were pooled and concentrated for analysis of base composition. The analytical poly- acrylamide gel electrophoresis patterns of these purified RNA’s are shown in Plate III. The fraction containing band 5 consisted of two closely moving bands which are also observed in whole nuclear 4 to 7 s RNA as band 5. Band 6 was electro- phoretically homogeneous.
The base compositions of the RNA of these purified fractions are shown in Table 4. B high content (26%) of uridylic acid was found in RNA of both fractions. The content of adenylic acid was higher in band 6 and that of cytidylic acid was higher in band 5.
(j) Effect of heat on extraction of nzcclear RNA
The polyacrylamide gel electrophoresis patterns of the nuclear 4 to 7 s RNA extracted with the hot phenol used in present studies and with cold phenol are
NUCLEAR 4 TO 7 s RNA
O-5
5
5s N 0.4 x .?? Et
%
2 0.2 “a 0
0 60 80 100 I20 140
347
Fraction number
Fro. 10. Electrophoretic pattern on a preparative polyacrylamide gel of the RNA of Sephadex peak II. The numerals on the peaks correspond to the band numbers of Fig. 4. The RNA’s of the shadowed areas were pooled.
TABLE 4
Ultraviolet nucleotide compositions of uridylic acid-rich fractions puri$ed by preparative gel electrophoresis
Fraction AMP UMP GMP
Rand 5 18.7 & 0*05 25.7 & 0.0 32.7 + 0.1 22.8 & 0.15 0.80 Band 6 22.2 f 1.1 26.1 -& 0.05 32.9 * 0.9 18.8 f 0.2 0.94
The values are averages of two experiments. The amounts of nucleotides were analyzed by opt,&1 density aft’er chromatography on Dowex 1-format’e columns (Muramatsu et al., 1966).
shown in Plate III slots 1 and 2. Since uridylic acid-rich RNA was extracted equally well in both cases, it cannot be a, degradation product caused by heating (Pene et al., 1968).
4. Discussion (a) Fractionation of the nuclear 4 to 7 s BNA
To fractionate and characterize nuclear 4 to 7 s RNA, a satisfactory source was
sought, and Novikoff hepatoma nuclei isolated by the cit,ric acid procedure (Higashi, Adams & Busch, 1966; Higashi, Narayanan, Adams & Busch, 1966) were found to have the advant,age of ready isolation, a large content of this RNA and a low content of RNase (Chakravorty & Busch, 1967).
The 4 to 7 s RKA, isolated from whole nuclear RNA of the Novikoff ascites hepatoma cells by zonal ultracentrifugation, is a complex mixture of RNA’s tha,t can be separated into a number of subspecies by chromatography on Sephadex Cl00 and by polyacryIamide gel electrophoresis. Whole nuclear 4 to 7 s RKA from NovikofT hepatoma cells is apparently more complex than the nucleolar 4 to 7 s RNA since 11 bands were found in the whole nuclear RNA compared to the three major a,nd three minor ba’nds found in the nucleolar RNA (Nakamura et al., 1968):
348 Y. MORIYAMA ET AL.
Partial purification of the 4 to 7 s RNA was carried out using exclusion chromato- graphy on Sephadex GlOO, and the peak II which was the most prominent peak was found to contain uridylic acid-rich RNA. Polyacrylamide gel electrophoresis of RNA in peak II showed that at least five bands were present. The uridylic acid-rich RNA’s contained in bands 5 and 6 were separated by preparatory disc electrophoresis. The RNA of band 6 was isolated as an electrophoretically homogeneous fraction which had a higher content of adenylic acid and a lower content of uridylic acid than the RNA in band 5.
(b) Intranuclear localization of the ~uridylic acid-rich low molecular weight RNA
Similar types of low molecular weight uridylic acid-rich RNA’s were found in the nucleoli of Novikoff hepa,toma cells (Nakamura et al., 1968) and in salt-extracted chromatin fractions of rat liver and Novikoff hepatoma cells (Prestayko & Busch, 1968). Since these uridylic acid-rich RNA’s seem to be located only in the nuclei, they apparently differ from the low molecular weight RNA’s found in cytopla’smic fractions such as rat liver microsomes (Gardner & Hoagland, 1968) and HeLa cell ribosomes (Pene et al., 1968). Although the tot,al number of these RNA’s has not yet been defined, it seems clear that a number of different uridylic acid-rich RNB’s are present in the nuclei. Moreover, different bands are apparently concentrated in the nucleolar (Nakamura et al., 1968) and the whole nuclear RNA.
(c) Evidence that the uridylic acid-rich RNA is not a degradation product
Slthough the possibility was considered that the uridylic acid-rich RNA might be a degradation product of high molecular weight RNA, such as 18,28,35 or 45 s RNA, this is excluded by a number of studies. (1) The amount of the RNA in peak II eluted from Sephadex GlOO was almost constant and the polyacrylamide gel electro- phoresis patterns of nuclear 4 to 7 s RNA were reproducible ; (2) the base composition of uridylic acid-rich RNA was different, from that of the other high molecular weight RNA; (3) the content of RNase in Novikoff hepatoma nuclei is low; (4) RNase activity was inhibited at the low pH of the citric acid used for the isolation of nuclei; (5) no degradation of added ribosomal 18 s and 28 s RNA was observed during extraction of RNA with phenol; (6) since uridylic acid-rich RNA bands 5 and’ 6 were found in whole cell, they cannot be a breakdown product during the process of isolation of nuclei; (7) uridylic acid-rich RNA wa’s extracted from nuclei equally well with hot or cold phenol; (8) the labeling patterns of the uridylic acid-rich RNA with either sodium [14C]formate or [methyZ-14C]methionine differs from that of tRNA, 5 s RNA and high molecular weight RNA; (9) similar types of uridylic acid-rich RNA were obtained from rat liver nuclei isolated by the sucrose procedure of Chauveau, Moule & Rouiller (1956) in other studies in this laboratory (Hodnett & Busch, 1968).
(d) Punctions of the low molecular weight uridylic acid-rich RNA
The functions of uridylic acid-rich RNA’s in the nuclei are unknown at present. Although the uridylio acid-rich RNA’s may be messenger RNA’s, their turnover rates are quite slow and the amount of this RNA is relatively large. It can be assumed that, the uridylic acid-rich RNA’s are relatively stable components of nuclei which may have a structural role or a role in the control of gene expression.
SUCLEAR 4 TO 7 s RNA 349
We thank Dr Sudhir Kumar for assays of amino acid accept,or activity, Mr Joe Arendeil for the zond ultracentrifuge runs and Mrs Rose K. Busch for supplying the transplanted tumors.
These studies were supported in part by grants from the American Cancer Society, the Jane Coffin Childs Fund, the Xational Science Foundat,ion and the U.S. Public Health Service (CA08182 CAlOS93).
REFERENCES
Brownlee, 61. G., Sanger, F. & Barrell, B. G. (1968). J. MoZ. Biol. 34, 379. Busch, H. (1967). In Methods in Enzymology, ed. by L. Grossman & K. Moldave, vol. 12,
p. 434. Busch, H. & Desjardins, A. (1965). Exp. Cell Res. 40, 127. Chakravorty, A. K. & Busch, H. (1967). Cancer Res. 27, 789. Chauveau, J., Moule, Y. & Rouiller, C. (1956). Exp. Cell Res. 11, 3 17. Davidson, J. N. & Smellie, R. M. S. (1952), Biochem. J. 52, 594. Forget, B. G. B: Weissman, S. M. (1967). Science, 158, 1695. Gardner, J. A. A. & Hoagland, M. B. (1968). J. BioZ. Chem. 243, 10. Gould, H. (1966). Biochemistry, 5, 1103. Bigashi, K., Adams, H. R. & Busch, H. (1966). Cancer Res. 26, 2196. Bigashi, K., Narayanan, K. S., Adams, II. R. & Busch, H. (1966). Cn.ncer Res. 26, 1582. Hodnett, J. L. & Busch, H. (1968). J. Biol. Chem. 243, 6336. Holley, R. W., Apgar, J., Everett, G. A., Madison, J. T., Narquisee, M., Merrill, S. II.,
Penswick, J. R. & Zamir, A. (1965a). Science, 147, 1462. .Holley, R. W., Everett, G. A., Madison, J. T. & Zamir, A. (1965b). J. Biol. Chem. 240, 2122. Larsen, C. J., Galibert, F., Lelong, J. C. & Boiron, M. (1967). C. R. -4cad. Sci. Pwis,
204, 1523. Loening, U. E. (1967). B&hem. J. 102, 251. Mummatsu, M. & Busch, H. (1965). J. Biol. Chem. 240, 3960. Murarnat,su, M., Hodnett, J. L. & Busch, H. (1966). J. Biol. Chem. 241, 1544. h’akamura, T., Prestayko, A. W. & Busch, H. (1968). J. Biol. Chem. 243, 1368. Okamura, Iz’. & Busch, H. (1965). Cuncer Res. 25, 693. Peacock, A. C. & Dingman, C. W. (1967). Biochemistry, 6, 1818. Prestayko, A. W. & Busch, H. (1968). Biochim. biophys. Acta, 169, 339. Pene, J. J., Knight, E., Jr. $ Darnell, J. E., Jr. (1968). J. Mol. BioZ. 33, 609. Richards, E. G., Coil, J. A. & Gratzer, W. B. (1965). Analvt. &o&em,. 12, 452. Steele, W. J., Okamura, N. & Busch, H. (1965). J. BioZ. Ch)em. 240, 1742. Ta,kanami, M. (I 961). Biochim. biophys. Acta, 51, 81.