Evidence that the insertion events of IS2 transposition are biased towards abrupt compositional shifts in target DNA and modulated by a diverse set of culture parameters Geisa A. L. Gonçalves 1,7,‡ , Pedro H. Oliveira 2,3,‡ , Ana G. Gomes 4 , Kristala L. J. Prather 5,7 , Leslie A. Lewis 6 , Duarte M. F. Prazeres 1,7 and Gabriel A. Monteiro 1,7,* 1 Institute for Biotechnology and Bioengineering (IBB), Centre for Biological and Chemical Engineering, Department of Bioengineering, Instituto Superior Técnico, Lisbon, Portugal. 2 Institut Pasteur, Microbial Evolutionary Genomics, Département Génomes et Génétique, F- 75015 Paris, France. 3 CNRS, UMR3525, F-75015 Paris, France. 4 Instituto Politécnico de Setúbal, ESTBarreiro, 2839-001 Barreiro, Portugal. 5 Department of Chemical Engineering, Massachusetts Institute of Technology, Room E17- 504G, Cambridge, MA 02139, USA. 6 Department of Biology, York College of the City University of New York, Jamaica, New York 11451, USA. 7 MIT-Portugal Program ‡ Equal contribution from both authors * Corresponding author: Gabriel A. Monteiro, Ph.D. Phone: +351 218 419 195 Fax: +351 218 419 062 E-mail: [email protected]Supplementary Material Applied Microbiology and Biotechnology
Transcript
Evidence that the insertion events of IS2 transposition are biased towards abrupt
compositional shifts in target DNA and modulated by a diverse set of culture
parameters
Geisa A. L. Gonçalves1,7,‡, Pedro H. Oliveira2,3,‡, Ana G. Gomes4, Kristala L. J.
Prather5,7, Leslie A. Lewis6, Duarte M. F. Prazeres1,7 and Gabriel A. Monteiro1,7,*
1 Institute for Biotechnology and Bioengineering (IBB), Centre for Biological and Chemical
Engineering, Department of Bioengineering, Instituto Superior Técnico, Lisbon, Portugal. 2 Institut Pasteur, Microbial Evolutionary Genomics, Département Génomes et Génétique, F-
75015 Paris, France. 3 CNRS, UMR3525, F-75015 Paris, France. 4 Instituto Politécnico de Setúbal, ESTBarreiro, 2839-001 Barreiro, Portugal. 5 Department of Chemical Engineering, Massachusetts Institute of Technology, Room E17-
504G, Cambridge, MA 02139, USA. 6 Department of Biology, York College of the City University of New York, Jamaica, New
Supplementary material S1 List of references from which IS2 target sequences were obtained Bernard N, Ferain T, Garmyn D, Hols P, Delcour J (1991) Cloning of the D-lactate
dehydrogenase gene from Lactobacillus delbrueckii subsp. bulgaricus by complementation in Escherichia coli. FEBS Lett 290(1-2):61-4
Cohen SP, Hachler H, Levy SB (1993) Genetic and functional analysis of the multiple antibiotic resistance (mar) locus in Escherichia coli. J Bacteriol 175(5):1484-92
Kim M, Ryu S (2011) Characterization of a T5-like coliphage, SPC35, and differential development of resistance to SPC35 in Salmonella enterica serovar typhimurium and Escherichia coli. Applied and environmental microbiology 77(6):2042-50
Kobayashi K, Tsukagoshi N, Aono R (2001) Suppression of hypersensitivity of Escherichia coli acrB mutant to organic solvents by integrational activation of the acrEF operon with the IS1 or IS2 element. J Bacteriol 183(8):2646-53
Lewis LA, Gopaul S, Marsh C (1994) The non-random pattern of insertion of IS2 into the hemB gene of Escherichia coli. Microbiol Immunol 38(6):461-5
Oliveira PH, Prazeres DM, Monteiro GA (2009) Deletion formation mutations in plasmid expression vectors are unfavored by runaway amplification conditions and differentially selected under kanamycin stress. J Biotechnol 143(4):231-8
Parker LL, Hall BG (1990) Mechanisms of activation of the cryptic cel operon of Escherichia coli K12. Genetics 124(3):473-82
Rijavec M, Budic M, Mrak P, Muller-Premru M, Podlesek Z, Zgur-Bertok D (2007) Prevalence of ColE1-like plasmids and colicin K production among uropathogenic Escherichia coli strains and quantification of inhibitory activity of colicin K. Applied and environmental microbiology 73(3):1029-32
Sengstag C, Arber W (1983) IS2 insertion is a major cause of spontaneous mutagenesis of the bacteriophage P1: non-random distribution of target sites. The EMBO journal 2(1):67-71
Szeverenyi I, Hodel A, Arber W, Olasz F (1996) Vector for IS element entrapment and functional characterization based on turning on expression of distal promoterless genes. Gene 174(1):103-10
Takimoto K, Tachibana A, Ayaki H, Yamamoto K (1997) Spectrum of spontaneous mutations in the cyclic AMP receptor protein gene on chromosomal DNA of Escherichia coli. Journal of radiation research 38(1):27-36
List of references from which IS1 target sequences were obtained Barker CS, Pruss BM, Matsumura P (2004) Increased motility of Escherichia coli by
insertion sequence element integration into the regulatory region of the flhD operon. J Bacteriol 186(22):7529-37
Feeney MA, Ke N, Beckwith J (2012) Mutations at several loci cause increased expression of ribonucleotide reductase in Escherichia coli. J Bacteriol 194(6):1515-22
Jovanovic M, Lilic M, Savic DJ, Jovanovic G (2003) The LysR-type transcriptional regulator CysB controls the repression of hslJ transcription in Escherichia coli. Microbiology 149(Pt 12):3449-59
Karow M, Fayet O, Georgopoulos C (1992) The lethal phenotype caused by null mutations in the Escherichia coli htrB gene is suppressed by mutations in the accBC operon, encoding two subunits of acetyl coenzyme A carboxylase. J Bacteriol 174(22):7407-18
Kobayashi K, Tsukagoshi N, Aono R (2001) Suppression of hypersensitivity of Escherichia coli acrB mutant to organic solvents by integrational activation of the acrEF operon with the IS1 or IS2 element. J Bacteriol 183(8):2646-53
La Ragione RM, Collighan RJ, Woodward MJ (1999) Non-curliation of Escherichia coli O78:K80 isolates associated with IS1 insertion in csgB and reduced persistence in poultry infection. FEMS microbiology letters 175(2):247-53
Manna D, Wang X, Higgins NP (2001) Mu and IS1 transpositions exhibit strong orientation bias at the Escherichia coli bgl locus. J Bacteriol 183(11):3328-35
Petersen C, Moller LB, Valentin-Hansen P (2002) The cryptic adenine deaminase gene of Escherichia coli. Silencing by the nucleoid-associated DNA-binding protein, H-NS, and activation by insertion elements. The Journal of biological chemistry 277(35):31373-80
Umenhoffer K, Feher T, Baliko G, Ayaydin F, Posfai J, Blattner FR, Posfai G (2010) Reduced evolvability of Escherichia coli MDS42, an IS-less cellular chassis for molecular and synthetic biology applications. Microbial cell factories 9:38
Zerbib D, Gamas P, Chandler M, Prentki P, Bass S, Galas D (1985) Specificity of insertion of IS1. J Mol Biol 185(3):517-24
Fig. S1: Skew analysis and insertion site in pCIneo. (A) Profiles of GC and AT skew
for plasmid pCIneo. A large shift in DNA composition is observed in the region
upstream of the neoR gene. PCR amplification of this region led to the identification
of a spontaneous IS2 insertion (red arrow). (B) Three-dimensional representation of a
0.4 kb region flanking the insertion site (shown in green). The region shows intrinsic
curvature close to the insertion site, and concomitantly, close to the sequences
showing abrupt compositional shifts. Start and end positions of the DNA fragment are
indicated. (C) Schematic representation of the pCIneo::IS2 plasmid, highlighting the
hybrid promoter generated between the 3’ end of IS2 (purple) and the neoR gene
(green). Capital letters match the E. coli promoter consensus sequence TTGACAN16–
18TATAAT. The 5 bp duplication generated upon insertion is shown as stippled box.
Two putative -10 regions are indicated.
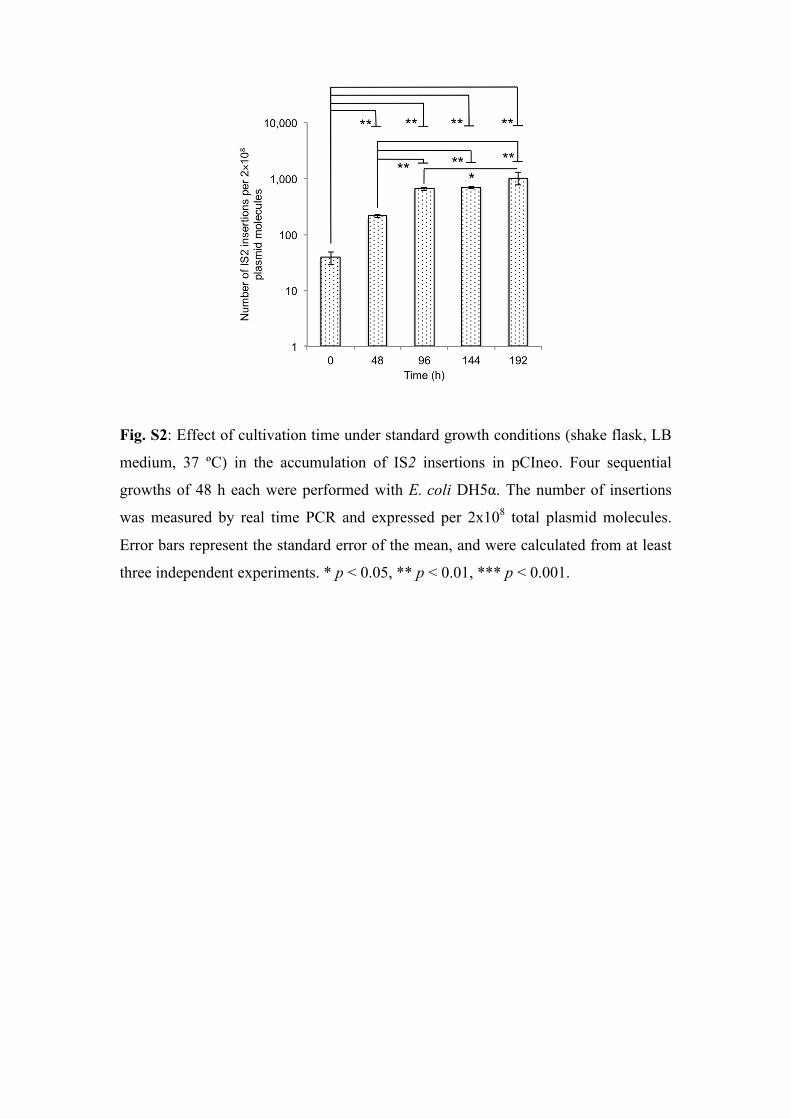
Fig. S2: Effect of cultivation time under standard growth conditions (shake flask, LB
medium, 37 ºC) in the accumulation of IS2 insertions in pCIneo. Four sequential
growths of 48 h each were performed with E. coli DH5α. The number of insertions
was measured by real time PCR and expressed per 2x108 total plasmid molecules.
Error bars represent the standard error of the mean, and were calculated from at least
three independent experiments. * p < 0.05, ** p < 0.01, *** p < 0.001.
Fig. S3: Relation between DNA compositional asymmetries and IS1 transposition. A
total of 15 different IS1 target sites from genomic and plasmid DNA were used (see
Suppl. Material S1). (A) Average profiles of GC and AT skew 550 bp upstream and
downstream of the insertion point of IS1. (B) Average of maximum amplitude in GC
skew and AT skews in the same window length used for skew analysis at observed
and random IS1 target sites. Error bars represent the standard error of the mean. A
more permissive threshold (p < 0.1) was allowed in this particular analysis to allow
for increased sensitivity and in order not to miss positive results.* p < 0.1.