The Oligoadenylate Synthetase Family: An Ancient Protein Family with Multiple Antiviral Activities Helle Kristiansen, 1 Hans Henrik Gad, 2 Signe Eskildsen-Larsen, 1,3 Philippe Despres, 2 and Rune Hartmann 1 The 2 0 -5 0 oligoadenylate synthetases (OAS) are interferon-induced antiviral enzymes that recognize virally produced dsRNA and initiate RNA destabilization through activation of RNase L within infected cells. How- ever, recent evidence points toward several RNase L–independent pathways, through which members of the OAS family can exert antiviral activity. The crystal structure of OAS led to a novel insight into the catalytic mechanism, and revealed a remarkable similarity between OAS, Polyadenosine polymerase, and the class I CCA-adding enzyme from Archeoglobus fulgidus. This, combined with a variety of bioinformatic data, leads to the definition of a superfamily of template independent polymerases and proved that the OAS family are ancient proteins, which probably arose as early as the beginning of metazoan evolution. Introduction T he 2 0 -5 0 oligoadenylate synthetase (OAS) family of proteins are members of an ancient class of template- independent RNA polymerases, along with the eukaryotic polyadenosine polymerase (PAP) and the class I CCA add- ing enzyme (CCA) from Archaea (Torralba and others 2008). We presume that the OAS proteins diverged from PAP and CCA at the beginning of metazoan (multicellular animals) evolution. This view is supported by the presence of OAS proteins in some of the oldest metazoans, sea sponges (Por- ifera) (Wiens and others 1999). The important role played by the OAS proteins in the defense against viral infections in mammalians is well documented, whereas the biological role of recently identified OAS proteins in lower animals, par- ticularly invertebrates, is less clear. The OAS proteins identify viral infection by binding to (viral derived) dsRNA. This results in an allosteric activation of OAS followed by synthesis of 2 0 -5 0 oligoadenylates (2-5A), which in turn bind and activate the latent RNase, RNase L. Upon 2-5A binding, RNase L dimerizes and degrades RNA present in the cell, cellular as well as viral, leading to the inhibition of protein synthesis (Clemens and Williams 1978; Dong and Silverman 1997). This is the classical OAS/RNase L pathway. Recently, additional exciting information regard- ing RNase L–independent antiviral pathways has become available. Here, we review both the RNase L–dependent and RNase L–independent functions of the OAS family, as well as providing an overview of the structure, function, and evolution of this interesting family of proteins. The Oligoadenylate Synthetase Family: Genome Organization, Protein Structure, and Catalytic Mechanism The OAS genes The human OAS family is a multigene family that consists of 4 genes, OAS1, OAS2, OAS3, and OASL. All 4 genes are actively induced by interferon. In addition, OASL, but not OAS1, is induced directly in response to viral infection (interferon-regulatory factor [IRF]-3 dependent) (Melchjor- sen and others 2009). Further, traces of an OASL2 pseudo- gene is found next to the OASL gene (Eskildsen and others 2003). The OAS genes are located on the human chromosome 12 with OAS1 to -3 clustered in the 12q24.1 region and OASL at 12q24.2 (Hovnanian and others 1998; Justesen and others 2000). All mammalian OAS genes share a general exon/in- tron structure, consisting of a basal OAS unit, encoded by 5 exons. This unit is repeated once in OAS1 and OASL, twice in OAS2, and thrice in OAS3. All the OAS genes contain 1 (OAS3 and OASL) or 2 (OAS1 and OAS2)3 0 exons, and these give rise to splice variants of OAS1 (designated p42, p44, p46, and p48) and OAS2 (designated p69 and p71) (Benech and others 1985; Ghosh and others 1991; Marie and Hova- nessian 1992). Only 1 splice variant of OAS3 has been identified (Rebouillat and others 1999). Two groups simul- taneously identified the fourth member of the OAS family, OASL (Hartmann and others 1998b; Rebouillat and others 1998). The OASL gene shares high homology with the other OAS family members; however, it is devoid of enzymatic activity (Marques and others 2008). Further, it contains a 3 0 1 Centre for Structural Biology, Department of Molecular Biology, Aarhus University, Aarhus, Denmark. 2 Unite ´ Interactions Mole ´culaires Flavivirus-Ho ˆ tes, Institut Pasteur, Paris, France. 3 Novozymes A/S, Copenhagen, Denmark. JOURNAL OF INTERFERON & CYTOKINE RESEARCH Volume 31, Number 1, 2011 ª Mary Ann Liebert, Inc. DOI: 10.1089/jir.2010.0107 41
Transcript
The Oligoadenylate Synthetase Family: An Ancient ProteinFamily with Multiple Antiviral Activities
Helle Kristiansen,1 Hans Henrik Gad,2 Signe Eskildsen-Larsen,1,3 Philippe Despres,2 and Rune Hartmann1
The 20-50 oligoadenylate synthetases (OAS) are interferon-induced antiviral enzymes that recognize virallyproduced dsRNA and initiate RNA destabilization through activation of RNase L within infected cells. How-ever, recent evidence points toward several RNase L–independent pathways, through which members of theOAS family can exert antiviral activity. The crystal structure of OAS led to a novel insight into the catalyticmechanism, and revealed a remarkable similarity between OAS, Polyadenosine polymerase, and the class ICCA-adding enzyme from Archeoglobus fulgidus. This, combined with a variety of bioinformatic data, leads tothe definition of a superfamily of template independent polymerases and proved that the OAS family are ancientproteins, which probably arose as early as the beginning of metazoan evolution.
Introduction
The 20-50 oligoadenylate synthetase (OAS) family ofproteins are members of an ancient class of template-
independent RNA polymerases, along with the eukaryoticpolyadenosine polymerase (PAP) and the class I CCA add-ing enzyme (CCA) from Archaea (Torralba and others 2008).We presume that the OAS proteins diverged from PAP andCCA at the beginning of metazoan (multicellular animals)evolution. This view is supported by the presence of OASproteins in some of the oldest metazoans, sea sponges (Por-ifera) (Wiens and others 1999). The important role played bythe OAS proteins in the defense against viral infections inmammalians is well documented, whereas the biological roleof recently identified OAS proteins in lower animals, par-ticularly invertebrates, is less clear.
The OAS proteins identify viral infection by binding to(viral derived) dsRNA. This results in an allosteric activationof OAS followed by synthesis of 20-50 oligoadenylates (2-5A),which in turn bind and activate the latent RNase, RNase L.Upon 2-5A binding, RNase L dimerizes and degrades RNApresent in the cell, cellular as well as viral, leading to theinhibition of protein synthesis (Clemens and Williams 1978;Dong and Silverman 1997). This is the classical OAS/RNaseL pathway. Recently, additional exciting information regard-ing RNase L–independent antiviral pathways has becomeavailable. Here, we review both the RNase L–dependent andRNase L–independent functions of the OAS family, as wellas providing an overview of the structure, function, andevolution of this interesting family of proteins.
The Oligoadenylate Synthetase Family:Genome Organization, Protein Structure,and Catalytic Mechanism
The OAS genes
The human OAS family is a multigene family that consistsof 4 genes, OAS1, OAS2, OAS3, and OASL. All 4 genes areactively induced by interferon. In addition, OASL, but notOAS1, is induced directly in response to viral infection(interferon-regulatory factor [IRF]-3 dependent) (Melchjor-sen and others 2009). Further, traces of an OASL2 pseudo-gene is found next to the OASL gene (Eskildsen and others2003). The OAS genes are located on the human chromosome12 with OAS1 to -3 clustered in the 12q24.1 region and OASLat 12q24.2 (Hovnanian and others 1998; Justesen and others2000). All mammalian OAS genes share a general exon/in-tron structure, consisting of a basal OAS unit, encoded by 5exons. This unit is repeated once in OAS1 and OASL, twice inOAS2, and thrice in OAS3. All the OAS genes contain 1(OAS3 and OASL) or 2 (OAS1 and OAS2) 30 exons, and thesegive rise to splice variants of OAS1 (designated p42, p44,p46, and p48) and OAS2 (designated p69 and p71) (Benechand others 1985; Ghosh and others 1991; Marie and Hova-nessian 1992). Only 1 splice variant of OAS3 has beenidentified (Rebouillat and others 1999). Two groups simul-taneously identified the fourth member of the OAS family,OASL (Hartmann and others 1998b; Rebouillat and others1998). The OASL gene shares high homology with the otherOAS family members; however, it is devoid of enzymaticactivity (Marques and others 2008). Further, it contains a 30
1Centre for Structural Biology, Department of Molecular Biology, Aarhus University, Aarhus, Denmark.2Unite Interactions Moleculaires Flavivirus-Hotes, Institut Pasteur, Paris, France.3Novozymes A/S, Copenhagen, Denmark.
JOURNAL OF INTERFERON & CYTOKINE RESEARCHVolume 31, Number 1, 2011ª Mary Ann Liebert, Inc.DOI: 10.1089/jir.2010.0107
41
exon that shares homology with 2 consecutive ubiquitinrepeats (Hartmann and others 1998b).
Oas gene organization in mice. The rodent Oas gene locushas undergone a recent series of duplications event, leadingto the presence of 8 mOas1 genes in the murine genome(Eskildsen and others 2002; Mashimo and others 2002; Per-elygin and others 2002). In addition to the 8 complete mOas1genes, we found a group of 3 exons (exon C, D, and E) and asingle exon E within the Oas loci (Eskildsen and others 2002).These duplications appear to have occurred after the sepa-ration of the rodent order, and are thus specific to rodents.For a comprehensive study of the mammalian OAS1 geneloci, see Perelygin and others (2006). We also observed thatthe mOas1a and mOas1g genes, which are almost identical inamino acid composition, are located within 2 18 kb DNArepeat sequences, suggesting that they originate from a re-cent DNA duplication. Further, 4 of the 8 mOas1 genes aretranscribed in the centromere to telomere direction (mOas1a,mOas1c, mOas1e, and mOas1g) and 4 in the opposite direction
(mOas1b, mOas1d, mOas1f, and mOas1h) as indicated by theirposition either above or below the chromosome in Fig. 1.
Sequence analysis of the active site of the mOas1 proteinsencoded by the various genes revealed that they could bedivided into 2 groups. The sequence of mOas1a and mOas1gproteins suggests that they are active enzymes, which alsoagree with experiential information. On the contrary, theremaining mOas1 proteins contain changes within the activesite, which are considered to be incompatible with activity.Experiential evidence from Kakuta and others (2002) sup-ports this division of mOas1 proteins into a group of active(mOas1a and -g) and a group of catalytic inactive proteins(mOas1b, -c, -d, -e, -f, and -h). However, real-time polymerasechain reaction analysis revealed that all 8 mOas1 genes areactively transcribed and is induced by interferon (Eskildsenand others 2002).
In a pioneering effort using positional cloning, 2 groupssimultaneously identified the murine Oas1b gene as respon-sible for restricting West Nile virus (WNV) replication in
FIG. 1. Genomic structure of the human and murine OAS genes. (A) The human OAS genomic structure. (B) The murineOas genomic structure. Squares above the line indicate genes transcribed in the centromere-to-telomere direction; squaresbelow the line indicate genes transcribed in the telomere-to-centromere direction. Genes are not drawn to scale. OAS, 20-50
oligoadenylate synthetase; TEL, telomere; CEN, centromere; CDE, Exon duplications, see text for further explanation.
FIG. 2. Structure and active site of OAS. (A) Crystal structure of the porcine OAS1. (B) Close-up of the active site of OAS1.Residues demonstrated to be involved in catalysis by site-directed mutagenesis are shown in ball and stick, yellow indicatesimpaired activity if mutated to alanine, and red indicates no activity if mutated to alanine. A model, based on mutagenesisdata, of the assumed position of the donor ATP is shown.
42 KRISTIANSEN ET AL.
wild mice (Mashimo and others 2002; Perelygin and others2002). In the majority of laboratory mouse strains, this genecontains a premature stop codon, leading to the productionof a truncated protein. The antiviral activity of the mOas1bprotein is discussed later.
Structure of the OAS proteins
The determination of the porcine OAS1 crystal structureprovided valuable insight into the enzymatic mechanism ofthe OAS family. One OAS unit forms 1 defined globularstructure, containing 2 domains: an N-terminal b-sheet do-main, containing 5 anti-parallel b-strands, and a C-terminal, a-helical domain (Hartmann and others 2003). The 2 domainsare connected by a linker region and a short N-terminal ex-tension. The N-terminal extends across to the C-terminal do-main, and secures the 2 domains together (Fig. 2A).
In the following, we discuss the structure of OAS and theimplications for the catalytic function. Before doing so, wewould like to clarify the nomenclature used by us. OASbinds 2 molecules of ATP: the donor and acceptor ATP. Thedonor ATP is the molecule of ATP added to the growingchain of 2-5As, whereas the acceptor ATP provides the free 20
hydroxyl to which the AMP moiety of the donor ATP isadded. The acceptor ATP can be replaced by a growing chainof 2-5As. In terms of catalysis, the acceptor ATP is equivalentof the primer in other polymerases.
The active site of OAS is located on the interface betweenthe N-terminal and C-terminal domains, and consists ofresidues from both domains (Fig. 2B). The N-terminal do-main contains 3 aspartic acids, D74, D76, and D147 (num-bered according to the porcine OAS1), which coordinate the2 Mg2þ ions critical for catalysis (Hartmann and others 2003).The coordination of 2 catalytic Mg2þ ions is a feature con-served among all nucleotidyl transferase enzymes (see alsoThe unique 20 polymerase activity and the origin of the OASproteins section) (Holm and Sander 1995). The N-terminaldomain also contains a b–turn between strand 1 and 2 (alsoknown as a phosphate binding loop or the Walker A motif),responsible for binding the triphosphate of the incomingdonor ATP. The C-terminal domain contains 2 basic residues(R209 and K212 in the porcine OAS1), which are presumedto neutralize the positive charge on the phosphates of thedonor ATP. These residues are conserved in all catalytic ac-tive members of the OAS family, as well as in both PAP andCCA. Mutation of these residues leads to a strong impair-ment of catalytic activity in the OAS enzymes (Sarkar andothers 1999b; Hartmann and others 2003), as well as PAPand CCA (Martin and others 1999; Cho and others 2005).
The C-terminal domain also contains several residues,which, based upon mutational studies, are believed to interactwith the base moiety of the donor ATP (Torralba and others2008) (see Fig. 2 for details). These residues are found withinthe second helix and a small b-loop-b motif, located betweenhelix 3 and 4, and are conserved in all OAS family members, aswell as in CCA, but not in PAP. Interestingly, the active siteresidues in the C-terminal 4-helix bundle domain are all con-served in the inactive OASL protein, whereas the N-terminalb-sheet domain in OASL has diverged substantially from theother members of the OAS family; in particular, the last of theessential aspartic acids is absent from OASL.
At present, we know little about the interaction with theacceptor nucleotide. Obviously, the 20 hydroxyl has to be in a
position to react with the a-phosphate of the donor, and thusthe ribose of the acceptor must be positioned in a differentway from that seen in 30 specific polymerases.
The unique 2 0 polymerase activity and the originof the OAS proteins
The formation of 20 linked oligonucleotides is a uniqueproperty of the OAS proteins. Nevertheless, the fundamentalchemical reaction between the formation of 20 and 30 linkedoligonucleotides is similar. A unified mechanism utilizing 2metal ions was proposed for the phosphoryl transfer reaction ofall polymerases (Delarue and others 1990; Beese and Steitz 1991;Steitz and others 1994; Steitz 1999). This naturally raised thequestion if the OAS family would adhere to this mechanism.
First part of the answer came from bioinformatics. Usingstructure-guided alignments, the OAS family was classifiedinto the nucleotidyl transferase superfamily based on theconservation of a specific sequence motif in the active site(Holm and Sander 1995). Martin and Keller worked exten-sively on the characterization of PAP, and noted a conserva-tion of several specific motifs between the OAS family and thefamily of eukaryotic PAPs (Martin and others 1999; Martinand Keller 2007). PAP and OAS are both template-indepen-dent, RNA-specific ribonucleotidyl transferases using ATP assubstrate. The enzymatic activity of OAS differs from PAP in 2important ways: they add nucleotides to the 20 hydroxyl in-stead of the 30, and the OAS enzymes are primer independent,whereas PAP adds the polyA tail to the 30 end of mRNA.
The crystal structure of porcine OAS1 confirmed the re-lation between OAS and other nucleotidyl transferases, andhelped define a new family of structurally related template-independent polymerases. It revealed a remarkable similar-ity to the structure of the catalytic core of both PAP and classI CCA adding enzymes. These 3 enzymes have a common 2domain catalytic core, equivalent to 1 OAS unit as describedabove. The classical 5-stranded b-sheet domain is found in allnucleotidyl transferases (the palm domain in the handanalogy used by Steitz and others) and the C-terminal do-main is a 4-helix bundle domain, which replaces the thumbsand finger domains found in template dependent polymer-ases. Together, these 3 families are proposed to constitute asuperfamily of template independent polymerases (Torralbaand others 2008), which, we believe, shares a common evo-lutionary origin. Structure-guided mutagenesis confirmedthat catalysis is performed in a highly similar way in these 3enzymes, despite OAS being 20 specific. This leads to theconclusion that the 20 specificity of the OAS enzymes arisesfrom a different orientation of the ribose of the acceptorsubstrate in such a way that the 20 hydroxyl, not the 30 hy-droxyl, is posed for catalysis (Torralba and others 2008).
A search for the 2-5A binding protein, RNase L, only gavepositive results in tetrapods, leading to the assumption thatthe OAS system only exists in tetrapods (Cayley and others1982). This might still hold true for the classical OAS/RNaseL pathway. However, it is now clear that the OAS proteinsare part of an ancient family of template-independent poly-merases. With the progress in genomic sequencing, homo-logs of OAS have been identified throughout a number ofanimal phyla (Torralba and others 2008; Kjaer and others2009), including sponges, which are some of the oldestmetazoans (Wiens and others 1999). At the same time, it ap-pears that OAS has been secondarily lost in certain animals, as
THE OLIGOADENYLATE SYNTHETASE FAMILY 43
it is absent from the genomes of Caenorhabditis elegans andDrosophila melanogaster. In vertebrates, members of the OASfamily have been identified in all classes, except from bonyfishes (Osteichthyes). On the basis of the evidence presentedabove, we presume that the 20-specific OAS arose at the be-ginning of metazoan evolution (Torralba and others 2008).
Detection of Viral Infection: Binding of dsRNAand Activation of the OAS Enzymatic Activity
The activation of OAS1 by dsRNA and certain single-stranded RNAs has been characterized extensively in vitro.There is a tendency toward better activation of OAS1 bylonger dsRNAs (Desai and Sen 1997). This requirement forlonger dsRNA is somewhat enigmatic, as the RNA bindinggroove on OAS only covers 18 to 20 base pairs. It was sug-gested that the active form of OAS1 is tetrameric (Marie andothers 1990; Ghosh and others 1997), which could explain thepreference of OAS1 for longer dsRNAs. However, evidencefrom both our own work and that of others clearly demon-strated that monomeric OAS1 is fully active (Wathelet andothers 1986; Hartmann and others 2001, 2003). Thus, at leastfor OAS1, multimerization cannot explain the preference forlonger dsRNA. Contrary to this, relatively strong data havebeen generated to support a dimeric model for activatedOAS2 (Sarkar and others 1999a, 2002).
The mechanistic details of RNA activation have primarilybeen studied using OAS1, and the discussion in this para-graph refers to data obtained using OAS1. At present,we believe that the basic mechanism is similar for bothOAS2 and OAS3. The RNA binding site of OAS has beenidentified as a positively charged groove, found at theinterface of the N- and C- terminal domains (Hartmann andothers 2003). Single charge-swap mutations (Lys/Arg toGlu) within this area prevent the protein from entering theactivated state but have limited effect on RNA binding.However, multiple mutations within the basic RNA bindinggrove have an additive effect upon RNA binding. Thesedata lead to the conclusion that the activation of OAS is a2-step process: OAS binds RNA, and if the RNA is a suit-able activator, the protein will undergo a conformationalchange, involving, but not limited to, a rotation of theN- and C-terminal domains relative to each other, assem-bling the active site (Hartmann and others 1998a, 2003).Binding of dsRNA is not equal to activation, as the OASprotein can bind to a number of single-stranded RNAs,which does not lead to activation. Our current under-standing of the molecular details involved in OAS recog-nition of, and activation by, dsRNA is poor, in particulardue to the lack of structural information on the activatedOAS-dsRNA complex.
It is presumed that, in vivo, OAS recognizes viral dsRNA,which will activate OAS and initiate RNA degradation withinthe infected cell via the RNase L pathway. Thus, OAS servesas a combined pattern recognition receptor and antiviralprotein. However, few in vivo data exist to aid in the identi-fication of what kind of viral RNA is recognized by OAS.
RNase L–Dependent and RNase L–IndependentAntiviral Effects of the OAS Family Members
A number of studies have been performed over the years,documenting the important role played by the members of
the OAS family in controlling viral replication within mam-malian cells. Unfortunately, space does not permit us to gothrough all these studies systematically, but a recent reviewcan be found in (Silverman 2007). Here, we will focus uponthe different fundamental mechanisms by which the OASproteins can exert antiviral activity.
The first direct demonstration of an antiviral effect by amember of the OAS family was performed by Judith Che-bath and others (1987). They measured the replication ofEncephalomyocarditis virus (EMCV, a picornavirus) in cellstransfected with a plasmid overexpressing human OAS1,and compared it to cells transfected with the empty vector.Expression of OAS1 led to 10- to 100-fold reduction of viraltiters. Similar experiments were carried out for human OAS2with similar results (Marie and others 1999; Ghosh andothers 2000). In both cases, the authors also tested a differentvirus, Vesicular Stomatitis virus, (VSV a rhabdovirus) andfound no effect of OAS expression on the replication of thisvirus. Recent experiments with cells overexpressing OAS3reduced the viral titers of alphaviruses such as Chikungunyavirus, Sindbis virus, and Semliki Forest virus (Brehin andothers 2009).
Beside the demonstrated antiviral effects, polymorphismsin the human OAS gene have been also shown to be asso-ciated with susceptibility to infection with West Nile Virus(WNV, a flavivirus) (Yakub and others 2005; Lim and others2009) and Hepatitis C virus (a flavivirus) (Knapp and others2003) as well as SARS coronavirus (a coronavirus) (Hamanoand others 2005; He and others 2006).
RNase L–dependent antiviral effectof the OAS family
The classical OAS/RNase L pathway was described al-ready in the late 1970s (Roberts and others 1976; Hova-nessian and others 1977; Kerr and others 1977; Baglioni andothers 1978; Zilberstein and others 1978). Activation of thispathway results in RNA degradation within infected cells,both viral and cellular, including ribosomal RNA. Thus, ineffect, protein translation is inhibited in cells where theOAS/RNase L pathway is active, which effectively preventsviral replication. The activity of the OAS/RNase L pathwaycan be monitored by following degradation of ribosomalRNA in virus-infected cells or cells transfected with 2-5As.Generation of RNase L–deficient mice (RNase L�/�) con-firmed the importance of RNase L in the replication of pi-cornaviruses as these mice displayed an increased sensitivitytoward EMCV infection (Zhou and others 1997).
A recent study investigated the role of the OAS familyagainst dengue virus (a flavivirus) and found that both OAS1and OAS3 possess antiviral activity against dengue virus inan RNase L–dependent manner (Lin and others 2009).
RNase L–independent antiviral effectof the OAS family
Recently, evidence has been accumulating about at least 1,and probably several RNase L–independent pathways, bywhich members of the OAS family can prevent viral repli-cation. The human OASL and murine Oas1b are both devoidof enzymatic activity and thus incapable of activating RNaseL. Nevertheless, both proteins display antiviral activity ifexpressed in mammalian cells.
44 KRISTIANSEN ET AL.
The murine Oas1b gene. Mice have multiple duplicationsof their Oas1 gene, leading to 8 Oas1 genes in mice. In 2002, 2groups simultaneously identified 1 of the 8 genes, Oas1b, asthe gene responsible for the relative resistance of wild micetoward infection with WNV (Mashimo and others 2002;Perelygin and others 2002). All inbred mouse strains, except1, were susceptible to infection with WNV, whereas all of thestrains derived from wild mice were resistant to infection.Contrary to wild mice, inbred mice contained an Oas1b genewith a premature stop codon, hence encoding a truncatedversion of Oas1b, lacking *30% of its C-terminal domain.This truncated protein is not expected to fold properly, and isexpected to be unstable within the cell. The resistant mice allharbored a full-length version of Oas1b. Additional experi-ments confirmed the importance of the Oas1b gene in resis-tance to WNV. Knock-in of the full-length Oas1b from aresistant strain results in the reversal of the flavivirus-susceptible phenotype (Scherbik and others 2007). Further,expression of Oas1b protein suppresses WNV replication incell culture (Kajaste-Rudnitski and others 2006).
So why is this listed under the RNase L–independent ef-fect? First, the inability of any of the other 7 genes to com-pensate suggests a different mechanism. Second, mutationswithin the active site of the Oas1b protein are incompatiblewith enzymatic activity (Hartmann and others 2003). Usingcell lines with inducible expression of Oas1b, we demon-strated that the suppression of WNV was restricted tothe early phases of infection, and knock-out of RNase L bysiRNA does not affect the inhibition of WNV replicationby Oas1b, clearly indicating an RNase L–independentmechanism (Kajaste-Rudnitski and others 2006).
The OASL gene. OASL is a unique member of the OASfamily that lacks the enzymatic activity but has a C-terminaldomain, which displays homology to ubiquitin. OASL dis-plays antiviral activity against EMCV in a manner requiring
the ubiquitine-like domain, but is independent of RNase L(Marques and others 2008). The ubiqutin-like domain ofOASL does not appear to function as an ubiqutin molecule,as we were unable to detect any conjugation. Further, theubiqutin-like domain does not have any effect on the stabilityof the OASL protein (Hartmann, unpublished). The OASLgene appears to be conserved in mammals, with some spe-cies harboring 2 genes of the OASL type. This is best studiedin mice, where both genes have been characterized. ThemOasl1 gene is the ortholog of the human OASL gene andenzymatically inactive (Eskildsen and others 2003). ThemOasl2 gene is enzymatically active, which is reflected in thesequence of their active sites. Further, the mOasl1 and mOasl2genes differ substantially in the sequence of their ubiqutin-like domains: the mOasl1 gene is closer to its human orthologthan to its murine paralog. In humans, the OASL2 gene is apseudogene. We believe that the mOasl2 gene represents anevolutionary intermediate between the active OAS1/mOas1genes and the inactive OASL/mOasl1 genes.
Concluding Remarks
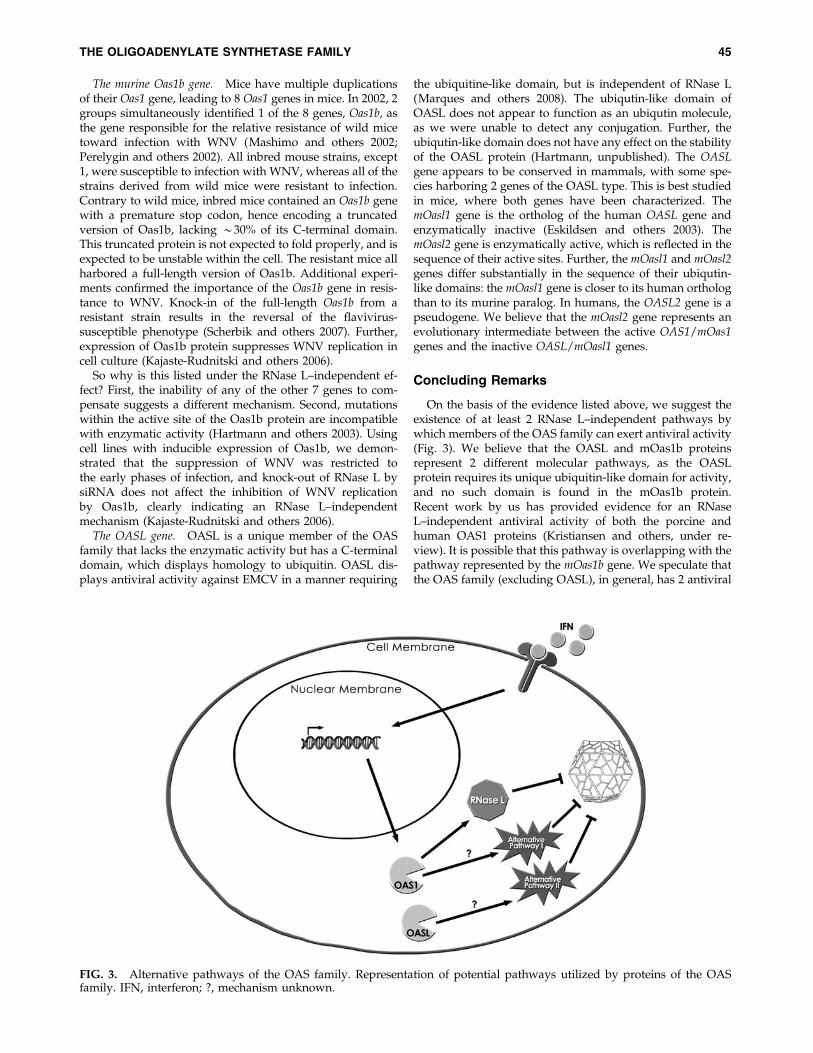
On the basis of the evidence listed above, we suggest theexistence of at least 2 RNase L–independent pathways bywhich members of the OAS family can exert antiviral activity(Fig. 3). We believe that the OASL and mOas1b proteinsrepresent 2 different molecular pathways, as the OASLprotein requires its unique ubiquitin-like domain for activity,and no such domain is found in the mOas1b protein.Recent work by us has provided evidence for an RNaseL–independent antiviral activity of both the porcine andhuman OAS1 proteins (Kristiansen and others, under re-view). It is possible that this pathway is overlapping with thepathway represented by the mOas1b gene. We speculate thatthe OAS family (excluding OASL), in general, has 2 antiviral
FIG. 3. Alternative pathways of the OAS family. Representation of potential pathways utilized by proteins of the OASfamily. IFN, interferon; ?, mechanism unknown.
THE OLIGOADENYLATE SYNTHETASE FAMILY 45
pathways, the classical RNase L–dependent pathway and ayet uncharacterized RNase L–independent pathway, whichcan act independently of each other. The mOas1b gene is theresult of a recent duplication event that has so far only beenidentified in the muroidea (mice and rats), and it is possi-ble that the mOas1b gene has specialized in the RNase L–independent pathway. However, this is purely speculativeand experiential evidence, in particular a clarification of themolecular details of the novel RNase L–independent path-way, is required.
Acknowledgments
Research in the author’s laboratory was supported bygrants from the Danish Cancer Society, The Danish MedicalResearch Council (Grant no. 22-04-0704), Novo NordiskFoundation, and the Carlsberg Foundation.
Author Disclosure Statement
No financial competing interests exist.
References
Baglioni C, Minks MA, Maroney PA. 1978. Interferon action maybe mediated by activation of a nuclease by pppA2’p5’A2’-p5’A. Nature 273:684–687.
Beese LS, Steitz TA. 1991. Structural basis for the 3’-5’ exonu-clease activity of Escherichia coli DNA polymerase I: a twometal ion mechanism. EMBO J 10:25–33.
Benech P, Mory Y, Revel M, Chebath J. 1985. Structure of twoforms of the interferon-induced (2’-5’) oligo A synthetase ofhuman cells based on cDNAs and gene sequences. EMBO J4:2249–2256.
Brehin AC, Casademont I, Frenkiel MP, Julier C, Sakuntabhai A,Despres P. 2009. The large form of human 2’,5’-OligoadenylateSynthetase (OAS3) exerts antiviral effect against Chikungunyavirus. Virology 384:216–222.
Cayley PJ, White RF, Antoniw JF, Walesby NJ, Kerr IM. 1982.Distribution of the ppp(A2’p)nA-binding protein and inter-feron-related enzymes in animals, plants, and lower organ-isms. Biochem Biophys Res Commun 108:1243–1250.
Chebath J, Benech P, Revel M, Vigneron M. 1987. Constitutiveexpression of (2’-5’) oligo A synthetase confers resistance topicornavirus infection. Nature 330:587–588.
Cho HD, Verlinde CL, Weiner AM. 2005. Archaeal CCA-addingenzymes: central role of a highly conserved beta-turn motif inRNA polymerization without translocation. J Biol Chem280:9555–9566.
Clemens MJ, Williams BR. 1978. Inhibition of cell-free proteinsynthesis by pppA2’p5’A2’p5’A: a novel oligonucleotidesynthesized by interferon-treated L cell extracts. Cell 13:565–572.
Delarue M, Poch O, Tordo N, Moras D, Argos P. 1990. An at-tempt to unify the structure of polymerases. Protein Eng3:461–467.
Desai SY, Sen GC. 1997. Effects of varying lengths of double-stranded RNA on binding and activation of 2’-5’-oligoadenylatesynthetase. J Interferon Cytokine Res 17:531–536.
Dong B, Silverman RH. 1997. A bipartite model of 2-5A-dependent RNase L. J Biol Chem 272:22236–22242.
Eskildsen S, Hartmann R, Kjeldgaard NO, Justesen J. 2002. Genestructure of the murine 2’-5’-oligoadenylate synthetase family.Cell Mol Life Sci 59:1212–1222.
Eskildsen S, Justesen J, Schierup MH, Hartmann R. 2003. Char-acterization of the 2’-5’-oligoadenylate synthetase ubiquitin-like family. Nucleic Acids Res 31:3166–3173.
Ghosh A, Sarkar SN, Guo W, Bandyopadhyay S, Sen GC. 1997.Enzymatic activity of 2’-5’-oligoadenylate synthetase is im-paired by specific mutations that affect oligomerization of theprotein. J Biol Chem 272:33220–33226.
Ghosh A, Sarkar SN, Sen GC. 2000. Cell growth regulatory andantiviral effects of the P69 isozyme of 2–5 (A) synthetase.Virology 266:319–328.
Hamano E, Hijikata M, Itoyama S, Quy T, Phi NC, Long HT, HaLD, Ban VV, Matsushita I, Yanai H, and others. 2005. Poly-morphisms of interferon-inducible genes OAS-1 and MxAassociated with SARS in the Vietnamese population. BiochemBiophys Res Commun 329:1234–1239.
Hartmann R, Justesen J, Sarkar SN, Sen GC, Yee VC. 2003. Crystalstructure of the 2’-specific and double-stranded RNA-activatedinterferon-induced antiviral protein 2’-5’-oligoadenylate syn-thetase. Mol Cell 12:1173–1185.
Hartmann R, Norby PL, Martensen PM, Jorgensen P, James MC,Jacobsen C, Moestrup SK, Clemens MJ, Justesen J. 1998a.Activation of 2’-5’ oligoadenylate synthetase by single-stran-ded and double-stranded RNA aptamers. J Biol Chem273:3236–3246.
Hartmann R, Olsen HS, Widder S, Jorgensen R, Justesen J. 1998b.p59OASL, a 2’-5’ oligoadenylate synthetase like protein: anovel human gene related to the 2’-5’ oligoadenylate synthe-tase family. Nucleic Acids Res 26:4121–4128.
Hartmann R, Walko G, Justesen J. 2001. Inhibition of 2’-5’ oligoa-denylate synthetase by divalent metal ions. FEBS Lett 507:54–58.
He J, Feng D, de Vlas SJ, Wang H, Fontanet A, Zhang P, Plan-coulaine S, Tang F, Zhan L, Yang H, and others. 2006. Asso-ciation of SARS susceptibility with single nucleic acidpolymorphisms of OAS1 and MxA genes: a case-controlstudy. BMC Infect Dis 6:106.
Holm L, Sander C. 1995. DNA polymerase beta belongs to anancient nucleotidyltransferase superfamily. Trends BiochemSci 20:345–347.
Hovnanian A, Rebouillat D, Mattei MG, Levy ER, Marie I, MonacoAP, Hovanessian AG. 1998. The human 2’,5’-oligoadenylatesynthetase locus is composed of three distinct genes clusteredon chromosome 12q24.2 encoding the 100-, 69-, and 40-kDaforms. Genomics 52:267–277.
Hovanessian AG, Brown RE, Kerr IM. 1977. Synthesis of lowmolecular weight inhibitor of protein synthesis with enzymefrom interferon-treated cells. Nature 268:537–540.
Justesen J, Hartmann R, Kjeldgaard NO. 2000. Gene structureand function of the 2’-5’-oligoadenylate synthetase family.Cell Mol Life Sci 57:1593–1612.
Kajaste-Rudnitski A, Mashimo T, Frenkiel MP, Guenet JL, LucasM, Despres P. 2006. The 2’,5’-oligoadenylate synthetase 1b is apotent inhibitor of West Nile virus replication inside infectedcells. J Biol Chem 281:4624–4637.
Kakuta S, Shibata S, Iwakura Y. 2002. Genomic structure of themouse 2’,5’-oligoadenylate synthetase gene family. J Inter-feron Cytokine Res 22:981–993.
Kerr IM, Brown RE, Hovanessian AG. 1977. Nature of inhibitorof cell-free protein synthesis formed in response to interferonand double-stranded RNA. Nature 268:540–542.
Kjaer KH, Poulsen JB, Reintamm T, Saby E, Martensen PM,Kelve M, Justesen J. 2009. Evolution of the 2’-5’-oligoadenylate
46 KRISTIANSEN ET AL.
synthetase family in eukaryotes and bacteria. J Mol Evol 69:612–624.
Knapp S, Yee LJ, Frodsham AJ, Hennig BJ, Hellier S, Zhang L,Wright M, Chiaramonte M, Graves M, Thomas HC, andothers. 2003. Polymorphisms in interferon-induced genes andthe outcome of hepatitis C virus infection: roles of MxA, OAS-1 and PKR. Genes Immun 4:411–419.
Lim JK, Lisco A, McDermott DH, Huynh L, Ward JM, Johnson B,Johnson H, Pape J, Foster GA, Krysztof D, and others. 2009.Genetic variation in OAS1 is a risk factor for initial infectionwith West Nile virus in man. PLoS Pathog 5:e1000321.
Lin RJ, Yu HP, Chang BL, Tang WC, Liao CL, Lin YL. 2009.Distinct antiviral roles for human 2’,5’-oligoadenylate syn-thetase family members against dengue virus infection. J Im-munol 183:8035–8043.
Marie I, Hovanessian AG. 1992. The 69-kDa 2-5A synthetase iscomposed of two homologous and adjacent functional do-mains. J Biol Chem 267:9933–9939.
Marie I, Rebouillat D, Hovanessian AG. 1999. The expression ofboth domains of the 69/71 kDa 2’,5’ oligoadenylate synthetasegenerates a catalytically active enzyme and mediates an anti-viral response. Eur J Biochem 262:155–165.
Marie I, Svab J, Robert N, Galabru J, Hovanessian AG. 1990.Differential expression and distinct structure of 69- and 100-kDa forms of 2-5A synthetase in human cells treated withinterferon. J Biol Chem 265:18601–18607.
Marques J, Anwar J, Eskildsen-Larsen S, Rebouillat D, PaludanSR, Sen G, Williams BR, Hartmann R. 2008. The p59 oligoa-denylate synthetase-like protein possesses antiviral activitythat requires the C-terminal ubiquitin-like domain. J Gen Virol89:2767–2772.
Martin G, Jeno P, Keller W. 1999. Mapping of ATP binding re-gions in poly(A) polymerases by photoaffinity labeling and bymutational analysis identifies a domain conserved in manynucleotidyltransferases. Protein Sci 8:2380–2391.
Martin G, Keller W. 2007. RNA-specific ribonucleotidyl trans-ferases. RNA 13:1834–1849.
Mashimo T, Lucas M, Simon-Chazottes D, Frenkiel MP, Mon-tagutelli X, Ceccaldi PE, Deubel V, Guenet JL, Despres P. 2002.A nonsense mutation in the gene encoding 2’-5’-oligoadenylatesynthetase/L1 isoform is associated with West Nile virussusceptibility in laboratory mice. Proc Natl Acad Sci USA99:11311–11316.
Melchjorsen J, Kristiansen H, Christiansen R, Rintahaka J,Matikainen S, Paludan SR, Hartmann R. 2009. Differentialregulation of the OASL and OAS1 genes in response to viralinfections. J Interferon Cytokine Res 29:199–207.
Perelygin AA, Scherbik SV, Zhulin IB, Stockman BM, Li Y,Brinton MA. 2002. Positional cloning of the murine flavivirusresistance gene. Proc Natl Acad Sci USA 99:9322–9327.
Perelygin AA, Zharkikh AA, Scherbik SV, Brinton MA. 2006.The mammalian 2’-5’ oligoadenylate synthetase gene family:evidence for concerted evolution of paralogous Oas1 genes inRodentia and Artiodactyla. J Mol Evol 63:562–576.
Rebouillat D, Hovnanian A, Marie I, Hovanessian AG. 1999. The100-kDa 2’,5’-oligoadenylate synthetase catalyzing preferen-tially the synthesis of dimeric pppA2’p5’A molecules iscomposed of three homologous domains. J Biol Chem 274:1557–1565.
Rebouillat D, Marie I, Hovanessian AG. 1998. Molecular cloningand characterization of two related and interferon-induced 56-kDa and 30-kDa proteins highly similar to 2’-5’ oligoadenylatesynthetase. Eur J Biochem 257:319–330.
Roberts WK, Hovanessian A, Brown RE, Clemens MJ, Kerr IM.1976. Interferon-mediated protein kinase and low-molecular-weight inhibitor of protein synthesis. Nature 264:477–480.
Sarkar SN, Bandyopadhyay S, Ghosh A, Sen GC. 1999a. Enzy-matic characteristics of recombinant medium isozyme of 2’-5’oligoadenylate synthetase. J Biol Chem 274:1848–1855.
Sarkar SN, Ghosh A, Wang HW, Sung SS, Sen GC. 1999b. Thenature of the catalytic domain of 2’-5’-oligoadenylate synthe-tases. J Biol Chem 274:25535–25542.
Sarkar SN, Pal S, Sen GC. 2002. Crisscross enzymatic reactionbetween the two molecules in the active dimeric P69 form ofthe 2’-5’ oligodenylate synthetase. J Biol Chem 277:44760–44764.
Scherbik SV, Kluetzman K, Perelygin AA, Brinton MA. 2007.Knock-in of the Oas1b(r) allele into a flavivirus-induced dis-ease susceptible mouse generates the resistant phenotype.Virology 368:232–237.
Silverman RH. 2007. Viral encounters with 2’,5’-oligoadenylatesynthetase and RNase L during the interferon antiviral re-sponse. J Virol 81:12720–12729.
Steitz TA, Smerdon SJ, Jager J, Joyce CM. 1994. A unified poly-merase mechanism for nonhomologous DNA and RNApolymerases. Science 266:2022–2025.
Torralba S, Sojat J, Hartmann R. 2008. 2’-5’ oligoadenylate syn-thetase shares active site architecture with the archaeal CCA-adding enzyme. Cell Mol Life Sci 65:2613–2620.
Wathelet M, Moutschen S, Cravador A, DeWit L, Defilippi P,Huez G, Content J. 1986. Full-length sequence and expressionof the 42 kDa 2-5A synthetase induced by human interferon.FEBS Lett 196:113–120.
Wiens M, Kuusksalu A, Kelve M, Muller WE. 1999. Origin of theinterferon-inducible (2’-5’)oligoadenylate synthetases: cloningof the (2’-5’)oligoadenylate synthetase from the marinesponge Geodia cydonium. FEBS Lett 462:12–18.
Yakub I, Lillibridge KM, Moran A, Gonzalez OY, Belmont J,Gibbs RA, Tweardy DJ. 2005. Single nucleotide polymor-phisms in genes for 2’-5’-oligoadenylate synthetase and RN-ase L inpatients hospitalized with West Nile virus infection. JInfect Dis 192:1741–1748.
Zhou A, Paranjape J, Brown TL, Nie H, Naik S, Dong B, ChangA, Trapp B, Fairchild R, Colmenares C, and others. 1997. In-terferon action and apoptosis are defective in mice devoid of2’,5’-oligoadenylate-dependent RNase L. EMBO J 16:6355–6363.
Zilberstein A, Kimchi A, Schmidt A, Revel M. 1978. Isolation oftwo interferon-induced translational inhibitors: a protein ki-nase and an oligo-isoadenylate synthetase. Proc Natl Acad SciUSA 75:4734–4738.
Address correspondence to:Dr. Rune Hartmann
Centre for Structural BiologyDepartment of Molecular Biology