The role of abiotic factors, cultivation practices and soil fauna in the dispersal of genetically modified microorganisms in soils
John Dighton a,*, Helen E. Jones b, Clare H. Robinson c, John Beckett b a Diuision of Pinelands Research, Department of Biology, Rutgers Uni~,ersity, Camden, NJ 08102, USA
h The Institute of Terrestrial Ecology, Merlewood Research Station, Grange-over-Sands, Cumbria, LA11 6JU, UK c Department of Animal and Plant Science, Sheffield Centre for Arctic Ecology, Universi~ of Sheffield, Sheffield SIO 2UQ, UK
Accepted 5 October 1996
Abstract
Any study of the fate of genetically modified microorganisms (GMMs), released into the environment, must examine how far they can disperse from their release site. The likelihood of dispersal will depend on two key factors: 1 . . Speed and distance of dispersal. 2 . . Survival and/or reproduction, enabling them to reach new environments or hosts in a viable state.
This paper reviews factors influencing the speed and distance of dispersal of soil microbes. These will depend on soil management practices, soil physico-chemical properties, and on the activities and movement of members of the soil biota acting as vectors.
Models of viral and bacterial dispersal are evaluated. Their limited relevance to natural soils, with their inherent heterogeneity, and applicability to actinomycete and fungal dispersal is discussed. Cultivation practices cause physical disturbance of the soil by ploughing, tyning and hoeing. The movement of soil during these activities can disperse microorganisms within 20-30 cm of the plough depth and several metres horizontally. Ploughing and tyning increase porosity, but heavy traffic compacts the soil and destroys macropores. The former thus aid dispersal and the latter retards it. Plant roots can act as food for microorganisms, and their growth will disperse the microorganisms deeper into the soil. Dispersal may be further increased as root death creates new macropores. As dispersal models have been mainly developed from studies of homogenised soils, many of these factors have not been adequately incorporated in predictions of microbial dispersal.
Their are few studies directly pertaining to faunal movement of GMMs. This review highlights our current knowledge in identifying potential key faunal groups which may be influential in enhancing GMM transport in soil. The soil faunal community present a range of species diversity and function. The potential importance of the major soil faunal groups in dispersal of microorganisms is discussed. Larger, mobile animals, such as lumbricids are the main agents of vertical and horizontal dispersal of GMMs. Large epigeic fauna such as diplopods and isopods may be important in horizontal dispersal at the soil surface but information on their abundance in different ecosystems is limited. Geographic distribution of faunal groups inflences their importance at any one location. Generally, the more numerically abundant, but smaller-sized protozoa and nematodes play an insignificant part. The dispersal of microorganisms by soil fauna may be of greater importance than
110 J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131
abiotic factors in soils of high clay content or impeded drainage. Models based on artificial microcosms have shown the importance of soil particle size in downward movement of the drainage water which will carry the microbial propagules.
This review highlights two major areas where our understanding of interactions of soil structure, biotic components and microbial dispersal is lacking. Firstly, soil heterogeneity is not included as a factor in model systems of abiotic dispersal of microbes. This is a major drawback for their use as predictive tools in natural field soils. Secondly, our lack of knowledge of the interactions between many soil faunal taxa and the microbial community requires further attention.
There have been very few intentional releases of genetically modified microorganisms (GMMs) into the environment (Lindow and Panopoulos, 1988), so little is known of their ability to survive and persist under natural conditions. The key questions posed by Barnthouse and Palumbo (1985) and Tiedje et al. (1989) regarding the ability of a GMM to disperse to an independent geographical site or ecological niche are: • How likely is it to disperse away from the release
site? • How far and how rapidly will the transported
microbe move? • Can it survive long enough to reach target envi-
ronments or hosts? A released organism will only become of ecologi-
cal concern if it survives and reproduces. The sur- vival and functioning of released species, including GMMs, will depend on the competitive balance be- tween them and the indigenous species for limiting resources, and on the balance between reproduction and mortality (Taub, 1992). It has been stated that the addition of extra genes reduces competitiveness, due to the added cost of synthesising and maintain- ing additional nucleic acids and the production of proteins (eg Lenski and Nguyen, 1988). However, Hartl et al. (1983); Winstanley et al. (1989) and Marshall et al. (1988) showed that modified microor- ganisms may have unchanged or even enhanced survival in both the laboratory and the field, depend- ing on the nature of the recombinant DNA and its expression.
Altered genetic information may also persist by being transferred to native species that have a greater
capacity to survive and multiply than the released organism (Alexander, 1986). Genetic exchange by conjugation, transduction or transformation (Lenski, 1987; Miller, 1988) to the native population, may allow the persistence of a modified gene even after the death of the released organism. Although this is an important means of transfer of the information contained within the GMM, we have omitted discus- sion of it in this review.
The released organism will differ from the native population due to the addition or modification of one or few genes, in the source pool size and, because of extensive laboratory culture, reduced competitive ability. In other respects the effectiveness of the dispersal agent should be similar for both the native population and for the released organism. Three factors will be important in extensive dispersal and spread: 1. The size of the source pool. 2. The effectiveness of the dispersal agent. 3. The susceptibility of the GMM to the environ-
ment. We have therefore reviewed the role of soil fauna
and tillage practices in the dispersal of soil microor- ganisms, and have made no distinction between the native microflora and released species.
We compare the role of soil fauna in the dispersal of microorganisms with the role of abiotic factors such as soil type, water movement, and different tillage practices. Where interactions between faunal activity and changes in soil structure are significant, the effects have been identified. The influence of the rhizosphere may also be important, as this is a site of enhanced microbial and faunal activity. We have assessed the importance of physical movement in both saturated and unsaturated soils, but not the
J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131 I 11
movement by mass flow of water in groundwater courses and rivers and mass movement of soil by erosion (aeolean and water). We also analyse the role of the microfauna, mesofauna and invertebrate macrofauna. We have deliberately excluded verte- brates, even though in some ecosystems their under- ground activities may be significant and they may be involved in long distance transfer of microorganisms above ground. Differences in the dispersal potential of viruses, bacteria, actinomycetes and fungi have been examined as the review by Gammack et al. (1992) is restricted mainly to bacteria and agricul- tural soils.
2. The role of abiotic factors in microbial disper- sal
The most important factor in the abiotic dispersal of microorganisms is the flow of percolating soil water. Soil porosity depends on the ratio of the volume of void spaces to the total soil volume. Generally, the porosity of clay soils (small particle size) is larger than that of sandy soils (large particle size), but, because of the arrangement of particles, not all of the void space is available for water flow. Table 1 gives the range of sizes of soil particles, voids and organisms inhabiting the soil. There is
Table 1
Sizes of soil structures, soil microorganisms and soil fauna (after Metting, 1992; Goodey , 1951; Evans et al., 1961; Wal lwork , 1976 and Coleman, 1994)
Collembola Surface/litter inhabitants 3 - 4 m m length ca 2.4 m m 3
Organic fractions 1 - 3 m m ca 0.4 m m 3
Soil dwellers 1 m m length ca 2 X 10 -4 m m 3
Acari Mesostigmata 0 . 2 - 2 m m length 4 x 1 0 - 3 - 4 m m 3
Cryptostigmata 0 . 2 - 1 . 5 m m length 4 x 1 0 - 3_ 1.8 m m 3
Astigmata 0 . 2 - 1 . 2 m m length 5 X 1 0 - 4 - 0 . 9 m m 3
Prostigmata 0 .1-1 .6 nun length 5 X 1 0 - 4 - 2 . 0 m m 3
Enchytraeidae ca 1 m m 1 0 - 2 0 m m 2
Lumbricidae 2 - 1 0 m m Variable on length ( > 10 m m )
112 J. Dighton et al. / Applied Soil Ecology 5 (1997) 109-131
greater potential for microbial movement in coarse than fine soils because of the larger pore spaces (Bitton et al., 1974).
Surface chemistry of soil particles and soil water chemistry influence surface charges on soil particles and adherence of microorganisms to them. Organic matter and clay surfaces both posess large, nega- tively charged, surface areas and increase microbial adsorption (Marshall, 1971). The effective diameter of the microorganism in relation to the pore neck radius may also affect its ability to move. The rate of passage of a microorganism through soil may be effected either as a Brownian movement or its own motility (Griffin and Quail, 1968).
Void space and rate of water flow will vary with soil type and tillage practices. Soil aggregates devel- oping from microbial activity (Lynch and Elliott, 1983) also influences physical characteristics of the soil, modifying the water flow and, hence, microbial movement.
To summarise, the influence of soil type on mi- crobial transport will be due both to its physical structure (relative proportion of sand, silt, clay and organic matter) and to variation in adsorptive capac- ity of its colloidal material. The adsorptive properties are thought to be important for viral retention, whereas for bacteria, physical properties are also significant (Goyal and Gerba, 1979).
Using physico-chemical properties of soils, mod- els have been developed to describe microbial flux in soil. Gerba et al. (1991) considered that five key factors determine the physical transport of microor- ganisms through soil: 1. Advection - - the process of transport in bulk
motion of flowing water with carriage at a rate equal to the average velocity of the water flow.
2. D i s p e r s i o n - - spreading of the organisms as they pass through the sub-surface medium. This is a function of the variation in actual pore water velocity compared with the average pore water velocity, and the effect of Brownian movement.
3. Adsorption - - adhesion to soil particles. 4 . F i l t r a t i o n - - removal of organisms from the
pore water by size exclusion. 5. Decay or die-off - - inactivation by environmen-
tal stress, such as temperature or lack of nutrients. Using these parameters, Dickinson (1991) devel-
oped a hydrodynamic advection/dispersion equation
to describe the general movement of organisms in both saturated and unsaturated groundwater flow.
Flux of microorganisms through soil is, however, reduced if the effects of adsorption and retardation are taken into account (Gerba et al., 1991). They stated that, at flow rates of less than 1 m h -1, the adsorption equilibria of viruses and bacteria are es- tablished within 1 to 24 h. Using stepwise additions of advection, dispersion, adsorption and decay into their model, Yates and Yates (1991) demonstrated that interactions of these factors led to fewer organ- isms being transported and over a shorter distance.
A hydrodynamic advection-dispersion equation was used to develop two models (Van Genuchten, 1978; Van Genuchten, 1986) called Sumatra-I and WORM, the former of which is more applicable for modelling microorganism transport in unsaturated porous media, such as soils (Dickinson, 1991). Yates and Ouyang (1992) elaborated dispersal models which included expressions for water and heat trans- port through soils and a virus transport model which they combined in a computer model called VIRTUS. The component of virus flow was a function of conduction in solid, aqueous, and air and water vapour components, which influenced the survival of virions. The model gave good predictions for both saturated and unsaturated soils.
A limitation of most of these models, however, is that they are based on parameters measured in recon- stituted soil cores. The soils are usually dried and sieved and then repacked to a uniform density (Lance et al., 1976; Duboise et al., 1976; Funderburg et al., 1979; Landry et al., 1979; Wang et al., 1981; Powet- son et al., 1990). Thus, many of the heterogenous physical characteristics of field soils, (e.g. the pres- ence of macropores, soil aggregates, earthworm bur- rows, plant root channels, cracks due to soil drying etc) are deliberately removed. It is likely that these variables will be the key ones in causing variation in the extent of dispersal.
2.1. Viruses
Provided viral particles can withstand the physical constraints of the soil environment, they will remain there unaltered. The physical stability, equivalent to decay or die-off in the key factors considered by
J. Dighton et al. /Appl ied Soil Ecology 5 (1997) 109-131 I 13
Gerba et al. (1991), of GMM baculoviruses (to be used as insecticides) has been tested by Bishop et al. (1988). These modified viruses were inoculated into sterile soil collected from the field and virus particles were monitored by plaque assays over 2 weeks. Viral infectivity declined by two orders of magnitude within the first 3 days which was thought to be a factor of the genetic manipulation. Genetic modifica- tion of the viruses appears to reduce their ability to withstand the physical constraints of the environ- ment. Dispersion was not measured.
Despite the limitations of the studies involving reconstituted cores, certain conclusions can be drawn from them. Iionic strength and chemical composition of the soil solution appears to be a key factor. Elution of a soil column with deionised water caused dispersal of adsorbed viral particles at least 80 cm down the column, whereas elution with 0.01 N cal- cium chloride did not (Lance et al., 1976). Also there was less elution with applied sewage effluent than with rainwater. Thus, release of adsorbed viral parti- cles may he more likely in soils of low fertility, where soil solution strength is low. Arable soils, where deliberate release of GMMs may be most likely to occur, are usually fertile, i.e. the conditions in which the adsorbed viral particles would be more likely to remain in situ. Wang et al. (1981) showed that greater release of viral particles occurs at low pH. Seasonal and spatial rainfall patterns are impor- tant as high rates of flow cause high rates of viral removal. A critical volume of water is necessary for appreciable movement of bacteria and viral particles to occur (Griffin and Quail, 1968). Generally, exper- imental treatments tend to be more extreme than natural variation of rainfall events and chemistry of soil solution, so the findings would need to be confirmed by treatments more closely related to con- ditions occurring in the field.
In a study comparing virus movement in intact versus reconstituted soil cores, Landry et al. (1979) showed that retention was much greater in the natu- ral soil. It appeared that the disturbance caused by drying and sieving had a greater effect in increasing viral dispersal than the natural variation in the field. This may have implications in relation to tillage treatments in arable soils.
Experimental findings from microorganisms and soil cores might have more relevance to field prob-
lems if conditions in them more closely resembled those found in the natural soil. Intact soil cores would seem to be a preferable medium to reconsti- tuted ones. The interaction between soil structure and faunal activity should also be considered in terms of the animals' burrowing activity and their role in forming aggregates. Concern with field testing should not focus solely on GMMs though, as organisms should be considered in respect of their novelty to the environment into which they are released rather than because of the means by which they have become new (Miller et al., 1987).
2.2. B a c t e r i a
Abiotic movement of bacteria has recently been reviewed by Murphy and Tate (1996) and Wiebers et al. (1995), who conclude that existing models cannot be universally applied due to differences in soil properties between soils and within soils depending on management. Bacterial movement is restricted by soil pore size (Liu, 1982; Patni et al., 1984; Breiten- beck et al., 1988). Bacteria also play their own part in soil formation by secreting mucilage which aids soil aggregation (Vandevivere and Baveye, 1992). The role of soil aggregates is difficult to quantify: they may assist dispersal by allowing greater free space between the aggregates, or they may hinder if the bacteria become trapped inside. Soil pore size is a critical factor, as there is little bacterial dispersal below a pore neck radius of 1.5 ~m (Griffin and Quail, 1968) for bacterial cells of 0.5 /~m diameter. Scott Smith et al. (1983) showed in intact soil cores that nearly four times as many bacteria were eluted from a silt loam containing large vertical cracks as from fine crumb silt loam without cracks. Thus pore size distribution and the pattern of pore connectivity will both affect bacterial movement.
It may be necessary to distinguish between verti- cal and horizontal dispersal, and of necessity, soil cores used to simulate dispersal will be restricted to the study of vertical movement. Once percolating drainage water reaches zones of groundwater move- ment, the organisms it is carrying may be moved large horizontal distances. However, that depth may be below the zone where soil fauna are found, except perhaps for some of the deeper burrowing lumbri- cids. Percolating rainwater will affect downward dis-
114 J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131
persal, to the extent that Trevors et al. (1990) showed negligible movement of Pseudomonasfluorescens in soil columns in the absence of downward water flow, but bacterial cells were present throughout the col- umn and in the leachate, when water was added. The soil water content itself (matric potential) is an im- portant factor influencing bacterial movement (Postma et al., 1989; Murphy and Tate, 1996). Postma et al. (1989) inoculated Rhizobium leguminosum cells into soil aggregates and, as the water content at the time of inoculation increased, so the number of bacterial cells in aggregates decreased. They ex- plained that the water in the soil pores was prevent- ing bacterial penetration.
Horizontal dispersal in the upper layers is more likely to be brought about by the soil fauna than water flux, except in areas of sloping ground. Rahe et al. (1978) showed that drainage water movement down-slope enabled inoculated antibiotic resistant Escherichia coli to be horizontally dispersed 20 m in 12 h, far in excess of any rate of movement that would be possible by biotic means. The poorly- drained soil in which this occurred might be envis- aged as having two layers, a deeper saturated zone into which the bacteria-laden water could not flow, and a surface layer through which bacterial dispersal was possible because of the slope. Topography, therefore, was the critical factor, so that GMMs released into poorly-drained fiat ground may have little opportunity for dispersal, but a slope of only 10% is sufficient to greatly increase the risk. Mawds- ley et al. (1995) considered that lateral movement is a particular problem in soils with impermeable sub- strata.
Predictions of bacterial movement in soil have often been made with reconstituted soil cores, though movement in these has sometimes been compared with intact ones (e.g. Scott Smith et al., 1983). The importance of retaining structure of the natural soil in microcosm studies was recognised by Bentjen et al. (1989), who evaluated their relevance in the transport and survival of genetically modified Azospirillum lipoferum. Intact cores of a silt loam soil were planted with wheat and maize and inocu- lated with A. lipoferum suspension at 9 × 108 colony forming units (CFU) ml -~. After 9 weeks rhizo- sphere populations varied from undetectable to 10 6
CFU g-1. Replicate variability was explained as
inherent variation in the intact soil cores, and was therefore representative of field complexity. No GMMs were detected in the leachate, xylem exudate or sap sucking insects that were introduced in one treatment. Earthworms were inadvertently introduced into a few of the microcosms and these contained none of the GMMs either. In addition there was no niche displacement of rhizosphere populations of nitrifying bacteria. They concluded that intact soil core microcosms were a useful simulation as they included many features of ecosystem complexity, were easy to replicate and could be readily manipu- lated to introduce treatment effects.
Chemical properties of soil solution are also im- portant in bacterial movement. Twice as many bacte- ria were retained in microcosms of the same soil adjusted to pH 3.3 instead of pH 6.6 (Bitton et al., 1974), and alteration of the ionic strength of the added solution with calcium chloride also resulted in more retention (Tan et al., 1991). Kemp et al. (1992) showed significantly fewer genetically modified Pseudomonas fluorescens cells in the leachate from soil at pH 4.5 than at 7.5. These factors may have implications with regard to soil fertility, inorganic fertilisation treatment and, possibly, atmospheric pol- lution. The first two of these are likely to be particu- larly important variables in arable soils where GMM release is most likely.
Drahos et al. (1988) showed very limited move- ment of a modified root-colonising Pseudomonas aureofaciens, inoculated into the rhizosphere of wheat on a 0.4 ha plot in South Carolina. After 10 weeks, the maximum recorded distance moved by the released GMM was 18 cm (equivalent to 2-3 mm day- l ) . This maximum distance moved was only recorded in one sample out of 96, and no GMMs were isolated from the non-planted border areas, or drainage water. They concluded that the pattern of survival of the GMMs in the field was predicted very well by the pre-release microcosm tests, that GMMs did not out-compete native micro- bial populations and remained largely confined to their place of introduction. Amarger et al. (1991) also used field trials to assess the risks in releasing modified Rhizobium leguminosarum inoculated into peas. Bacteria were detected after several weeks at a distance of 15 cm, probably due to a spread along the roots. In the same study, R. cicer was used in
J. Dighton et al . /Applied Soil Ecology 5 (1997) 109-131 l 15
field experiments at Rothamsted with chickpea. After one year it had moved 60 cm, possibly in surface water after heavy winter rain. Mechanical soil culti- vation also dispersed it throughout the experimental plot. Where lucerne or cereals were grown subse- quently, the modified R. leguminosarum was found at a depth of 80 cm after 3 years, but not in plots planted with the shallower-rooting peas or beans. Ploughing had spread a very few colonies up to 2 m from the original perimeter (P. Hirsch, personal com- munication).
There are few studies of the direct effects of tillage on bacterial activity and dispersal, but they generally show that the accompanying disturbance leads to increased bacterial activity except where compaction is caused by heavy agricultural machin- ery. Deep cultivation increased bacterial numbers in soil (Franz, 1976), with a persistent effect still appar- ent 13 years later. Donegan et al. (1991) showed that tillage resulted in significantly higher numbers of Erwinia herbicola compared with an untilled treat- ment and the effect lasted for several weeks. Soil composition also influences water infiltration rate and is affected by heavy machinery (Meek et al., 1992). Root growth alleviates compaction as natural channels are formed by the death of tap roots. Living roots may also assist bacterial dispersal. Parke et al. (1986) showed that when Pseudomonas fluorescens was inoculated on wheat seedling roots, it was car- ried down the root channel at about 1.5 cm day -~, but never advanced beyond the root tip.
Bacterial dispersal by their own motility in soil water films will be very restricted. The largest recorded distance moved by self-propulsion was 60 mm day- ~ (Wong and Griffin, 1976) but generally it is below 10 mm day -1 (Hamdi, 1971; Soby and Bergman, 1983; Misaghi et al., 1992). Most studies on chemotaxis of soil bacteria, in which they migrate to sites of high nutrient concentration, have been conducted in vitro (see the review by Gammack et al., 1992), but Howie et al. (1987) showed that mutant strains of P. fluorescens without flagella colonized roots in a soil system as effectively as wild-type strains that possessed flagella. Thus either these wild strains were only flagellated in vitro, or it was not possible for them to make active flagellar movements in the soil.
Where a new population of bacteria is added to
soil, it is likely that most of the adsorption sites will be occupied by indigenous bacteria. There may be some exchange between these and the added popula- tion, but it is likely that the new population will be more readily redistributed by water fluxes. Thus added GMMs may be more mobile than indigenous organisms but more vulnerable to predation.
2.3. Actinomycetes
There are relatively few studies concerning acti- nomycete dispersal in soil, either by abiotic or inver- tebrate means, although the survival of recombinant forms has been monitored in sterile and non-sterile soil (eg Wang et al., 1989; Wang et al., 1991; Wellington et al., 1990). Actinomycetes exist pre- dominantly in soil as spores which are tolerant of prolonged starvation and desiccation (Wellington et al., 1990). It is as spores that dispersal is most likely to occur and subject to the same constriants as bacteria. Wang et al. (1989) showed that modified strains of Streptomyces lividans could survive in non-sterile soil for 30 days as both spore and mycelial inoculants, but some inoculated spores were able to survive at least 10 months. Wellington et al. (1990) showed that there was an initial short mycelial growth phase of 2 to 3 days in S. lividans and S. violaceola- tus, after which sporulation occurred. Therefore, to assess the survival of genetically modified actino- mycetes after their release into non-sterile soil, it will be necessary to monitor the rate of sporulation, and determine the conditions under which it occurs.
Crawford et al. (1993) studied the effect of a modified S. lividans strain on the numbers and activities of existing microorganisms in microcosms of an air-dried sieved silt loam. The survival and persistence of the modified organism was similar to its parent form, but its effects on existing microor- ganisms differed. The numbers of fungal propagules were transiently, but significantly, depressed but cel- lulose-degrading bacterial populations were: en- hanced.
If the spore form predominates in the soil, it is clear from the work of Ruddick and Williams (1972) that its structure and surface characteristics will be the factors determining how far the spore can be dispersed. These characteristics will affect both abi- otic and biotic dispersal and the fate of any geneti- cally modified forms after release. The single smooth
116 J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131
spores of Micromonospora sp. are much more likely to be moved downwards by percolating water than the spiny-sheathed spore chains of S. viridochromo- genes.
2.4. Fungi
Fungal hyphae are not discrete entities but form a connected network which extends beyond and be- tween soil particles and voids. The dispersal of fungi through soil can, therefore, only be modelled as spores, when they will behave in a similar way to bacteria. Only fragmented hyphae are able to act as propagules and, because of loss of physiological function of small fragments, many of these will not be viable. Fungal spores are larger than bacteria (2 -200 /zm diameter) and are, therefore, more easily retained by soils with small pore spaces. Ignoffo et al. (1977) found 75% of the conidia of Nomuraea rileyi were recovered from the filtrate of a sand after the equivalent of 16.25 cm of rain was applied. By contrast, 90% were retained in the top 2 cm of a silt loam. The degree of ornamentation, wettability and mucilaginous production of fungal spores influence the degree of adsorption to soil particles (Hepple, 1960). In undisturbed (non-tilled) soils, the vertical movement of fungal spores is normally small (cm) but this can be increased by tillage where spores and hyphal fragments may be dragged to the bottom of a plough layer (20-30 cm) and over tens of metres in the horizontal plane.
Motility of fungal zoospores of the order Mastigomycotina can account for small movements both upwards and downwards in the soil profile and are ineffective in dispersal. Mehrotra (1970) showed zoospore migration of Phytophthora spp. was largely dependent on water movement, and Duniway (1976) found that flooded soils were more suitable for ac- tive dispersal of Phytophthora cryptogea zoospores. Zoospores act as agents of long distance dispersal only i f they reach surface water. In a field study, most strawberry plants infected with P. fragariae occurred in a shallow depression running the length of the field, and receiving drainage water from an infected site where there had been standing water for much of the winter (Duncan, 1979). Plants adjacent to the central depression were infected within 2 months; those 18 m away were disease-free for 18 months.
Salas and Stack (1991) found that total Fusarium populations isolated from soil in a spring wheat cropping system were similar under conventional and reduced tillage, but the species composition was different. Fusarium equiseti was only 16% of the total in reduced tillage plots but it comprised 27% under conventional tillage. Fusarium culmorum, F. graminearum and F. avenaceum were isolated more frequently under reduced tillage than under a con- ventional regime. Herman (1990) found Gaeuman- nomyces graminis var. tritici was more common under conventional tillage than in minimal- and zero-tillage systems in a Chernozem soil. However, the number of acervuli of Colletotrichum gramini- cola developing in spring on Zea mays residues buried by autumn ploughing to 5-15 cm depth was significantly less than on residues left on the soil surface (Lipps, 1983). Conventional tillage of crop residues may also cause an increase in free fungal propagules in the soil (Angle et al., 1982).
Tillage redistributes fungal propagules vertically in the ploughed layer. Propagules of Fusarium solani sub sp. phaseoli (a root pathogen) were found dis- tributed through the ploughed layer (20-30 cm depth) of sandy loam and silt loam bean fields in Central Washington (Burke et al., 1972), but were seldom isolated from sub-soil (33-41 cm depth). Smith (1978) found that spores of arbuscular mycorrhizas were common in surface layers of soil under wheat and pasture, declining in numbers down the profile until they were rare at 50 cm. Tillage had no effect on total spore numbers, but direct drilled wheat contained more spores in the surface 8 cm and fewer from 8-15 cm than conventionally tilled plots, due to mechanical inversion of soil during cultivation. Zoospores of Peronosclerospora sorghi occurred at 0-20 cm in soil with conventional tillage, but dis- tributed to 20-40 cm with deep tillage (Janke et al., 1983).
3. The role of soil fauna in dispersal of microor- ganisms
The role of soil fauna in the potential movement of microbes is dependent on the ability of the animal to pick up or ingest and subsequently deposit mi- croorganism and the distance over which the animal is able to move. Microorganisms can be carried on
J. Dighton et al. / Applied Soil Ecology 5 (1997) 109-131 117
the animal's surface. Distance of transport may be related to the size of the animal; larger animals are able to move greater distances and are, therefore, of greater potential importance. Other fauna may ex- hibit seasonal or diurnal vertical migrations in soil, making them potentially important vectors. The na- ture of the dominant faunal taxa is dependent on soil type and varies by geographical region (Heal and Dighton, 1986; Petersen and Luxton, 1982); for ex- ample, earthworms are scarce in soils of low pH. It is, therefore, important to consider the potential role of all soil dwelling faunal groups in the dispersal of genetically modified microorganisms.
The literature on the rote of soil animals in dis- persing viruses relates mainly to the transmission and spread of plant diseases. Viruses do not have mechanisms to penetrate plant cell walls, so must either infect through wounds, or by vectors such as insects, mites, nematodes or fungi. Plant viruses are not spread in aerosols, but can be airborne in vectors, either arthropods or fungal spores (Brakke, 1992). The ability to be transmitted by a vector is a genetic property of the virus, and non-transmissible mutants are common. Nearly 400 species of animals are known to transmit at least one plant virus, of these about 94% are arthropods (mainly insects), and 6% are nematodes (Harris, 1981). Many of the insects, however, are aerial sap-suckers and so outside the limits of this review. With the exception of nema- todes and some mites, the role of the soil fauna appears to be limited, and even in those groups, their role is generally restricted to plant parasitic or root feeding species. It is possible for a vector to ingest a virus which can survive gut passage and be released in the faeces at a new site. There it would also require a suitable host in which to replicate, and/or another appropriate vector. However, plant viruses cannot survive gut passage and this method of dis- persal will generally only occur in insect-parasitic viruses (Miller et al., 1987). Since viral dispersal appears to be a special case, the following discussion will be limited to dispersal of bacteria, actinomycetes and fungi.
3.1. Protozoa
Naked amoebae, testate amoebae, Flagellata and Ciliata are all important members of the soil fauna
(Petersen and Luxton, 1982). Petz and Foissner (1988) found that active ciliates were abundant in the litter layer of a spruce forest, whereas testaceans peaked in the F and H horizon. There were very few active ciliates in meadows, or sites without litter layers. Protozoa accounted for less than 2% of the total biomass of soil organisms in field plots of winter wheat (Brussaard et al., 1990).
The activity of protozoa is closely related to soil moisture conditions (Bamforth, 1988). Soil pore size will also affect their distribution, as flagellates and small amoebae can occupy pore spaces down to 8 /zm in diameter and thus could be more important vectors in compacted soils. Laminger and Bucher (1984) measured the foraging activity of the tes- tacean Quadrulella symmetrica which reached a mean speed of 3.2 X 10 -2 mm min -1. Four food vacuoles were observed in 10 min. By extrapolation, this might be equivalent to ingestion of about 600 bacteria and movement over a distance of 5 cm day 1. Zwart and Darbyshire (1992) noted that a soil flagellate, Spumella sp. egested some intact Pseu- domonas fluorescens bacteria, but the amount of movement involved would be minimal. Protozoan effects are more likely to be those of reducing microbial numbers through grazing (Rutherford and Juma, 1992a and Rutherford and Juma, 1992b).
Fungal spore dispersal was demonstrated by naked amoebae (Heal, 1963) in vitro over a few millimeters and germination of the spores of Mucor hiemalis was shown to be unaffected by ingestion.
Movement of GMMs by protozoa is likely to be small, so it may be concluded that Protozoa are of limited importance in dispersal of GMMs and re- stricted to the order of centimetres at the most.
3.2. Nematoda
Nematodes are an abundant and heterogenous group, with up to 119 identified species of soil nematodes (Yeates, 1984). The free-living microbial feeders may be important dispersal agents, although, compared with larger invertebrates, their movement within the soil is limited. Reproductive strategies allow drought-resistant species to respond quickly to favourable conditions and dominate in dry soils. Nematode densities in wooded and non-wooded ecosystems are similar, 2.9 X 1 0 7 m - 2 in a mixed
118 J. Dighton et al. / Applied Soil Ecology 5 (1997) 109-13l
deciduous forest (Sohlenius, 1980); 4 to 6.3 X 106
m -2 temperate deciduous forest soils; 1 to 2.5 X 106 m -2 in temperate coniferous forests (Petersen and Luxton, 1982). Densities are greater in mull soils than in soils with a strongly developed organic hori- zon, and in mesic soils rather than 'wet' or 'dry' soils (Petersen and Luxton, 1982). Ingham et al. (1989) found bacterial-feeding nematodes to be dom- inant in meadows and prairie, while micro-arthro- pods dominated in lodgepole pine forest. In more extreme environments, such as tundra and alpine soils, nematodes account for more biomass and res- piration than any other soil fauna (Procter, 1990) with some species surviving in anaerobic conditions (Por and Masry, 1968).
Data concerning nematode movement in soil are mainly related to parasitic species. Georgis and Poinar (1983) found that the insect parasite, Het- erorhabditis bacteriophora migrated to the surface soil within 5 days from inoculation points at 28 cm depth, although most surface-inoculated animals tended to remain within the 0 -2 cm layer. The plant parasite, Meloidogyne arenaria, can move vertically 50 mm d- l (Freckman, 1988), and in a greenhouse experiment where they were placed at the bottom of a soil column, Meloidogyne chitwoodi individuals moved vertically upwards at an initial maximum of 80 mm days -l , to reach 400 mm in 9 days, 44 mm days-l overall (Pinkerton et al., 1987).
Bacterial-feeding nematodes play a key role in decomposition and nutrient cycling (Freckman, 1988), where part of this role may be to disperse the microorganisms, either through adherence to their cuticles (Bird, 1987) or through egestion of viable microbial cells. Up to 60% of ingested bacteria may survive gut passage (Jatala et al., 1974). Cayrol et al. (1987) used Cephalobus part, us as a cartier of the bacterium Rhizobium japonicum to inoculate field- grown soybeans. Compared with inoculated seeds, nematode-manipulated plants had a more even distri- bution of nodules along their roots, which increased with increasing nematode density. There is therefore potential to use them to inoculate useful bacteria into deficient soils.
Beare et al. (1989) observed higher numbers of bacterivorous nematodes in a field site modified with clover residues than in a no-tillage system, because of higher bacterial numbers in the former, and Over-
hoff et al. (1991) also showed that nematodes are affected by soil tillage. On the contrary, numbers of bacterial feeding nematodes were generally reduced in untilled soils and increased after ploughing at 12.5-25 cm depth (Sohlenius and Sandor, 1989). Christensen et al. (1992) found 30 times more nema- todes in the rhizosphere of barley roots than in the surrounding bulk soil, again as a result of higher bacterial activity. Griffiths (1990) related this in- crease to the ability of nematodes to migrate more readily than protozoa into the rhizosphere zone. The effects of soil management treatments, increasing nematode populations influences the degree of spread of released GMMs. Clarholm et al. (1981) used the number of bacterivorous nematodes as a measure of previous bacterial production as they considered it provided a better indicator than standing crop mea- surements of bacteria, which varied little, despite substantial bacterial production.
Fungal feeding nematodes tend to puncture hy- phae with their stylets and evacuate the cell contents, behaviour which is not conducive to dispersal of hyphal propagules. There is little evidence to support the claim that spores are ingested or transported on the body surface of these animals.
Nematodes may be causative agents for widespread microbial dispersal in their anhydrobiotic state (Demeure and Freckman, 1981), as they can be transported by the wind over several kilometres (Carroll and Viglierchio, 1981). Because of the high abundance of nematodes in many soils and their ability to move over distances of tens to hundreds of centimetres, they have the potential to disperse GMMs over short and long distances.
3.3. Acari
Four of the seven orders of Acari are generally represented in the soil fauna (Wallwork, 1967). Cryptostigmata are numerically the most abundant acarine group, and occur in greatest numbers in forest soils. Luxton (1982) found cryptostigmatid mites predominated in a beechwood soil; 3.4 × 10 4
m -2 out of a total mite population of 6.7 X 104.
Although numerous, they only represent a small proportion of the total biomass of soil animals in forest soils (Schaefer, 1982). However, even in soils with mull humus formations, the Cryptostigmata re-
J. Dighton et aL /Applied Soil Ecology 5 (1997) 109-131 119
main the dominant group, although cultivated soils are usually sparsely populated by them (Wallwork, 1967). Curry (1986) also found that species numbers and diversity were higher in grassland than ploughed soil, although Mueller et al. (1990) noted that con- ventional tillage may favour bacterivorous species. Lussenhop (1981) showed oribatid mites (a sub-group of Cryptostigmata) to be more numerous in raked than undisturbed prairie soil, and concluded that physical disturbance of the soil increased the number of oribatid, prostigmatid and astigmatid mites. Per- due and Crossley (1990) found no difference in the vertical distribution of mites in agricultural plots under no-tillage or conventional tillage, with the highest density at 0-5 cm in both systems.
Cryptostigmata are more sedentary than the Prostigmata and Mesostigmata. Berthet (1964) found that certain oribatid (Cryptostigmata) species could travel up to 11 to 20.5 cm day -1 A number of hemiedaphic species such as Scheloribates laeviga- tus show regular vertical movements into and from the epigeal zone, associated with humidity fluctua- tions (Wallwork and Rodriguez, 1961). There is likely to be seasonal vertical migration among some species in every order of mites. Prostigmatic mites are generally more abundant in the litter layer in summer, and cryptostigmatic mites in spring (Lux- ton, 1982). Gravid Cryptostigmata and Prostigmata may migrate to different parts of the soil profile to deposit eggs.
Mites may also carry bacterial, actinomycete and fungal propagules on their body surfaces. Oribatid mites have highly sculptured integuments which function as effective traps (Lussenhop, 1992). How- ever, they are not an important component of the arable soil faunal community where bacterial GMMs are most likely to be released. The role of soil arthropods in transporting actinomycetes has been examined by Ruddick and Williams (1972). They found that Acari and Collembola were more likely to transport smooth-spored Streptomyces griseus than the hairy spores of S. finlayi, with hydrophobic properties being more significant than the surface patterns. Fungi producing spore chains rather than single spores were more likely to be transorted, as the chains wrap around the cuticular hairs of acarine species (Ruddick and Williams, 1972). Approxi- mately 50% of the Acari and Collembola extracted
from pine litter or garden soil were found to carry the smooth-spored S. griseus; those isolated from beech litter did not. Behan and Hill (1978) isolated fungal colonies from the bodies and tracks of 58% of oribatid mites studied, but from only 14% of the faecal pellets. However, 90% of the pellets from Caloglyphus michaeli feeding on Pythium myrioty- lum, contained viable fungal remains (Shew and Beute, 1979).
Acari may be regarded as potentially important microbial vectors because of their range of trophic groups and their ability to trap bacterial and fungal propagules on body integuments and survival of propagules in the gut. Their vertical and horizontal migratory responses to microclimatic variation is also highly relevant.
3.4. Collembola
Collembola occur in nearly all soils, often in many thousands per m 2 (Anderson and Healey, 1972). They usually represent only a small percent- age of animal biomass in more fertile soils, e.g. only 1,4% of the biomass in a limestone beech forest where the macrofauna, lumbricids in particular, con- tributed the greatest majority (Schaefer, 1982). Tem- perate coniferous forests contain the highest average densities, between 3.2 × 104 and 2.1 × 105 m - : (Hagvar, 1982), and temperate deciduous forests and grasslands 4 to 7 × 104 (Petersen and Luxton, 1982).
Little is known about the extent of lateral move- ment of collembola in soil, but vertical migration has been studied by Takeda (1978); Hassall et al. (1986a) and Hassall et al. (1986b). They showed that thou- sands of Onychiurus subtenuis moved from the hu- mus layer to the litter, a distance of several cms in less than 18 h, in response to moisture and fungal food availability. This vertical movement would al- low the Collembola to disperse microbial spores to new substrate surfaces.
Collembola have great potential for GMM disper- sal, but mainly occur in coniferous soils with well- developed surface organic layers, where fungal food-webs predominate. Vannier (1975) demon- strated the important role they play in dispersing microbial propagules by heat-treating a forest soil and studying recolonisation. Large numbers of sur- face-dwelling Collembola with different diet prefer-
120 J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131
ences invaded, and many microorganisms that they had ingested were voided in a viable form into the sterilised soil. Although collembola are mainly fun- gal feeders (Hanlon and Anderson, 1979; Visser, 1985); Gomez et al. (1990) found large numbers of viable bacteria in the faeces of Proisotoma minuta, and attached to the integument. Stimulation of bacte- rial activity by collembolan grazing of the fungi could be important, but there does not appear to be much known about collembolan/bacterial interac- tions as such.
Collembola extracted from a range of habitats have approximately 10-20 fungal species associated with them (Christen, 1975; Pherson and Beattie, 1979; Wiggins and Curl, 1979). Visser et al. (1987) isolated over 100 species of fungi from Collembola collected from an aspen woodland soil, with the number of propagules and taxa per animal decreas- ing from the L to H layers. Wiggins and Curl (1979) found no evidence of transport of a non-sporulating pathogen, Rhizoctonia solani, on the body surface of Collembola. Visser et al. (1987) isolated fungi which did not sporulate on agar media, but it is unclear whether these would have been carried as spores or hyphae.
The vertical migrations of Collembola through the soil horizons and their ability to carry bacterial and fungal propagules in the gut and on the integument, suggests that they could be important vectors of GMMs over distances of tens to hundreds of cen- timetres.
3.5. Enchytraeidae
Enchytraeid worms rarely measure more than 2 cm in length. They are found in all habitats, with highest numbers recorded from Juncus peat, 1.45 N 105 m -2 (Peachey, 1963)and deciduous forest, 1.43 × 105 m -2 (Heck and R~Smbke, 1990). Numbers in arable soils are considerably lower, between 2 X 103 and 3 × 104 m -2 (Didden, 1993). They tolerate acid conditions, and establish large populations in organic soils of woodlands and moorlands (Wallwork, 1976; Huhta et al., 1967). Abundance is similar in conifer- ous and deciduous forest soils (Petersen and Luxton, 1982).
Soil moisture changes may induce vertical migra- tion (Ellenberg et al., 1986) but vertical distribution may also be species dependent, e.g. Cognettia
sphagnetorum lives mainly in the Scots pine litter layer and Achaeta brevivasa deeper down (GriSngriSft and Miehlich, 1983). Most enchytraeid worms are unable to burrow, but species of the genus Frederi- cia can burrow to a depth of 3.5-4.0 cm. Standen (1984) observed that a few other species, e.g. Hen- lea ventriculosa, transported material up to 0.5 cm. Ellenberg et al. (1986) found most enchytraids in the F layer in a German beech forest, and, in ploughed agricultural soils, the largest numbers are found at the ploughing depth (Didden, 1990).
Dispersal of GMMs by enchytraeid worms is likely to be limited to acidic soils and natural or semi-natural ecosystems where fungal food webs predominate and Latter and Howson (1978) found fungal hyphae in the faeces of many of them. Ponge (1990) showed that bacterial penetration of Scots pine litter was increased by enchytraeid tunnelling activities. They probably do not have great potential as vectors of microorganisms.
3.6. Lumbricidae
Earthworms are widely distributed in temperate ecosystems and may be present in high densities (Bishop, 1989). These large, mobile invertebrates are potentially important in transporting GMMs and their role in the introduction of beneficial soil microorgan- ism has recently been reviewed by Doube et al. (1994a).
Edwards and Lofty (1977) and Edwards and Bohlen (1996) reveiew earthworm populations and biomass for a range of temperate habitats. These reviews indicate a wide range in numbers; their density in some moorland soils may be only 102 m -2, in pig slurry they have been recorded at 9.6 × 102 m 2. Smaller numbers and lower biomass occur in acid mor soils, fallow soils, temperate coniferous soils and moorland soils than in mull soils (Petersen and Luxton, 1982) and they are generally scarce below pH 5.5 (Satchell, 1955; Piearce, 1972; Robin- son et al., 1992). Lee (1985) reported lower numbers in arable soils than in pastures. Different species tend to be associated with different soil types.
Earthworms of the different ecological categories (Bouch6, 1977) will affect microbial dispersal in different ways (Lavelle et al., 1989). Epigeic surface litter dwellers make no burrows, no recognizable casts and ingest little or no soil, anecic species cast
J. Dighton et aL /Applied Soil Ecology 5 (1997) 109-131 121
at the entrance of their burrows and move leaf litter from the surface into their burrows. Endogeic species form extensive burrows in which they cast and feed on soil organic matter, litter form the surface and roots. Thus the latter two groups are more likely to be important in movement of modified microorgan- isms.
Soil physical changes resulting from earthworm activity have many secondary effects on the micro- bial populations. Burrowing increases pore volume, enhances aeration and field moisture capacity and reduces compaction and waterlogging (Edwards et al., 1990; Kirkham, 1981; Lee, 1985). Endogeic worms which inhabit the deeper horizons are proba- bly only important in short distance dispersal (Re- ddell and Spain, 1991), the degree of which will depend on their feeding and casting behaviour. Some species, such as A. longa and L. terrestris, build vertical burrows, others, such as A. caliginosa and L. rubellus, move further horizontally, so the species distribution in different soil types will affect micro- bial dispersal in a range of ways. As mean burrow length can vary from 160 mm (6 mm diameter) for O. cyaneum to 450 mm (3 mm diameter) for L. rubellus (Boulton and Phillipson, 1976), there will be important differential effects on soil water move- ment and associated microbial propagules.
There is little information on the horizontal dis- persal of earthworms, though Hamblyn and Dingwall (1945) claimed A. caliginosa could advance by 10 m per year from inoculation points in recently limed grasslands. Stein et al. (1992) studied the horizontal dispersal of A. caliginosa, A. longa and L. rubellus in permanent grassland between 1983 and 1990, and found an overall rate of 13 m per year. L. terrestris has been found to move up to 19 m on the soil surface in one night (Mather and Christensen, 1988).
The interactions between earthworm numbers and tillage practice may also be important. Direct drilling results in more earthworms than mould-board ploughing (Carter et al., 1988) or conventional tillage (Haines and Uren, 1990; Doube et al., 1994b). Mad- sen and Alexander (1982) and Fredericksen et al. (1989) showed a link between tillage, earthworms and bacterial dispersal. Vertical movement of Rhizo- bium japonicum and Pseudomonas putida in perco- lating water was increased by the presence of L. rubellus and also by ploughing. Bacteria were dis-
persed throughout the 60 cm core by percolating water and passage through earthworm guts.
Oligochaetes, particularly the lumbricids, are probably the most important agents of fungal disper- sal in soil. A large number of fungal species are able to survive passage through the gut as either spores or hyphal fragments (Murray et al., 1985; Edwards and Fletcher, 1988; Rabatin and Stinner, 1988; Reddeil and Spain, 1991). They can then increase rapidly in the casts (Shaw and Pawluk, 1986). Earthworms may be selective in their feeding. Lumbricus terrestris, Apporectodea longa and Allelobophora chlorotica when offered straw colonised by different fungal communities, preferred early colonising fungal com- munities (capable of utilising soluble sugars and cellulose) to lignin decomposers (Moody et al., 1995). In a parallel study, Moody et al. (1997) showed that fungal spores survived passage through the gut of L. terrestris and A. longa and survival varied with fungal species. After passage through the gut, Fusar- ium lateritium but Agrocybe temulenta spores failed to germinate; Trichoderma sp. and Mucor heimalis spore germination was reduced, but germination of Chaetomium globosum spores was enhanced. Thus, earthworms can be important in modifying the com- munity structure of fungal populations and the ability to act as a vector will depend on the species of fungus.
Many studies have shown higher numbers of bac- teria in faeces than in bulk soil (e.g. Edwards and Lofty, 1977; Shaw and Pawluk, 1986). Pedersen and Hendriksen (1993) showed significant changes, both quantitative and qualitative, in the bacterial flora: Escherichia coli and Pseudomonas putida declined during gut passage, but Enterobacter cloacae and Acromonas hydrophila survived to a similar degree. Retention time in the gut and site of defaecation will also affect how far an earthworm can disperse an ingested bacterium. Retention time varies from 3 to 4 h in E. fetida to 12 to 20 h in L. terrestris (Edwards and Lofty, 1977). In microcosms, Henschke et al. (1989) fed L. terrestris with a genetically modified bacterium, Pseudomonas fluorescens. Only 20% of the original inoculum of 5 x 106 cells were viable in the worm faeces after 44 days and, after 50 days, they were below the limits of detection. They con- cluded that the risk of dispersal of GMMs by L. terrestris was limited to 50 days after relase in those
122 J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131
experimental conditions, but the GMMs might also survive in the secretions lining the burrows. Exten- sive studies on the movement of Pseudomonas and Rhizobium (Doube et al., 1994b and Doube et al., 1994c) show evidence of bacterial dispersal over 15-20 cm by endogeic earthworms. Of the two species of worm, Aporrectodea trapezoides was a more effective vector than A. rosea. Burrowing activity of worms and, hence, their potential to carry microorganisms appears to be incresead by surface soil organic matter (Hughes et al., 1996).
The interaction between earthworms and soil mi- crobial communities has recently been reviewed by Brown (1995) where their importance as microbial regulators has been highlighted. To summarize, the movement of worms over large distances (metres) in both the vertical and horizontal plane allows for effective long-distance dispersal. In addition, the construction of burrows increases the macropore density of soil, enhancing movement by abiotic fac- tors.
3. 7. Other invertebrates
A number of other larger soil invertebrates may be significant in dispersing soil microorganisms, al- though they are considerably less abundant than the mesofauna (Collembola, Nematoda and Acari). Diplopoda and Isopoda occur in numbers of 500 m -2 in temperate grassland soils (Macfadyen, 1968), two or three orders of magnitude lower than Acari or Collembola. There are about 300 m -2 in permanent grassland to a depth of 15 cm, and about 200 m -2 in ploughed arable soils (Wallwork, 1976). Populations of three small ant species, Lasius alienus, L. niger and Tetramoriurn caespitosum in sandy heath soils, contain about 5 × 10 3 worker ants m -2. Similarly high estimates of populations of Lasius flavus have been found in English neutral grassland. In temperate sites, ants are practically absent on the forest floor (Petersen and Luxton, 1982). Although there is no reported work on their potential to disperse microor- ganisms, it may be significant because of their mo- bility. Their role will presumably be to carry the propagules on their external body surfaces rather than by enhancing microbial growth during gut pas- sage, but there are no data to verify this.
Macroarthropods may cause less damage to the microbial propagule during passage through the gut
than smaller invertebrates (Visser, 1985). Particle sizes in the faecal pellets may be a factor in their suitability for colonisation by microorganisms; in- gested plant material is reduced to particles of 200- 300 /zm by medium-sized arthropods and 20-50 /xm by Collembola and Acari (Swift et al., 1979). Anderson and Bignall (1980) showed 10- to 100-fold increases in soil bacterial numbers after egestion by the millipede Glomeris marginata. Hanlon (1981) studied gut content and faeces of G. marginata and the isopod Oniscus asellus and recorded a 7-fold increase in bacterial numbers during gut passage. Similar results were reported by Griffiths et al. (1989) for the isopod Porcellio scaber and by Gunnarsson and Tunlid (1986) for O. asellus. They found that growth and lysis of bacteria occurred in the gut and that gram-positive and gram-negative bacteria were differentially affected.
Although these other invertebrates may be impor- tant fungal feeders (particularly the Diptera and Isopoda) there are few data available on their ability to distribute fungal propagules. Rabatin and Stinner (1988) noted the presence of vesicular arbuscular mycorrhizal spores in the guts of carabid and scarab beetles and isopods from agricultural plots, and found some of them to be viable. In the same study, chlamydospores of Glomus intradices survived gut t ransport in the mil l ipede Cylindroiulus coeruleocinctus, and so spread to infect previously uninfected roots of alfalfa. In acidic organic soils, dipteran larvae of the Tipulidae are a dominant group of the soil fauna. Their potential for dispersal of fungal propagules is high, but there are no data on their actual abilities to disperse microorganisms and it is unlikely that GMMs would be released into such sites.
A secondary factor influencing dispersal by faunal vectors is the role of Oligochaeta and Diptera larvae as food for other animals, particularly birds. These invertebrates may be carried over distances of many kilometres especially when being delivered as food for the young.
There appears to be a lack of information on the potential for larger mobile soil fauna (other than earthworms) to move fungal propagules. Litter in- habiting dipteran larvae, isopods and ants may be agents for long distance dispersal but little is known about the extent of this.
J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131 123
4. Discussion
Movement of viruses and bacteria through soil by percolating water has been extensively modelled (see reviews by Murphy and Tare (1996) and Wiebers et al. (1995)). However, these models have been con- structed from data obtained from homogenised soils. The importance of heterogeneity in natural soils (vertical stratification, root channels, soil faunal bur- rows, etc) are complicating factors which are ex- cluded from the models. Soil management practices (ploughing, tyning etc) and selection of crop species (rooting depth) can alter soil physical properties and also physically move microorganisms for distances ranging from metres to kilometres.
Interactions between microbes and soil fauna can influence microbial dispersal by substantially alter- ing the physical structure of soil throught the cre- ation of continuous voids (earthworm burrows) or discontinuous voids (change of soil structure by Collembola and mites (Rusek, 1975; Rusek, 1978) or by acting as vectors of microbial propagules.
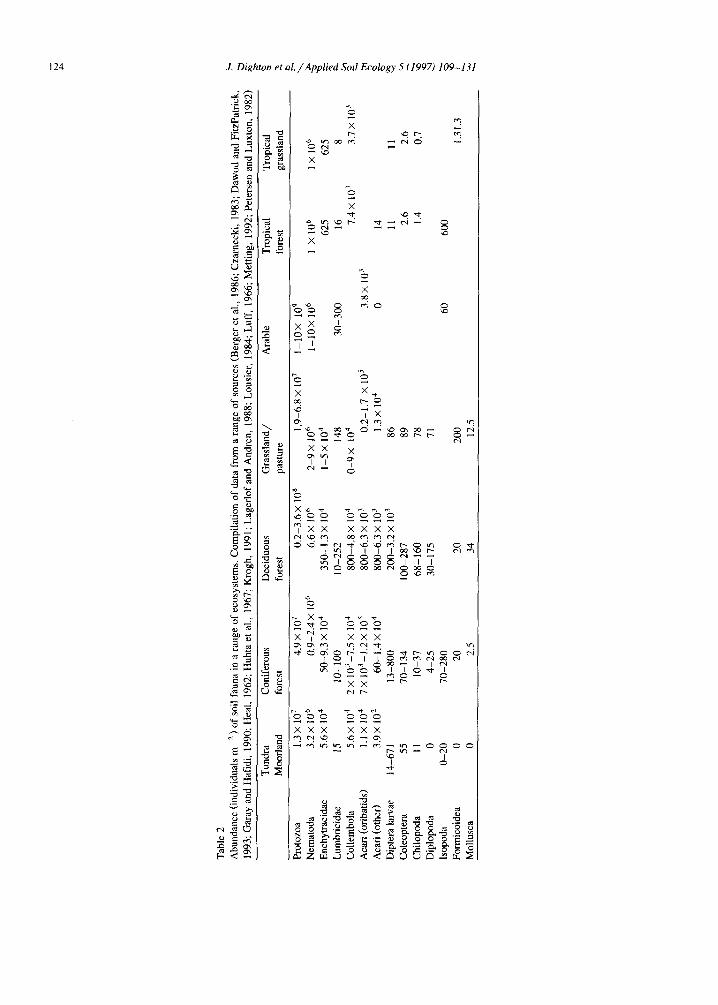
The importance of soil fauna as vectors of soil microorganisms depends on the species composition of the fauna and the nature of the dominant micro- bial component of the soil. As soils become more acid and less disturbed, organic residues accumulate, favouring fungal growth over bacterial. The domi- nant soil faunal taxon also changes, with Lumbrici- dae dominating in grassland and deciduous wood- land compared with Enchytraeidae in acidic soils of coniferous forests and tundra. Acari are more prolific in wooded than non-wooded systems. Collembola are widespread, but are a less important component of fertile soils. Faunal biomas is greatest in temper- ate deciduous and steppe ecosystems, declining to- wards both warm dry and cool systems (Heal and Dighton, 1986). Details of soil faunal species in different ecosystems and their activities was sum- marised by Wallwork (1976) and an index of differ- ences between ecosystems is given in Table 2.
As different animal taxa have differing capacities to move through soil, their potential to carry mi- croorganisms also differs from ecosystem to ecosys- tem. Macrofauna such as lumbricids, which domi- nate in many temperate and tropical forests and grasslands, may move much larger distances than their smaller counterparts, the dominant meso- and
microfauna of temperate forest and tundra regions (e.g. Enchytraeidae, Nematoda and Collembola). GMM release is more likely in arable than semi-nat- ural soils and these are more likely to contain high lumbricid numbers.
Differences in behaviour of soil fauna also influ- ence their potential to carry microflora. Van der Drift (1965) compared behaviour of animals feeding on woodland leaf litter and their attendant microfloral communities. Where free water exists between dead leaves on the woodland floor, Nematoda (micro- and mesofuna) are likely to be among the initial grazers on leaf surface microflora. Collembola, Acari (mesofauna) and, to some extent, Enchytraeidae are more tolerant of water stress and will graze leaf surfaces in the absence of free water. Larger animals, Mollusca, Diplopoda and Isopoda may take large pieces of leaf litter and have the capacity to move large distances (metres) over the soil surface.
From the available evidence, Protozoa are un- likely to be important in dispersing microorganisms. Nematode motility is restricted and, except in the wind-blown anhydrobiotic state, they are probably not of great significance. Their effect is probably limited to dispersal over centimetres. Movement of microarthropods within the soil is greater, so their potential effect in moving microorganisms is; also greater.
Collembola or Acari densities of 1-7 × l05 m -2 in protorendzina and ranker soils in Czechoslovakia, where soil structure is composed of faecal material derived from the activities of these animals (Rusek, 1975; Rusek, 1978). The nature of the faecal pellets derived from the different animals associated with these soils (Table 3) affects pore size, void surface roughness and the physicochemical nature of the surface and hence its adsorption characteristics. On a more local scale, the activities of termites have been shown to have similar effect on soil structure in Kenya (Wielemaker, 1984), and can enhance soil erosion potential as well as influencing water perco- lation patterns. Acari and Collembola have orna- mented integuments which are likely to trap microor- ganisms and facilitate their dispersal. Spores and hyphal fragments can survive gut passage.
Enchytraeid dispersal is probably fairly limited and is also more likely to occur in natural or semi- natural ecosystems of low pH, but the potential role
Tab
le 2
A
bund
ance
(in
divi
dual
s m
2 )
of
soil
fau
na i
n a
rang
e of
eco
syst
ems.
Com
pila
tion
of
data
fro
m a
ran
ge o
f so
urce
s (B
erge
r et
al.,
198
6; C
zarn
ecki
, 19
83;
Daw
od a
nd F
itzP
atri
ck,
"~
1993
; G
aray
and
Haf
idi,
1990
; H
eal,
1962
; H
uhta
et
al.,
1967
; K
rogh
, 19
91;
Lag
erlo
f an
d A
ndre
n, 1
988;
Lou
sier
, 19
84;
Luf
f, 1
966;
Met
ting
, 19
92;
Pete
rsen
and
Lux
ton,
198
2)
~'
Tun
dra
Con
ifer
ous
Dec
iduo
us
Gra
ssla
nd/
Ara
ble
Tro
pica
l T
ropi
cal
Moo
rlan
d fo
rest
fo
rest
pa
stur
e fo
rest
gr
assl
and
.,-..,
Prot
ozoa
1.
3X 1
07
4.9X
107
0
.2-3
.6x
10 s
1.
9-6.
8 X
107
1-
10X
10
9 ~
. N
emat
oda
3.2x
106
0.
9-2.
4 x
l06
6.6X
106
2
-9x
10
6 1
-10
x 1
06
1 X
106
1X
106
E
nchy
trae
idae
5.
6 x
104
50-9
.3 x
104
35
0-1.
3 x
104
1-5
X 1
04
625
625
"~m
L
umbr
icid
ae
15
10-1
00
10-2
52
148
30-3
00
16
8 C
olle
mbo
la
5.6x
104
2X
103
-7.5
X 1
04
800-
4,8X
l04
0
-9X
10
4 7.
4X 1
03
3.7x
103
~z
A
cari
(or
ibat
ids)
1.
1X 1
04
7X 1
03-1
.2x
105
800-
6.3
X 1
03
0.2-
1.7
x 10
5 3.
8 X
103
A
cari
(ot
her)
3.
9 X
102
60
-1.4
X 1
04
800-
6.3
X 1
03
1.3
X 1
04
0 14
D
ipte
ra l
arva
e 14
-671
13
-800
20
0-3.
2 X
103
86
11
11
C
oleo
pter
a 55
70
-134
10
0-28
7 89
2.
6 2.
6 C
hilo
poda
11
10
-37
68-1
60
78
1.4
0.7
~e
Dip
lopo
da
0 4-
25
30-1
75
71
lsop
oda
0-20
70
-280
60
60
0 ~e
Fo
rmic
oide
a 0
20
20
200
1.31
.3
Mol
lusc
a 0
2.5
34
12.5
~'
J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131 125
Table 3 Characteristics of faecal pellets of Collembola, Acari and Enchytraeidae which, when forming a major portion of soil, influence water flow and retention patterns (from Rusek, 1975)
Size ( # m ) 30-90 30-50 120-200 Shape Irregular, round Oval, round Extremely irregular Surface Uneven Smooth Uneven Mineral particles Present Usually Absent Present Feature Compact Compact Often divided into primary components
of lumbricids will depend on the ecosystem, soil type and species of worm. Earthworms are predominant in fertile agricultural soils dominated by bacteria. Moreover the widely reported ability for bacteria to multiply in the earthworm gut, worm casts and bur- row linings suggests that their activity in soils, where GMM releases are being considered, should be in- vestigated. Another important aspect of lumbricid dispersal is their role in the formation of soil struc- ture through their casting and burrowing activities. In this way they interact with the abiotic factors affect- ing dispersal and exacerbate drainage effects. These factors are discussed in detail by Brown (1995).
Other larger invertebrate groups such as Diplopoda and Isopoda may also be important in dispersing bacteria, particularly as the ingested bacteria will be more likely to survive gut passage than in the small Nematoda, for example. However data on the role these animals may play in microbial dispersal are negligible.
Finally, most of the experimental studies with microcosms involve packing them with air-dried and sieved soils. Treatment comparisons are often made between sterile and non-sterile soils. In spite of the potential shortcomings, in restricting water move- ment to vertical flow for example, microcosms are a useful abstraction from natural conditions. However, intact, undisturbed soil cores containing a native microflora, with and without key groups of soil fauna, would provide more useful information in relation to the potential of the faunal groups to disperse GMMs under field conditions.
Acknowledgements
This paper is a summary of the findings reported to the Department of the Environment (UK) under
contract PECD 7 / 8 / 2 3 4 to review the potential of soil fauna and abiotic factors to disperse genetically modified microorganisms (DOE, 1995).
We would also like to thank two anonymous reviewers for their constructive comments on an earlier version of this paper.
References
Alexander, M., 1986. Ecological concerns relative to genetically engineered microorganisms. In: V. Jensen, A. Kjoller and L.H. Sorensen (Editors). Microbial Communities in Soil. Elsevier; London, pp. 347-354.
Amarger, N., Hirsch, P. and Klingmiiller, W., 1991. Assessing the risks involved in the release of genetically manipulated mi- croorganisms. In: I. Economides (Editor). Biotechnology R and D in the EC. Part IL Detailed Final Report of BAP contractors in Risk Assessment (1985-1990), pp. 55-59.
Anderson, J.M. and Bignall, D.E., 1980. Bacteria in the food gut contents and faeces of the litter feeding millipede Glomeris marginata. Soil Biol. Biochem., 12: 251-254.
Anderson, J.M. and Healey, I.N., 1972. Seasonal and inter-specific variation in major components of the gut contents of some woodland Collembola. J. Anim. Ecol., 41: 359-368.
Angle, J.S., Dunn, K.A. and Wagner, G.H., 1982. Effect of cultural practices on the soil population of Aspergillus flavus and Aspergillus parasiticus. Soil Sci. Soc. Am. J., 46: 301- 304.
Bamforth, S.S., 1988. Interactions between protozoa and other organisms. Agric. Ecosys. Environ., 24: 229-234.
Barnthouse, L.W. and Palumbo, A.V., 1985. Assessing the trans- port and fate of bioengineered microorganisms in the environ- ment. In: J.R. Fiksel and V. Covel!o. (Editors). Biotechnology Risk. Assessment Issues and Methods for Environmental In- troduction. Pergamon, Oxford, pp. 109-128.
Beare, M.H., Blair, J.M. and Parmelee, R.W., 1989. Resource quality and trophic responses to simulated throughfall effects on decomposition and nutrient flux in a non-tillage agroe- cosystem. Soil Biol. Biochem., 21: 1027-1036.
Behan, V.M. and Hill, S.B., 1978. Feeding habits and spore dispersal of oribatid mites in the north American arctic. Rev. Ecol. Biol. Sol, 15: 497-516.
126 J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131
Bentjen, S.A., Fredrickson, J.K., Van Voris, P. and Li, S.W., 1989. Intact soil-core microcosms for evaluating the fate and ecological impact of the release of genetically engineered microorganisms. Appl. Environ. Microbiol., 55: 198-202.
Berger, H., Foissner, W. and Adam, H., 1986. Field experiments on the effects of fertilizers and lime on the root microfauna of an alpine pasture. Pedobiologia, 29: 261-272.
Berthet, P., 1964. L'activite des oribatids d'une chenaie. Mem- oires de l'lnstitut Royale des Sciences Naturelles de Belgique, 152: 1-152.
Bird, A.F., 1987. Adhesion of microorganisms to nematodes. J. Nematol., 19: 514.
Bishop, D.H.L., Entwistle, P.F., Cameron, I.R., Allen, C.J. and Possee, R.D., 1988. Field trials of genetically-engineered bac- ulooirus insecticides. In: M. Sussman, C.H. Collins, F.A. Skinner and D.E. Stewart-Tully (Editors). The Release of Genetically-engineered Microorganisms, Academic Press, London: pp. 143-180.
Bishop, G.P., 1989. Review of Biosphere Information: Biotic Transport of Radionuclides as a Result of Mass Movement of Soil by Burrowing Animals. (NSS/RI94). Nirex Ltd, Didcot.
Bitton, G., Lahav, N. and Henis, Y., 1974. Movement and reten- tion of Klebsiella aerogenes in soil columns. PI. Soil, 40: 373-380.
Bouch6, M.B., 1977. Stratigies Lombriciens. In: U. Lohm and T. Persson (Editors). Soil Organisms as Components of Ecosys- tems, (Ecological Bulletin No. 25). Swedish Natural Science Research Council, Stockholm: pp. 122-132.
Boulton, P.J. and Phillipson, J., 1976. Burrowing, feeding, eges- tion and energy budget of AUolopobophora rosea (Savigny) (Lumbricidae). Oecologia, 23: 225-245.
Brakke, M.K., 1992. Containment, decontamination and mitiga- tion of plant viruses in the environment. In: M.A. Levin, R.J. Scidler and M. Regnal (Editors). Microbial Biology: Princi- ples, Methods and Applications. McGraw-Hill, Maidenhead.
Breitenbeck, G.A., Yang, H. and Dunigan, E.P., 1988. Water- facilitated dispersal of inoculant Bradyrhizobium japonicum in soils. Biol. Fert. Soils, 7: 58-62.
Brown, G.G., 1995. How do earthworms affect microflora and faunal community diversity? Plant Soil., 170: 209-231.
Brussaard, L., Bouwman, L.A., Geurs, M., Hassink, J. and Zwart, K.B., 1990. Biomass composition and temporal dynamics of soil organisms of a silt loam soil under conventional and integrated management. Netherlands J. Agric. Sci., 38: 283- 302.
Burke, D.M., Holmes, L.D. and Barker, A.W., 1972. Distribution of Fusarium solari f.sp. phaseoli and bean roots in relation to tillage and soil compaction. Phytopathology, 62: 550-554.
Carroll, J.J. and Viglierchio, D.R., 1981. On the transport of nematodes by the wind. J. Nematol., 13: 476-483.
Carter, M.R., Johnston, H.W. and Kimpinski, J., 1988. Direct drilling and soil loosening for spring cereals on a fine sandy loam in atlantic Canada. Soil Till. Res,, 12: 365-384.
Cayrol, J.C., Frankowski, J.P. and Quiles, C., 1987. Cephalobus parvus as a carrier of Rhizobium japonicum in field experi- ment on soybean cultures. Rev. Nematol., 10: 57-59.
Christen, A.A., 1975. Some fungi associated with Collembola. Rev. Ecol. Biol. Sol, 12: 723-728.
Christensen, H., Griffiths, B. and Christensen, S., 1992. Bacterial incorporation of tritiated thymidine and populations of bacte- riophagous fauna in the rhizosphere of wheat. Soil Biol. Biochem., 24: 703-709.
Clarholm, M., Popovic, B., Rosswall, T., Soderstrom, B., Sohle- nius, B., Staaf, H. and Wiren, A., 1981. Biological aspects of nitrogen mineralization in humus from a pine forest podzol incubated under different moisture and temperature conditions. Oikos, 37: 137-145.
Coleman, D.C., 1994. Development and consequences of commu- nity structure in soil systems: is there room in the middle? In: K. Ritz, J. Dighton and K. Giller (Editors). Beyond the Biomass: Compositional and Functional Analysis of Soil Mi- crobial Communities. Wiley, Chichester, pp. 201-220.
Crawford, D.L., Doyle, J.D., Wang, Z., Hendricks, C.W., Bentjen, S.A., Bolton, H., Frederickson, J.K. and Bleakley, B.H., 1993. Effects of a lignin peroxidase-expressing recombinant Strepto- myces lividans TK23.1, on biogeochemical cycling and the numbers and activities of micro-organisms in soil. Appl. Envi- ron. Microbiol., 59: 508-518.
Curry, J.P., 1986. Effects of management on soil decomposers and decomposition processes in grasslands and croplands. In: M.J. Mitchell and J.P. Nakas (Editors). Microfloral and Faunal Interactions in Natural and Agro-ecosystems, Nijhoff/Junk, Dordrecht, pp. 349-398.
Czarnecki, A.J., 1983. Springtails (Apterygota, Collembola) as index of forest in the development. In: Ph. Lebrun, H.M. Andre, A. De Medts, C. Gregoire-Wibo and G. Wanthy (Editors). New Trends in Soil Biology. Dieu-Brichart, Lou- vain-la-Neave, pp. 643-644.
Dawod, V. and FitzPatrick, E.A., 1993. Some population sizes and effects of the Enchytraeidae (Oligochaeta) on soil struc- ture in a selection of Scottish soils. Geoderma, 56: 173-178.
Demeure, Y. and Freckman, D., 1981. Recent advances in the study of anhydrobiosis in nematodes. In: B.M. Zuckerman and R.A. Rohde (Editors). Plant Parasitic Nematodes, Vol 3. Aca- demic Press, NY, pp. 204-225.
Dickinson, R.A., 1991. Problems with using existing transport models to describe microbial transport in porous media. In: C.J. Hurst (Editor). Modelling the Environmental Fate of Microorganisms. American Society of Microbiology, Wash- ington DC, pp. 21-47.
Didden, W.A.M., 1990. Involvement of the Enchytraeidae (Oligochaeta) in soil structure evolution in agricultural fields. Biol. Fert. Soil., 9: 152-158.
Department of the Environment, 1995. Dispersal of Genetically Modified Microorganisms by Abiotic Factors and Soil Inverte- brates. Research Report 4; Genetically Modified Organisms Research Report, 134 pp.
Donegan, K., Seidler, R. and Matyac, C., 1991. Physical and chemical control of released microorganisms at field sites, Can. J. Microbiol., 37: 708-712.
J. Dighton et aL /Applied Soil Ecology 5 (1997) 109-131 127
Doube, B.M., Stephens, P.M., Davoren, C.W. and Ryder, M.H., 1994a. Earthworms and the introduction and management of beneficial soil microorganisms. In: C.E. Pankhurst, B.M. Doube, V.V.S.R. Gupta and P.R. Grace (Editors). Soil Biota: Management in Sustainable Farming Systsems, CSIRO, pp. 32-41.
Doube, B.M., Stephens, P.M., Davoren, C.W. and Ryder, M.H., 1994b. Interactions between earthworms, beneficial soil mi- crorganisms and root pathogens. Appl. Soil Ecol., 1: 3-10.
Doube, B.M., Ryder, M.H., Davoren, C.W. and Stephens, P.M., 1994c. Enhanced root nodulation of subterranean clover (Tri-
folium subterraneum) by Rhizobium leguminosum biovar tri- folii in the presence of the earthworm Aporrectodea trape- zoides (Lumbricidae). Biol. Fert. Soils, 18: 169-174.
Drahos, D.J., Barry, G.F., Hemming, B.C., Bran&, E.J., Skipper, H.D., Kline, E.L., Kluepfel, D.A., Hughes, T.A. and Gooden, D.T., 1988. Prerelease testing procedures: US field test of LacZY-engineered bacterium. In: M. Sussman, C.H. Collins, F.A. Skinner and D.E. Stewart-Tully (Editors). Release of Genetically-engineered Organisms. Academic Press, London, pp. 181-192.
Duboise, S.M., Moore, B.E. and Sagik, B.P., 1976. Poliovirus survival and movement in a sandy forest soil. Appl. Environ. Microbiol., 31: 536-543.
Duncan, J.M., 1979. In-situ bait plants as a method for detecting red core Phytophthora fragariae in strawberry fields. Ann. Appl. Biol., 92: 307-312.
Duniway, J.M., 1976. Movement of zoospores of Phytophthora cryptogea in soils of various textures and matric potentials. Phytopathology, 66: 877-882.
Edwards, C.A. and Bohlen, P.J., 1996. Biology and Ecology of Earthworms, 3rd edn. Chapman and Hall, London.
Edwards, C.A. and Fletcher, K.E., 1988. Interactions between earthworms and micro-organisms in organic-matter break- down. Agric. Ecosys. Environ., 24: 235-247.
Edwards, C.A. and Lofty, J.R., 1977. Biology of Earthworms, 2nd edn. Chapman and Hall, London, 333 pp.
Edwards, W.M., Shipitalo, M.J., Owens, L.B. and Norton, L.D., 1990. Effect of Lumbricus terrestris L. burrows on hydrology of continuous no-till corn fields. Geoderma, 46: 73-84.
Ellenberg, H., Mayer, R. and Schauermann, J., 1986. Okosystem- forschung: Ergebnisse des Sollingprojekts, 1966-1986. Ulmer, Stuttgart.
Evans, G.O., Sheal, J.G. and Macfarlane, D., 1961. The Terres- trial Acari of the British Isles: an Introduction to their Mor- phology, Biology and Classification. British Museum (Natural History), London, 219 pp.
Franz, G., 1976. The influence of deep loosening of pseudogley on the microbial activity of the soils and the stability of this soil treatment. Landwirtschaftliche Forschung, 29: 77-87.
Fredericksen, J.K., Bentjen, S.A., Bolton, H.J., Li, S.W. and Van, V.P., 1989. Fate of TN5 mutants of root growth-inhibiting Pseudomonas sp. in intact soil-core microcosms. Can. J. Mi- crobiol., 35: 867-873.
Funderburg, S.W., Moore, B.E., Sorber, C.A. and Sagik, B.P., 1979. Soil column preparation for evaluation of viral transport. Appl. Environ. Microbiol., 38: 102-107.
Gammack, S.M., Paterson, E., Kemp, J.S., Cresser, M.S. and Killham, K., 1992. Factors affecting the movement of mi- croorganisms in soils. In: G. Stotsky and J.M. Bolland (Edi- tors). Soil Biochem., 7: 263-305.
Garay, I. and Hafidi, N., 1990. Study of a mixed forest litter of Lorabeam ( Carpinus betulus L.) and oak ( Quercus sessiliflora Smith) III. Organization of the edaphic microarthropod com- munity as a function of litter quality. Acta Oecol., 11: 43-60.
Georgis, R. and Poinar, G.O.J., 1983. Vertical migration of Het- erorhabditis bacteriophora and Heterorhabditis heliothidis (Nematoda, Heterorhabditidae) in sandy loam soil. J. Nematol., 15: 652-654.
Gerba, C.P., Yates, M.V. and Yates, S.R., 1991. Quantitation of factors controlling viral and bacterial transport in the subsur- face. In: C.J. Hurst (Editor). Modelling the Environmental Fate of Microorganisms. Washington, DC, American Society for Microbiology.
Gomez, M.A., Perez, M.T. and Sagardoy, M.A., 1990. Bacteria living on the integument and the faeces of Proisotoma minuta (Collembola: Isotomidae). Rev. Ecol. Biol. Sol, 27: 449-457.
Goodey, T., 1951. Soil and Freshwater Nematodes: a Monograph. Methuen, London, 389 pp.
Goyal, S.M. and Gerba, C.P., 1979. Comparative adsorption of human enteroviruses, Simian Rotovirus and selected bacterio- phages to soils. Appl. Environ. Microbiol., 38: 241-247.
Griffin, D.M. and Quail, G., 1968. Movement of bacteria in moist particulate systems. Aust. J. Biol. Sci., 21: 579-582.
Griffiths, B.S., 1990. A comparison of microbial-feeding nema- todes and protozoa in the rhizosphere of different plants. Biol. Fert. Soils., 9: 83-88.
Griffiths, B.S., Wood, S. and Cheshire, M.V., 1989. Mineraliza- tion of carbon-14-1abelled plant material by Porcellio scaber. Pedobiologia, 33: 355-360.
Gr~Sngrtift, A. and Miehlich, G., 1983. Bedeutung der Boden- feuchte fur die Populationsdynamik von Enchytraeidae (Oligochaeta) und Oribatei (Acari). Untersuchung in der Ro- hhumusauflage in Kiefernforst Gartow (Kreis Luchnow-Dan- nenberg). Abhandlungen der Naturwissenschaftlichen Vereins in Hamburg, 25:115-131.
Gunnarsson, T. and Tunlid, A., 1986. Recycling of fecal pellets in isopods Microorganisms and nitrogen compounds as potential food for Oniscus asellus L. Soil Biol. Biochem., 18: 595-600.
Hagvar, S., 1982. Collembola in Norwegian coniferous forest soils. I. Relation to plant communities and soil fertility. Pedo- bio|ogia, 24: 255-296.
Haines, P.J. and Uren, N.C., 1990. Effects of conservation tillage farming on soil microbial biomass, organic matter and earth- worm populations, in north-eastern Victoria. Aust. J. Exp. Agric., 30: 365-371.
Hamdi, Y.A., 1971. Soil water tension and the movement of rhizobia. Soil Biol. Biochem., 3: 121-126.
128 J. Dighton et al. / Applied Soil Ecology 5 (1997) 109-131
Hanlon, R.D.G., 1981. Influence of grazing by Collembola on the activity of senescent fungal colonies grown on media of different nutrient concentration. Oikos, 36: 362-367.
Hanlon, R.D.G. and Anderson, J.M., 1979. The effects of Collem- bola grazing on microbial activity in decomposing leaf litter. Oecologia, 38: 93-99.
Harris, K.F., 1981. Arthropod and nematode vectors of plant diseases. A. Rev. Plant Physiol., 19: 391-426.
Hartl, D.L., Dykhuizen, D.E., Miller, R.D., Green, L. and deFra- mond, J., 1983. Transposable element IS 50 improves growth rate of E. coli cells without transposition. Cell, 35: 503-510.
Hassall, M. Parkinson, D. and Visser, S., 1986a. Effects of the collembolan Onychiurus subtenuis on decomposition of Popu- lus tremuloides leaf litter. Pedobiologia, 29: 219-225.
Hassall, M., Visser, S. and Parkinson, D., 1986b. Vertical migra- tion of Onychiurus subtenuis Collembola in relation to rainfall and microbial activity. Pedobiologia, 29: 175-182.
Heal, O.W., 1962. The abundance and microdistribution of testate amoebae (Rhizopoda: Testacea) in Sphagnum. Oikos, 13: 35-47.
Heal, O.W., 1963. Soil fungi as food for amoebae. In: J. Doeksen and J. van der Drift (Editors). Soil Organisms. North-Holland, Amsterdam, pp. 289-297.
Heal, O.W. and Dighton, J., 1986. Nutrient cycling and decompo- sition in natural and terrestrial ecosystems. In: M.J. Mitchell and J.P. Nakas (Editors). Microfloral and Faunal Interactions in Natural and Agro-ecosystems. Nijhoff/Junk, Dordrecht, pp. 14-73.
Heck, M. and RiSmbke, J., 1990. Enchytraeiden Gemeinschaften Berliner Forstandorte. Zoologische Beitrage N.F., 33: 433- 458.
Henschke, R.B., Nucken, E. and Schmidt, F.R.J., 1989. Fate and dispersal of recombinant bacteria in a soil microcosm contain- ing the earthworm Lumbricus terrestris. Biol. Fertil. Soils, 7: 374-376.
Hepple, S., 1960. The movement of fungal spores in soil. Trans. Br. Mycol. Soc., 43: 73-79.
Herman, M., 1990. Effect of tillage systems and crop sequence on the rhizosphere microflora of winter wheat. Soil Till. Res., 15: 297-306.
Howie, W.J., Cook, R.J. and WeUer, D.M., 1987. Effects of soil matric potential and cell mobility on wheat root colonizaion by fluorescent pseudomonads suppressive to take-all. Phy- topathology, 79: 286-292.
Hughes, M.S., Bull, C.M. and Doube, B.M., 1996. Microcosm investigations into the influence of sheep manure on the behaviour of the geophagous earthworms Aporrectodea trape- zoides and Microscolex dubius. Biol. Fertil. Soils, 22: 71-75.
Huhta, V., Karppinen, E., Nurminen, N. and Valpas, A., 1967. Effect of silvicultural practises upon arthropod, annelid and nematode populations in coniferous forest soil. Ann. Zool. Fenn., 4: 87-143.
Ignoffo, C.M., Garcia, C., Hostetter, D.L. and Pinnell, R.E., 1977. Vertical movement of conidia of Nomuraea rileyi through sand and loam soils. J. Econ. Entomol., 70: 163-164.
Ingham, E.R., Coleman, D.C. and Moore, J.C., 1989. An analysis of food-web structure and function in a shortgrass prairie, a
mountain meadow, and a lodgepole pine forest. Biol. Fert. Soils, 8: 29-37.
Janke, G.D., Pratt, R.G., Arnold, J.D. and Odvody, G.N., 1983. Effects of deep tillage and roguing of diseased plants on oospore populations of Peronosclerospora sorghi in soil and on incidence of downy mildew in grain sorghum Sorghum bicolor. Phytopathology, 73: 1674-1678.
Jatala, P., Jensen, H.J. and Russell, S.A., 1974. Pristionchus theritieri as a carrier of Rhizobium japonicum. J. Nematol., 6: 130-131.
Kemp, J.S., Paterson, E., Gammack, S.M., Cresser, M.S. and Killham, K., 1992. Leaching of genetically modified Pseu- domonas fluorescens through organic soils; influence of tem- perature, soil pH and roots. Biol. Fert. Soils, 13: 218-224.
Kirkham, M.B., 1981. Earthworms and soil physical conditions: Darwin to the present, and future research needs. In: Work- shop on the Role of Earthworms in the Stabilization of Or- ganic Matter Residues, Vol 1. Beech Leaf Press, Kalamazoo, pp. 86-115.
Krogh, P.H., 1991. Perturbation of the soil microarthropod com- munity with the pesticides benomyl and isofenphos. Pedobi- ologia, 35: 71-88.
Lagerlof, J. and Andren, O., 1988. Abundance and activity of soil mites (Acari) in four cropping systems. Pedobiologia, 32: 129-145.
Laminger, H~ and Bucher, M., 1984. Feeding behavior of some terrestrial Testacea, (Protozoa: Rhizopoda). Pedobiologia, 27: 313-322.
Lance, J.C., Gerba, C.P. and Melnick, J.L., 1976. Virus movement in soil columns flooded with secondary sewage effluent. Appl. Environ. Microbiol., 32: 520-526.
Landry, E.F., Vaughn, J.M., Thomas, M.Z. and Beckwith, C.A., 1979. Adsorption of enteroviruses to soil cores and their subsequent elution by artificial rain water. Appl. Environ. Microbiol., 38: 680-687.
Latter, P.M. and Howson, G., 1978. Studies on the microfauna of blanket bog with particular reference to Enchytraeidae II. Growth and survival of Cognettia sphagnetorum on various substrates. J. Anim. Ecol., 47: 425-448.
Lavelle, P.B., Barois, I., Martin, A., Zaidi, Z. and Schaefer, R., 1989. Management of earthworm populations in agro-ecosys- tems: a possible way to maintain soil fertility? In: M. Clarholm and L. Bergstrom (Editors). Ecology of Arable Land: Perspec- tives and Challenges. Kluwer, Dordrecht, pp. 109-122.
Lee, K., 1985. Earthworms: Their Ecology and Relationships with Soil and Land use. Academic Press, London, 411 pp.
Lenski, R.E., 1987. The infectious spread of engineered genes. In: J.R.I. Fowle (Editor). Application of Biotechnology: Environ- mental and Policy issues. American Association for the Ad- vancement of Science, Washington DC, pp. 99-124.
Lenski, R.E. and Nguyen, T.T., 1988. Stability of recombinant DNA and its effects on fitness. In: J. Hodgson and A.M. Sugden (Editors). Planned Release of Genetically Engineered Organisms. Elsevier, Barking, pp. 18-20.
Lindow, S.E. and Panopoulos, N.J., 1988. Field tests of recombi- nant ice - - Pseudomonas syringae for biological frost control in potato. In: M. Sussman, C.H. Collins, F.A. Skinner and
J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131 129
D.E. Stewart-Tully (Editors). Release of Genetically-en- gineered Organisms. Academic Press, London, pp. 121-138.
Lipps, P.E., 1983. Survival of CoUetotrichum graminicola in infested corn Zea mays residues in Ohio USA. Plant Dis., 67: 102-104.
Liu, D., 1982. The effect of sewage sludge land disposal on the microbiological quality of ground water. Water Res., 16: 957-962.
Lousier, J.D., 1984. Population dynamics and production studies of species of Euglyphidae (Testacea, Rhizopoda, Protozoa) in an aspen woodland soil. Pedobiologia, 26: 309-330.
Luff, M.L., 1966. The abundance and diversity of the beetle fauna of grass tussocks. J. Anim. Ecol., 35: 189-208.
Lussenhop, J., 1981. Microbial and microarthropod detrital pro- cessing in a prairie soil. Ecology, 62: 964-972.
Lussenhop, J., 1992. Mechanisms of microarthropod-microbial interactions in soil. Adv. Ecol. Res., 23: 1-33.
Luxton, M., 1982. Biology of mites from beech woodland soil. Pedobiologia, 23: 1-8.
Lynch, J.M. and Elliott, L.F., 1983. Aggregate stabilisation of volcanic ash and soil during microbial degradation of straw. Appl. Environ. Microbiol., 45: 1398-1401.
Macfadyen, A., 1968. The animal habitat of soil bacteria. In: T.R.G. Gray and D. Parkinson (Editors). The Ecology of Soil Bacteria. Liverpool University Press, Liverpool, pp. 66-75.
Madsen, E.L. and Alexander, M., 1982. Transport of Rhizobium and Pseudomonas through soil. Soil Sci. Soc. Am. J., 46: 557-560.
Marshall, B., Flynn, D., Kamely, D. and Levy, S.B., 1988. Survival of Escherichia coli with and without ColEI::Tn5 after aerosol dispersal in a laboratory and farm environment. Appl. Environ. Microbiol., 54: 1776-1783.
Marshall, K.C., 1971. Sorptive interactions between soil particles and microorganisms. In: A.D. McLaren and J. Skujins (Edi- tors). Soil Biochemistry, Vol. 2. Marcel Dekker, NY, pp. 409-445.
Mather, J.H. and Christensen, O., 1988. Surface movement of earthworms in agricultural land. Pedobiologia, 32: 399-405.
Mawdsley, J.L., Bardgett, R.D., Merry, R.J., Pain, B.F. and Theodorou, M.K., 1995. Pathogens in livestock waste, their potential for movement through soil and environmental pollu- tion. Appl. Soil Ecol., 2: 1-15.
Meek, B.D., Rechel, E.R., Carter, L.M., DeTar, W.R. and Rieu, A.L., 1992. Infiltration rate of a sandy loam soil: effects of traffic, tillage and plant roots. Soil Sci. Soc. Am. J., 56: 908-913.
Mehrotra, R.S., 1970. Techniques for demonstrating accumulation of zoospores of Phytophthora species on roots in soil. Can. J. Bot., 48: 879-882.
Metting, Jr., F.B., 1992. Structure and physiological ecology of soil microbial communities. In: F.B. Metting, Jr. (Editor). Soil Microbial Ecology: Applications in Agricultural and Environ- mental Management. Dekker, NY, pp. 3-25.
Miller, R.V., 1988. Potential for transfer and establishment of engineered genetic sequences. In: J. Hodgson and A.M. Sug- den (Editor). Planned Release of Genetically Engineered Organisms. Elsevier, Oxford, pp. $23-27.
Miller, D.W., Clark, E.C., Palukaitis, P., Fox, J., Giddings, V., Hollander, A., McCormick, D., Mellon, M., Moolenar, R. and Suter, G., 1987. Control of genetically modified virnses. In: Prospects for the Physical and Biological Containment of Genetically-engineered Organisms. Cornell University Ecosys- tem Research Centre Report No ERC-114, NY.
Misaghi, I.J., Olsen, M.W., Billotte, J.M. and Sonoda, R.M., 1992. The importance of rhizobacterial mobility in biocontrol of bacterial wilt of tomato. Soil Biol. Biochem., 24: 287-293.
Moody, S.A., Briones, M.J.I., Piearce, T.G. and Dighton, J., 1995. Selective consumption of decomposing wheat straw by earth- worms. Soil Biol. Biochem., 27: 1209-1213.
Moody, S.A., Piearce, T.G. and Dighton, J., 1997. The fate of some fungal spores associated with wheat straw decomposi- tion on passage through the guts of Lumbricus terrestris and Apporectodea longa. Soil Biol. Biochem. (in press).
Mueller, B.R., Beare, M.H. and Crossley, D.AJ,, 1990. Soil mites in detrital food webs of conventional and no-tillage agroe- cosystems. Pedobiologia, 34: 389-401.
Murphy, S.L. and Tate, R.L., 1996. Bacterial movement through soil. In: G. Stotzky and J.M. Bollag (Editors). Soil Biochem., 9: 253-286.
Murray, P.M., Feest, A. and Madelin, M.F., 1985. The number of viable myxomycete cells in the alimentary tracts of earth- worms and in earthworm casts. Bot. J. Linn. Soc., 91: 359-366.
Overhoff, A., Schaefer-Pregl, R. and Roessner, J., 1991. Tillage effects on the population density of nematodes. Zeitschrift fur Kulturtechnik und Landentwicklung, 32: 106-113.
Parke, J.L., Moen, R., Rovira, A.D. and Bowen, G.D., 1986. Soil water flow affects the rhizosphere distribution of a seed-borne biological control agent Pseudomonas fluorescens. Soil Biol. Biochem., 18: 583-588.
Patni, N.K., Toxopeus, R., Tennant, A.D. and Hore, F.R., 1984. Bacterial quality of tile drainage water from manured and fertilized cropland. Water Res., 18: 127-132.
Peachey, J.E., 1963. Studies on the Enchytraeidae (Oligochaeta) of moorland soil. Pedobiologia, 2: 81-95.
Pedersen, J.C. and Hendriksen, N.B., 1993. Effects of passage through the intestinal track of detritivore earthworms (Lumbricus spp. ) on the number of selected Gram-negative and total bacterial. Biol. Fert. Soils, 16: 227-232.
Perdue, J.C. and Crossley, D.A., 1990. Seasonal abundance of soil mites (Acari) in experimental agroecosystems: effects of drought in no-tillage and conventional tillage. Soil Till. Res., 15: 117-124.
Petersen, H. and Luxton, M., 1982. A comparative analysis of soil fauna populations and their role in decomposition processes. Oikos, 39: 288-388.
Petz, W. and Foissner, W., 1988. Spatial separation of terrestrial ciliates and testaceans protozoa: a contribution to soil cil- iatostasis. Acta Protozool., 27: 249-258.
Pherson, D.A. and Beattie, A.J., 1979. Fungal loads of inverte- brates in beech leaf litter. Rev. Ecol. Biol. Sol, 16: 325-335.
Piearce, T.G., 1972. Acid intolerant and ubiquitous Lumbricidae in selected habitats in north Wales. J. Anim. Ecol., 41: 397- 410.
Pinkerton, J.N., Mojtahedi, H., Santo, G.S. and Obannon. J.J.,
130 J. Dighton et al. / Applied Soil Ecology 5 (1997) 109-131
1987. Vertical migration of Meloidogyne chitwoodi and M.hapla under controlled temperature. J. Nematol., 19: 152- 157.
Ponge, J.F., 1990. Ecological study of a forest humus by observ- ing a small volume 1. Penetration of pine litter by mycorrhizal fungi. Eur. J. For. Pathol., 20: 290-303.
Por, F.D. and Masry, D., 1968. Survival of a nematode and an oligochaete species in the anaerobic benthos of Lake Tiberias. Oikos, 19: 388-391.
Postma, J., van Veen, J.A. and Walter, S., 1989. Influence of different initial soil moisture contents on the distribution and population dynamics of introduced Rhizobium leguminosum biovar trifolii. Soil Biol. Biochem., 21: 437-442.
Powelson, D.K., Simpson, J.R. and Gerba, C.P., 1990. Virus transport and survival in saturated and unsaturated flow through soil columns. J. Environ. Qual., 19: 396-401.
Procter, D.L.C., 1990. Global overview of the functional roles of soil-living nematodes in terrestrial communities and ecosys- tems. J. Nematol., 22: 1-7.
Rabatin, S.C. and Stinner, B.R., 1988. Indirect effects of interac- tions between VAM fungi and soil-inhabiting invertebrates on plant processes. Agric. Ecosys. Environ., 24: 135-146.
Rahe, T.M., Hagedorn, C., McCoy, E.L. and Kling, G.F., 1978. Transport of antibiotic resistant Escherichia coli through western Oregon USA hill slope soils under conditions of saturated flow. J. Environ. Qual., 7: 487-494.
Reddell, P. and Spain, A.V., 1991. Earthworms as vectors of viable propagation of mycorrhizal fungi. Soil Biol. Biochem., 23: 767-774.
Robinson, C.H., Piearce, T.G., Ineson, P., Dickson, D.A. and Nys, C., 1992. Earthworm communities of limed coniferous soils: field observations and implications for forest management. For. Ecol. Manage., 55:117-134.
Ruddick, S.M. and Williams, S.T., 1972. Studies on the ecology of actinomycetes in soil. 5. Some factors influencing the dispersal and adsorption of spores in soil. Soil Biol. Biochem., 4: 93-103.
Rusek, J., 1975. Die Bodenbildende Funktion von Collembolen und Acarina. Pedobiologia, 15: 299-308.
Rusek, J., 1978. Pedozootische Sukzessionen wahrend der En- twicklung von Okosystemen. Pedobiologia, 18: 426-433.
Rutherford, P.M. and Juma, N.G., 1992a. Influence of soil texture on protozoa-induced mineralization of bacterial C and N. Can. J. Soil Sci., 72: 183-200.
Rutherford, P.M. and Juma, N.G., 1992b. Influence of texture on habitable pore space and bacterial-protozoan populations in soil. Biol. Fert. Soils, 12: 221-227.
Salas, B. and Stack, R.W., 1991. Influence of tillage on soil populations of Fusarium species in a spring wheat cropping system. Phytopathology, 81: 1215-1216.
Satchell, J.E., 1955. Some aspects of earthworm ecology. In: D.K. McE. Kevan (Editor). Soil Zoology. Butterworths, London, pp. 180-201.
Schaefer, M., 1982. The function of saprophagous animals of the soil of a limestone beech forest a long-term study program in the Goettingen forest west Germany. Drosera, 82: 75-84.
Scott Smith, M., Thomas, G.W. and White, R.E., 1983. Move-
ment of bacteria through macropores to ground water. Re- search Report No. 139, Water Resources Research Institute, Lexington, KY.
Shaw, C. and Pawluk, S., 1986. Faecal microbiology of Octola- sion tyrtaeum, Aporrectodea turgida and Lumbricus terrestris and its relation to the carbon budgets of three artificial soils. Pedobiologia, 29: 377-389.
Shew, H.D. and Beute, M.K., 1979. Evidence for the involvement of soil-borne mites in Pythium pod rot of peanut. Phytopathol- ogy, 69: 204-207.
Smith, T.F., 1978. A note on the effect of soil tillage on the frequency and vertical distribution of spores of vesicular ar- buscular endophytes. Aust. J. Soil Res., 16: 359-362.
Soby, S. and Bergman, K., 1983. Motility and chemotaxis of Rhizobium meliloti in soil. Appl. Environ. Microbiol., 46: 995-998.
Sohlenius, B., 1980. Abundance, biomass and contribution to energy flow by soil nematodes in terrestrial ecosystems. Oikos, 34: 186-195.
Sohlenius, B. and Sandor, A., 1989. Plowing of a perennial grass ley effect on the nematode fauna. Pedobiologia, 33: 199-210.
Standen, V., 1984. Production and diversity of enchytraeids, earthworms and plants in fertilized hay meadow plots. J. Appl. Ecol., 21: 293-312.
Stein, A., Bekker, R.M., Blom, J.H.C and Rogaar, H., 1992. Spatial variability of earthworm populations in a permanent polder grassland. Biol. Fert. Soils, 14: 260-266.
Swift, M.J., Heal, O.W. and Anderson, J.M., 1979. Decomposi- tion in Terrestrial Ecosystems. Studies in Ecology, Vol. 5. Blackwell Scientific, Oxford, 372 pp.
Takeda, H., 1978. Ecological studies of collembola populations in a pine forest soil II. Vertical distribution of Collembola. Pedobiologia, 18: 22-30.
Tan, Y., Bond, W.J., Rovira, A.D., Brisbane, P.G. and Griffin, D.M., 1991. Movement through soil of a biological control agent, Pseudomonas fluorescens. Soil Biol. Biochem., 23: 821-826.
Taub, F.B., 1992. Synthetic microcosms as test systems for sur- vival and effects of genetically-engineered microorganisms. In: M.A. Levin, R.J. Scidler and M. Regnal (Editors). Micro- bial Biology: Principles, Methods and Applications. McGraw- Hill, Maidenhead, pp. 643-660.
Tiedje, J.M., Colwell, R.K., Grossman, Y.L., Hodson, R.E., Lenski, R.E., Mack, R.N. and Regal, P.J., 1989. The planned introduction of genetically engineered organisms: ecological considerations and recommendations. Ecology, 70: 298-315.
Trevors, J.T., van Elsas, J.D., van Overbeek, L.S. and Starodub, M.E., 1990. Transport of genetically engineered Pseudomonas fluorescens strain through a soil microcosm. Appl. Environ. Microbiol., 56: 401-408.
Van der Drift, J., 1965. The effects of animal activity in the litter layer. In: E.G. Hallsworth and D.V. Crawford (Editors). Ex- perimental Pedology. Butterworths, London, pp. 227-235.
Vandevivere, P. and Baveye, P., 1992. Effect of bacterial extracel- lular polymers on the saturated hydraulic conductivity of sand columns. Appl. Environ. Microbiol., 58: 1690-1698.
Van Genuchten, M.T., 1978. Mass Transport in Saturated-un-
J. Dighton et al. /Applied Soil Ecology 5 (1997) 109-131 131
saturated Media: One Dimensional Solutions. Water Resources Program, Department of Civil Engineering. New Princeton University, NJ.
Van Genuchten, M.T., 1986. A Numerical Model for Water and Solute Movement In and Below the Root Zone; Model De- scription and User Manual. Draft Report US Salinity Labora- tory, U.S.D.A., Riverside, CA.
Vannier, G., 1975. Fundamental methods for studying the micro arthropods of the soil. Ann. Univ. Abidjan, Ser. C Sci., 11: 123-173.European Microbiological Societies) Microbiol. Ecol. and 74: 73-78.European Microbiological Societies) Microbiol. Ecol., 74: 73-78.QUERY ABOVE
Visser, S., 1985. Role of the soil invertebrates in determining the composition of soil microbial communities. In: A.H. Fitter, D. Atkinson, D.J. Read and M.B. Usher (Editors). Ecological Interactions in Soil: Plants, Microbes and Animals. Blackwell Scientific, Oxford, pp. 297-317.
Visser, S., Parkinson, D. and Hassall, M., 1987. Fungi associated with Onychiurus subtenuis (Collembola) in an aspen wood- land. Can. J. Bot., 65: 635-642.
Wallwork, J.A., 1967. Acari. In: A. Burgess and F. Raw (Editors). Soil Biology. Academic Press, London, pp. 363-395.
Wallwork, J.A., 1976. The Distribution and Diversity of Soil Fauna. Academic Press, London, 283 pp.
Wallwork, J.A. and Rodriguez, J.G., 1961. Ecological studies on oribatid mites with particular reference to their role as interme- diate hosts of Anoplocephalid cestodes, J. Econ. Entomol., 54: 701-705.
Wang, D.S., Gerba, C.P. and Lance, J.C., 1981. Effect of soil permeability on virus removal through soil columns. Appl. Environ. Microbiol., 42: 83-88.
Wang, Z., Crawford, D.L., Pometto, A.L. and Rafh, F., 1989. Survival and effects of wild-type, mutant and recombinant Streptomyces in a soil ecosystem. Can. J. Microbiol., 35: 535-543.
Wang, Z., Crawford, D.L., Magnuson, T.S., Bleakley, B.H. and Hertel, G., 1991. Effects of bacterial peroxidase on organic
carbon mineralisation in soil, using recombinant Streptomyces strains. Can. J. Microbiol., 37: 287-294.
Wellington, M.H., Cresswell, N. and Saunders, V.A., 1990. Growth and survival of Streptomycete inoculants and extent of plasmid transfer in sterile and non-sterile soil. Appl. Environ. Microbiol., 56:1413-1419
Wiebers, J.L., Hill, R.L. and Angel, J.S., 1995. Genetically engi- neered microbial amendments in soils. In: J.E. Recheigl (Edi- tor). Soil Amendments: Impacts on Biotic Systems. Lewis Publishers, Boca Raton, FL, pp. 471-490.
Wielemaker, W.G., 1984. Soil Formation by Termites: A Study in the Kisii Area, Kenya. Ph.D. Thesis, University of Wagenin- gen.
Wiggins, E.A. and Curl, E.A., 1979. Interactions of Collembola and microflora of cotton rhizosphere. Phytopathology, 69: 244-249.
Winstanley, C., Morgan, J.A.W., Pickup, R.W., Jones, J.G. and Saunders, J.R., 1989. Differential regulation of Lambda-PL and Lambda-PR by a CI repressor in a broad host-range thermoregulated plasmid marker system. Appl. Environ. Mi- crobiol., 55: 771-777.
Wong, P.T.W. and Griffin, D.M., 1976. Bacterial movement at high matrix potentials part II. In fungal colonies. Soil Biol. Biochem., 8: 219-223.
Yates, M.V. and Ouyang, Y., 1992. Virtus: a model of virus transport in unsaturated soils. Appl. Environ. Microbiol., 58: 1609-1616.
Yates, M.V. and Yates, S.R., 1991. Modelling microbial transport in the subsurface: a mathematical discussion. In: C.J. Hurst (Editor). Modelling the Environmental Fate of Microorgan- isms. American Society for Microbiology, Washington DC, pp. 48-76.
Yeates, G.W., 1984. Variation in soil nematode diversity under pasture with soil and year. Soil Biol. Biochem., 16: 95-102.
Zwart, K.B. and Darbyshire, J.F., 1992. Growth and nitrogenous excretion of a common soil Flagellate Spumella sp. A labora- tory experiment. J. Soil Sci., 43: 145-157.