Page 1
Testing diagnostic bioindicators in Prairie streams: Are biological traits and delta
15N of aquatic insects able to detect agricultural impacts?
by
Sophie Nicole Cormier
Bachelor of Science, University of New Brunswick, 2013
A Thesis Submitted in Partial Fulfillment
of the Requirements for the Degree of
Master of Science
in the Graduate Academic Unit of Biology
Supervisor: Joseph M. Culp, Ph.D., Biology
Examining Board: Alexa Alexander-Trusiak, PhD, Biology, Chair
Kerry T.B. MacQuarrie, PhD, Civil Engineering
This thesis is accepted by the
Dean of Graduate Studies
THE UNIVERSITY OF NEW BRUNSWICK
January, 2017
© Sophie Cormier 2017
Page 2
ii
ABSTRACT
Agricultural activities in the Red River watershed of Manitoba, Canada, can be
significant sources of excess nutrients, sediments and pesticides leading to ecological
effects in streams and downstream Lake Winnipeg. In such multiple stressor
environments, it is difficult to identify, separate and diagnose the cause of
environmental impacts from different agricultural activities using traditional methods
(e.g., taxa assemblage). However, ecological function indicators (e.g., functional feeding
groups) have potential as diagnostic indicators because they lead to the identification of
ecological change pathways. This study evaluated the efficacy of two indicators of
ecological function: biological traits and nitrogen isotopic signatures (δ15N) of benthic
macroinvertebrate. Indicator sensitivity was evaluated by their association with human
activity gradients that define the type and intensity of human activities (i.e., livestock,
wastewater lagoon discharge, crop production). Results indicated that biological traits
and δ15N of BMI were effective diagnostic bioindicators for small scale impacts (e.g.,
riparian condition) and point sources of stressors (e.g., wastewater discharge). However,
catchment scale agricultural activities were not associated with the bioindicators likely
because of hydrological factors affecting the timing of stressor transportation in these
prairie catchments. This study also demonstrated the importance of testing pathways of
human impacts based on conceptual models including the type and magnitude of
exposure to human activities and natural gradients.
Page 3
iii
DEDICATION
To the little being that experienced this process from within.
Page 4
iv
ACKNOWLEDGEMENTS
Completing this thesis would not have been possible without the help and advice
of many. First, I would like to thank my supervisor, Joseph Culp, for his wisdom,
patience, and his understanding that life gets in the way. Thank you to my committee
members who also provided invaluable support throughout this process: Wendy Monk,
and Patricia Chambers. I would also like to thank Adam Yates and Bob Brua for their
guidance, especially with the study design, and I am grateful for contributions from
Armin Namayandeh and Edward Krynak towards the development of the trait database.
I was lucky to have great technical assistance from colleagues at the Canadian
Rivers Institute, Environment Canada and the Yates lab. A special thank you goes to
Eric Luiker, Dave Hryn and Courtney Thompson for enduring field work in muddy
Manitoban streams with me. I am also thankful for logistical and technical support from
Kim Rattan and Zoey Duggan. In addition, I want to acknowledge the last minute field
assistance from Alistair Brown, Jacqueline Freeman, Alexandra Loeppky, Jezuele
Milanez, Matt Remple and Doug Watkinson. I would also like to thank Melanie
Deschènes, Kristie Heard, Craig Logan, Kirk Roach, and the SINLAB team for their
help with sample processing, as well as Linley Jesson, Alexa Alexander, Jennifer Lento,
Chris Tyrell, and Bruce Webb for taking the time to answer my questions and offer
advice.
Last but not least, I extend my gratitude to my family and close colleagues for
their encouragement and open ears, especially my parents, Katherine, Brianna,
Stephanie and Aleatha. I also thank my husband for his support, patience and
understanding.
Page 5
v
Table of Contents
ABSTRACT ..................................................................................................................... ii
DEDICATION ................................................................................................................ iii
ACKNOWLEDGEMENTS ........................................................................................... iv
Table of Contents .............................................................................................................v
List of Tables ................................................................................................................ viii
List of Figures ...................................................................................................................x
1.0 General introduction ..................................................................................................1
1.1 Background ............................................................................................................. 1
1.2 Diagnostic bioindicators.......................................................................................... 4
1.2.1 Structural vs. functional bioindicators ............................................................. 4
1.2.2 Functional bioindicators: biological traits and δ15N ........................................ 5
1.2.3 Testing bioindicators ........................................................................................ 7
1.3 Objectives ................................................................................................................ 8
1.4 Research significance ............................................................................................ 11
1.5 References ............................................................................................................. 12
2.0 Effects of agriculture on stream benthic macroinvertebrate community and
trait composition in Prairie streams .............................................................................19
2.1 Introduction ........................................................................................................... 19
Page 6
vi
2.2 Methods ................................................................................................................. 24
2.2.1 Study area ....................................................................................................... 24
2.2.2 BMI collection and trait assignment .............................................................. 25
2.2.3 Natural factors and HAGs .............................................................................. 27
2.2.4 Data analysis .................................................................................................. 29
2.3 Results ................................................................................................................... 31
2.3.1 Macroinvertebrate and trait assemblage structure.......................................... 31
2.3.2 Association with environmental variables ..................................................... 33
2.4 Discussion ............................................................................................................. 36
2.4.1 Drivers of trait and taxa beta diversity ........................................................... 36
2.4.2 Biological traits as bioindicators for the RRV ............................................... 40
2.4.3 Conclusions .................................................................................................... 42
2.5 References ............................................................................................................. 43
3.0 Δ15N as a tracer of anthropogenic nitrogen sources in Landscapes of Southern
Manitoba, Canada. .........................................................................................................70
3.1 Introduction ........................................................................................................... 70
3.2 Methods ................................................................................................................. 74
3.21 Study area and experimental design ................................................................ 74
3.22 Data analysis ................................................................................................... 76
3.3 Results ................................................................................................................... 78
Page 7
vii
3.4 Discussion ............................................................................................................. 79
3.4.1 Response of δ15N to point and non-point sources of nitrogen ....................... 79
3.4.2 δ15N of BMI and POM as bioindicator in the RRV ....................................... 83
3.4.3 Conclusion ..................................................................................................... 84
3.5 References ............................................................................................................. 85
4.0 General conclusion .................................................................................................101
4.1 Stated objectives.................................................................................................. 101
4.2 Drivers of trait and taxa beta diversity ................................................................ 101
4.3 Response of δ15N of BMIs to anthropogenic nitrogen ........................................ 103
4.4 Research implications ......................................................................................... 104
4.5 Conclusions and recommendations ..................................................................... 106
4.6 References ........................................................................................................... 110
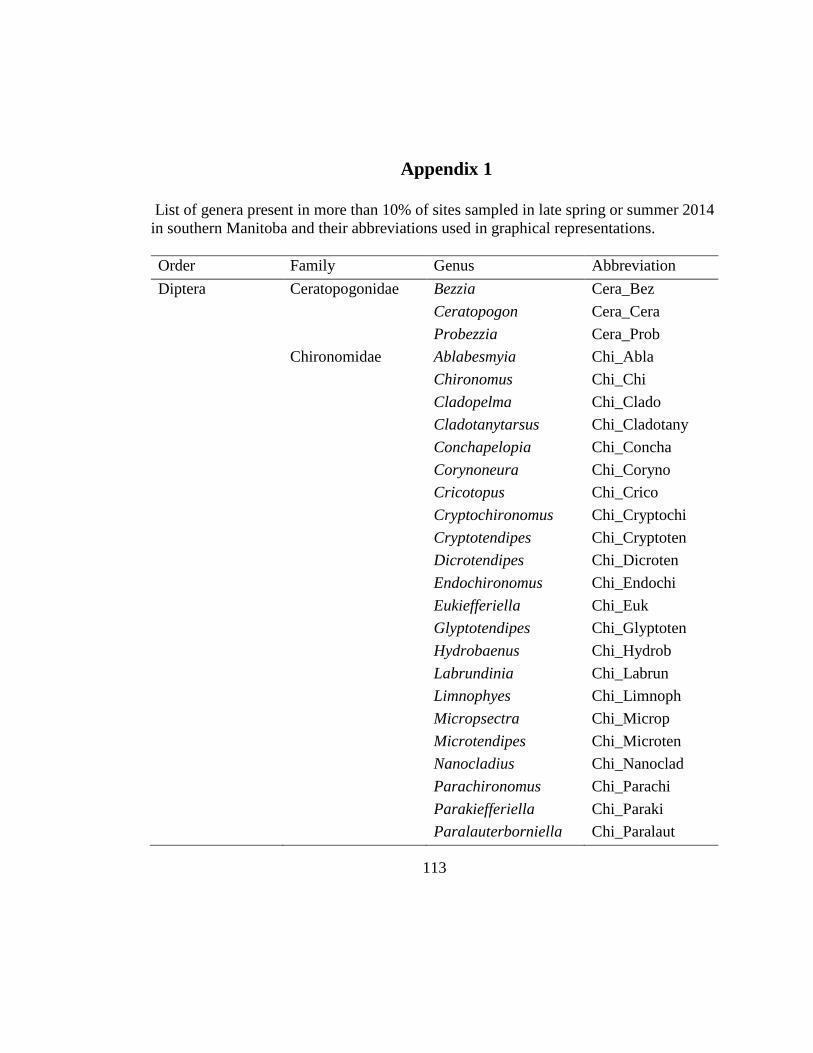
Appendix 1 ....................................................................................................................113
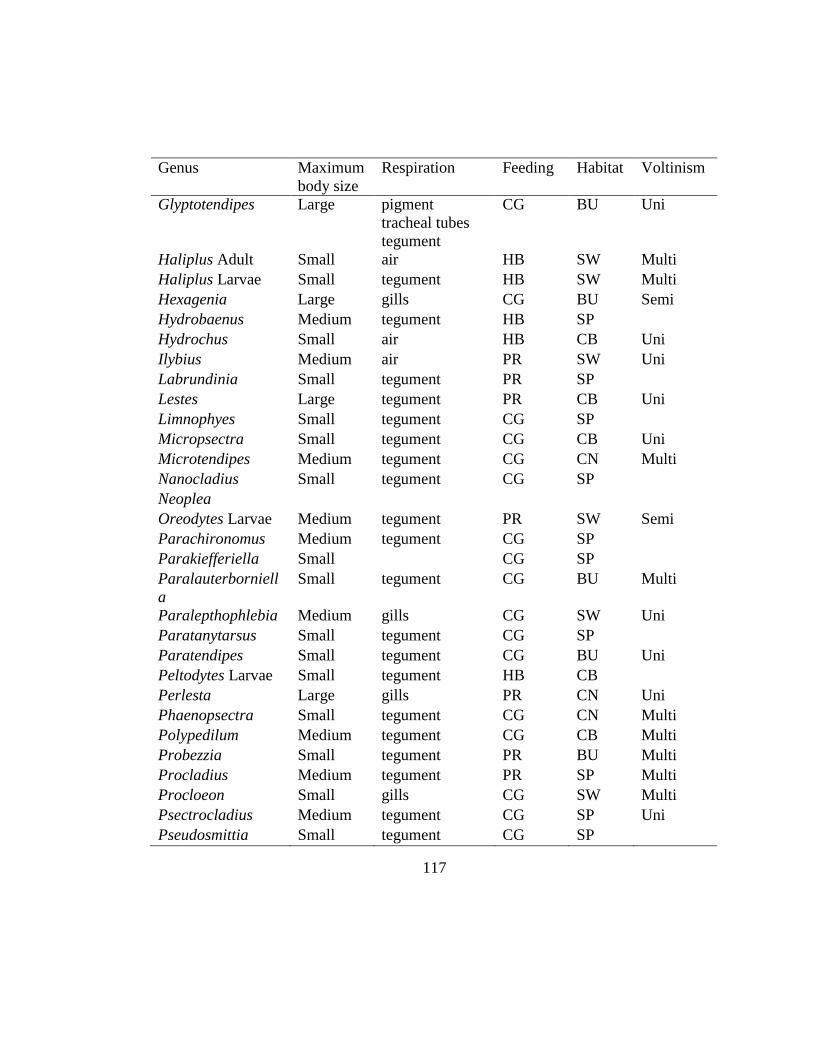
Appendix 2 ....................................................................................................................116
Appendix 3 ....................................................................................................................119
Appendix 4 ....................................................................................................................120
Curriculum Vitae
Page 8
viii
List of Tables
Table 2.1: Name and catchment description of 20 sites sampled in the Red River Valley
in southern Manitoba, Canada. .................................................................. 51
Table 2.2: Trait categories and trait states included in trait analyses of benthic
macroinvertebrate genera in southern Manitoba and their abbreviations (based on
Poff et al. 2006). ........................................................................................ 52
Table 2.3: Site and reach scale variables sampled in late spring (sp) or summer (su) 2014
for sites in the Red River Valley, southern Manitoba, Canada. Variables not used
in analyses were considered redundant based on Pearson correlation with other
variables of the same scale. ........................................................................ 53
Table 2.4: Catchment scale human activity included in each Principal Component
Analysis (PCA) to determine Human Activity Gradients in southern Manitoba,
Canada. High correlation was determined by a correlation coefficient higher than
0.8. .............................................................................................................. 54
Table 2.5: Distribution (% frequency of occurrence among 20 catchments) and
descriptive statistics of human activity in study catchments of the Red River
Valley, in southern Manitoba, Canada. ...................................................... 55
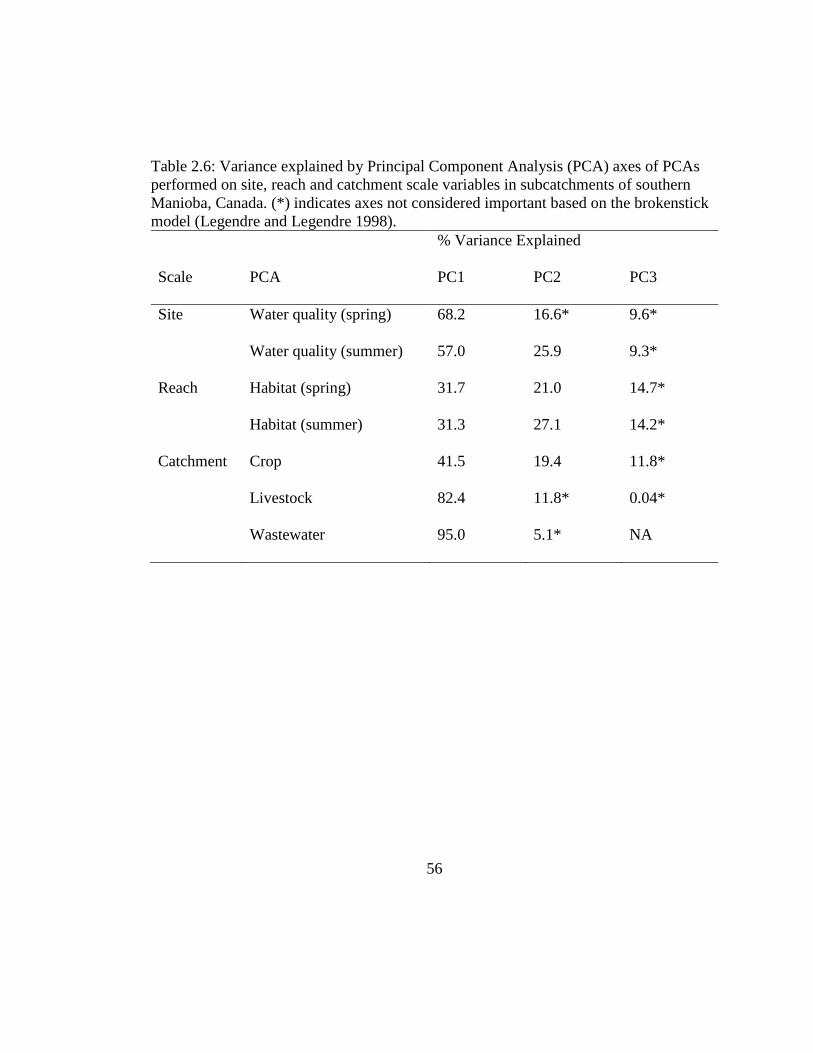
Table 2.6: Variance explained by Principal Component Analysis (PCA) axes of PCAs
performed on site, reach and catchment scale variables in subcatchments of
southern Manioba, Canada. (*) indicates axes not considered important based on
the brokenstick model (Legendre and Legendre 1998). ............................. 56
Table 2.7: Variance explained by axes (PC) of Principal Component Analyses of taxa or
trait assemblages sampled in spring or summer in southern Manitoba, Canada.
.................................................................................................................... 57
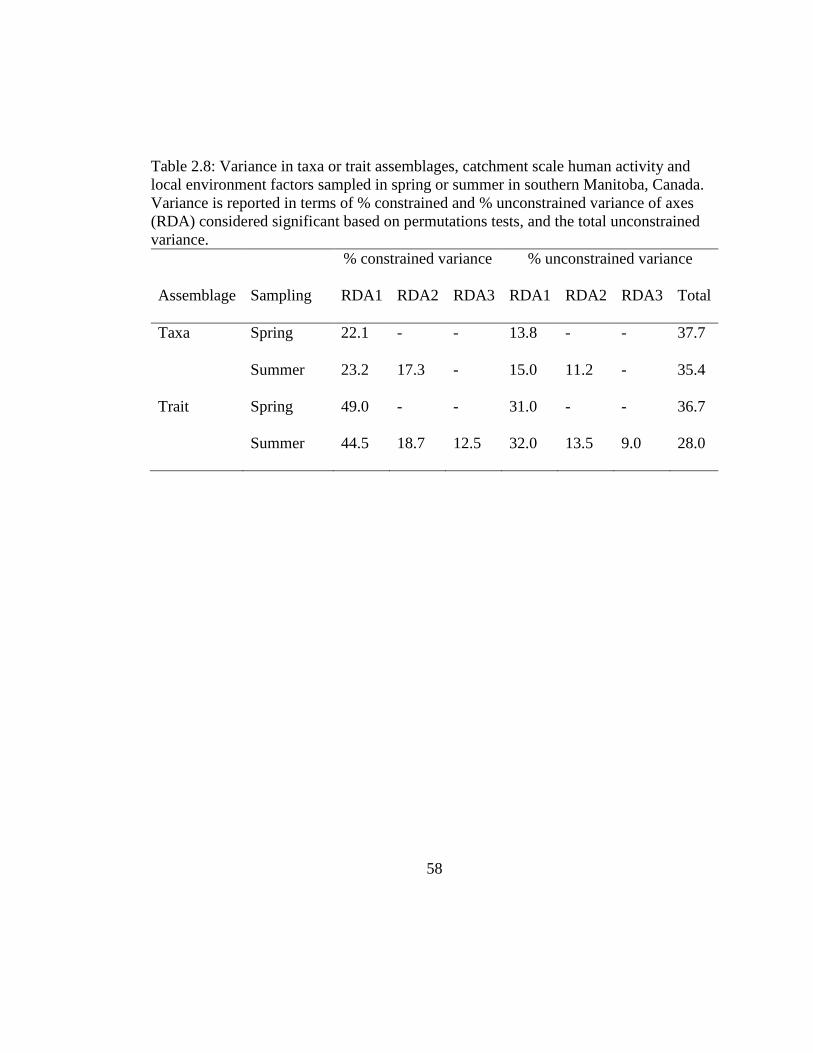
Table 2.8: Variance in taxa or trait assemblages, catchment scale human activity and
local environment factors sampled in spring or summer in southern Manitoba,
Canada. Variance is reported in terms of % constrained and % unconstrained
variance of axes (RDA) considered significant based on permutations tests, and
the total unconstrained variance. ................................................................ 58
Table 3.1: A priori hypotheses of how δ15N of particulate organic matter and primary
consumers may be affected by human activity gradients in subcatchments of the
Red River Valley of southern Manitoba, Canada. The ordinate intercept is
identified by (I). The PCA axes used as predictors are coded using “crop” (crop
types), “live” (type of livestock densities) and “wwt” (wastewater) with PC1
indicating the first axis or PC2, the second axis. ....................................... 90
Page 9
ix
Table 3.2: Descriptive statistics of δ15N values (‰) for particulate organic matter and
primary consumers sampled during both spring and summer in streams of
southern Manitoba, Canada. ....................................................................... 91
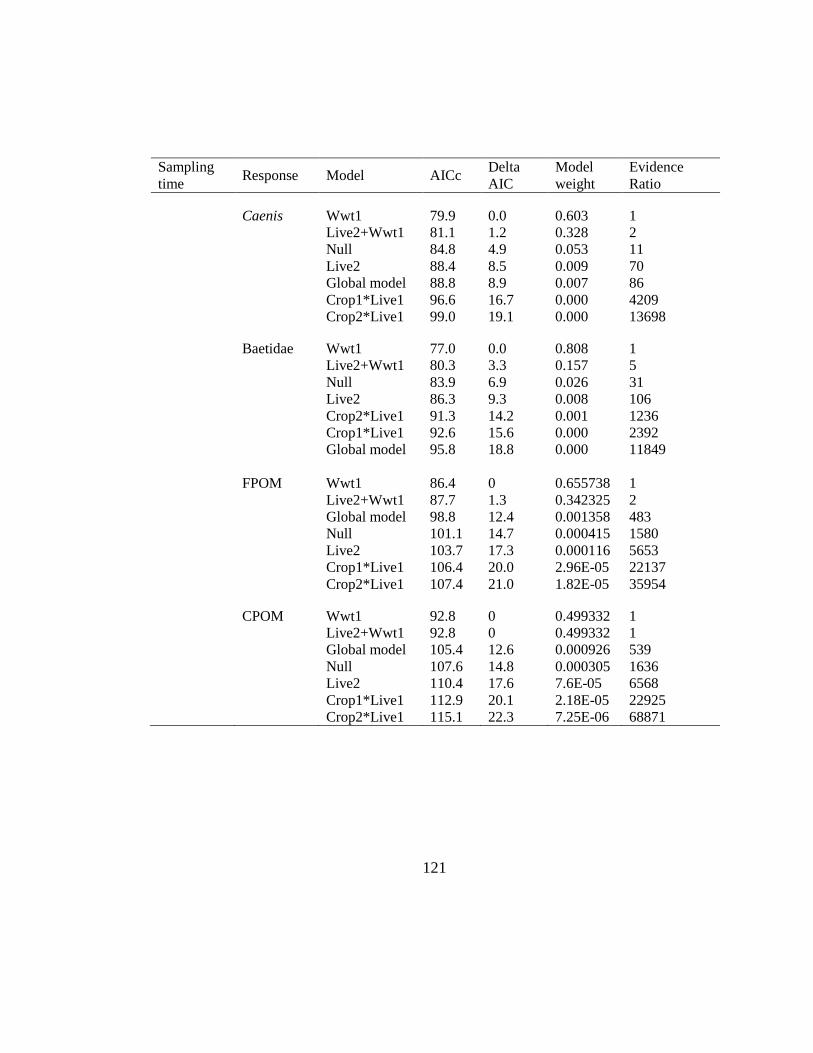
Table 3.3: Comparison of a priori tested models for predicting δ15N changes in primary
consumer taxa sampled in late spring for subcatchments in southern Manitoba,
Canada, using corrected Akaike Information Criterion (AICc). ................ 92
Table 3.4: Comparison of a priori tested models for predicting δ15N changes in primary
consumer taxa sampled in summer for subcatchments in southern Manitoba,
Canada, using corrected Akaike Information Criterion (AICc). ................ 93
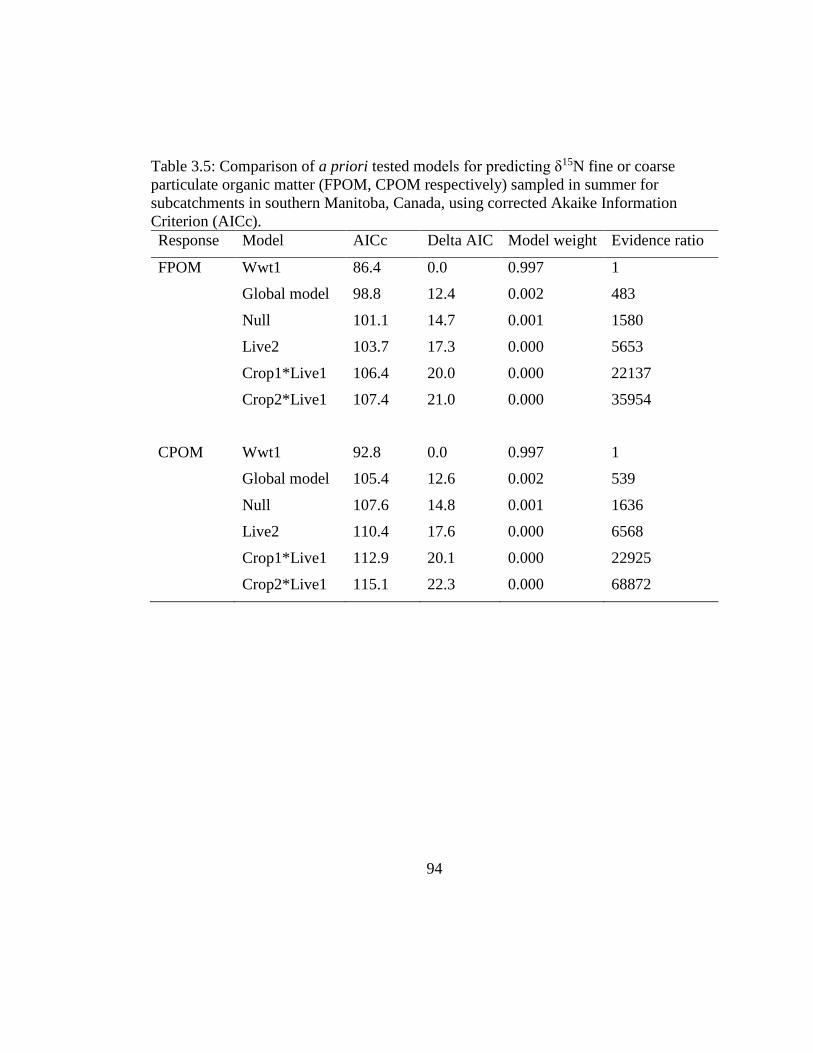
Table 3.5: Comparison of a priori tested models for predicting δ15N fine or coarse
particulate organic matter (FPOM, CPOM respectively) sampled in summer for
subcatchments in southern Manitoba, Canada, using corrected Akaike
Information Criterion (AICc). .................................................................... 94
Page 10
x
List of Figures
Figure 2.1: Conceptual diagram of broad mechanistic links between catchment scale
human activities, reach scale habitat, site scale water quality and trait categories
in freshwater streams.................................................................................. 59
Figure 2.2: Location of 20 sites and their subcatchments in southern Manitoba, Canada,
sampled in this study. Gray lines represent elevation contours. (Layers were
extracted from Geogratis.ca). ..................................................................... 60
Figure 2.3: Principal Component Analysis (PCA) of the type and intensity of a) crops,
b) livestock, and c) wastewater lagoons for 20 subcatchments in southern
Manitoba, Canada. Abbreviations are listed in Table 2.4. ......................... 61
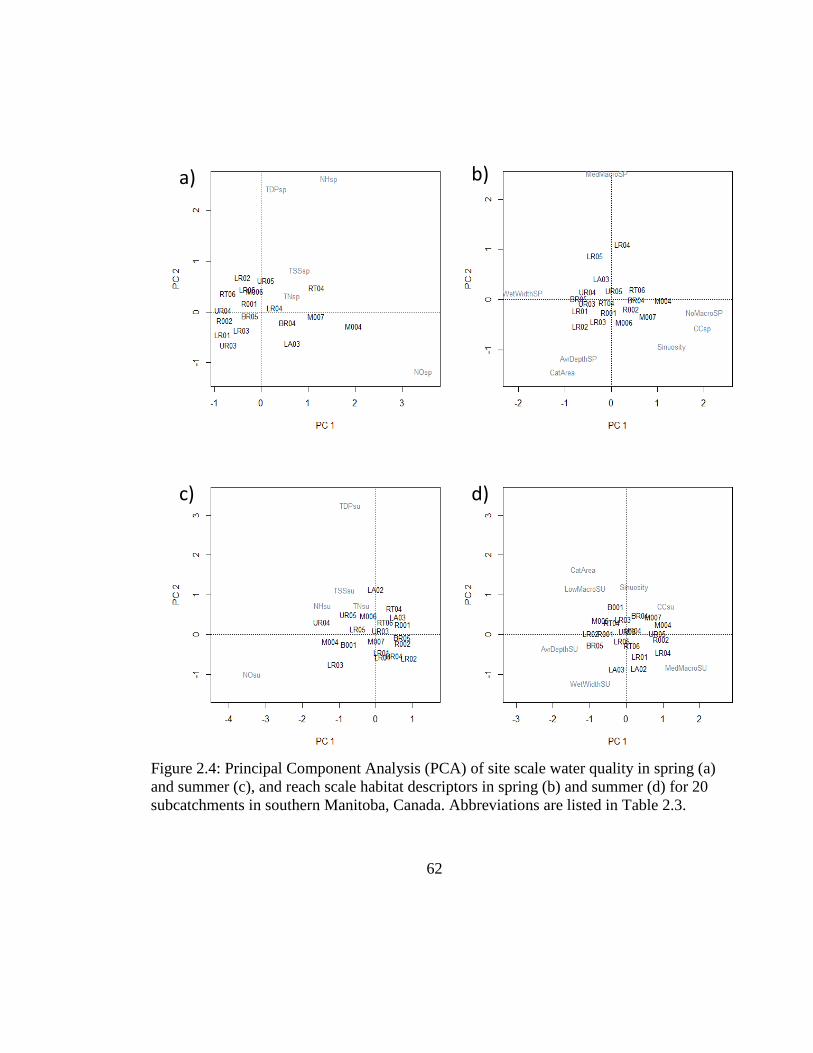
Figure 2.4: Principal Component Analysis (PCA) of site scale water quality in spring (a)
and summer (c), and reach scale habitat descriptors in spring (b) and summer (d)
for 20 subcatchments in southern Manitoba, Canada. Abbreviations are listed in
Table 2.3. .................................................................................................... 62
Figure 2.5: Principal Component Analyses (PCA) of taxa assemblage in spring (a) and
summer (b) (closed circles) measured at 18 (spring) or 20 (summer) study sites
(open circles) in southern Manitoba in 2014. Taxa abbreviation are listed in
Appendix 1. Taxa were labeled by order of importance to limit label overlaps.
.................................................................................................................... 63
Figure 2.6: Principal Component Analyses (PCA) of trait assemblage in spring (a) and
summer (b) summer assemblage (closed circles) sampled at 18 (spring) or 20
(summer) study sites (open circles) in southern Manitoba in 2014. Trait
abbreviation are listed in Table 2.2. ........................................................... 64
Figure 2.7: Redundancy Analysis (RDA) of presence-absence of aquatic insect genera
sampled in late spring 2014 in 18 sites of southern Manitoba, Canada. Taxa
abbreviations are listed in Appendix 1. ...................................................... 65
Figure 2.8: Redundancy Analysis (RDA) of presence-absence of aquatic insect genera
sampled in summer 2014 in 20 sites of southern Manitoba, Canada. Taxa
abbreviations are listed in Appendix 1. ...................................................... 66
Figure 2.9: Redundancy Analysis (RDA) of presence-absence of aquatic insect’ traits
sampled in late spring 2014 in 18 sites of southern Manitoba, Canada. Trait
abbreviations are listed in Table 2.2. ......................................................... 67
Page 11
xi
Figure 2.10: Redundancy Analysis (RDA) of presence-absence of aquatic insect’ traits
sampled in summer 2014 in 20 sites of southern Manitoba, Canada. Trait
abbreviations are listed in Table 2.2. ......................................................... 68
Figure 2.11: Redundancy Analysis (showing RDA2 and RDA3) of presence-absence of
aquatic insect’ traits sampled in summer 2014 in 20 sites of southern Manitoba,
Canada. Trait abbreviations are listed in Table 2.2. ................................... 69
Figure 3.1: Conceptual model of δ15N changes from landscape nitrogen sources to the
food web. (+) indicates an enrichment in δ15N, while (-) indicates a depletion.
Because the influence of both crops is dependent upon whether livestock manure
or synthetic fertilizer is the dominant source of nitrogen applied on the fields, the
relationship between crops and δ15N of the food web is difficult to predict. This
uncertainty is denoted by (?). ..................................................................... 95
Figure 3.2: Mean δ15N (± 1SD) of primary consumers and particulate organic matter
sampled during spring and summer 2014 in streams of southern Manitoba,
Canada. ....................................................................................................... 96
Figure 3.3: Relationship between the δ15N of all BMI and the wastewater HAG
(wwtPC1) sampled in spring or summer in subcatchments of southern Manitoba,
Canada. ....................................................................................................... 97
Figure 3.4: Relationship between the δ15N of particulate organic matter (fine, FPOM;
coarse, CPOM) and the wastewater HAG (wwtPC1) sampled in spring or
summer in subcatchments of southern Manitoba, Canada. ........................ 98
Figure 3.5: Conditional plots of δ15N of Hyalella in spring and (a) beef cattle production
(livePC2) while controlling for wwtPC1, or (b) wastewater HAG (wwtPC1)
while controlling for livePC2. Shaded areas represent confidence bands
(Breheny and Burchett 2013). .................................................................... 99
Figure 3.6: Conditional plot of δ15N of Coarse Particulate Organic Matter (CPOM) and
beef cattle production (livePC2) while controlling for wwtPC1 with (a) and
without (b) UR05. Shaded areas represent confidence bands (Breheny and
Burchett 2013). ......................................................................................... 100
Page 12
1
1.0 General introduction
1.1 Background
Agricultural land use alters the natural landscapes of watersheds and causes the
degradation of stream ecosystem health (Moss 2008). The major environmental stressors
associated with agriculture are excess nutrients, fine sediment and pesticides (Allan
2004), all of which can affect aquatic ecosystems directly (e.g., pesticide-related
mortality, Shulz and Liess 1999) and indirectly (e.g., altered herbivory rates; Delong and
Brusven 1998). Synthetic fertilizer applied on crops and manure from livestock
production causes increased nutrient inputs to streams through runoff (Carpenter et al.
1998, Chambers et al. 2009). This excess nutrient input can affect aquatic productivity
causing bottom-up effects through the food web, such as a change in invertebrate
density (Correll 1998, Rabalais 2002). Sediment inputs from tillage (Lobb et al. 2007),
livestock trampling (Kyriakeas and Watzin 2006) and clearing of riparian vegetation
(Meador and Goldstein 2003) can also have significant deleterious effects on stream
communities. For example, fine sediments may interfere with respiration or feeding
mechanisms (e.g., gills), reduce food source availability through smothering or light
reduction, and decrease the quality of benthic habitats (Wood and Armitage 1997,
Henley et al. 2000). Such impacts are difficult to quantify because of possible
Page 13
2
interactions among stressors (Townsend et al. 2008, Matthaei et al. 2010, Magbanua et
al. 2013), and are difficult to attribute to specific agricultural practices in large
watersheds because several types of crops and livestock production co-occur within the
landscape creating multiple stressor environments (Allan 2004).
The Red River Valley (RRV) in southern Manitoba, Canada, is an area with
intensive agriculture and, thus, a multiple stressor environment (Yates et al. 2012). This
watershed drains into Lake Winnipeg, the 10th largest lake in the world, and is the main
contributor of nutrients to the lake leading to increased algal blooms and related
ecosystem shifts (Jones and Armstrong 2001, Bunting et al. 2011, Schindler et al.
2012). The hyper-eutrophic state of Lake Winnipeg has serious repercussions for the
fisheries and tourism industries of the area (Environment Canada and Manitoba Water
Stewardship 2011) and has led to intensive research and monitoring to describe these
ecosystem changes in the hopes of developing appropriate mitigation measures
(Wassenaar and Rao 2012). As the stressors that affect the lake, most notably excess
nutrients, arise from land based activities in the RRV, managing these human activities
in the upstream watershed will be critical to address environmental changes in the lake
(Dodds and Oaks 2008, Schindler et al. 2012). An important step that still remains for
successful monitoring and mitigation of impacts is the development of monitoring tools
capable of diagnosing stressor-specific impacts to understand how to effectively manage
Page 14
3
each of these, and to test the effectiveness of management actions (Yates et al. 2012,
2014).
Indicators based on biota (i.e., bioindicators) have been widely used to describe
changes in the general health of ecosystems, but there is a need for developing
diagnostic bioindicators to better serve management and monitoring efforts (Downes
2010, Rapport and Hilden 2013). Bioindicators can integrate effects over longer
timeframes and have advantages over point samples of water quality (Cairns et al.
1993). A common group of organisms used as bioindicators are benthic
macroinvertebrates (BMI; Bonada et al. 2006). These organisms characterize the effects
of local environmental conditions because of their relatively limited dispersal and short
generation time (Cairns et al. 1993). Furthermore, BMI may be appropriate as
diagnostic indicators because they have diverse ecological roles (Wallace and Webster
1996) and stressor tolerance (Bonada et al. 2006), and thus may be useful in a multiple
stressor watershed such as the RRV. Therefore, the aim of this thesis is to use BMI to
develop diagnostic indicators relevant to agricultural activities in the RRV.
Page 15
4
1.2 Diagnostic bioindicators
1.2.1 Structural vs. functional bioindicators
Reliable diagnostic bioindicators need to detect specific stressor-effect
relationships based on a priori mechanistic hypotheses (Dale and Bayeler 2001, Bonada
et al. 2006, Downes 2010). Structural indicators, such as taxa metrics summarizing BMI
communities, have been extensively used to identify general degradation of stream
health related to human activities (Karr 1991, Cairns et al. 1993, Allan 2004). For
example, communities exposed to agricultural pollutants commonly experience a loss of
intolerant taxa resulting in decreased taxa evenness (Delong and Brusven 1998, Friberg
et al. 2003, Sutherland et al. 2012). However, it can be difficult to infer causation with
structural bioindicators in multiple stressor environments because they often respond in
a similar way to a wide range of environmental stressors (Dolédec et al. 2006, Palmer
and Febria 2012) and may include noise arising from biotic factors such as recruitment
and dispersal (Bunn and Davies 2000).
Indicators of ecological function describe ecosystem processes, such as nutrient
cycling (Dale and Beyeler 2001), and are considered to have greater diagnostic power
than structural indicators because they are more directly linked to pathways of
ecological change (Palmer and Febria 2012). Furthermore, because simple but powerful
predictions about specific processes can be tested using functional indicators, it is
Page 16
5
possible to infer causal relationships and better isolate the effects of independent
stressors (Bunn and Davies 2000, Culp et al. 2010). Thus, it is not surprising that
functional indicators commonly demonstrate a stronger response to stressor gradients
than structural indicators (Sandin and Solimini 2009). Functional indicators may also be
better suited for studying impacts in large watersheds because they provide more
consistent relationships to human impacts over large spatial scales (Bêche et al. 2006,
Pollard and Yuan 2010, Dolédec et al. 2011). Two functional bioindicators are
considered in this thesis based on recommendations by Yates et al. (2012, 2014),
namely biological traits and the nitrogen isotopic signature of BMI (δ15N).
1.2.2 Functional bioindicators: biological traits and δ15N
Biological traits include characteristics of organisms associated with adaptations
to their environment (e.g., functional feeding groups, maximum body size, respiration,
locomotion). Traits have the potential to be good diagnostic bioindicators in multiple
stressor environments because their relationship with pathways of environmental change
are predictable (Poff 1997) and they often respond independently to different stressors
(Townsend and Hildrew 1994, Poff 1997, Culp et al. 2010, Statzner and Bêche 2010).
For example, Lange et al. (2014) found that life-history traits (e.g., life duration of
adults) of BMI in New Zealand streams were associated with farming intensity, while
feeding habitats and respiration traits were indicative of water abstraction. Although this
Page 17
6
approach does not consider variation of traits within individual organisms (e.g.,
maximum body size of taxa based on the literature vs. average body size of individuals
collected), aggregating species to create biological trait categories simplifies the study of
multi-species assemblages (Poff 1997) while retaining more information than composite
indexes (Statzner and Bêche 2010). In addition, by only including traits that can be
mechanistically linked to the stressor of interest, it is possible to avoid confounding
interpretation from traits that are evolutionary linked while strengthening the ability to
discriminate stressors (Poff et al. 2006, Statzner and Bêche 2010).
In addition to biological traits, the δ15N signature of BMI, a stable isotope
approach, is a promising diagnostic bioindicator because it may allow identification of
independent sources of anthropogenic nitrogen (Anderson and Cabana 2005, Niemi et
al. 2011, Winemiller et al. 2011, Clapcott et al. 2012). Human activities produce
nitrogen with different δ15N signatures. For example, synthetic fertilizers often have low
nitrogen isotopic values compared to manure or wastewater (Bedard-Haughn et al.
2003). When these nitrogen inputs with relatively unique δ15N signatures reach the
aquatic environment and are assimilated within the food web, they change the
proportion of heavy to light nitrogen isotopes in tissues of primary producers and
consumers in a predictable manner (Peterson and Fry 1987). Moreover, because the
δ15N signature of BMI integrates temporal variation in exposure to excess nitrogen, it
Page 18
7
may help infer the source in addition to the magnitude of nitrogen assimilation from
human sources in the food chain (Bedard-Haughn et al. 2003).
1.2.3 Testing bioindicators
Because bioindicators can be affected by natural factors in addition to human
impacts, the effectiveness of the proposed bioindicators must be tested in the RRV prior
to implementing them in monitoring programs. For example, natural variation unrelated
to human activities, such as catchment geology, may affect traits independently and
confound conclusions about human impacts (Poff 1997, Yates and Bailey 2010, Heino
et al. 2013). In addition, local factors relating to nitrogen transformation and cycling
affect the natural δ15N baseline and the δ15N of anthropogenic sources, thus
necessitating local testing and validation to ensure the effectiveness of this tool in
identifying specific nitrogen sources (Bedard-Haughn et al. 2003, Diebel and Vander
Zanden 2009). Interpreting changes in biological traits or δ15N signature of BMI based
on associations from other geographical regions may result in falsely attributing
differences to human impacts.
A key characteristic of a successful diagnostic bioindicator is that it exhibits a
known and consistent response to specific anthropogenic stressors (Dale and Beyeler
2001, Bonada et al. 2007). Thus, a multiple catchment design has been proposed as a
powerful approach for testing the effectiveness of bioindicators over gradients of
Page 19
8
exposure to human activities (Yates and Bailey 2007). Bailey et al. (2007) proposed the
use of conceptual models, representing the pathways of effects in response to specific
human land use, to enable inferences of causal relationships between bioindicators and
human activities. Quantifying exposure to human activities has often involved using
coarse descriptions of land use (e.g., % agricultural land cover; Allan 2004), but these
may not be appropriate for diagnostic studies in catchments with intensive agriculture
because catchments with a similar extent of agriculture may differ in specific
agricultural practices, the connectivity of agricultural land with the stream, and the
consequent ecological effects (e.g., unequal contribution of nutrients from different crop
types; Yates and Bailey 2010). Instead, Human Activity Gradients (HAGs) that quantify
the type and magnitude of specific human activities (e.g., types of crops such as small
grain production) have been shown to be an effective and comprehensive measure of
exposure to anthropogenic stressors in multiple stressor watersheds (Yates et al. 2012).
Natural gradients can be included to support hypotheses about the pathway of effects of
human impacts (Bailey et al. 2007) and to identify possible covariation between human
impacts and unrelated natural variation (Yates and Bailey 2006).
1.3 Objectives
This thesis aims to determine the utility of two bioindicators based on BMI
assemblages (i.e., biological traits and δ15N signatures) to diagnose human impacts in
Page 20
9
the agricultural watershed of the RRV by testing their response along HAGs as
described by Yates et al. (2012). Previous work in southern Manitoba revealed that the
δ15N signature of BMI may be a good diagnostic bioindicator for the region because of
relationships between some human activities and primary producers (Yates et al. 2014).
However, the potential of biological traits of BMI to be used as diagnostic bioindicators
has not been tested in the RRV.
The first objective, examined in Chapter 2, compared the sensitivity of biological
traits and community structure of BMI to specific human activities with the goal of
determining if traits have better diagnostic power than the structural approach, as
predicted by the literature. It is expected that a trait-based approach is more sensitive to
stressors of interest because of a mechanistic linkage with pathways of interest, greater
seasonal stability and consistency across broad spatial scales when compared to a
taxonomic approach (Culp et al. 2010). The trait-based approach included the
formulation of a priori mechanistic hypotheses based on a conceptual diagram
describing predicted relationships among human activities, local habitat and biological
traits (Figure 2.1). It was predicted that: (1) high exposure to pesticides favors traits that
promote resilience to stress, more specifically smaller body size and short generation
time (Townsend and Hildrew 1994, Liess et al. 2005); (2) excess nutrients increases
herbivores, shredders and/or collector-filterers if accompanied with higher food
Page 21
10
availability and less tegument respiration if the effect decreases dissolved oxygen
concentrations (Compin and Céréghino 2007, Carlisle and Hawkins 2008, Feio and
Dolédéc 2012); and (3) excess fine sediment decreases the occurrence of traits such as
aquatic respiration, filter feeding, scraping and shredding because of the interference
from sediment particles or reduction in food source availability, coupled with an
increase in burrowers which prefer fine sediment bottoms (Rabeni et al. 2005,
Magbanua et al. 2013).
The second objective, explored in Chapter 3, determined whether the δ15N
signature of BMI in the RRV could be used to identify sources of anthropogenic
nitrogen in streams of the RRV. Primary consumers were chosen as the food web
component to evaluate δ15N as a potential tracer of nitrogen sources in order to limit the
influence of fractionation due to trophic position within a food web (Peterson and Fry
1987). In addition, the δ15N signature of particulate organic matter (POM) was included
to represent the base of the food web. A conceptual model describing the predicted
influence of different anthropogenic nitrogen sources on the baseline δ15N assimilated in
the food web was used to develop hypotheses (Figure 3.1). This model is based on
literature observations and predicts that point sources of anthropogenic nitrogen, such as
livestock feedlots and wastewater lagoons, would enrich δ15N of POM and BMI because
both manure and wastewater discharge tend to have high δ15N compared to the natural
Page 22
11
baseline (Bedard-Haughn et al. 2003, Anderson and Cabana 2006, Diebel and Vander
Vander Zanden 2009). Furthermore, the relationship between cropland and δ15N of BMI
in southern Manitoba was predicted to be dependent on the dominant fertilizer, synthetic
or manure, used in a particular catchment. In contrast to manure that has a high δ15N,
synthetic fertilizers tend to have a low δ15N compared to the natural baseline (Bedard-
Haughn et al. 2003). Because information about the amount and type of fertilizer
applied in catchments of the study area was not available, more specific predictions
about the influence of nitrogen from crop runoff could not be formulated.
1.4 Research significance
Indicator and baseline studies, such as the current thesis research, are an
important step toward understanding the effects of agriculture in prairie ecosystems and
improving diagnostic techniques in multiple stressor watersheds. This thesis will
contribute to research for the Tobacco Creek Model Watershed Research Consortium
funded by the Canadian Water Network in its goal of developing localized monitoring
frameworks that can be used to monitor the efficiency of new regulations and best
management practices. In addition, the study contributes to the Lake Winnipeg Basin
Initiative, led by Environment Canada, by assessing the suitability of nutrient
monitoring using aquatic animals as indicators.
Page 23
12
The thesis is written in manuscript format and consequently, there is some
redundancy between this section and the introduction of the next two chapters.
1.5 References
Allan J.D. (2004) Landscapes and riverscapes: the influence of land use on stream
ecosystems. Annual Review of Ecology, Evolution, and Systematics 35, 257–284.
Anderson C. & Cabana G. (2005) 15 N in riverine food webs: effects of N inputs from
agricultural watersheds. Canadian Journal of Fisheries and Aquatic Sciences 340,
333–340.
Anderson C. & Cabana G. (2006) Does delta 15N in river food webs reflect the intensity
and origin of N loads from the watershed? The Science of the total environment
367, 968–978.
Bailey R.C., Reynoldson T.B., Yates A.G., Bailey J. & Linke S. (2007) Integrating
stream bioassessment and landscape ecology as a tool for land use planning.
Freshwater Biology 52, 908–917.
Bêche L.A., McElravy E.P. & Resh V.H. (2006) Long-term seasonal variation in the
biological traits of benthic-macroinvertebrates in two Mediterranean-climate
streams in California, U.S.A. Freshwater Biology 51, 56–75.
Bedard-Haughn A., van Groenigen J.W. & van Kessel C. (2003) Tracing 15N through
landscapes: potential uses and precautions. Journal of Hydrology 272, 175–190.
Bonada N., Prat N., Resh V.H. & Statzner B. (2006) Developments in aquatic insect
biomonitoring: a comparative analysis of recent approaches. Annual review of
entomology 51, 495–523.
Bonada N., Rieradevall M. & Prat N. (2007) Macroinvertebrate community structure
and biological traits related to flow permanence in a Mediterranean river network.
Hydrobiologia 589, 91–106.
Page 24
13
Bunn S. & Davies P. (2000) Biological processes in running waters and their
implications for the assessment of ecological integrity. Hydrobiologia 422, 61–70.
Bunting L., Leavitt P., Wissel B., Laird K.R., Cumming B.F., St. Amand A., et al.
(2011) Sudden ecosystem state change in Lake Winnipeg, Canada, caused by
eutrophication arising from crop and livestock production during the 20th century.
Winnipeg, Canada.
Cairns J., McCormick P. V. & Niederlehner B.R. (1993) A proposed framework for
developing indicators of ecosystem health. Hydrobiologia 263, 1–44.
Carlisle D.M. & Hawkins C.P. (2008) Land use and the structure of western US stream
invertebrate assemblages: predictive models and ecological traits. Journal of the
North American Benthological Society 27, 986–999.
Carpenter S.R., Caraco N.F., Correll D.., Howarth R.W., Sharpley A.N. & Smith V.H.
(1998) Nonpoint pollution of surface waters with phosphorus and nitrogen.
Ecological Applications 8, 559–568.
Chambers P.A., M. G., Dixit S.S., Benoy G.A., Brua R.B., Culp J.M., et al. (2009)
Nitrogen and phosphorus standards to protect the ecological condition of
Canadian streams, rivers and coastal waters. National Agri-Environment
Standards Initiative Synthesis Report No. 11. Environment Canada, Gatineau,
Quebec.
Clapcott J.E., Collier K.J., Death R.G., Goodwin E.O., Harding J.S., Kelly D., et al.
(2012) Quantifying relationships between land-use gradients and structural and
functional indicators of stream ecological integrity. Freshwater Biology 57, 74–90.
Compin A. & Céréghino R. (2007) Spatial patterns of macroinvertebrate functional
feeding groups in streams in relation to physical variables and land-cover in
Southwestern France. Landscape Ecology 22, 1215–1225.
Correll D.L. (1998) The role of phosphorus in the eutrophication of receiving waters: a
review. Journal of Environment Quality 27, 261-266.
Page 25
14
Culp J.M., Armanini D.G., Dunbar M.J., Orlofske J.M., Poff N.L., Pollard A.I., et al.
(2010) Incorporating traits in aquatic biomonitoring to enhance causal diagnosis
and prediction. Integrated environmental assessment and management 7, 187–197.
Dale V.H. & Beyeler S.C. (2001) Challenges in the development and use of ecological
indicators. Ecological Indicators 1, 3–10.
Delong M.D. & Brusven M.A. (1998) Macroinvertebrate community structure along the
longitudinal gradient of an agriculturally impacted stream. Environmental
management 22, 445–457.
Diebel M.W. & Vander Zanden M.J. (2009) Nitrogen stable isotopes in streams: effects
of agricultural sources and transformations. Ecological applications 19, 1127–
1134.
Dodds W.K. & Oakes R.M. (2008) Headwater Influences on Downstream Water
Quality. Environmental Management 41, 367–377.
Dolédec S., Phillips N., Scarsbrook M., Riley R.H. & Townsend C.R. (2006)
Comparison of structural and functional approaches to determining landuse effects
on grassland stream invertebrate communities. Journal of the North American
Benthological Society 25, 44–60.
Dolédec S., Phillips N. & Townsend C.R. (2011) Invertebrate community responses to
land use at a broad spatial scale: trait and taxonomic measures compared in New
Zealand rivers. Freshwater Biology 56, 1670–1688.
Downes B.J. (2010) Back to the future: little-used tools and principles of scientific
inference can help disentangle effects of multiple stressors on freshwater
ecosystems. Freshwater Biology 55, 60–79.
Environment Canada & Manitoba Water Stewardship (2011) State of Lake Winnipeg:
1999 to 2007. Winnipeg, MB.
Feio M.J. & Dolédec S. (2012) Integration of invertebrate traits into predictive models
for indirect assessment of stream functional integrity: A case study in Portugal.
Ecological Indicators 15, 236–247.
Page 26
15
Friberg N., Lindstrøm M., Kronvang B. & Larsen S.E. (2003)
Macroinvertebrate/sediment relationships along a pesticide gradient in Danish
streams. Hydrobiologia 494, 103–110.
Heino J., Schmera D. & Erős T. (2013) A macroecological perspective of trait patterns
in stream communities. Freshwater Biology 58, 1539–1555.
Henley W.F., Patterson M.A., Neves R.J. & Lemly A.D. (2000) Effects of sedimentation
and turbidity on lotic food webs: a concise review for natural resource managers.
Reviews in Fisheries Science 8, 125–139.
Jones G. & Armstrong N. (2001) Long-term trends in total nitrogen and total
phosphorus concentrations in Manitoba streams. Winnipeg, MB.
Karr J.R. (1991) Biological integrity : a long-neglected aspect of water resource
management. Ecological Applications 1, 66–84.
Kyriakeas S.A. & Watzin M.C. (2006) Effects of adjacent agricultural activities and
watershed characteristics on stream macroinvertebrate communities. Journal of the
American Water Resources Association 42, 425–441.
Liess M. & Von Der Ohe P.C. (2005) Analyzing effects of pesticides on invertebrate
communities in streams. Environmental Toxicology and Chemistry 24, 954.
Lobb D.A., Huffman E. & Reicosky D.C. (2007) Importance of information on tillage
practices in the modelling of environmental processes and in the use of
environmental indicators. Journal of Environmental Management 82, 377–387.
Magbanua F.S., Townsend C.R., Hageman K.J. & Matthaei C.D. (2013) Individual and
combined effects of fine sediment and the herbicide glyphosate on benthic
macroinvertebrates and stream ecosystem function. Freshwater Biology 58, 1729–
1744.
Matthaei C.D., Piggott J.J. & Townsend C.R. (2010) Multiple stressors in agricultural
streams: interactions among sediment addition, nutrient enrichment and water
abstraction. Journal of Applied Ecology 47, 639–649.
Page 27
16
Meador M.R. & Goldstein R.M. (2003) Assessing water quality at large geographic
scales: Relations among land use, water physicochemistry, riparian condition, and
fish community structure. Environmental Management 31, 504–517.
Moss B. (2008) Water pollution by agriculture. Philosophical transactions of the Royal
Society of London. Series B, Biological sciences 363, 659–666.
Niemi G.J., Reavie E.D., Peterson G.S., Kelly J.R., Johnston C.A., Johnson L.B., et al.
(2011) An integrated approach to assessing multiple stressors for coastal Lake
Superior. Aquatic Ecosystem Health & Management 14, 356–375.
Palmer M.A. & Febria C.M. (2012) Ecology. The heartbeat of ecosystems. Science 336,
1393–1394.
Peterson B.J. & Fry B. (1987) Stable isotopes in ecosystem studies. Annual Review of
Ecology and Systematics 18, 293–320.
Poff N.L. (1997) Landscape filters and species traits: towards mechanistic understanding
and prediction in stream ecology. Journal of the North American Benthological
Society 16, 391–409.
Poff N.L., Olden J.D., Vieira N.K.M., Finn D.S., Mark P., Kondratieff B.C., et al.
(2006) Functional trait niches of North American lotic insects: traits-based
ecological applications in light of phylogenetic relationships. Journal of the North
American Benthological Society 25, 730–755.
Pollard A.I. & Yuan L.L. (2010) Assessing the consistency of response metrics of the
invertebrate benthos: a comparison of trait- and identity-based measures.
Freshwater Biology 55, 1420–1429.
Rabalais N.N. (2002) Nitrogen in aquatic ecosystems. Ambio 31, 102–112.
Rabení C.F., Doisy K.E. & Zweig L.D. (2005) Stream invertebrate community
functional responses to deposited sediment. Aquatic Sciences 67, 395–402.
Rapport D.J. & Hildén M. (2013) An evolving role for ecological indicators: from
documenting ecological conditions to monitoring drivers and policy responses.
Ecological Indicators 28, 10–15.
Page 28
17
Sandin L. & Solimini A.G. (2009) Freshwater ecosystem structure-function
relationships: from theory to application. Freshwater Biology 54, 2017–2024.
Schindler D.W., Hecky R.E. & McCullough G.K. (2012) The rapid eutrophication of
Lake Winnipeg: greening under global change. Journal of Great Lakes Research
38, 6–13.
Schulz R. & Liess M. (1999) A field study of the effects of agriculturally derived
insecticide input on stream macroinvertebrate dynamics. Aquatic Toxicology 46,
155–176.
Statzner B. & Bêche L.A. (2010) Can biological invertebrate traits resolve effects of
multiple stressors on running water ecosystems? Freshwater Biology 55, 80–119.
Sutherland A.B., Culp J.M. & Benoy G.A. (2012) Evaluation of deposited sediment and
macroinvertebrate metrics used to quantify biological response to excessive
sedimentation in agricultural streams. Environmental management 50, 50–63.
Townsend C.R. & Hildrew A.G. (1994) Species traits in relation to a habitat templet for
river systems. Freshwater Biology 31, 265–275.
Townsend C.R., Uhlmann S.S. & Matthaei C.D. (2008) Individual and combined
responses of stream ecosystems to multiple stressors. Journal of Applied Ecology
45, 1810–1819.
Wallace J.B. & Webster J.R. (1996) The role of macroinvertebrates in stream ecosystem
function. Annual Review of Entomology 41, 115–139.
Wassenaar L.I. & Rao Y.R. (2012) Lake Winnipeg: the forgotten great lake. Journal of
Great Lakes Research 38, 1–5.
Winemiller K.O., Hoeinghaus D.J., Pease A.A., Esselman P.C., Honeycutt R.L.,
Gbanaador D., et al. (2011) Stable isotope analysis reveals food web structure and
watershed impacts along the fluvial gradient of a mesoamerican coastal river. River
Research and Application 27, 791–803.
Wood P. & Armitage P. (1997) Biological effects of fine sediment in the lotic
environment. Environmental Management 21, 203–217.
Page 29
18
Yates A.G. & Bailey R.C. (2010) Improving the description of human activities
potentially affecting rural stream ecosystems. Landscape Ecology 25, 371–382.
Yates A.G. & Bailey R.C. (2006) The stream and its altered valley: integrating
landscape ecology into environmental assessments of agro-ecosystems.
Environmental monitoring and assessment 114, 257–271.
Yates A.G., Brua R.B., Culp J.M., Chambers P.A. & Wassenaar L.I. (2014) Sensitivity
of structural and functional indicators depends on type and resolution of
anthropogenic activities. Ecological Indicators 45, 274–284.
Yates A.G., Culp J.M. & Chambers P.A. (2012) Estimating nutrient production from
human activities in subcatchments of the Red River, Manitoba. Journal of Great
Lakes Research 38, 106–114.
Page 30
19
2.0 Effects of agriculture on stream benthic macroinvertebrate
community and trait composition in Prairie streams
2.1 Introduction
Agricultural activities can increase the exposure of stream biota to multiple
stressors, most notably through the input of excess nutrients, fine sediment and
pesticides (Moss 2008, Blann et al. 2009). These stressors can alter both the structure
(e.g., abundance of species) and function (e.g., energy flow through food web) of
aquatic communities through multiple pathways of effect (Allan 2004). Catchment scale
production of crops can contribute excess nutrient inputs to nearby streams through
leaching of fertilizers (Carpenter et al. 1998), resulting in increased aquatic production
and changes in the aquatic food web (Correll 1998, Rabalais 2002). Crop and livestock
land use can also cause changes in habitat at smaller spatial scales which then affects
local scale water quality. For example, crop and livestock land use can increase stream
bank erosion as a result of removal of riparian vegetation, resulting in a local scale
increase in suspended fine sediment (Meador and Goldstein 2003). In turn, this fine
sediment input directly affects aquatic organisms by interfering with respiration or
feeding mechanisms, and indirectly through reduced availability of food sources or
benthic habitat (Wood and Armitage 1997, Henley et al. 2000). Such impacts on stream
Page 31
20
biota in watersheds with intensive agriculture are often difficult to attribute to specific
causes because multiple agricultural and other human activities often co-occur in
catchments.
To discern the pathways of effects of different agricultural activities in multiple
stressor landscapes, a diagnostic approach using bioindicators may be most appropriate
(Bonada et al. 2006). To be effective, diagnostic bioindicators must respond to
particular stressors in the presence of many others and have associations with stressors
that are easily predictive (Dale and Beyeler 2001, Bonada et al. 2006). Structural
metrics (e.g., taxa richness and evenness) are common bioindicators used to describe
changes in environmental conditions due to human impacts (Karr 1991, Cairns et al.
1993, Heik and Kowarik 2010). However, taxonomic-based approaches can be limited
in diagnostic ability because structural bioindicators are often responsive to a wide range
of disturbances and not definitely associated with particular stressors (Culp et al. 2010).
Instead, functional bioindicators have been proposed as better diagnostic tools because
they can be independently linked with specific pathways of effects using testable a
priori mechanistic hypotheses (Bunn and Davies 2000, Menezes et al. 2010).
Biological traits represent the functional relationship between aquatic biota and
their environment, and are thought to be predictable because specific traits can respond
independently to a given stressor (Poff 1997, Townsend and Hildrew 1994). Therefore,
Page 32
21
biological traits may be used to diagnose specific human impacts (Culp et al. 2010,
Menezes et al. 2010, Statzner and Bêche 2010). In addition, a trait-based approach can
simplify the study of complex multi-species assemblages by aggregating taxonomic
information in functional groups while being more consistent over large spatial scales
(Heino et al. 2013). Although biological traits are expected to allow causal inferences
better than a taxonomic approach, it is not yet clear if this is generally the case in all
geographic regions (Heino et al. 2007). Therefore, it is important to consider both
approaches in the development of biomonitoring programs (Yates et al. 2014b).
Taxonomic- and trait-based bioindicators respond to both natural and
anthropogenic factors across different spatial scales (Poff 1997, Heino et al. 2013).
Therefore, to develop suitable diagnostic bioindicators for an area of interest, it is
important to ensure that traits or taxa vary predictively to human impacts with minimal
effect from natural variation (Dale and Beyeler 2001, Bonada et al. 2006, Downes et al.
2010). Multiple catchment studies are considered most appropriate in designing a study
representative of the full range of human exposure and natural gradients (Yates and
Bailey 2006, Bailey et al. 2007), and thus the use of beta diversity (differences in
assemblage among catchments) of taxa and biological traits has been proposed as a
powerful diagnostic approach (Yates et al. 2012b, Heino et al. 2013). Furthermore,
human activity gradients (HAGs) that describe human exposure by quantifying the types
Page 33
22
and intensity of all human activities in a watershed (% land cover of different crop types
instead of simplifying the activity to percent agricultural land cover) have been found to
provide a comprehensive description of human exposure in multiple stressor catchments
(Yates and Bailey 2010, Yates et al. 2012b). Environmental variables, such as nutrient
concentration in streams, are not used as HAGs because they can represent the resulting
response to variation in human activities (Bailey et al. 2007). Instead, environmental
variables are best used alongside HAGs to identify natural gradients or support
hypotheses about the mechanism causing environmental changes as a result of human
activities (Bailey et al. 2007).
This study aimed to evaluate the use of beta diversity of taxa and trait
assemblages to diagnose agricultural impacts in the Red River Valley (RRV) in southern
Manitoba, Canada, using HAGs as an estimate of human activity exposure. While the
RRV watershed is dominated by agriculture (Environment Canada and Manitoba Water
Stewardship 2011), the varying intensities of crops, livestock production and wastewater
discharge from rural villages present throughout the watershed make it difficult to
attribute impacts to specific causes (Yates et al. 2012b). Therefore, the objective of this
study was to compare the sensitivity of trait versus taxa beta diversity to anthropogenic
stressors in the RRV and thus determine if trait beta diversity responded to these in an
more predictable manner. BMIs were chosen as an ecological effect endpoint because
Page 34
23
they have relatively low mobility and short generation times, and thus are indicative of
local environmental conditions (Cairns et al. 1993). HAGs and environmental variables
were grouped by spatial scale to help formulate mechanistic hypotheses about possible
pathways of human impact on beta diversity (Figure 2.1). HAGs based on types of
crops, livestock and wastewater represented catchment scale factors, reach scale factors
were based upon geomorphological and riparian environmental variables describing
stream habitat over a distance of six times the bankfull width, while site scale factors
were characterized by local water quality measurements with a focus on sediments and
nutrients. Based on the literature, crop and livestock agricultural activities were
hypothesized to be associated with changes in stream hydrology and riparian condition
that would alter water quality (Allan 2004, Moss 2008, Blann et al. 2009; Figure 2.1).
Furthermore, wastewater discharge from rural lagoons was predicted to be a direct
source of nutrients to streams (Yates et al. 2012b). Changes in local water quality
caused by catchment scale activities were expected to act as filters constraining beta
diversity of traits and thus taxa (Poff 1997). Moreover, the hypothesized mechanistic
pathways were used to create predictions about trait beta diversity trends. It was
predicted that: (1) high exposure to pesticides would favor smaller body size and short
generation time that increases resilience (Townsend and Hildrew 1994, Liess et al.
2005); (2) excess nutrients would increase herbivores, shredders and/or collector-
Page 35
24
filterers if accompanied with higher food availability, but would include less tegument
respiration in the BMI assemblage if the effect decreases dissolved oxygen levels
(Compin and Céréghino 2007, Carlisle and Hawkins 2008, Feio and Dolédéc 2012); and
(3) excess fine sediment would decrease aquatic respiration, collector-filterers,
herbivores and shredders because of the interference from sediment particles or
reduction in food source availability, coupled with an increase in burrowers which
prefer fine sediment bottoms (Rabeni et al. 2005, Magbanua et al. 2013).
2.2 Methods
2.2.1 Study area
The study was conducted in 20 streams of medium-sized subcatchments (Table
2.1) of the Red River Valley (RRV), part of the watershed of Lake Winnipeg located in
southern Manitoba, Canada (Figure 2.2). The RRV was originally composed of wetlands
that have been drained for agriculture by early settlers; thus streams are predominantly
channelized, U-shaped and without riffle and pool pattern (Yates et al. 2014a). In
addition, the RRV has low topographic relief between the Manitoba Escarpment and the
Canadian Shield, which is also an area characterized by vermiculite clay, silt and fine
sand (Environment Canada and Manitoba Water Stewardship 2011). This flat landscape
Page 36
25
is dominated by agriculture with small rural villages but no major industries (Table 2.1;
Yates et al. 2012b).
Sampling sites on 20 study streams were chosen systematically so as to be
similar in stream order (2-4 Strahler order), roadside accessible (defined as 100 m
upstream or 200 m downstream of a road crossing), and wadable during late spring and
early summer. All 20 sites were on independent tributaries within the RRV (Figure 2.2).
Sites were sampled over a two week period in late spring (late May through early June)
and again in summer (late July through August) to permit determination of the best time
for sampling the BMI community in the RRV because phenological shifts in species
composition may affect community response to land use activity (Carlson et al. 2013).
Two sites could not be sampled in spring because of time limitation (BO01) or low
water depth coupled with dense macrophyte cover (LA02).
2.2.2 BMI collection and trait assignment
BMIs were collected using a 3 minute travelling kick and sweep using a kicknet
(400 μm mesh) at 18 sites in spring 2014 and 20 sites in summer 2014 (Figure 2.2). All
available habitats were sampled proportionately (i.e., areas with or without
macrophytes) and samples were preserved using 95% denatured Ethyl alcohol
(Histoprep). Each sample was subsampled following the Canadian Aquatic
Biomonitoring Network (CABIN) protocol (Reynoldson et al. 2006). Subsampling
Page 37
26
consisted of using a Marchant box to divide the sample (100 cell sub-sampling box;
Marchant 1989), then randomly choosing cells for sorting until at least 300 organisms
were counted. Abundances of organisms in a sample were then estimated by
extrapolating abundances from subsamples to estimate the abundance in the entire
samples. All BMIs were identified by a Society of Freshwater Science-certified
taxonomist to lowest taxonomic level possible. However, only aquatic insect genera
were used for data analysis because of the greater availability of trait information for
this taxonomic group. Quality assurances were completed for 10% of samples to ensure
a minimum of 95% taxonomic accuracy. A site-by-taxa matrix of aquatic insect genera
was created by pooling species- and genus-level abundances for each site (Appendix 1).
Organisms not identified to genus- or species-level and taxa present in less than 10% of
sites were excluded from the matrix used for further analyses.
Traits were assigned to aquatic insect genera using a binary approach (0 = genus
does not have the trait state, 1=genus does have the trait state) summed across all genera
for each site (Table 2.2, Appendix 2). Trait information was obtained from a database
developed by the National Center for Environmental Assessment (Vieira et al. 2006). If
trait information differed between studies reported in the database, a “majority wins”
approach was used to decide which trait state to assign to the genus. However, if for a
given genus, two trait states were stated in equal frequency by studies in the database,
Page 38
27
trait states information from studies in northern US were given priority, or information
from species known to be present in the RRV was used to break the tie. When no
information was provided for a trait category in the database, trait states represented by
the majority of species were assigned to that genus; if information was missing for
species, the literature was used to assign a trait state to genera (Meyer 2006, Merritt and
Cummins 2008, Armitage et al. 2012).
2.2.3 Natural factors and HAGs
Site scale water chemistry, reach characteristics and the magnitude of catchment
scale human activities (HAGs) were recorded for all sites (Table 2.3, 2.4, Appendix 3).
Water samples were collected once to three times during each of the two week sampling
periods and analysed by the Biogeochemical Analytical Service Laboratory, Edmonton,
Canada, for nutrients following standard protocols (United States Environmental
Protection Agency (USEPA) 1993a,b; American Public Health Associated (APHA),
2012). Water samples with nutrient concentrations below measurement detection levels
were recorded as 0, and measurements were averaged for each of the two sampling
periods. Total suspended solids (TSS) samples were measured by weighing dried solids
filtered on GF/F filters (pre-ashed for 1 hour at 550ᵒC) at 104ᵒC for 1 hour. The TSS
sample for site BO01 in summer was lost, and the average of all sites during that period
Page 39
28
was used as an estimate of summer TSS for BO01 to avoid a missing data point in
further analyses (Quinn and Keough 2002).
On each sampling trip, reach characteristics were measured at every site (canopy
cover (%), wetted width (m), average depth (cm), macrophytes cover (categories) and
riparian vegetation (categories); Table 2.3). Canopy cover estimates were made with a
densitometer and calculated as the average of four readings (upstream, downstream,
right and left banks) of the degree of coverage of the spherical densitometer by
overstory vegetation. Wetted width and water depth were averaged across three stream
cross sections along the sampling site. Macrophytes and riparian vegetation were
described using categories similar to reach characteristics as defined in the CABIN
protocol (i.e., 0-24%, 25-50%, 50-75%, or 75-100% macrophyte cover, dominant
riparian vegetation and list of all vegetation present; Reynoldson et al. 2006). In
addition, sinuosity of the channels was calculated from digital images for a 0.5 km
section above and below the BMI sampling site and stream catchment area was
estimated using geospatial information as described below (Table 2.3).
The amount and type of human activity in each catchment was estimated from
geospatial data using Quantum GIS 2.6.1 (Table 2.5, QGIS development team 2014).
First, site catchments were delimited using an ASTER (Advanced Spaceborne Thermal
Emission and Reflection Radiometer) DEM (Digital Elevation Model; version 2,
Page 40
29
product of METI and NASA) with a resolution of 1 arc-second, and GRASS tools
(GRASS Development Team 2014). Land cover of different crops were compiled from
2013 Agriculture and Agri-food Canada crop type map of Canada, a 30-m-resolution
raster created from remote-sensing images (STB-EO 2014). Livestock density in each
watershed was estimated using 2011 Canadian census data following the methods of
Yates et al. (2012). Density of each type of livestock was converted to amount of
manure produced by each based on nutrient coefficients developed by Ontario Ministry
of Agriculture, Food and Rural Affairs (2007). Wastewater discharge was calculated as
the density of people served by sewage lagoons that either discharge to streams or are
pumped and applied to crops as fertilizer, based on population estimates from the 2011
Canadian census or personal communications from municipalities.
2.2.4 Data analysis
Spring and summer assemblages were analysed separately to avoid variability
due to life history phenology as explained above, and presence/absence data was used to
represent the taxa assemblages to avoid the influence of statistical outliers due to most
genera having many zeroes and a few non-zero points (Quinn and Keough, 2002). The
same was done with trait data for consistency. Predictors were divided into categories
(site, reach and catchment scales) and correlation among them were explored using
separate Principal Component Analyses (PCA) on transformed and standardized data
Page 41
30
(Tables 2.3, 2.4). To avoid statistical overfitting, only predictors considered important
based on the PCAs and predictors directly related to a priori hypotheses were included
in further analyses. Therefore, site scale water quality was represented by total dissolved
phosphorus, nitrite and nitrate, and TSS (Table 2.6, Figure 2.3); reach scale variables
included were % canopy cover, % macrophytes cover and wetted width (Table 2.6,
Figure 2.3); and catchment scale human activity were represented by the percentage of
small grains, swine nutrient density and population served by wastewater lagoons
discharging to streams (Table 2.3, Figure 2.6).
Detrended Correspondence Analyses (DCA) were performed on taxa and trait
matrices of either sampling times to determine if the community data were linear or
unimodal, using a first gradient length of 3.5 standard deviations as the upper threshold
(Legendre and Legendre 1998). Because all datasets were linear, PCAs were run to
explore the site-based association of taxa and trait assemblages, followed by
Redundancy Analyses (RDA) to examine the association between beta diversity
(variation in assemblages among sites) and predictors (Anderson et al. 2011). RDA
correlation triplots were also used to visualize these associations and axes were tested
for statistical significance using Monte Carlo permutations. All analyses were performed
using R 3.1.3 (R Core Team 2014).
Page 42
31
2.3 Results
2.3.1 Macroinvertebrate and trait assemblage structure
The first PCA axes of both the taxa and trait matrices accounted for a similar
proportion of community variance between the two sampling seasons (Table 2.7);
however, more variance was explained by the first PCA axis for traits than taxa. For the
two sampling seasons, variance explained by the first PCA axis average 16% for the
taxa matrix compared to 59% for the trait matrix. Collectively, the first three axes of the
taxa assemblage explained 37% and 42% of the total variation among sites in late spring
and summer respectively, whereas the same three axes accounted for a total of 82 and
81% of the variation in the trait matrix. In fact, the first PCA axis of the trait assemblage
alone accounted for 62% of variation in late spring, and 56% of variance in summer
(Table 2.7).
In late spring, the PCA biplot of taxa assemblage showed some clustering of
sites along the first two axes (Figure 2.5a). The cluster of sites associated with the first
PCA axis (PC1) was positively associated with Simulium, (Diptera), Triaenodes
(Trichoptera) and Perlesta (Plecoptera) and two Ephemeroptera genera (Procloeon and
Paraleptophlebia). The cluster of sites positively associated with the second PCA axis
(PC2) was associated with Ceratopogon (Diptera) whereas the cluster of sites negatively
associated with PC2 was associated with Baetis and Caenis (Ephemeroptera) (Figure
Page 43
32
2.5a). In addition, Chironomidae genera (Diptera) were present along both PC1 and PC2
axes.
In summer, the PCA biplot of taxa assemblages appeared centrally clustered
(Figure 2.5b). The spatial separation of site along the first axis (PC1) was determined by
the presence of Coleoptera and Ephemeroptera genera: Coleoptera genera were
positively associated with PC1, whereas Ephemeroptera genera had a negative
association with that axis (Figure 2.5b). The second PCA axis (PC2) was positively
associated with Simulium, Thienemaniella and Cricotopus (Diptera), and negatively
associated with Procloeon (Ephemeroptera) as well as other Diptera genera (Figure
2.5b). As seen in spring, Chironomidae (Diptera) genera were present along both PC1
and PC2 in summer.
In contrast with the taxa PCAs, the trait PCAs in late spring and summer had a
wide spread of sites along the first PCA axis (PC1). In both seasons, this gradient
represented trait richness, from sites highly associated with collector-gatherers,
tegument respiration and multiple generations per year, to sites with low presence of all
traits included in the analysis (Figure 2.6 a, b). The second PCA axis (PC2) in late
spring and summer was positively associated with medium sized insects and predators,
and negatively associated with small insects (Figure 2.6a,b). Although traits had similar
patterns between the two sampling times, sites along the two PCA axes differed.
Page 44
33
2.3.2 Association with environmental variables
Taxa and trait assemblages were correlated with environmental variables for
both sampling periods. In spring, only the first RDA axis (RDA1) was significant for
both taxa (22% variance explained, p<0.05)) and trait (49% variance explained, p<0.05,
Table 2.8) beta diversity. In summer, the first two axes (RDA1, RDA2) were significant
for taxa beta diversity (23 and 17% variance explained respectively, p<0.05), whereas
the first three RDA axes (RDA1-RDA3) explained significant (p<0.05) variation in trait
beta diversity (44, 19 and 12% variance explained respectively; Table 2.8). In addition,
significant RDA axes consistently explained more variation in trait beta diversity
compared to taxa beta diversity (Table 2.8).
Taxa beta diversity in late spring was associated with both small grain crop
production and channel wetted width along the first RDA gradient (RDA1, Figure 2.7).
Small grain crop production and wetted width were positively associated with
Chironomidae and Coleoptera genera but negatively associated with Simulium (Diptera,
loading = -0.40) and Procloeon (Ephemeroptera, loading = -0.42). Other
Ephemeroptera, Tricoptera and Plecoptera (EPT) genera were also negatively associated
with small grain production and wetted width. In addition, canopy cover and wastewater
appeared to be the least important variables affecting taxa assemblages in the spring
(Figure 2.7).
Page 45
34
In contrast, taxa beta diversity was strongly associated with canopy cover and
other reach and site scale factors in summer (Figure 2.8). RDA1 in summer represented
a gradient from high canopy cover to low canopy cover with abundant macrophytes and
high total dissolved phosphorus. The RDA1 gradient was associated with EPT genera,
present in sites with mid- to high % canopy cover. Furthermore, Somatochlora
(Odonata) had the highest correlation with % canopy cover (loading = -0.62), whereas
Peltodytes larvae (Coleoptera) was highly correlated with % macrophytes and
phosphorus concentration (loading = 0.52; Figure 2.8). In addition, taxa diversity in
summer was driven by catchment scale agricultural activities and TSS, represented by
the second RDA axis (RDA2). Moreover, small grain crop production was positively
associated with total suspended solids and negatively correlated with livestock
production along RDA2 (Figure 2.8). Hydrochus (Coleoptera) had the strongest
correlation with small grain crops and TSS (loading = 0.52), whereas Simulium
(Diptera) was correlated with swine production (loading = -0.52).
Trait beta diversity had a similar association to human activity and
environmental variables as taxa beta diversity. Catchment scale agriculture and wetted
width were important drivers of trait beta diversity in late spring, whereas local scale
environmental factors were the primary drivers in summer. In late spring, the first RDA
axis (RDA1) was positively associated with wetted width and to a lesser extend
Page 46
35
negatively with swine and total dissolved phosphorus (Figure 2.9). Most traits had a
strong positive loading on RDA1, and thus were positively associated with wetted width
(loadings >0.60). Furthermore, only insect with gills and collector-filterers were
negatively associated with RDA1 in spring (Figure 2.9). In summer, trait beta diversity
was strongly associated to a gradient of canopy cover and total dissolved phosphorus
along the first RDA axis (RDA1; Figure 2.10). Predators (loading = -0.89) and
collector-gatherers (loading = -0.84) had the strongest positive association with %
canopy cover. In contrast, herbivores and shredders were found in sites with low %
canopy cover (Figure 2.10). The second RDA axis (RDA2) was positively associated
with small grain crops and TSS, and negatively with swine production (Figure 2.10).
Furthermore, air breathers (loading = 0.70) had the highest positive loading on this axis,
present in sites with high small grain crop production and TSS, whereas clingers
(loading = -0.48) and collector-filterers (loading = -0.40) had the highest negative
loadings. Sites with high TSS were also positively associated with burrowers, but
negatively associated with shredders (Figure 2.10). The third RDA axis (RDA3) in
summer appeared to be a gradient of water quality associated with small grain
production (Figure 2.11).
Page 47
36
2.4 Discussion
2.4.1 Drivers of trait and taxa beta diversity
The sensitivity of trait and taxa beta diversity to catchment scale human
activities and natural factors at different spatial scales revealed that local environmental
factors were the main drivers of aquatic insect assemblages in the RRV. Reach scale
factors were strongly associated with biological patterns in both sampling periods (i.e.,
channel wetted width in spring and canopy cover in summer), while catchment scale
agriculture was associated with beta diversity in spring, but only weakly in summer.
Previous studies have concluded that seasonal differences in natural hydrological factors
act as a strong selection pressure (e.g., high discharge and flooding during spring, Bêche
et al. 2006, Horrigan and Baird 2008) and can create seasonal differences in ecological
sensitivity to agricultural land use (Carlson et al. 2013, Botwe et al. 2015). Thus, the
strong association of trait and taxa beta diversity with channel wetted width in the spring
supports the conclusion that the hydrological gradient is an overriding environmental
filter in the RRV. Based on genera level taxonomic information in the North American
trait database (Viera et al. 2006), EPT taxa found in narrow streams are known to
tolerate strong water currents, whereas genera predominantly present in depositional
habitats dominated assemblages in wide streams. Trait beta diversity in the RRV
exhibited this skewed distribution and association of EPT genera with stream
Page 48
37
geomorphology: specifically insects with gills were associated with narrow streams,
whereas all other traits were more common in wide streams. Snowmelt in early spring is
the major discharge event in the RRV (Corriveau et al. 2011, McCullough et al. 2012)
and may have a lasting effect on aquatic insect assemblages. Thus, the effects of channel
width in these U-shaped prairie streams is likely associated with geomorphological
differences created during these high flow periods as discharge during the spring
sampling period (post-snowmelt) was very low (near zero) and did not vary
considerably among sites.
Trait beta diversity was associated with both swine production and TDP in
spring, providing support for the hypothesized mechanistic link between catchment
scale agriculture and local water quality. Hydrological conditions during the spring
snowmelt period in the RRV are known to result in greater connectivity between
catchments and their streams (McCullough et al. 2012) which results in higher runoff of
nutrients from a greater area than in summer (Yates et al. 2014a). Therefore, trait and
taxa beta diversity may be affected by catchment scale agricultural activities in spring
through changes in water quality resulting from an increase in hydrological connectivity
within catchments during this period. Although an increase in occurrence of certain
functional feeding groups (FFG; i.e., herbivores, shredders and collector-filterers) was
predicted in sites with higher nutrient concentration (Comping and Céréghino 2007,
Page 49
38
Carlisle and Hawkins 2008), FFG traits were either unrelated (i.e., collector-filterers) or
present in sites with low TDP. Furthermore, tegument respiration was negatively
associated with TDP as was predicted if excess nutrients affected dissolved oxygen
levels (Feio and Dolédéc 2012), but an autocorrelation among traits may have created
this association (i.e., trait syndromes, Poff et al. 2006). Moreover, tegument respiration
was the most common respiratory method in the study area (i.e., more than 50% of
genera used tegument respiration), and most genera were found in sites with low TDP
levels. The occurrence of genera with gills in sites with high TDP provides further
evidence that the association among swine production, TDP and traits may not have
been cause by changes in dissolved oxygen (Statzner and Bêche 2010). Therefore, the
effect of high TDP and swine production on BMI assemblages in the RRV is unclear.
The importance of local natural gradients for both trait and taxa beta diversity
was not limited to the spring. In summer, trait and taxa assemblages were strongly
associated with reach and site scale factors, such as canopy cover and TDP, and weakly
related to catchment land use. More specifically, both trait and taxa beta diversity were
associated with a primary gradient of riparian condition, ranging from sites of high
canopy cover to sites with high macrophytes cover and TDP, whereas a secondary
gradient of catchment scale land use and TSS explained a lower amount of variation in
assemblages. FFGs were distributed along the primary gradient, with collector-gatherers
Page 50
39
in streams with high canopy cover, and herbivores in sites with grass dominated riparian
vegetation and higher in-stream TDP. Although I predicted that excess nutrients would
increase herbivory due to higher food availability (Comping and Céréghino 2007,
Carlisle and Hawkins 2008), the importance of canopy cover suggests that light
availability was also a critical factor. Shading can limit algal production and result in
stream food webs that are more dependent on allochthonous energy sources, such as leaf
litter than autochthonous algal production (Lyons et al. 2000). Indeed, streams with high
canopy cover were found to have lower gross primary production in the RRV (Yates et
al. 2012a). Therefore, light availability likely has a strong effect on food availability in
the RRV and thus constrains the distribution of herbivores.
Many other studies have found that reach scale factors, such as riparian
condition, were an important driver of trait and taxa diversity because they affect food
and microhabitat availability (e.g., Richards et al. 1997, Sponseller et al. 2001), which
in turn filter traits and constrain taxa found in a specific area (Poff 1997, Strayer et al.
2003, Heino 2004). Often, these small scale differences in riparian condition are caused
by agricultural practices and can elucidate the mechanism of agricultural impacts (Roth
et al. 1996, Kyriakeas and Watzin 2006, Hutchens et al. 2009, Carlson et al. 2013).
However, this was not the case in the RRV because agricultural activities were not
associated with reach scale riparian condition. Rather, catchment scale agricultural
Page 51
40
activities were an independent secondary driver of beta diversity associated with TSS,
supporting the prediction that traits are affected by agriculture through changes in water
quality. As predicted, this gradient was associated with the presence of burrowers and
air breathers at sites of high small grain crop production and TSS, but an absence in
shredders and, to a certain extent, collector-filterers, likely because of the effect of TSS
on habitat and food source availability (Wood and Armitage 1997, Henley et al. 2000).
However, this gradient only explained a small proportion of the variance in trait
assemblages, indicating that there are minimal impacts on stream communities in the
RRV from catchment scale agriculture in summer. Additional predictions about the
effect of wastewater discharge, nutrients and pesticides on traits were not supported by
results, further suggesting a limited impact of anthropogenic pollutants in summer on
aquatic insect assemblages.
2.4.2 Biological traits as bioindicators for the RRV
Biological traits are predicted to have greater seasonal stability and be more
sensitive to stressors of interest compared to taxonomic approaches (Culp et al. 2010,
Menezes et al. 2010). However, the similarity in responses between trait and taxa beta
diversity in the RRV suggest that both functional and structural approaches can show
similar trends in temporal stability and sensitivity to anthropogenic stressors. Moreover,
drivers trait beta diversity in this study were seasonally dependent. The RRV is a system
Page 52
41
driven by snowmelt and surface runoff, creating large hydrological differences among
seasons (Corriveau et al. 2011, McCullough et al. 2012, Yates et al. 2014a). Therefore,
results suggest that in environments with major seasonal changes in habitat, traits can
exhibit seasonal patterns. This matches the findings from Bêche et al. (2006) and
Bonada (2003) that showed temporal variation of BMI trait composition in streams with
strong seasonal patterns, such as ephemeral or intermittent streams in the Mediterranean
region. In addition, trait and taxa beta diversity exhibited similar sensitivity to catchment
scale agricultural activities, in contrast to what was expected. Although trait and taxa
beta diversity were equally sensitive to land use, the association between trait beta
diversity and important local habitat variables was stronger, as shown previously by
Heino et al. (2007).
Results of this study also demonstrates the advantages of using a trait-based
approach because of its ability to test a priori mechanistic hypotheses about the pathway
of human impacts and the ease in interpreting results compared to a taxonomic approach
(Poff et al. 1997, Culp et al. 2010, Statzner and Bêche 2010, Lange et al. 2014). For
example, traits responded as predicted, albeit weakly, to increased TSS associated with
catchment scale agriculture in summer. In addition, when predictions about human
impacts were not supported by relationships with catchment scale gradients, it was
possible to infer causal relationships between traits and local habitat factors. Moreover,
Page 53
42
the response of functional feeding groups to riparian condition was easily interpretable
due to the known relationship between FFG and habitat variables. In contrast,
interpreting taxa beta diversity in spring without relying on trait information would not
have allowed the formulation of new hypotheses about the possible confounding effect
of hydrology. Therefore, a trait-based approach can aid in inferring causal links between
aquatic insect assemblages and environmental stressors at different scales.
2.4.3 Conclusions
Trait and taxa beta diversity were not effective diagnostic bioindicators of
catchment scale agricultural activities in the RRV because of the strong influence of
natural gradients and a lack of relationship between local habitat factors and catchment
scale land use. In spring, a hydrological gradient appeared to confound the effects of
agriculture, while in summer, reach scale riparian condition was the major driver of beta
diversity because of its strong filtering of FFG traits. The lack of relationship between
catchment scale agriculture and local habitat may be due to historical stream
modification having a lasting influence on stream habitat and thus aquatic assemblages
(Hardings et al. 1998). This is likely in the study area because of the extensive
channelization of streams during settlement and long history of agriculture in this
watershed. In addition, the seasonality of discharge in the RRV may allow stressors to
bypass local habitat because of high discharge during snowmelt and early spring high
Page 54
43
flows (Corriveau et al. 2011), while limiting runoff of stressors in summer because of
the disconnection of streams and their catchment during this dry period (Yates et al.
2014a). It is also possible that the highly modified nature of streams in this study (Table
2.5) has created a species pool dominated by tolerant aquatic insect taxa, limiting its
sensitivity to agricultural impacts (Heino 2013). Therefore, monitoring efforts in the
RRV should carefully consider whether the use of aquatic insects as bioindicators of
catchment scale agricultural impacts is appropriate largely because of the influence of
natural and historical factors. However, the strong relationship between trait beta
diversity and local habitat factors suggest that a trait-based approach is appropriate for
future studies wanting to identify local habitat changes in the RRV and this study
underscores the ability of biological traits to help identify important factors affecting
aquatic communities in multiple stressors environment.
2.5 References
Allan J.D. (2004) Landscapes and riverscapes: the influence of land use on stream
ecosystems. Annual Review of Ecology, Evolution, and Systematics 35, 257–284.
Anderson M.J., Crist T.O., Chase J.M., Vellend M., Inouye B.D., Freestone A.L., et al.
(2011) Navigating the multiple meanings of B diversity: a roadmap for the
practicing ecologist. Ecology Letters 14, 19–28.
Armitage P.D., Cranston P.S. & Pinder L.C. V. (2012) The Chironomidae: biology and
ecology of non-biting midges. Springer Science & Business Media.
Page 55
44
American Public Health Association. (2012) Standard methods for the examination of
water and wastewater. American Public Health Association, Washington, DC.
Bailey R.C., Reynoldson T.B., Yates A.G., Bailey J. & Linke S. (2007) Integrating
stream bioassessment and landscape ecology as a tool for land use planning.
Freshwater Biology 52, 908–917.
Bêche L.A., McElravy E.P. & Resh V.H. (2006) Long-term seasonal variation in the
biological traits of benthic-macroinvertebrates in two Mediterranean-climate
streams in California, U.S.A. Freshwater Biology 51, 56–75.
Blann K.L., Anderson J.L., Sands G.R. & Vondracek B. (2009) Effects of agricultural
drainage on aquatic ecosystems: a review. Critical Reviews in Environmental
Science and Technology 39, 909–1001.
Bonada N. (2003) Ecology of the macroinvertebrate communities in Mediterranean
rivers at different scales and organization levels. University of Barcelona, Spain.
Bonada N., Prat N., Resh V.H. & Statzner B. (2006) Developments in aquatic insect
biomonitoring: a comparative analysis of recent approaches. Annual review of
entomology 51, 495–523.
Botwe P.K., Barmuta L.A., Magierowski R., McEvoy P., Goonan P. & Carver S. (2015)
Temporal patterns and environmental correlates of macroinvertebrate communities
in temporary streams. Plos One 10.
Bunn S. & Davies P. (2000) Biological processes in running waters and their
implications for the assessment of ecological integrity. Hydrobiologia 422, 61–70.
Cairns J., McCormick P. V. & Niederlehner B.R. (1993) A proposed framework for
developing indicators of ecosystem health. Hydrobiologia 263, 1–44.
Carlisle D.M. & Hawkins C.P. (2008) Land use and the structure of western US stream
invertebrate assemblages: predictive models and ecological traits. Journal of the
North American Benthological Society 27, 986–999.
Page 56
45
Carlson P.E., Johnson R.K. & McKie B.G. (2013) Optimizing stream bioassessment:
habitat, season, and the impacts of land use on benthic macroinvertebrates.
Hydrobiologia 704, 363–373.
Carpenter S.R., Caraco N.F., Correll D.L., Howarth R.W., Sharpley A.N. & Smith V.H.
(1998) Nonpoint pollution of surface waters with phosphorus and nitrogen.
Ecological Applications 8, 559–568.
Compin A. & Céréghino R. (2007) Spatial patterns of macroinvertebrate functional
feeding groups in streams in relation to physical variables and land-cover in
Southwestern France. Landscape Ecology 22, 1215–1225.
Correll D.L. (1998) The role of phosphorus in the eutrophication of receiving waters: a
review. Journal of Environment Quality 27, 261-266.
Corriveau J., Chambers P. A., Yates A.G. & Culp J.M. (2011) Snowmelt and its role in
the hydrologic and nutrient budgets of prairie streams. Water Science &
Technology 64, 1590–1596.
Culp J.M., Armanini D.G., Dunbar M.J., Orlofske J.M., Poff N.L., Pollard A.I., et al.
(2010) Incorporating traits in aquatic biomonitoring to enhance causal diagnosis
and prediction. Integrated environmental assessment and management 7, 187–197.
Dale V.H. & Beyeler S.C. (2001) Challenges in the development and use of ecological
indicators. Ecological Indicators 1, 3–10.
Downes B.J. (2010) Back to the future: little-used tools and principles of scientific
inference can help disentangle effects of multiple stressors on freshwater
ecosystems. Freshwater Biology 55, 60–79.
Environment Canada & Manitoba Water Stewardship (2011) State of Lake Winnipeg:
1999 to 2007. Winnipeg, MB.
Feio M.J. & Dolédec S. (2012) Integration of invertebrate traits into predictive models
for indirect assessment of stream functional integrity: a case study in Portugal.
Ecological Indicators 15, 236–247.
Page 57
46
GRASS Development Team (2014) Geographic Resources Analysis Support System
(GRASS) Software, Version 6.4.3. Open Source Geospatial Foundation.
Harding J.S., Benfield E.F., Bolstad P. V, Helfman G.S. & Jones E.B. (1998) Stream
biodiversity: the ghost of land use past. Proceedings of the National Academy of
Sciences of the United States of America 95, 14843–14847.
Heink U. & Kowarik I. (2010) What are indicators? On the definition of indicators in
ecology and environmental planning. Ecological Indicators 10, 584–593.
Heino J. (2013) The importance of metacommunity ecology for environmental
assessment research in the freshwater realm. Biological Reviews 88, 166–178.
Heino J., Louhi P. & Muotka T. (2004) Identifying the scales of variability in stream
macroinvertebrate abundance, functional composition and assemblage structure.
Freshwater Biology 49, 1230–1239.
Heino J., Mykrä H., Kotanen J. & Muotka T. (2007) Ecological filters and variability in
stream macroinvertebrate communities: do taxonomic and functional structure
follow the same path? Ecography 30, 217–230.
Heino J., Schmera D. & Erős T. (2013) A macroecological perspective of trait patterns
in stream communities. Freshwater Biology 58, 1539–1555.
Henley W.F., Patterson M.A., Neves R.J. & Lemly A.D. (2000) Effects of sedimentation
and turbidity on lotic food webs: a concise review for natural resource managers.
Reviews in Fisheries Science 8, 125–139.
Horrigan N. & Baird D.J. (2008) Trait patterns of aquatic insects across gradients of
flow-related factors: a multivariate analysis of Canadian national data. Canadian
Journal of Fisheries and Aquatic Sciences 65, 670–680.
Hutchens J.J., Schuldt J.A., Richards C., Johnson L.B., Host G.E. & Breneman D.H.
(2009) Multi-scale mechanistic indicators of Midwestern USA stream
macroinvertebrates. Ecological Indicators 9, 1138–1150.
Karr J.R. (1991) Biological integrity : a long-neglected aspect of water resource
management. Ecological Applications 1, 66–84.
Page 58
47
Kyriakeas S.A. & Watzin M.C. (2006) Effects of adjacent agricultural activities and
watershed characteristics on stream macroinvertebrate communities. Journal of the
American Water Resources Association 42, 425–441.
Lange K., Townsend C.R. & Matthaei C.D. (2014) Can biological traits of stream
invertebrates help disentangle the effects of multiple stressors in an agricultural
catchment? Freshwater Biology 59, 2431–2446.
Legendre P. & Legendre L. (1998) Numerical ecology, Second English edition. Elsevier
Science B.V., Amsterdam.
Liess M. & Von Der Ohe P.C. (2005) Analyzing effects of pesticides on invertebrate
communities in streams. Environmental Toxicology and Chemistry 24, 954.
Lyons J., Thimble S.W. & Paine L.K. (2000) Grass versus trees: managing riparian
areas to benefit streams of central North America. Journal of the American Water
Resources Association 36, 919–930.
Magbanua F.S., Townsend C.R., Hageman K.J. & Matthaei C.D. (2013) Individual and
combined effects of fine sediment and the herbicide glyphosate on benthic
macroinvertebrates and stream ecosystem function. Freshwater Biology 58, 1729–
1744.
Marchant R. (1989) A subsampler for samples of benthic invertebrates. Bulletin of the
Australian Society for Limnology 12, 49–52.
McCullough G.K., Page S.J., Hesslein R.H., Stainton M.P., Kling H.J., Salki A.G., et al.
(2012) Hydrological forcing of a recent trophic surge in Lake Winnipeg. Journal of
Great Lakes Research 38, 95–105.
Meador M.R. & Goldstein R.M. (2003) Assessing water quality at large geographic
scales: relations among land use, water physicochemistry, riparian condition, and
fish community structure. Environmental Management 31, 504–517.
Menezes S., Baird D.J. & Soares A.M.V.M. (2010) Beyond taxonomy: a review of
macroinvertebrate trait-based community descriptors as tools for freshwater
biomonitoring. Journal of Applied Ecology 47, 711–719.
Page 59
48
Merritt R.W. & Cummins K.W. (2008) An Introduction to the Aquatic Insects of North
America. Kendall Hunt.
Meyer J.R. Respiration in Aquatic Insects. Department of Entomology NC State
University.
Moss B. (2008) Water pollution by agriculture. Philosophical Transactions of the Royal
Society of London. Series B, Biological sciences 363, 659–666.
Ontario Ministry of Agriculture Food and Rural Affairs (2007) Nutrient Management
Protocol.
Poff N.L. (1997) Landscape filters and species traits: towards mechanistic understanding
and prediction in stream ecology. Journal of the North American Benthological
Society 16, 391–409.
Poff N.L., Olden J.D., Vieira N.K.M., Finn D.S., Mark P., Kondratieff B.C., et al.
(2006) Functional trait niches of North American lotic insects: traits-based
ecological applications in light of phylogenetic relationships. Journal of the North
American Benthological Society 25, 730–755.
QGIS development team (2014) QGIS Geographic Information System. Open Source
Geospatial Foundation Project.
Quinn G.P. & Keough M.J. (2002) Experimental design and data analysis for biologists.
Cambridge University Press, New York, USA.
R Core Team (2015) R: A language and environment for statistical computing.
Rabalais N.N. (2002) Nitrogen in aquatic ecosystems. Ambio 31, 102–112.
Rabení C.F., Doisy K.E. & Zweig L.D. (2005) Stream invertebrate community
functional responses to deposited sediment. Aquatic Sciences 67, 395–402.
Reynoldson T.B., Logan C., Pascoe T. & Thompson S.P. (2006) Invertebrate
biomonitoring field and laboratory manual for running water habitats. National
Water Research Institute, Environment Canada, Burlington, ON.
Page 60
49
Richards C., Haro R., Johnson L.B. & Host G.E. (1997) Catchment and reach-scale
properties as indicators of macroinvertebrate species traits. Freshwater Biology 37,
219–230.
Roth N.E., Allan J.D. & Erickson D.L. (1996) Landscape influences on stream biotic
integrity assessed at multiple spatial scales. Landscape Ecology 11, 141–156.
Sponseller R., Benfield E. & Valett M. (2001) Relationships between land use, spatial
scale and stream macroinvertebrate communities. Freshwater Biology 46, 1409 –
1424.
Statzner B. & Bêche L.A. (2010) Can biological invertebrate traits resolve effects of
multiple stressors on running water ecosystems? Freshwater Biology 55, 80–119.
STB-EO (2014) 2013 AAFC Crop Type Map of Canada, version 1.
ftp://ftp.agr.gc.ca/pub/outgoing/aesb-eos-gg/Crop_Type_Mapping/2013/
Strayer D.L., Beighley R.E., Thompson L.C., Brooks S., Nilsson C., Pinay G., et al.
(2003) Effects of land cover on stream ecosystems: roles of empirical models and
scaling issues. Ecosystems 6, 407–423.
Townsend C.R. & Hildrew A.G. (1994) Species traits in relation to a habitat templet for
river systems. Freshwater Biology 31, 265–275.
United States Environmental Protection Agency (1993) Determination of nitrate-nitrite
nitrogen by automated colorimetry, Method 353.2. Cincinnati, OH.
United States Environmental Protection Agency (1993) Determination of total nitrogen
by automated colorimetry, Method 353.2. Cincinnati, OH.
Vieira N.K.M., Poff N.L., Carlisle D.M., Moulton II S., Koski M. & Kondratieff B.
(2006) A database of lotic invertebrate traits for North America. Geological
Survey Data Series 187, Reston, Virginia.
Wood P. & Armitage P.D. (1997) Biological effects of fine sediment in the lotic
environment. Environmental Management 21, 203–217.
Page 61
50
Yates A.G. & Bailey R.C. (2010) Improving the description of human activities
potentially affecting rural stream ecosystems. Landscape Ecology 25, 371–382.
Yates A.G., Brua R.B., Corriveau J., Culp J.M. & Chambers P.A. (2014a) Seasonally
driven variation in spatial relationships between agricultural land use and in-stream
nutrient concentration. River Research and Applications 30, 476–493.
Yates A.G., Brua R.B., Culp J.M. & Chambers P.A. (2012a) Multi-scaled drivers of
rural prairie stream metabolism along human activity gradients. Freshwater
Biology 58, 675–689.
Yates A.G., Brua R.B., Culp J.M., Chambers P.A. & Wassenaar L.I. (2014b) Sensitivity
of structural and functional indicators depends on type and resolution of
anthropogenic activities. Ecological Indicators 45, 274–284.
Yates A.G., Culp J.M. & Chambers P.A. (2012b) Estimating nutrient production from
human activities in subcatchments of the Red River, Manitoba. Journal of Great
Lakes Research 38, 106–114.
Page 62
51
Table 2.1: Name and catchment description of 20 sites sampled in the Red River Valley in southern Manitoba, Canada.
Subwatershed Stream name Code Area
(km2)
Crop
(%)
Livestock
(nut. density/km2)
Population using WWT
lagoons (ind./km2)
Boyne River Boyne River BO01 276.5 76.4 24.5 2.2
Brokenhead River Bears Creek BR04 136.8 25.9 2.9 0.0
Brokenhead River Unnamed Drain BR05 163.4 49.5 3.0 0.7
LaSalle River West Branch Drain LA02 64.9 89.4 13.8 1.7
LaSalle River Elm River LA03 105.3 80.1 7.8 4.4
Lower Red Drainage Cooks Creek LR01 176.5 43.2 47.0 0.6
Lower Red Drainage Gamby Drain LR02 544.8 72.0 16.6 11.7
Lower Red Drainage Devils Creek LR03 305.6 58.2 5.9 3.3
Lower Red Drainage Edie Creek LR04 80.4 26.0 3.7 0.0
Lower Red Drainage Satans Creek LR05 140.9 30.6 4.3 0.9
Morris River South Tobacco Creek MO04 149.5 73.8 13.9 4.2
Morris River North Shannon Creek MO06 189.6 91.2 7.4 0.8
Morris River Shannon Creek MO07 157.4 79.1 18.7 0.0
Roseau River Jordan River RO01 124.8 37.0 11.9 0.0
Roseau River Casson-Stewart Drain RO02 156.5 24.1 11.1 0.0
Rat/Touround Drainages Manning Canal RT04 375.4 53.2 141.9 39.1
Rat/Touround Drainages Unnamed Drain RT06 88.7 56.3 104.7 0.0
Upper Red Drainage Hespeler Drain UR03 255.8 81.1 21.4 0.0
Upper Red Drainage Deadhorse Creek UR04 360.2 81.4 21.3 53.6
Upper Red Drainage Krongsgart Drain UR05 208.0 95.2 16.3 0.0
Page 63
52
Table 2.2: Trait categories and trait states included in trait analyses of benthic
macroinvertebrate genera in southern Manitoba and their abbreviations (based on Poff et
al. 2006).
Trait
Group Trait category Trait state (modality) Abbreviation
Life
History Voltinism Multiple generations per year Multi
One generation per year Uni
Less than one generation per
year Semi
Morphol
ogy Maximum Body size Large L
Medium M
Small S
Respiration Tegument teg
Gills gills
Pigment (and tegument and/or
tracheal tubes) pigP
Aerial (plastron, spiracle) air
Ecology Functional Feeding
Group (FFG) Collector-filterer CF
Collector-gatherer CG
Herbivore (scraper, piercer, and
shedder) HB
Predator (piercer and engulfer) PR
Shredder (detritivore) SH
Habitat relation Burrow BU
Climb CB
Cling CN
Sprawl SP
Swim SW
Page 64
53
Table 2.3: Site and reach scale variables sampled in late spring (sp) or summer (su) 2014
for sites in the Red River Valley, southern Manitoba, Canada. Variables not used in
analyses were considered redundant based on Pearson correlation with other variables of
the same scale.
Scale Variable Units Transformation Abbreviation
Site Total Nitrogen μg/L Natural log +1
Ammonia μg/L Natural log +1
Nitrite and Nitrate μg/L Natural log +1 NOsp or su
Total Phosphorus μg/L Natural log +1
Total Dissolved
Phosphorus
μg/L Natural log +1 TDPsp or su
Total Suspended
Solids
mg/L Natural log +1 TSSsp or su
Reach Canopy Cover % Logit CCsp or su
Wetted Width m Natural log +1 WetWidthSP or
SU
Average Depth m Natural log +1
Percent
Macrophytes Cover
Categories low to
high cover
None NoMacro,
LowMacro,
MedMacro,
FullMacro
Dominant Riparian
Vegetation
Categories None
Riparian Vegetation
Present
Categories None
Sinuosity channel length
over 1 km
distance (ratio)
None
Catchment area km Natural log + 1
Page 65
54
Table 2.4: Catchment scale human activity included in each Principal Component
Analysis (PCA) to determine Human Activity Gradients in southern Manitoba, Canada.
High correlation was determined by a correlation coefficient higher than 0.8.
PCA Human
activity
Units Transformation Correlation Abbreviation
Crop Wheat % Logit Barley
Canola Barley
Barley TBarley
Soybean TSoybean
Corn TCorn
Oats TOats
Forage TForage
Flaxseed TFlaxseed
Sunflowers TSunflower
Fallow TFallow
Beans TBeans
Potatoes TPotatoes
Livestock Beef nutrient
units/km2
Natural log+1 TBeef
Dairy cows TDairy.cows
Sheep dairy cow,
chicken
Pig TPig
Horse THorse
Goat TGoat
Bison dairy cow,
chicken
Chicken TChicken
Turkeys dairy cow,
chicken
Wastewater Lagoon
discharge
to streams
Pop/km2 Natural log+1 TWater
Lagoon
discharge
to land
TLand
Page 66
55
Table 2.5: Distribution (% frequency of occurrence among 20 catchments) and
descriptive statistics of human activity in study catchments of the Red River Valley, in
southern Manitoba, Canada.
Category Type Frequency
(%) Mean Median Max Min
Std.
Dev.
Crop (%) Forage 100 10.86 9.84 21.66 2.39 5.40 Wheat 100 16.78 16.13 33.81 4.99 8.88 Corn 100 4.54 1.76 17.58 0.22 5.34 Canola 100 13.35 10.98 29.92 1.11 10.74 Soybeans 100 11.94 11.89 28.88 1.77 8.21 Oats 95 0.66 0.44 2.50 0 0.70 Fallow 85 0.23 0.15 1.03 0 0.27 Barley 80 1.49 0.37 8.34 0 2.38 Flaxseed 80 0.19 0.10 0.93 0 0.26 Sunflowers 70 0.58 0.21 3.25 0 0.94 Beans 35 0.13 0.00 0.87 0 0.27 Potatoes 35 0.17 0.00 1.47 0 0.36
Livestock (NU*) Beef 100 1.65 1.49 3.93 0.11 1.24
Dairy
cows 100 1.29 0.51 7.53 0.00 2.07
Horse 100 0.31 0.29 0.72 0.08 0.20 Goat 100 0.02 0.01 0.11 0.00 0.02 Sheep 95 0.15 0.08 1.06 0 0.27 Pig 85 14.64 5.28 105.55 0 26.88 Chicken 85 0.97 0.18 8.15 0 2.22 Turkeys 60 0.07 0.00 0.84 0 0.22 Bison 15 0.01 0.00 0.10 0 0.03
Wastewater
(population/km2)
Discharge
to stream 45 5.93 0 53.64 0 14.28
Discharge
to land 25 0.22 0 1.69 0 0.48
*nutrient units/km2
Page 67
56
Table 2.6: Variance explained by Principal Component Analysis (PCA) axes of PCAs
performed on site, reach and catchment scale variables in subcatchments of southern
Manioba, Canada. (*) indicates axes not considered important based on the brokenstick
model (Legendre and Legendre 1998).
% Variance Explained
Scale PCA PC1 PC2 PC3
Site Water quality (spring) 68.2 16.6* 9.6*
Water quality (summer) 57.0 25.9 9.3*
Reach Habitat (spring) 31.7 21.0 14.7*
Habitat (summer) 31.3 27.1 14.2*
Catchment Crop 41.5 19.4 11.8*
Livestock 82.4 11.8* 0.04*
Wastewater 95.0 5.1* NA
Page 68
57
Table 2.7: Variance explained by axes (PC) of Principal Component Analyses of taxa or
trait assemblages sampled in spring or summer in southern Manitoba, Canada.
% variance explained
Assemblage Sampling time PC1 PC2 PC3
Taxa spring 15.0 12.7 9.9
summer 16.7 13.9 11.3
Trait spring 61.5 11.2 8.7
summer 56.4 14.8 10.1
Page 69
58
Table 2.8: Variance in taxa or trait assemblages, catchment scale human activity and
local environment factors sampled in spring or summer in southern Manitoba, Canada.
Variance is reported in terms of % constrained and % unconstrained variance of axes
(RDA) considered significant based on permutations tests, and the total unconstrained
variance.
% constrained variance % unconstrained variance
Assemblage Sampling RDA1 RDA2 RDA3 RDA1 RDA2 RDA3 Total
Taxa Spring 22.1 - - 13.8 - - 37.7
Summer 23.2 17.3 - 15.0 11.2 - 35.4
Trait Spring 49.0 - - 31.0 - - 36.7
Summer 44.5 18.7 12.5 32.0 13.5 9.0 28.0
Page 70
59
Figure 2.1: Conceptual diagram of broad mechanistic links between catchment scale
human activities, reach scale habitat, site scale water quality and trait categories in
freshwater streams. 1Total Suspended Solids
Page 71
60
Figure 2.2: Location of 20 sites and their subcatchments in southern Manitoba, Canada,
sampled in this study. Gray lines represent elevation contours. (Layers were extracted
from Geogratis.ca)
Page 72
61
Figure 2.3: Principal Component Analysis (PCA) of the type and intensity of a) crops,
b) livestock, and c) wastewater lagoons for 20 subcatchments in southern Manitoba,
Canada. Abbreviations are listed in Table 2.4.
a) b)
c)
Page 73
62
Figure 2.4: Principal Component Analysis (PCA) of site scale water quality in spring (a)
and summer (c), and reach scale habitat descriptors in spring (b) and summer (d) for 20
subcatchments in southern Manitoba, Canada. Abbreviations are listed in Table 2.3.
a)
d) c)
b)
Page 74
63
Figure 2.5: Principal
Component Analyses
(PCA) of taxa assemblage
in spring (a) and summer
(b) (closed circles)
measured at 18 (spring) or
20 (summer) study sites
(open circles) in southern
Manitoba in 2014. Taxa
abbreviation are listed in
Appendix 1. Taxa were
labeled by order of
importance to limit label
overlaps.
a)
b)
Page 75
64
Figure 2.6: Principal
Component Analyses
(PCA) of trait assemblage
in spring (a) and summer
(b) summer assemblage
(closed circles) sampled at
18 (spring) or 20 (summer)
study sites (open circles) in
southern Manitoba in 2014.
Trait abbreviation are listed
in Table 2.2.
a)
b)
Page 76
65
Figure 2.7: Redundancy Analysis (RDA) of presence-absence of aquatic insect genera
sampled in late spring 2014 in 18 sites of southern Manitoba, Canada. Taxa
abbreviations are listed in Appendix 1.
Page 77
66
Figure 2.8: Redundancy Analysis (RDA) of presence-absence of aquatic insect genera
sampled in summer 2014 in 20 sites of southern Manitoba, Canada. Taxa abbreviations
are listed in Appendix 1.
Page 78
67
Figure 2.9: Redundancy Analysis (RDA) of presence-absence of aquatic insect’ traits
sampled in late spring 2014 in 18 sites of southern Manitoba, Canada. Trait
abbreviations are listed in Table 2.2.
Page 79
68
Figure 2.10: Redundancy Analysis (RDA) of presence-absence of aquatic insect’ traits
sampled in summer 2014 in 20 sites of southern Manitoba, Canada. Trait abbreviations
are listed in Table 2.2
Page 80
69
Figure 2.11: Redundancy Analysis (showing RDA2 and RDA3) of presence-absence of
aquatic insect’ traits sampled in summer 2014 in 20 sites of southern Manitoba, Canada.
Trait abbreviations are listed in Table 2.2.
Page 81
70
3.0 Δ15N as a tracer of anthropogenic nitrogen sources in Landscapes of
Southern Manitoba, Canada.
3.1 Introduction
Agricultural activities are a source of excess nutrients that can alter stream
ecosystems, creating water quality concerns and altering the composition and abundance
of stream communities (Rabalais 2002, Allan 2004, Chambers et al. 2009). In
agricultural landscapes, nutrients are derived from both diffuse sources, primarily
synthetic and manure fertilizer, and direct pollution sources, in particular wastewater
discharge from rural villages and intensive livestock operations involving feedlots or
dairy barns (Carpenter et al. 1998). Surface waters receiving excess nutrients from
agricultural landscapes are often transformed to eutrophic, highly productive, algal-
dominated ecosystems. This is the case in southern Manitoba where intensive
agriculture in the Red River Valley (RRV) has contributed to eutrophication of Lake
Winnipeg (Bourne et al. 2002, Bunting et al. 2011, McCullough et al. 2012) and
negatively affected the fishery and tourism industries (Environment Canada and
Manitoba Water Stewardship 2011).
Watersheds with intensive agriculture, such as the RRV, typically have
subcatchments with varying degrees of human activities as a result of differences in both
Page 82
71
the types and magnitude of anthropogenic nutrient sources, and thus are considered to be
multiple stressor landscapes (Yates and Bailey 2010, Yates et al. 2012b). In the case of
the RRV, subcatchments are exposed to a varying extent of cash crops, swine and cattle
production, and rural villages (Yates et al. 2012b). Although the coverage of agricultural
land (e.g., % agriculture) in a subcatchment has often been used to represent the relative
exposure of a stream to anthropogenic stressors (Bailey et al. 2007), this metric may not
allow the identification and isolation of important sources of nutrients. Different
agricultural practices contribute different amount of nutrients (e.g., fertilizer application
on crops vs. runoff of manure from livestock production, Yates et al. 2012b), and
biochemical processes may affect the quantity and forms of nutrient lost from specific
sources (e.g., storage of swine manure in lagoons vs. manure from cattle spread on
pastures; Carpenter et al. 1998, Diebel and Vander Zanden 2009, Yang et al. 2011).
Moreover, nutrients reaching surface waters can be reduced by retention in soils, taken
up by terrestrial plants or lost to the atmosphere through nitrogen volatilization
(Carpenter et al. 1998, Chambers et al. 2001). Therefore, efforts to reduce excess
nutrients in downstream waterbodies need to consider identification of specific sources
of anthropogenic nutrient in a watershed.
Nitrogen isotopic signatures (δ15N), calculated as the ratio of light to heavy
nitrogen isotopes in a substance, is a useful tool for identifying the types and magnitude
Page 83
72
of anthropogenic nitrogen sources (Peterson and Fry 1987). Studies have shown that
δ15N in water, organic matter or aquatic animals can be used to identify exposure to
excess nitrogen from agriculture (e.g. Chang et al. 2002, Anderson & Cabana 2006,
Lefebvre et al. 2007, Peterson et al. 2007, Clapcott et al. 2012), and wastewater (e.g.
DeBruyn & Rasmussen 2002, Cole et al. 2005, Kaushal et al. 2011). However, few
studies have applied δ15N in multiple stressor environments to identify anthropogenic
nitrogen sources using fine spatial scale land use categories (but see Anderson &
Cabana 2006). Synthetic fertilizer, manure and wastewater frequently have different
δ15N signatures, making it possible to individually identify these sources if their isotope
signatures are sufficiently distinct from one another and the natural baseline (Bedard-
Haughn et al. 2003). In addition, biogeochemical transformations of nitrogen during
transport may widen the gap in δ15N among sources even before they are incorporated
into the food web (Bedard-Haughn et al. 2003, Anderson and Cabana 2005, Diebel and
Vander Zanden 2009). Moreover, Yates et al. (2014) showed that δ15N is a potential
tracer of anthropogenic nitrogen sources in catchments of southern Manitoba, Canada,
although they indicated that a broader gradient of exposure to human activities was
needed to validate the approach.
The objective of this study was to determine if δ15N of particulate organic matter
(POM) and benthic macroinvertebrates (BMI) could be used to identify excess nitrogen
Page 84
73
inputs from independent agricultural activities in streams of southern Manitoba, Canada.
BMI primary consumers were chosen as the food web component to evaluate δ15N as a
potential tracer of anthropogenic nitrogen sources in order to limit the influence of
fractionation due to trophic position within a food chain (Peterson and Fry 1987, Diebel
and Vander Zanden 2009). In addition, POM was used to represent the base of the food
chain, which is comprised of biofilm, periphyton, plant detritus and algae. Human
activity gradients (HAGs, Yates 2012b), representing the magnitude of different
nitrogen-producing activities (crops, livestock and wastewater) present in the RRV,
were developed to test mechanistic hypotheses throughout the possible range of
exposure (Figure 3.1). I predicted that direct sources of anthropogenic nitrogen would
enrich δ15N of BMIs and POM because cattle manure accumulated in pastures and
wastewater discharge tend to have a high δ15N compared to surface waters (Bedard-
Haughn et al. 2003). Furthermore, I expected that wastewater would have a stronger
relationship with δ15N because it is a direct source of nutrients, whereas manure in
pastures can lose nitrogen through biochemical processes (Diebel & Vander Zanden
2009, Yang et al. 2011). In addition, I did not predict a specific response from exposure
to excess nitrogen from cash crops because: (1) synthetic fertilizer has a low signature
compared to manure fertilizer (Bedard-Haughn et al. 2003); and (2) farmers may choose
to rely unequally on synthetic or manure fertilizer based on the price of synthetic
Page 85
74
fertilizer and the price of transporting manure between their crop and the closest manure
storage facility (Wiens et al. 2008).
3.2 Methods
3.21 Study area and experimental design
The study was conducted during spring (late May through early June) and
summer (late July through August) 2014 in 20 catchments located in the agriculturally-
dominated landscape of the RRV in southern Manitoba, Canada (previously described in
Section 2.2). POM and BMI were sampled at all sites with a three-minute travelling kick
and sweep method using a kick net (400 µm mesh) in all available habitats including
those colonized by macrophytes. The samples were preserved in 95% denatured Ethyl
alcohol (Histoprep) and later processed in the laboratory where they were subsampled
and the BMI sorted and identified to lowest taxonomic level possible (i.e., most often
genus or species) (Section 2.2).
Primary consumers were represented by the genera Hyalella and Caenis, and
family Baetidae, all of which are collector-gatherers, and were selected because these
taxonomic groups were present at most sites with sufficient biomass to perform stable
isotope analysis (approximately 1 mg of dried tissue; Jardine et al. 2003). Baetidae
isotopic values were comprised of organisms that could not be identified to a lower
Page 86
75
taxonomic resolution or represented the average isotopic values of organisms identified
at family and genus levels. Fine and coarse POM (FPOM, CPOM respectively; FPOM
>250 μm to ≤ 1 mm and CPOM > 1 mm) were sampled by sieving vegetative matter
collected in BMI kick samples after removal of all animals. Because of time limitation,
only summer POM samples were analysed. However, the signature of POM was not
expected to change significantly over the year based on findings from Cole et al. (2005).
Periphyton or benthic algae were not sampled because of a lack of hard surfaces in the
streams. BMI primary consumer and POM samples were rinsed with distilled water,
dried at 60ᵒC for a minimum of 48 h, ground and analyzed for δ15N using a Carlo Erba
NC2500 or Costech 4010 Elemental Analyzer and a Delta Plus or Delta XP continuous
flow isotope-ratio mass spectrometer (CF-IRMS, Thermo-Finnigan, Bremen, Germany;
samples processed by the Stable Isotopes in Nature Laboratory at the University of New
Brunswick, Fredericton, Canada).
Human activity gradients were calculated based on geospatial information for the
type and extent of human activity in the study catchments (Section 2.2). Each category
of human activity (crops, livestock or wastewater) was summarized with separate
Principal Component Analyses (PCA, Table 2.6, and Figure 2.3). Axis scores were used
as quantitative gradients (Yates et al. 2012b) and were selected using the broken stick
model (Jackson 1993). The resulting five gradients represent specific types of activity
Page 87
76
with varying intensity among sites: small grain (barley and correlated wheat, canola;
Crop1); sunflower and oat crops (Crop 2) (Figure 2.3a); swine production (Live1); beef
cattle production (Live2) (Figure 2.3b); and wastewater lagoons discharging to streams
(WWT) (Figure 2.3c). The second axis of the wastewater PCA, which represented
lagoons serving small settlements that use the waste as manure fertilizer, was not
included as a gradient in further analyses. This decision was made because small
lagoons that served a maximum of 190 people represented a relatively small source of
nitrogen compared to large wastewater lagoons (i.e., serving a population of 122 to
14000, Table 2.5) and this HAG (lagoon waste applied as fertilizer) only explained 5%
of the variation in sites exposed to wastewater (Table 2.6).
3.22 Data analysis
Single and multiple linear regression analyses were used to evaluate a priori
hypotheses describing the relationship among HAGs and the δ15N of POM or primary
consumers (Table 3.1). Spring and summer samples were analysed separately to limit
the influence of possible seasonal variation in nitrogen signature of BMI tissue (Jardine
et al. 2003). All statistical assumptions were checked using standard graphical methods
(Quinn and Keough 2002). The data were skewed due to statistical outliers in the
response variables, particularly for the WWT HAG where most sites were at one end of
the gradient. Raw values of δ15N were used because the statistical outliers could not be
Page 88
77
considered incorrect values and transforming response variables did not improve
normality of the data (Quinn and Keough 2002). The absence or lack of sufficient tissue
for stable isotope analysis resulted in a decrease in sample size for all primary consumer
groups, with the largest reduction being 8 sites for Caenis in the summer (i.e., from 20
to 12 sites; Table 3.2). In addition, the CPOM sample for LR05 was lost because of a
technical error during stable isotope analysis.
For each POM type and primary consumer taxa, 7 models including a null and
global model were evaluated using the corrected Akaike Information Criterion (AICc),
which is appropriate for small sample sizes (Anderson 2010). Interactions between the
swine (Live1) and two crop (Crop1 and 2) HAGs were used to represent the non-
independence between manure from swine production and crops because swine
operations store all manure to apply to nearby fields as fertilizer (Figure 3.1). Some beef
cattle operations also store manure to apply as fertilizer, but as it was not always the
case (e.g., feedlots where manure is spread on pasture), and because of small sample
size, three way interactions with the beef HAG (Live2) were not included. The model
with lowest AICc score was considered best, although other models were plausible if
they remained within 7 Δ AICc units of the best model (Anderson 2010). In addition,
models with possible pretending variables (i.e., within approximately 2 Δ AICc units of
the best model) were removed if they were considered redundant based on guidelines
Page 89
78
from Anderson (2010; see Appendix 4 for results including redundant models). All data
analyses were completed with the statistical software R version 3.1.3 (R Core Team
2015).
3.3 Results
The range of δ15N of POM and primary consumers was, on average, from 3.9 to
17.2 ‰ among sites (Figure 3.2). Sites with an average δ15N greater than 10 ‰ were
LR02, RT04 and UR04, the three sites with the highest WWT scores, and UR05 with
high crop production (Table 2.1). The mayfly genus Caenis when sampled in summer
had the greatest range of δ15N among sites compared to other primary consumers and
POM (Table 3.2). In contrast, the mayfly family Baetidae had the smallest range across
both sampling times (8 ‰ range in spring, Table 3.2).
With the exception of Baetidae and Hyalella in spring, variation in δ15N of
primary consumer taxa and POM was best explained by the WWT model, which
described the intensity of wastewater discharge in catchments (Tables 3.3 - 3.5).
Moreover, WWT was positively associated with the δ15N of Caenis in spring, as well as
all BMI in summer (Figure 3.3) and both types of POM (Figure 3.4). Furthermore, there
was no support for models ascribing an influence of crops or swine production (Tables
3.3 - 3.5). Baetidae in spring also had the weakest relationship with any human activity
because the best model was the null model (Table 3.3). In contrast, the Live2 + WWT1
Page 90
79
model was by far the best model explaining variation in δ15N of Hyalella in spring
(Table 3.3). While WWT enriched the δ15N of spring Hyalella (Figure 3.3), Live2 was
negatively associated with this genus’ δ15N when controlling for WWT (Figure 3.5). All
other models were more than 7 Δaic from the best model and thus not considered
plausible for Hyalella in spring (Table 3.3). Although Live2+WWT1 also appeared to
be a plausible model for CPOM (Appendix 4), diagnostic plots revealed that the
relationship was created by a point with high leverage (Figure 3.6). Removing this
outlier from the model set demonstrated that Live2 was a pretending variable, and thus
the redundant model, Live2+WWT1, was removed from the analysis for CPOM
(Anderson 2010).
3.4 Discussion
3.4.1 Response of δ15N to point and non-point sources of nitrogen
Analysis of nitrogen pathways in 20 catchments in the agriculturally-dominated
landscape of the Red River Valley of southern Manitoba revealed that wastewater
appears to be a more important source of nitrogen than either manure or synthetic
fertilizer for these small streams. In fact, δ15N values of particulate organic matter (both
CPOM and FPOM) and most primary consumers (e.g., collector-gatherers) were only
associated with the wastewater HAG. The high nitrogen signature of both POM and
Page 91
80
primary consumers supports the hypothesis that wastewater discharge is a source of
bioavailable nitrogen that enriches the nitrogen signature of food sources. This
conclusion is consistent with observations by Chesworth et al. (in review) that
wastewater discharge from lagoons in the RRV contributed substantial quantities of
bioavailable nutrients to a RRV stream. Wastewater discharge was also shown to be a
more important source of nutrients to Dead Horse Creek than non-point sources
(corresponding to catchment UR04 of this study; Carlson et al. 2013). The results from
this study also corroborate studies in other aquatic ecosystems that have shown a
positive relationship between exposure to wastewater discharge and δ15N of biota (Cole
et al. 2005, Ulseth & Hershey 2005, Kaushal et al. 2011). Wastewater lagoon discharge
in southern Manitoba streams is episodic, permitted in only spring and summer
(Government of Manitoba, personal communication), and results in a temporary
increase in nutrients (Chesworth et al. in review). Nevertheless, results indicate that
these episodic increases in nitrogen are of sufficient magnitude and availability to create
a lasting influence on the food web because δ15N values of both spring and summer
organisms demonstrated clear enrichment from wastewater.
Occasional exceptions to the unique association of δ15N to wastewater were
observed in this study. In spring, the δ15N of the collector-gatherer, Hyalella, was
influenced by both wastewater discharge and manure from beef cattle operations. This
Page 92
81
suggests that manure from cattle operations also contributes to nitrogen assimilated by
biota in southern Manitoba. Manure from beef cattle operation is considered a point
source of nitrogen in the study area because it is not often stored and applied as fertilizer
for cash crops, but rather left on feedlots or pasture (Government of Manitoba personal
communication, Cade-Menun et al. 2013) and has the potential to runoff directly to
streams. This source of nitrogen was associated with a depletion of δ15N of Hyalella
tissues, in contrast to what was predicted. Manure has been found to enrich δ15N of biota
in other ecosystems because biochemical processes such as volatilization during storage
create a high δ15N signature of manure before it reaches streams (Bedard-Haughn et al.
2003, Anderson and Cabana 2006). Therefore, it is possible that the negative association
between δ15N and beef cattle production in this study is due to a lower rate of
volatilization from lack of storage or fast transportation to streams. However, this source
of nitrogen does not appear to be of uniform importance for the stream ecosystems of
the RRV: only one collector gatherer (i.e., Hyalella) was associated with manure from
beef cattle operation. In addition, the effect of cattle operation was important in spring
only, suggesting that nitrogen fractionation and transport of this point source may differ
between spring, when peak flows due to snowmelt and surface runoff have just
subsided, and summer, when low or no flow limits transport of nutrients (Yates et al.
2014). Another exception was the lack of association between δ15N of Baetidae in spring
Page 93
82
and any human activity. This finding was surprising because Baetidae were expected to
have a similar relationship with the HAGs as other collector-gatherers, such as Caenis,
that were strongly associated with wastewater discharge.
Unlike wastewater discharge, results suggest that synthetic and manure fertilizer
did not contribute significantly to nitrogen incorporation into food webs of southern
Manitoba streams. The lack of response in δ15N values of POM and most primary
consumers to these two agricultural practices suggests either that a considerable quantity
of nitrogen is lost from these sources before reaching the streams, or that nitrogen from
these sources is not incorporated into food webs once it reaches the streams.
Biochemical and microbial processes during storage, transport, and application of
fertilizer often result in a loss of nitrogen through retention in soil or ground water,
nitrogen volatilization, and uptake by terrestrial plants (Chambers et al. 2001). For
example, volatilization can cause nitrogen losses of up to 70% for swine manure stored
in lagoons (Yang et al. 2011). The δ15N pool of manure and synthetic fertilizer after
such biochemical or microbial transformation, combined with blurring of the δ15N
values when both manure and synthetic fertilizer are applied to the same field, makes it
difficult to isolate these two sources using δ15N (Diebel and Vander Zanden 2009). In
addition, severe hydrologic events may move nitrogen out of a subcatchment before it
can be assimilated. In southern Manitoba, snowmelt accounts for the majority of annual
Page 94
83
discharge and nitrogen concentrations at that time are considerably higher compared to
other seasons (Corriveau et al. 2011, Rattan et al. under review). Dissolved forms of
nitrogen in snowmelt waters are likely not assimilated by the biota because of minimal
biological activity at that time due to cold temperatures (Yates et al. 2012b). Therefore,
catchment scale agricultural activities may not be an important source of nitrogen for
food webs in southern Manitoba streams because of a disconnect between the timing of
supply (snowmelt) versus demand (summer), yet these sources likely contribute to the
overall nutrient status of downstream Lake Winnipeg.
3.4.2 δ15N of BMI and POM as bioindicator in the RRV
Results of this study show that δ15N of BMI and POM are useful in detecting
point sources of nitrogen, notably WWT, in southern Manitoba, but watershed-specific
factors limits its use in identifying catchment scale fertilizer application. The success of
δ15N as a source-tracking method relies heavily on watershed-specific parameters, such
as the difference in isotopic signatures of nitrogen sources and sinks (Bedard-Haughn et
al. 2003), and the potential for biogeochemical processes (Anderson & Cabana 2006,
Diebel and Vander Zanden 2009). The clear enrichment effect on δ15N of BMI primary
consumers and POM in response to wastewater lagoon discharge showed that this point
source is easily identified by this bioindicator. Wastewater discharge is a direct source
of nutrient to streams in the RRV (Yates 2012b, Chesworth 2016), and results of this
Page 95
84
study indicate that δ15N is able to identify this source likely because of its direct input to
streams, and its independence from catchment hydrology. Although manure from cattle
did not have as important an effect as wastewater, it was detected by δ15N of Hyalella in
spring, suggesting that δ15N can identify sources of manure from feedlots or pastures.
Moreover, future studies may be able to detect this nitrogen source using smaller scale
designs similar to Karr et al. 2001. In contrast, catchment scale synthetic and manure
fertilizer application was not reflected in the δ15N signatures of BMI primary consumers
and POM. These diffuse sources of nitrogen are therefore either not an important source
of nitrogen to RRV streams ecosystems (due to loss before reaching the streams) or their
signatures were not distinct enough from the natural δ15N baseline to be identified.
3.4.3 Conclusion
Results showed that δ15N of primary consumers and POM can serve to identify
the nitrogen signature of wastewater and beef cattle operations in the RRV, but are not
good bioindicators for nitrogen applied to crops. I found evidence that wastewater
discharge is a significant source of nitrogen for biota in small streams in the RRV, and
that nitrogen from cattle operations may also be important. However, I was unable to
detect the contribution of synthetic fertilizers or manure fertilizer applied to crops to
nitrogen assimilated in the food web. Therefore, findings suggest that δ15N is an
efficient diagnostic bioindicator of wastewater discharge, but future studies are needed
Page 96
85
to clarify the importance of other potential nitrogen sources. It is possible that the
contribution of nitrogen from agricultural activities in the RRV is limited, or that the
signature from these activities cannot be distinguished in these catchments. In addition,
it is not yet clear if wastewater discharge is a significant source of nutrients for Lake
Winnipeg, or if it is completely assimilated within the stream network. However, this
study demonstrates that δ15N of biota is a promising tool to answer these questions.
3.5 References
Allan J.D. (2004) Landscapes and riverscapes: the influence of land use on stream
ecosystems. Annual Review of Ecology, Evolution, and Systematics 35, 257–284.
Anderson C. & Cabana G. (2006) Does delta 15N in river food webs reflect the intensity
and origin of N loads from the watershed? The Science of the total environment
367, 968–978.
Anderson C. & Cabana G. (2005) 15 N in riverine food webs: effects of N inputs from
agricultural watersheds. Canadian Journal of Fisheries and Aquatic Sciences 340,
333–340.
Anderson D.R. (2008) Model based inference in the life sciences: a primer on evidence.
Springer Science and Business Media, New York, USA.
Bailey R.C., Reynoldson T.B., Yates A.G., Bailey J. & Linke S. (2007) Integrating
stream bioassessment and landscape ecology as a tool for land use planning.
Freshwater Biology 52, 908–917.
Bedard-Haughn A., van Groenigen J.W. & van Kessel C. (2003) Tracing 15N through
landscapes: potential uses and precautions. Journal of Hydrology 272, 175–190.
Page 97
86
Bourne A., Armstrong N. & Jones G. (2002) A preliminary estimate of total nitrogen
and total phosphorus loading to streams in Manitoba, Canada. Winnipeg, Canada.
Bunting L., Leavitt P., Wissel B., Laird K.R., Cumming B.F., St. Amand A., et al.
(2011) Sudden ecosystem state change in Lake Winnipeg, Canada, caused by
eutrophication arising from crop and livestock production during the 20th century.
Winnipeg, Canada.
Cade-Menun B.J., Bell G., Baker-Ismail S., Fouli Y., Hodder K., McMartin D.W., et al.
(2013) Nutrient loss from Saskatchewan cropland and pasture in spring snowmelt
runoff. Canadian Journal of Soil Science 93, 445–458.
Carlson J.C., Anderson J.C., Low J.E., Cardinal P., MacKenzie S.D., Beattie S.A., et al.
(2013) Presence and hazards of nutrients and emerging organic micropollutants
from sewage lagoon discharges into Dead Horse Creek, Manitoba, Canada. Science
of The Total Environment 445-446, 64–78.
Carpenter S.R., Caraco N.F., Correll D.L., Howarth R.W., Sharpley A.N. & Smith V.H.
(1998) Nonpoint pollution of surface waters with phosphorus and nitrogen.
Ecological Applications 8, 559–568.
Chambers P.A., Guy M., Dixit S.S., Benoy G.A., Brua R.B., Culp J.M., et al. (2009)
Nitrogen and phosphorus standards to protect the ecological condition of
Canadian streams, rivers and coastal waters. National Agri-Environment
Standards Initiative Synthesis Report No. 11. Environment Canada, Gatineau,
Quebec.
Chambers P.A., Guy M., Roberts E.S., Charlton M.N., Kent R., Gagnon C., et al. (2001)
Nutrients and their impact on the Canadian environment. Agriculture and Agri-
Food Canada, Environment Canada, Fisheries and Oceans Canada, Health Canada
and Natural Resources Canada.
Chang C.C.., Kendall C., Silva S.R., Battaglin W.A. & Campbell D.H. (2002) Nitrate
stable isotopes: tools for determining nitrate sources among different land uses in
the Mississippi River Basin. Canadian Journal of Fisheries and Aquatic Sciences
59, 1874–1885.
Page 98
87
Chesworth C.T. (2016) Lagoon wastewater effluent impacts stream metabolism in Red
River tributaries. Electronic Thesis and Dissertation Repository Paper 3483.
University of Western Ontario.
Chesworth C.T., Brua R.B., Chambers P.A., Culp J.M. & Yates A.G. (in review)
Stream metabolism is affected by pulse release of municipal wastewater effluent.
Clapcott J.E., Collier K.J., Death R.G., Goodwin E.O., Harding J.S., Kelly D., et al.
(2012) Quantifying relationships between land-use gradients and structural and
functional indicators of stream ecological integrity. Freshwater Biology 57, 74–90.
Cole M.L., Kroeger K.D., McClelland J.W. & Valiela I. (2005) Macrophytes as
indicators of land-derived wastewater: application of a δ 15 N method in aquatic
systems. Water Resources Research 41.
Corriveau J., Chambers P. A., Yates A.G. & Culp J.M. (2011) Snowmelt and its role in
the hydrologic and nutrient budgets of prairie streams. Water Science &
Technology 64, 1590–1596.
DeBruyn A.M.H. & Rasmussen J.B. (2002) Quantifying assimilation of sewage-derived
organic matter by riverine benthos. Ecological Applications 12, 511–520.
Diebel M.W. & Vander Zanden M.J. (2009) Nitrogen stable isotopes in streams: effects
of agricultural sources and transformations. Ecological applications 19, 1127–
1134.
Environment Canada & Manitoba Water Stewardship (2011) State of Lake Winnipeg:
1999 to 2007. Winnipeg, MB.
Jackson D.A. (1993) Stopping rules in principal components analysis: a comparison of
heuristical and statistical approaches. Ecology 74, 2204–2214.
Jardine T.D., McGeachy S., Paton C., Savoie M. & Cunjak R.A. (2003) Stable isotopes
in aquatic systems: sample preparation, analysis, and interpretation. Canadian
Manuscript Report of Fisheries and Aquatic Sciences No. 2656.
Page 99
88
Karr J.D., Showers W.J., Gilliam J.W. & Andres A.S. (2001) Tracing nitrate transport
and environmental impact from intensive swine farming using delta nitrogen-15.
Journal of Environment Quality 30, 1163–1175.
Kaushal S.S., Groffman P.M., Band L.E., Elliott E.M., Shields C.A. & Kendall C.
(2011) Tracking nonpoint source nitrogen pollution in human-impacted
watersheds. Environmental Science & Technology 45, 8225–8232.
Lefebvre S., Clément J.-C., Pinay G., Thenail C., Durand P. & Marmonier P. (2007) 15N-Nitrate Signature in Low-Order Streams : Effects of Land Cover and
Agricultural Practices. Ecological Society of America 17, 2333–2346.
McCullough G.K., Page S.J., Hesslein R.H., Stainton M.P., Kling H.J., Salki A.G., et al.
(2012) Hydrological forcing of a recent trophic surge in Lake Winnipeg. Journal of
Great Lakes Research 38, 95–105.
Peterson B.J. & Fry B. (1987) Stable isotopes in ecosystem studies. Annual Review of
Ecology and Systematics 18, 293–320.
Peterson G.S., Sierszen M.E., Yurista P.M. & Kelly J.R. (2007) Stable nitrogen isotopes
of plankton and benthos reflect a landscape-level influence on Great Lakes coastal
ecosystems. Journal of Great Lakes Research 33, 27–41.
Quinn G.P. & Keough M.J. (2002) Experimental design and data analysis for biologists.
Cambridge University Press, New York, USA.
R Core Team (2015) R: A language and environment for statistical computing.
Rabalais N.N. (2002) Nitrogen in aquatic ecosystems. Ambio 31, 102–112.
Rattan K.J., Corriveau J.C., Brua R.B., Culp J.M., Yates A.G. & Chambers P.A. (in
review) Quantifying seasonal variation in total phosphorus and nitrogen from
prairie streams in the Red River Basin, Manitoba Canada.
Ulseth A. & Hershey A. (2005) Natural abundances of stable isotopes trace
anthropogenic N and C in an urban stream. Journal of the North American
Benthological Society 24, 270–289.
Page 100
89
Wiens M.J., Entz M.H., Wilson C. & Ominski K.H. (2008) Energy requirements for
transport and surface application of liquid pig manure in Manitoba, Canada.
Agricultural Systems 98, 74–81.
Yang J.Y., Huffman E.C., Drury C.F., Yang X.M. & De Jong R. (2011) Estimating the
impact of manure nitrogen losses on total nitrogen application on agricultural land
in Canada. Canadian Journal of Soil Science 91, 107–122.
Yates A.G. & Bailey R.C. (2010) Improving the description of human activities
potentially affecting rural stream ecosystems. Landscape Ecology 25, 371–382.
Yates A.G., Brua R.B., Corriveau J., Culp J.M. & Chambers P.A. (2014) Seasonally
driven variation in spatial relationships between agricultural land use and in-stream
nutrient concentration. River Research and Applications 30, 476–493.
Yates A.G., Brua R.B., Culp J.M. & Chambers P.A. (2012a) Multi-scaled drivers of
rural prairie stream metabolism along human activity gradients. Freshwater
Biology 58, 675–689.
Yates A.G., Culp J.M. & Chambers P.A. (2012b) Estimating nutrient production from
human activities in subcatchments of the Red River, Manitoba. Journal of Great
Lakes Research 38, 106–114.
Page 101
90
Table 3.1: A priori hypotheses of how δ15N of particulate organic matter and primary
consumers may be affected by human activity gradients in subcatchments of the Red
River Valley of southern Manitoba, Canada. The ordinate intercept is identified by (I).
The PCA axes used as predictors are coded using “crop” (crop types), “live” (type of
livestock densities) and “wwt” (wastewater) with PC1 indicating the first axis or PC2,
the second axis.
Model Name Hypothesis Model Structure
Crop1*Live1 Influence of Small Grain
HAG
= β₁(cropPC1) + β₂(livePC1) +
β3(cropPC1) (livePC1) + β4(I)
Crop2*Live1 Influence of Sunflower and
Oats HAG
= β₁(cropPC2) + β₂(livePC1) +
β3(cropPC2)(livePC1) + β4(I)
Live2 Influence of Beef HAG =β₁(livePC2) + β₂(I)
WWT Influence of Wastewater
HAG =β₁(wwtPC1)+β₂(I)
Live2 +
WWT Influence of point sources = β₁(wwtPC1)+ β2(livePC2)+ β3(I)
Null δ15N does not respond to
HAGs = β₁(I)
Global Model with all predictors
= β₁(cropPC1) + β₂(cropPC2) +
β3(livePC1) + β4(livePC2) +
β5(wwtPC1)+ β6(I)
Page 102
91
Table 3.2: Descriptive statistics of δ15N values (‰) for particulate organic matter and
primary consumers sampled during both spring and summer in streams of southern
Manitoba, Canada.
Sampling period Sample Mean Median SD Min Max n
Spring Hyalella 8.10 7.02 3.89 2.97 14.27 13
Caenis 7.36 6.47 3.22 4.20 15.26 15
Baetidae 6.50 6.40 2.35 2.65 10.65 16
Summer Hyalella 8.62 7.82 5.52 2.54 24.68 14
Caenis 10.25 8.03 6.92 5.66 31.42 12
Baetidae 8.37 8.27 2.95 2.46 13.25 15
FPOM* 6.76 5.97 2.77 4.05 14.21 20
CPOM** 7.09 5.75 3.73 3.87 18.39 19
*Fine Particulate Organic Matter, ** Coarse Particulate Organic Matter
Page 103
92
Table 3.3: Comparison of a priori tested models for predicting δ15N changes in primary
consumer taxa sampled in late spring for subcatchments in southern Manitoba, Canada,
using corrected Akaike Information Criterion (AICc).
Response Model AICc Delta AIC Model weight Evidence ratio
Hyalella Live2+Wwt1 58.1 0.0 0.999 1
Wwt1 72.6 14.6 0.001 1466
Crop2*Live1 73.5 15.4 0.000 2231
Null 76.4 18.3 0.000 9557
Global model 76.7 18.6 0.000 11159
Live2 78.2 20.1 0.000 23272
Crop1*Live1 84.9 26.8 0.000 663312
Caenis Wwt1 71.0 0.0 0.994 1
Null 81.6 10.7 0.005 206
Live2 84.8 13.8 0.001 1007
Crop1*Live1 87.7 16.8 0.000 4337
Crop2*Live1 88.4 17.4 0.000 6063
Global model 88.5 17.5 0.000 6342
Baetidae Null 76.6 0.0 0.459 1
Wwt1 77.2 0.5 0.356 1
Live2+Wwt1 78.9 2.3 0.145 3
Crop2*Live1 82.9 6.2 0.020 23
Crop1*Live1 83.1 6.5 0.018 25
Global model 89.9 13.2 0.001 739
Page 104
93
Table 3.4: Comparison of a priori tested models for predicting δ15N changes in primary
consumer taxa sampled in summer for subcatchments in southern Manitoba, Canada,
using corrected Akaike Information Criterion (AICc).
Response Model AICc Delta AIC Model weight Evidence ratio
Hyalella Wwt1 78.8 0.0 0.998 1
Null 91.6 12.8 0.002 602
Live2 94.2 15.4 0.000 2208
Global model 96.4 17.6 0.000 6601
Crop1*Live1 100.1 21.3 0.000 42193
Crop2*Live1 102.6 23.7 0.000 142914
Caenis Wwt1 79.9 0.0 0.898 1
Null 84.8 4.9 0.079 11
Live2 88.4 8.5 0.013 70
Global model 88.8 8.9 0.010 86
Crop1*Live1 96.6 16.7 0.000 4209
Crop2*Live1 99.0 19.1 0.000 13698
Baetidae Wwt1 77.0 0.0 0.959 1
Null 83.9 6.9 0.031 31
Live2 86.3 9.3 0.009 106
Crop2*Live1 91.3 14.2 0.001 1236
Crop1*Live1 92.6 15.6 0.000 2392
Global model 95.8 18.8 0.000 11849
Page 105
94
Table 3.5: Comparison of a priori tested models for predicting δ15N fine or coarse
particulate organic matter (FPOM, CPOM respectively) sampled in summer for
subcatchments in southern Manitoba, Canada, using corrected Akaike Information
Criterion (AICc).
Response Model AICc Delta AIC Model weight Evidence ratio
FPOM Wwt1 86.4 0.0 0.997 1
Global model 98.8 12.4 0.002 483
Null 101.1 14.7 0.001 1580
Live2 103.7 17.3 0.000 5653
Crop1*Live1 106.4 20.0 0.000 22137
Crop2*Live1 107.4 21.0 0.000 35954
CPOM Wwt1 92.8 0.0 0.997 1
Global model 105.4 12.6 0.002 539
Null 107.6 14.8 0.001 1636
Live2 110.4 17.6 0.000 6568
Crop1*Live1 112.9 20.1 0.000 22925
Crop2*Live1 115.1 22.3 0.000 68872
Page 106
95
Figure 3.1: Conceptual model of δ15N changes from landscape nitrogen sources to the
food web. (+) indicates an enrichment in δ15N, while (-) indicates a depletion. Because
the influence of both crops is dependent upon whether livestock manure or synthetic
fertilizer is the dominant source of nitrogen applied on the fields, the relationship
between crops and δ15N of the food web is difficult to predict. This uncertainty is
denoted by (?).
Page 107
96
Figure 3.2:
Mean δ15N
(± 1SD) of
primary
consumers
and
particulate
organic
matter
sampled
during
spring and
summer
2014 in
streams of
southern
Manitoba,
Canada.
Page 108
97
Figure 3.3: Relationship between the δ15N of all BMI and the wastewater HAG (wwtPC1) sampled in spring or summer in
subcatchments of southern Manitoba, Canada.
Adj. R2=0.37 Adj. R
2=0.57 Adj. R
2=0.09
Adj. R2=0.65 Adj. R
2=0.46 Adj. R
2=0.42
Page 109
98
Figure 3.4: Relationship between the δ15N of particulate organic matter (fine, FPOM;
coarse, CPOM) and the wastewater HAG (wwtPC1) sampled in spring or summer in
subcatchments of southern Manitoba, Canada.
Adj. R2=0.58
Adj. R2=0.56
Page 110
99
Figure 3.5: Conditional plots of δ15N of Hyalella in spring and (a) beef cattle production
(livePC2) while controlling for wwtPC1, or (b) wastewater HAG (wwtPC1) while
controlling for livePC2. Shaded areas represent confidence bands (Breheny and Burchett
2013).
a) b)
Page 111
100
Figure 3.6: Conditional plot of δ15N of Coarse Particulate Organic Matter (CPOM) and
beef cattle production (livePC2) while controlling for wwtPC1 with (a) and without (b)
UR05. Shaded areas represent confidence bands (Breheny and Burchett 2013).
a) b)
Page 112
101
4.0 General conclusion
4.1 Stated objectives
This thesis aimed to evaluate the potential of two functional bioindicators,
biological traits and δ15N of benthic macroinvertebrates (BMI), to identify and diagnose
catchment scale agricultural impacts in streams of the Red River Valley (RRV). Chapter
2 compared and contrasted trait and taxa beta diversity, and examined the sensitivity of
these community measures to human activities and natural gradients in the RRV.
Chapter 3 identified and described the relationship of δ15N values of aquatic insects (i.e.,
collector-gatherers) to anthropogenic nitrogen input from specific sources. Human
activity gradients (HAGs) based on Yates et al. (2012) were used in Chapters 2 and 3 to
estimate exposure of the stream ecosystem to specific catchment scale agricultural
activities, such as land cover of different types of crops, and to wastewater from
municipal lagoons, which represented the only significant non-agricultural human
activity in the study area.
4.2 Drivers of trait and taxa beta diversity
I predicted in Chapter 2 that trait and taxa beta diversity would be affected by
catchment scale human activities through alteration of the local environment. Moreover,
human activities were expected to change aquatic insect assemblages by altering the
Page 113
102
reach scale factors of hydrology and riparian condition, and site scale water quality
(Allan 2004, Moss 2008, Blann et al. 2009). However, the predicted pathway of human
impact was not supported. Instead, I found that local habitat factors unrelated to
catchment scale human activity were the primary drivers of trait and taxa beta diversity.
Although aquatic assemblages were associated with catchment scale agriculture in
spring, traits did not respond as expected along this gradient of land use. Because wetted
width was also an important factor in spring, I hypothesize that historical channelization
of streams in the study area created a geomorphological gradient, represented by wetted
width that confounded the effects of land use. Flow conditions during the snowmelt
period may have masked agricultural effects as well, which is likely due to the
considerable discharge increase during this period (Corriveau et al. 2011). For summer
assemblages, riparian condition (i.e., canopy cover) and dissolved phosphorus were the
primary drivers of trait and taxa assemblages. As predicted, catchment scale agricultural
activities were an independent, secondary driver that affected trait assemblages through
changes in total suspended solid inputs. However, this secondary gradient explained
only a small amount of variation in trait assemblages. Therefore, local environment
factors have a stronger effect on aquatic insect communities in the RRV than catchment
land use during low flow conditions.
Page 114
103
Results of this study also revealed that biological traits had similar responses to
natural and anthropogenic gradients compared to the taxonomic approach, but provided
the advantage of being easily predictable and interpretable. Both approaches showed
responses to factors that were seasonally dependent, indicating that trait-based
approaches can vary across season in environments with strong seasonal habitat
patterns. In addition, trait beta diversity was not more sensitive to human impacts than
taxa beta diversity. Instead, it was more sensitive to important local environment factors.
These results are consistent with studies that have found local abiotic factors to be an
important filter of traits (Richards et al. 1997, Sponseller et al. 2001, Strayer et al. 2003,
Heino 2004, Heino et al. 2007).
4.3 Response of δ15N of BMIs to anthropogenic nitrogen
Testing the response of δ15N of BMI to gradients of human activities in the RRV
confirmed my prediction that wastewater was a direct source of nitrogen to these food
webs. Wastewater discharge had a clear enrichment effect on the δ15N of BMI, as found
in other ecosystems (Cole et al. 2005, Ulseth and Hershey 2005, Kaushal et al. 2011).
These results provide evidence that wastewater discharge is a source of biologically
available nitrogen, as confirmed by Chesworth (2016). In addition, I predicted that
manure runoff from beef cattle operation would also be a point source of nitrogen;
however, only the δ15N of Hyalella in spring was associated with this human activity,
Page 115
104
suggesting that manure left on pastures and feedlots is not as important as wastewater
discharge. Exposure to nitrogen from synthetic and manure fertilizer applied to crops (as
estimated by crop and swine production) was also not associated with δ15N of BMI. I
hypothesized that the lack of clear relationship between δ15N of BMI and catchment
scale agricultural activities was due to significant nitrogen loss during transport to
streams (e.g., through retention in soil, volatilization or uptake by terrestrial plants),
hydrological factors controlling the transport of nutrients, and/or the mixing of high and
low δ15N signatures from manure and synthetic fertilizer respectively. The hydrological
conditions in spring and summer may limit availability of nitrogen to stream
ecosystems. In spring, high discharge associated with snowmelt may transport nitrogen
out of the subcatchment before it can be assimilated (Yates et al. 2012b), while in
summer, there is little catchment scale transport of nutrients because of a lack of
hydrological connectivity within catchments (Yates et al. 2014a). Therefore, δ15N of
BMI is an efficient diagnostic bioindicator for wastewater discharge likely because this
source of nitrogen is independent from biochemical transformation during transport and
catchment hydrology.
4.4 Research implications
My results suggest that impacts of catchment scale agricultural activities on
aquatic communities in streams dissecting the RRV were seasonal, and also limited
Page 116
105
likely because of hydrological factors. Although the associations were not always clear,
both trait beta diversity and δ15N of BMI were related to catchment scale agriculture in
spring, while relationships were weaker or lacking in summer. These results provide
evidence that there is little impact from catchment scale agricultural activities during
periods of low flow likely because of a lack of hydrological connectivity within
catchments during this period as suggested by Yates et al. (2014). The weak or lack of
relationship between agricultural land use and stream ecosystems in summer has also
been found by studies assessing other indicators for southern Manitoba: stream
metabolism (Yates et al. 2012a), aquatic communities and δ15N of primary producers
and consumers (Yates et al. 2014b), aquatic macrophytes composition (Tyrrell 2015), or
macrophyte leaf shape and size (Standen 2016). In spring, the association between
catchment scale agriculture and bioindicators did not follow a priori hypotheses,
suggesting that agricultural impacts on stream ecosystems were also limited during that
period. Moreover, the relationship between trait beta diversity and catchment scale
agriculture was likely confounded by natural factors (i.e., site hydrology), and diffuse
sources of agricultural nitrogen were not associated with δ15N of BMI. Although
agricultural stressors are transported to streams and affect water quality during the high
flow period in early spring (Corriveau et al. 2011, Yates et al. 2014a), my results
suggest that these stressors are transported through the system without being assimilated
Page 117
106
in the food web, or other factors are masking their effects (i.e., influence from site
hydrology on traits or blurring of δ15N signatures of fertilizer).
In addition, analysis of biological traits and δ15N of BMI demonstrated the
importance of local habitat and point sources of stressors in defining aquatic insect
communities. More specifically, trait beta diversity was driven by reach scale factors in
both spring and summer (i.e., wetted width and canopy cover, respectively). Therefore,
this bioindicator was effective at detecting local habitat changes in the RRV. In addition,
wastewater discharge had a clear relationship with δ15N of BMI, likely because it is a
direct source of biological nitrogen. However, wastewater discharge was not associated
with trait beta diversity which suggests that this nutrient source did not alter local habitat
considerably although it was assimilated in the food web. This provides evidence that
small streams in the RRV have exceeded nutrient saturation levels as proposed by
Chesworth et al. (in review), leaving canopy cover as the main constraint on stream
productivity in the RRV.
4.5 Conclusions and recommendations
For the small streams traversing the RRV of Manitoba, biological traits and δ15N
of BMI were found to be effective diagnostic bioindicators of small scale impacts and
point source stressors. Catchment scale agriculture did not strongly affect biological
indicators because the study area has limited hydrological connectivity during the
Page 118
107
summer period of high biotic activity. In addition, only human activities in the stream
corridors or stressors that are unrelated to catchment hydrology appear to have an
impact on streams during summer low flow (Yates et al. 2014a). Therefore, both
bioindicators tested are better suited to isolate possible impacts of human activities at
smaller scales, such as agricultural land use in the stream corridor, or in upstream-
downstream studies examining the contribution of pollutants from a specific field or
discharge. Chapter 3 demonstrated that δ15N of primary producers and consumers are
effective bioindicators of nitrogen contribution from wastewater discharge and, to some
extent, manure runoff from beef cattle feedlots. Therefore, δ15N could be used to
understand the influence of point sources of nitrogen throughout specific catchments
when coupled with a gradient study design. This approach could help elucidate whether
these sources are important to Lake Winnipeg, or if their contribution is limited to
upstream ecosystems. Chapter 2 also demonstrated that trait and taxa beta diversity can
be used to detect changes in natural local scale factors, especially riparian conditions. It
may be possible to detect subtle impacts from agriculture on aquatic insect assemblages
in summer if agricultural activities in the stream corridor affect riparian conditions and
site water quality. However, if the goal is to use BMIs to monitoring pollutants reaching
Lake Winnipeg, then these small scale studies may be of limited use to current
Page 119
108
management efforts because the transport of agricultural pollutant is significantly more
important during when biotic activity is minimal (i.e., snowmelt).
My findings reiterate the importance for testing bioindicators over the entire
extent of human exposure and natural gradients in an area of interest before
implementing them in monitoring programs. For example, using broad categories of
human activity would have made a priori mechanistic hypotheses hard to develop in
Chapter 2, and would have hidden effects from cattle production in Chapter 3. In
addition, natural gradients unrelated to human activities had a great influence on
bioindicators as seen in Chapter 2, and revealed a possible lack of connectivity between
catchment scale activities and stream ecosystems. Natural gradients at different scales
may be more important in freshwater monitoring than previously thought (Bailey et al.
2007, Yates and Bailey 2006, Yates and Bailey 2010, Heino 2013), and thus they should
be considered when testing bioindicators (Downes et al. 2010). Bioindicators may still
be able to identify and isolate human impacts if their natural variation can be controlled
for in the study design (Dale and Beyeler 2001). However, in this study, bioindicators
were limited in their ability to detect of agricultural impacts in late spring because of a
possible confounding effect from an unrelated hydrology gradient, and there was simply
no major agricultural impacts to isolate in summer. Therefore, biomonitoring programs
based on aquatic communities may benefit from evaluating the sensitivity of
Page 120
109
bioindicators to human exposure and natural gradients pertinent to the study area to
understand potential mechanistic link between anthropogenic and natural variation, and
potential confounding relationships before interpreting changes in bioindicators as
indication of human impacts (Allan 2004, Yates and Bailey 2006).
In conclusion, future studies or monitoring efforts using BMI as bioindicators in
the study area should be careful in selecting a sampling season appropriate for their
goals and develop study designs controlling for known natural gradients. Moreover,
traditional metrics relying on aquatic insect communities should not be used uncritically
in the summer with the goal of identifying human impacts in the RRV because of the
influence of natural reach and site scale factors found in this study. Even with such
considerations, aquatic insect assemblages may not be an appropriate monitoring tool in
southern Manitoba where the aim is to validate management efforts to reduce pollutant
delivery to Lake Winnipeg, because this monitoring tool is not sensitive during the high
discharge snowmelt period. In contrast, δ15N is a promising technique that is best suited
to determine point sources of nitrogen in the RRV. Future studies should explore a
wider application of δ15N of aquatic insects and POM as bioindicators of wastewater
discharge to answer pertinent management questions in the study area. The relationship
between point sources of manure from beef cattle feedlots should also be explored using
a gradient study design to better understand the contribution of this source of nitrogen to
Page 121
110
streams and potentially to Lake Winnipeg. In addition, this research has implications for
biomonitoring in other geographical regions by demonstrating the importance of
estimating human impacts using fine resolution information about the types and
magnitude of all human activities in an area, and considering natural gradients
separately to test if they are related to human impacts or possible confounding factors.
4.6 References
Allan J.D. (2004) Landscapes and riverscapes: the influence of land use on stream
ecosystems. Annual Review of Ecology, Evolution, and Systematics 35, 257–284.
Bailey R.C., Reynoldson T.B., Yates A.G., Bailey J. & Linke S. (2007) Integrating
stream bioassessment and landscape ecology as a tool for land use planning.
Freshwater Biology 52, 908–917.
Blann K.L., Anderson J.L., Sands G.R. & Vondracek B. (2009) Effects of agricultural
drainage on aquatic ecosystems: a review. Critical Reviews in Environmental
Science and Technology 39, 909–1001.
Chesworth C.T. (2016) Lagoon wastewater effluent impacts stream metabolism in Red
River tributaries. Electronic Thesis and Dissertation Repository Paper 3483.
University of Western Ontario.
Chesworth C.T., Brua R.B., Chambers P.A., Culp J.M. & Yates A.G. (in review) Stream
metabolism is affected by pulse release of municipal wastewater effluent.
Cole M.L., Kroeger K.D., McClelland J.W. & Valiela I. (2005) Macrophytes as
indicators of land-derived wastewater: Application of a δ 15 N method in aquatic
systems. Water Resources Research 41.
Page 122
111
Corriveau J., Chambers P. A., Yates A.G. & Culp J.M. (2011) Snowmelt and its role in
the hydrologic and nutrient budgets of prairie streams. Water Science &
Technology 64, 1590–1596.
Dale V.H. & Beyeler S.C. (2001) Challenges in the development and use of ecological
indicators. Ecological Indicators 1, 3–10.
Downes B.J. (2010) Back to the future: little-used tools and principles of scientific
inference can help disentangle effects of multiple stressors on freshwater
ecosystems. Freshwater Biology 55, 60–79.
Heino J. (2013) The importance of metacommunity ecology for environmental
assessment research in the freshwater realm. Biological Reviews 88, 166–178.
Heino J., Louhi P. & Muotka T. (2004) Identifying the scales of variability in stream
macroinvertebrate abundance, functional composition and assemblage structure.
Freshwater Biology 49, 1230–1239.
Heino J., Mykrä H., Kotanen J. & Muotka T. (2007) Ecological filters and variability in
stream macroinvertebrate communities: do taxonomic and functional structure
follow the same path? Ecography 30, 217–230.
Kaushal S.S., Groffman P.M., Band L.E., Elliott E.M., Shields C.A. & Kendall C.
(2011) Tracking nonpoint source nitrogen pollution in human-impacted
watersheds. Environmental Science & Technology 45, 8225–8232.
Moss B. (2008) Water pollution by agriculture. Philosophical transactions of the Royal
Society of London. Series B, Biological sciences 363, 659–666.
Richards C., Haro R., Johnson L.B. & Host G.E. (1997) Catchment and reach-scale
properties as indicators of macroinvertebrate species traits. Freshwater Biology 37,
219–230.
Sponseller R., Benfield E. & Valett M. (2001) Relationships between land use, spatial
scale and stream macroinvertebrate communities. Freshwater Biology 46, 1409 –
1424.
Page 123
112
Standen K.M. (2013) Sagittaria as a bioindicator of stream nutrient status: effects of
nitrogen and phosphorus on leaf size and shape. University of New Brunswick.
Strayer D.L., Beighley R.E., Thompson L.C., Brooks S., Nilsson C., Pinay G., et al.
(2003) Effects of land cover on stream ecosystems: roles of empirical models and
scaling issues. Ecosystems 6, 407–423.
Tyrrell C.D. (2014) The utility of growth form for predicting and evaluating aquatic
plant nutrient relations. University of New Brunswick.
Ulseth A. & Hershey A. (2005) Natural abundances of stable isotopes trace
anthropogenic N and C in an urban stream. Journal of the North American
Benthological Society 24, 270–289.
Yates A.G. & Bailey R.C. (2010) Improving the description of human activities
potentially affecting rural stream ecosystems. Landscape Ecology 25, 371–382.
Yates A.G. & Bailey R.C. (2006) The stream and its altered valley: integrating
landscape ecology into environmental assessments of agro-ecosystems.
Environmental monitoring and assessment 114, 257–271.
Yates A.G., Brua R.B., Corriveau J., Culp J.M. & Chambers P.A. (2014a) Seasonally
driven variation in spatial relationships between agricultural land use and in-stream
nutrient concentration. River Research and Applications 30, 476–493.
Yates A.G., Brua R.B., Culp J.M. & Chambers P.A. (2012a) Multi-scaled drivers of
rural prairie stream metabolism along human activity gradients. Freshwater
Biology 58, 675–689.
Yates A.G., Brua R.B., Culp J.M., Chambers P.A. & Wassenaar L.I. (2014b) Sensitivity
of structural and functional indicators depends on type and resolution of
anthropogenic activities. Ecological Indicators 45, 274–284.
Yates A.G., Culp J.M. & Chambers P.A. (2012b) Estimating nutrient production from
human activities in subcatchments of the Red River, Manitoba. Journal of Great
Lakes Research 38, 106–114.
Page 124
113
Appendix 1
List of genera present in more than 10% of sites sampled in late spring or summer 2014
in southern Manitoba and their abbreviations used in graphical representations.
Order Family Genus Abbreviation
Diptera Ceratopogonidae Bezzia Cera_Bez
Ceratopogon Cera_Cera
Probezzia Cera_Prob
Chironomidae Ablabesmyia Chi_Abla
Chironomus Chi_Chi
Cladopelma Chi_Clado
Cladotanytarsus Chi_Cladotany
Conchapelopia Chi_Concha
Corynoneura Chi_Coryno
Cricotopus Chi_Crico
Cryptochironomus Chi_Cryptochi
Cryptotendipes Chi_Cryptoten
Dicrotendipes Chi_Dicroten
Endochironomus Chi_Endochi
Eukiefferiella Chi_Euk
Glyptotendipes Chi_Glyptoten
Hydrobaenus Chi_Hydrob
Labrundinia Chi_Labrun
Limnophyes Chi_Limnoph
Micropsectra Chi_Microp
Microtendipes Chi_Microten
Nanocladius Chi_Nanoclad
Parachironomus Chi_Parachi
Parakiefferiella Chi_Paraki
Paralauterborniella Chi_Paralaut
Page 125
114
Order Family Genus Abbreviation
Paratanytarsus Chi_Paratany
Paratendipes Chi_Paraten
Phaenopsectra Chi_Phaeno
Polypedilum Chi_Polyped
Procladius Chi_Proclad
Psectrocladius Chi_Psectroclad
Pseudosmittia Chi_Pseudosm
Rheotanytarsus Chi_Rheotany
Stempellina Chi_Stemp
Stempellinella Chi_Stempell
Tanytarsus Chi_Tany
Thienemanniella Chi_Thien
Zavreliella Chi_Zavre
Simulidae Simulium Sim_Sim
Psychodidae Psychoda Psy_Psy
Coleoptera Elmidae Dubiraphia Col_DubiA
Dubiraphia Larvae Col_DubiL
Haliplidae Haliplus Col_HaliA
Haliplus Larvae Col_HaliL
Peltodytes Col_Pelto
Peltodytes Larvae Col_PeltoL
Hydrochidae Hydrochus Col_Hydro
Dystiscidae Ilybius Col_Ily
Oreodytes Larvae Col_Oreod
Hydrophilidae Berosus Larvae Col_BeroL
Ephemeroptera Baetidae Acerpenna E_Acer
Beatis E_Bae
Procloeon E_Procl
Caeniidae Caenis E_Cae
Page 126
115
Order Family Genus Abbreviation
Ephemeridae Hexagenia E_Hexa
Leptophlebiidae Paralepthophlebia E_Paralepto
Heptageniidae Stenacron E_Stena
Heteroptera Corixidae Sigara H_Sig
Neoplea H_Neo
Megaloptera Sialidae Sialis Mega_Sia
Odonata Cordulidae Somatochlora O_Soma
Coenagrionidae Enallagma O_Enall
Lestidae Lestes O_Lest
Tricoptera Limnephilidae Anabolia T_Anab
Leptoceridae Triaenodes T_Triae
Plecoptera Perlidae Perlesta P_Perl
Page 127
116
Appendix 2
Trait matrix for genera based on the trait database from the National Center for
Environmental Assessment (Vieira et al. 2006), Meyer (2006), Merritt and Cummins
(2008), and Armitage et al. (2012). Trait abbreviations and descriptions provided in
Table 2.2.
Genus Maximum
body size
Respiration Feeding Habitat Voltinism
Ablabesmyia Medium tegument PR SP Multi
Acerpenna Small gills CG SW Multi
Anabolia Large
SH SP
Beatis Small gills CG SW Multi
Berosus Larvae Small gills HB SW Multi
Bezzia Small tegument PR BU Multi
Caenis Small gills CG SP Multi
Ceratopogon Small tegument PR BU Uni
Chironomus Large pigment
tracheal tubes
tegument
CG BU Uni
Cladopelma Small tegument CG BU Multi
Cladotanytarsus Small tegument CG CB Multi
Conchapelopia Small tegument PR SP Multi
Corynoneura Small tegument CG SP Multi
Cricotopus Small pigment
tegument
SH CN Multi
Cryptochironomus Medium pigment
tegument
PR SP Multi
Cryptotendipes Small tegument CG SP
Dicrotendipes Medium pigment
tegument
CG BU Multi
Dubiraphia Adult Small air HB CN Uni
Dubiraphia Larvae Small tegument CG CN Uni
Enallagma Large tegument PR SP Multi
Endochironomus Large tegument SH CN
Eukiefferiella Small tegument CG SP Multi
Page 128
117
Genus Maximum
body size
Respiration Feeding Habitat Voltinism
Glyptotendipes Large pigment
tracheal tubes
tegument
CG BU Uni
Haliplus Adult Small air HB SW Multi
Haliplus Larvae Small tegument HB SW Multi
Hexagenia Large gills CG BU Semi
Hydrobaenus Medium tegument HB SP
Hydrochus Small air HB CB Uni
Ilybius Medium air PR SW Uni
Labrundinia Small tegument PR SP
Lestes Large tegument PR CB Uni
Limnophyes Small tegument CG SP
Micropsectra Small tegument CG CB Uni
Microtendipes Medium tegument CG CN Multi
Nanocladius Small tegument CG SP
Neoplea
Oreodytes Larvae Medium tegument PR SW Semi
Parachironomus Medium tegument CG SP
Parakiefferiella Small
CG SP
Paralauterborniell
a
Small tegument CG BU Multi
Paralepthophlebia Medium gills CG SW Uni
Paratanytarsus Small tegument CG SP
Paratendipes Small tegument CG BU Uni
Peltodytes Larvae Small tegument HB CB
Perlesta Large gills PR CN Uni
Phaenopsectra Small tegument CG CN Multi
Polypedilum Medium tegument CG CB Multi
Probezzia Small tegument PR BU Multi
Procladius Medium tegument PR SP Multi
Procloeon Small gills CG SW Multi
Psectrocladius Medium tegument CG SP Uni
Pseudosmittia Small tegument CG SP
Page 129
118
Genus Maximum
body size
Respiration Feeding Habitat Voltinism
Psychoda
CG BU
Rheotanytarsus Small tegument CF CN Multi
Sialis Medium tegument PR BU Uni
Sigara Small air HB SW
Simulium Small tegument CF CN Multi
Somatochlora Large gills PR SP Semi
Stempellina Small tegument CG CB Multi
Stempellinella Small tegument CG SP Multi
Stenacron Medium gills CG CN Uni
Tanytarsus Medium tegument CG CN Uni
Thienemanniella Medium pigment
tegument
CG SP Multi
Triaenodes Large
SH SW Uni
Zavreliella Small tegument CG CB
Page 130
119
Appendix 3
Descriptive statistics of water quality sampled in late spring and summer 2014 in
streams of southern Manitoba, Canada.
Sampling
time Variable (μg/L) Mean Median SD Min Max n
Spring Total Nitrogen 1493.9 1356.3 841.2 775.0 4557.5 18
Ammonia 26.8 12.5 42.1 0.0 165.5 18
Nitrite and Nitrate 235.1 0.0 772.6 0.0 3294.0 18
Total Dissolved
Phosphorus 219.0 129.0 233.7 17.0 883.0 18
Total Phosphorus 282.0 222 260.4 33.5 909 18
Total Suspended
Solids* 26.8 11.3 46.0 1.7 159.4 18
Summer Total Nitrogen 1375.3 1191.0 641.6 604.7 3093.3 20
Ammonia 14.5 6.5 15.0 0.0 44.0 20
Nitrite and Nitrate 91.8 0.0 307.0 0.0 1343.3 20
Total Dissolved
Phosphorus 342.4 113.1 490.5 14.5 1785.7 20
Total Phosphorus 420.3 185.8 555.8 31.5 2132 20
Total Suspended
Solids* 20.2 15.1 21.0 0.9 85.4 20
*mg/L
Page 131
120
Appendix 4
Comparison of a priori tested models for predicting δ15N changes in primary consumer
taxa, fine and coarse particulate matter (FPOM, CPOM respectively) sampled in late
spring for subcatchments in southern Manitoba, Canada, using corrected Akaike
Information Criterion (AICc) and including redundant models with pretending variables.
Sampling
time Response Model AICc
Delta
AIC
Model
weight
Evidence
Ratio
Spring Hyalella Live2+Wwt1 58.1 0.0 0.999 1 Wwt1 72.6 14.6 0.001 1466 Crop2*Live1 73.5 15.4 0.000 2231 Null 76.4 18.3 0.000 9557 Global model 76.7 18.6 0.000 11159 Live2 78.2 20.1 0.000 23272 Crop1*Live1 84.9 26.8 0.000 663312
Caenis Wwt1 71.0 0.0 0.777 1 Live2+Wwt1 73.5 2.5 0.218 4 Null 81.6 10.7 0.004 206 Live2 84.8 13.8 0.001 1007 Crop1*Live1 87.7 16.8 0.000 4337 Crop2*Live1 88.4 17.4 0.000 6063 Global model 88.5 17.5 0.000 6342
Baetidae Null 76.6 0.0 0.410 1 Wwt1 77.2 0.5 0.318 1 Live2+Wwt1 78.9 2.3 0.130 3 Live2 79.3 2.7 0.108 4 Crop2*Live1 82.9 6.2 0.018 23 Crop1*Live1 83.1 6.5 0.016 25 Global model 89.9 13.2 0.001 739
Summer Hyalella Wwt1 78.8 0.0 0.800 1 Live2+Wwt1 81.6 2.8 0.198 4 Null 91.6 12.8 0.001 602 Live2 94.2 15.4 0.000 2208 Global model 96.4 17.6 0.000 6601 Crop1*Live1 100.1 21.3 0.000 42193 Crop2*Live1 102.6 23.7 0.000 142914
Page 132
121
Sampling
time Response Model AICc
Delta
AIC
Model
weight
Evidence
Ratio
Caenis Wwt1 79.9 0.0 0.603 1 Live2+Wwt1 81.1 1.2 0.328 2 Null 84.8 4.9 0.053 11 Live2 88.4 8.5 0.009 70 Global model 88.8 8.9 0.007 86 Crop1*Live1 96.6 16.7 0.000 4209 Crop2*Live1 99.0 19.1 0.000 13698
Baetidae Wwt1 77.0 0.0 0.808 1 Live2+Wwt1 80.3 3.3 0.157 5 Null 83.9 6.9 0.026 31 Live2 86.3 9.3 0.008 106 Crop2*Live1 91.3 14.2 0.001 1236 Crop1*Live1 92.6 15.6 0.000 2392 Global model 95.8 18.8 0.000 11849
FPOM Wwt1 86.4 0 0.655738 1 Live2+Wwt1 87.7 1.3 0.342325 2 Global model 98.8 12.4 0.001358 483 Null 101.1 14.7 0.000415 1580 Live2 103.7 17.3 0.000116 5653 Crop1*Live1 106.4 20.0 2.96E-05 22137 Crop2*Live1 107.4 21.0 1.82E-05 35954
CPOM Wwt1 92.8 0 0.499332 1 Live2+Wwt1 92.8 0 0.499332 1 Global model 105.4 12.6 0.000926 539 Null 107.6 14.8 0.000305 1636 Live2 110.4 17.6 7.6E-05 6568 Crop1*Live1 112.9 20.1 2.18E-05 22925 Crop2*Live1 115.1 22.3 7.25E-06 68871
Page 133
Curriculum Vitae
Candidate’s full name: Sophie Nicole Cormier
Universities attended:
University of New Brunswick, Bachelor of Science, 2009-2013
University of New Brunswick, Master of Science, 2013-2016
Conference Presentations:
Cormier S.N., Culp J.M., Yates A.G., Brua R.B. 2016. Can delta 15-N of aquatic
invertebrates identify sources of agricultural nitrogen in small Canadian streams?
Institute of Landscape Ecology and Resources Management Colloquium, Justus-Liebig-
Universität, Giessen, Germany. April 2016. Oral Presentation.
Cormier S.N., Culp J.M., Yates A.G., Brua R.B. 2015. Indicators of ecological function
to diagnose agricultural impacts in stream assemblages of southern Manitoba, Canada.
European Symposium on Fresh Water Sciences (SEFS), Geneva, Switzerland. July
2015. Oral Presentation.
Cormier S.N., Culp J.M., Yates A.G., Chambers P.A. 2014. Ecological indicators to
diagnose agricultural impacts in streams of southern Manitoba. Canadian Rivers
Institute AGM, Charlottetown, PE. October 2014. Poster Presentation.