TROPHIC STRUCTURE OF MIDWATER FISHES OVER COLD SEEPS IN THE NORTH CENTRAL GULF OF MEXICO Jennifer P. McClain-Counts A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment of the Requirements for the Degree of Master of Science Center for Marine Science University of North Carolina Wilmington 2010 Approved by Advisory Committee Steve W. Ross Lawrence B. Cahoon Chair Joan W. Willey Accepted by Dean, Graduate School
Transcript
TROPHIC STRUCTURE OF MIDWATER FISHES OVER COLD SEEPS IN THE NORTH CENTRAL GULF OF MEXICO
Jennifer P. McClain-Counts
A Thesis Submitted to the University of North Carolina Wilmington in Partial Fulfillment
of the Requirements for the Degree of Master of Science
Center for Marine Science
University of North Carolina Wilmington
2010
Approved by
Advisory Committee
Steve W. Ross Lawrence B. Cahoon Chair
Joan W. Willey
Accepted by
Dean, Graduate School
TABLE OF CONTENTS ABSTRACT....................................................................................................................... iv
ACKNOWLEDGMENTS ................................................................................................. vi
DEDICATION.................................................................................................................. vii
LIST OF TABLES........................................................................................................... viii
LIST OF FIGURES ........................................................................................................... xi
LITERATURE CITED ......................................................................................................37
iii
ABSTRACT
Midwater fishes are an important component of pelagic food webs and provide insight into
energy utilization and movement through the water column. In this study, the diets of midwater
fishes collected over cold seep habitats were examined to determine general feeding patterns and
whether size, depth, time of day or location affected diet composition within fish species. The
base of the midwater food web was also examined to determine whether chemosynthetic energy
in benthic cold seeps was incorporated into the midwater fish community. Discrete depth Tucker
trawling was conducted in August 2007 over three cold seep habitats (> 1000 m) in the north-
central Gulf of Mexico. Surface sampling was also conducted to provide a prey base
(zooplankton and POM) for stable isotope analyses (SIA). Gut content analysis (GCA) and SIA
(δ13C and δ15N) in conjunction with IsoSource software were utilized for diet reconstruction and
to determine trophic positions. SIA also aided efforts to determine chemosynthetic influences on
the midwater food web. GCA was performed on 31 species in the five most abundant families
(Gonostomatidae, Myctophidae, Phosichthyidae, Sternoptychidae and Stomiidae), with midwater
fishes classified into one of three guilds: piscivore, large crustacean consumer, or
zooplanktivore. SIA was performed on 6 fish families (Gonostomatidae, Myctophidae,
Phosichthyidae, Sternoptychidae, Stomiidae, and Melamphaidae), 13 invertebrate categories, and
3 primary producers (POM, Sargassum spp. and detritus), and classified all fishes as
zooplanktivores. Using IsoSource, more precise contributions of individual prey taxon were
documented, which did not always support results from GCA. Size, depth, time of day and
location did not affect diet composition within a species; however migration trends suggested
competition may be reduced by feeding over a range of depths and over a 24 hour period.
Significant differences in trophic position calculations between GCA and SIA highlighted the
iv
importance of using multiple techniques to describe trophic structure, as each method
characterized the diets differently.
v
ACKNOWLEDGMENTS
This project was largely funded by the Department of the Interior U.S. Geological Survey
under Cooperative Agreement No. 05HQAG0009, sub agreement 05099HS004. I thank the crew
of the R/V Cape Hatteras and all scientific personal for assisting with fishing operations and
sample processing. S. Artabane, A. Quattrini, and A. Roa-Varon assisted with fish
identifications and C. Ames assisted with invertebrate identifications. Guidance and support
during stable isotope analyses were provided by Drs. A. Demopoulos and C. Tobias, and K.
Duernberger. I would also like to thank S. Artabane, T. Casazza and A. Roa-Varón for their
assistance in dissecting and processing fish stomachs. Special thanks to my committee, Drs. S.
Ross, L. Cahoon, and J. Willey, for their guidance and support during the duration of this project.
I would additionally like to thank my advisor, Dr. S. Ross, for setting me up with this project and
Dr. L. Cahoon for his assistance with statistics. Finally, thanks to S. Ross, T. Casazza, A.
Demopoulos, A. Quattrini, L. Truxal and M. Carlson for their suggestions and edits provided
throughout the writing process of this thesis.
vi
DEDICATION
I would like to dedicate this thesis to my parents, who encouraged my early passion in
marine science and gave me the confidence to follow my dreams and overcome any obstacles.
Your constant love and support was unwavering and because of that, I can present this Masters
project.
vii
LIST OF TABLES
Table Page 1. Surface and midwater stations sampled over three cold seep sites
(AT340, GC852, and AC601) (see Fig.1) in the Gulf of Mexico (9-25 August 2007)................................................................................................48
2. The total number of all midwater fishes, invertebrates and
autotrophs examined in dietary analyses from the North-central Gulf of Mexico ......................................................................................................55
3. Results of ANOSIM comparing effects of size, time of day, depth
and location on the general prey categories consumed for each fish species.............................................................................................................58
4. Percent volume and frequency of prey items consumed by
Chauliodus sloani collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................59
5. Percent volume and frequency of prey items consumed by
Gonostoma elongatum collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................60
6. Percent volume and frequency of prey items consumed by
Stomiidae collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................62
7. Percent volume and frequency of prey items consumed by
Cyclothone alba collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................63
8. Percent volume and frequency of prey items consumed by
Cyclothone braueri collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................65
9. Percent volume and frequency of prey items consumed by
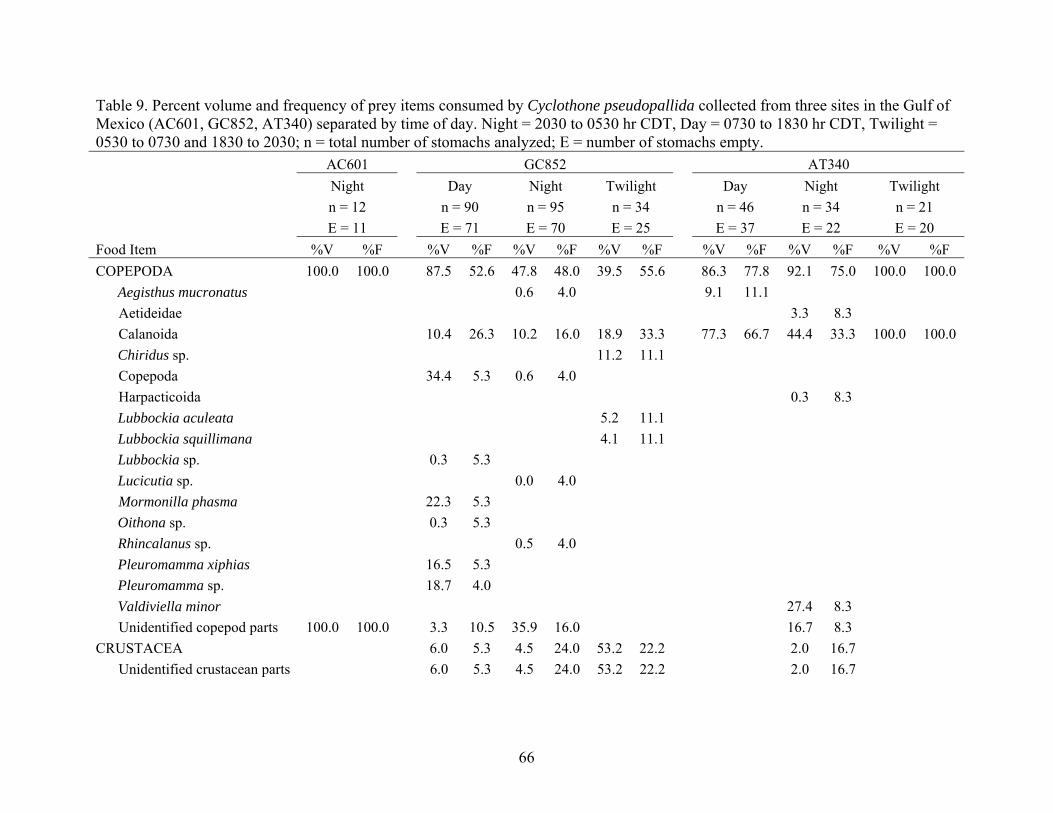
Cyclothone pseudopallida collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................66
10. Percent volume and frequency of prey items consumed by
Hygophum benoiti collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ............................68
viii
11. Percent volume and frequency of prey items consumed by Valenciennellus tripunctulatus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day....................69
12. Percent volume and frequency of prey items consumed by
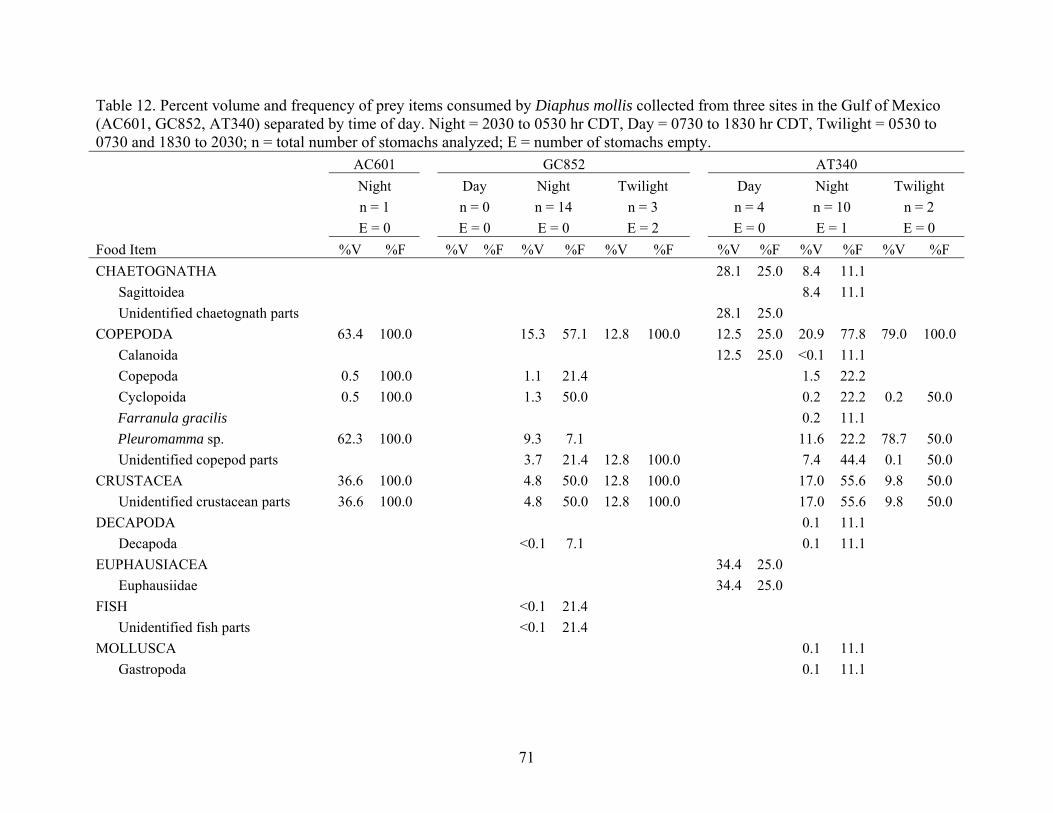
Diaphus mollis collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................71
13. Percent volume and frequency of prey items consumed by
Cyclothone pallida collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................73
14. Percent volume and frequency of prey items consumed by
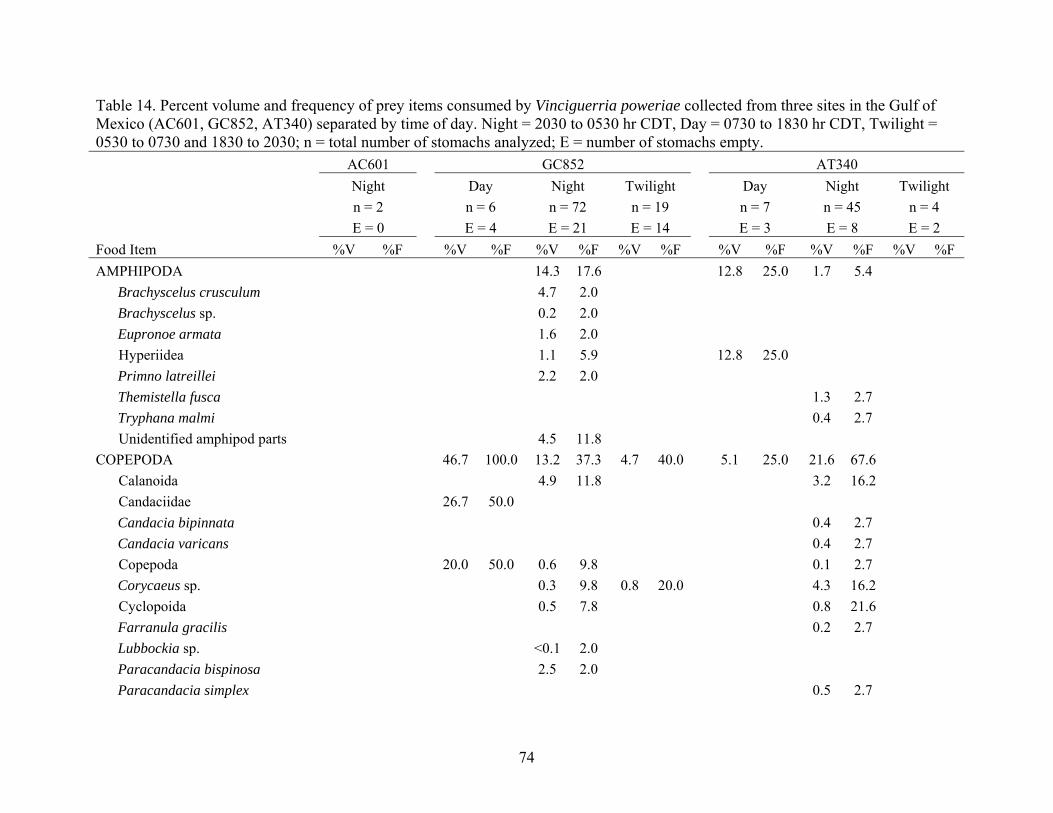
Vinciguerria poweria collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................74
15. Percent volume and frequency of prey items consumed by
Myctophum affine collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................77
16. Percent volume and frequency of prey items consumed by
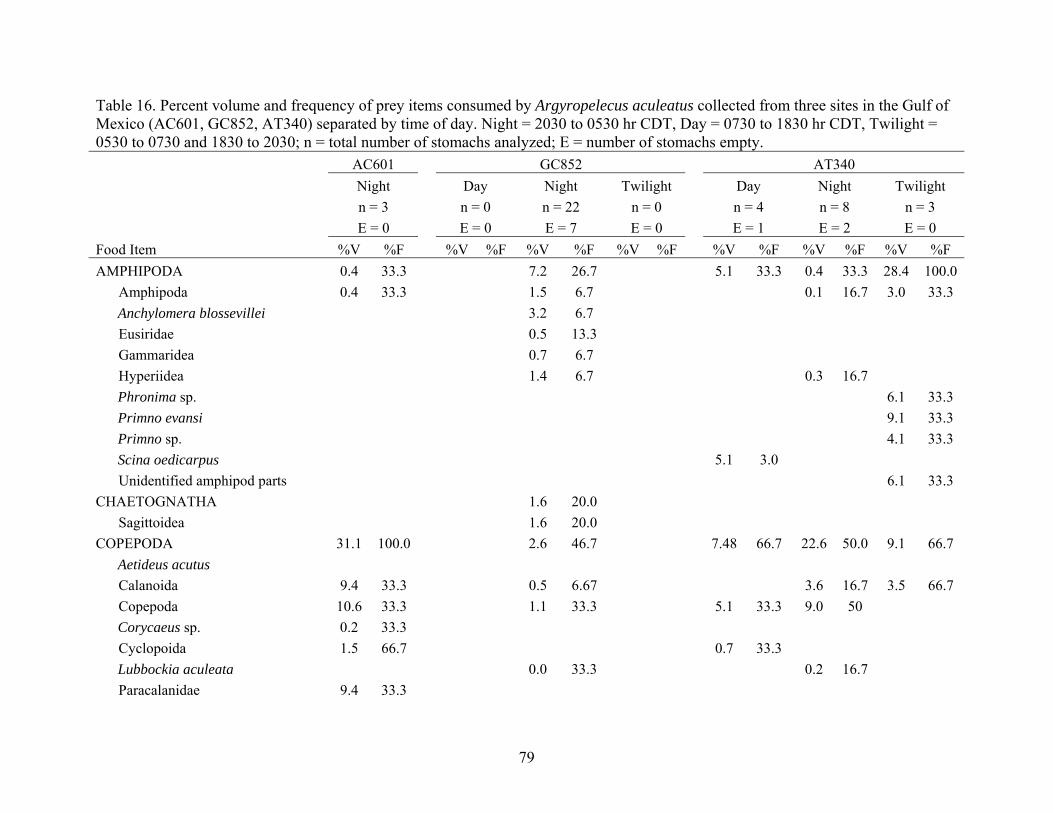
Argyropelecus aculeatus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................79
17. Percent volume and frequency of prey items consumed by Argyropelecus hemigymnus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ............................81
18. Percent volume and frequency of prey items consumed by Pollichthys mauli collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................82
19. Percent volume and frequency of prey items consumed by
Benthosema suborbitale collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................84
20. Percent volume and frequency of prey items consumed by
Lampanyctus alatus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................86
21. Percent volume and frequency of prey items consumed by
Lepidophanes guentheri collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day..............................................88
ix
22. Percent volume and frequency of prey items consumed by Notolychnus valdiviae collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ................................90
23. Percent volume and frequency of prey items consumed by Ceratoscopelus warmingii collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day ............................92
24. Mean (± 1 SE) δ13C and δ15N values for midwater fishes, invertebrates
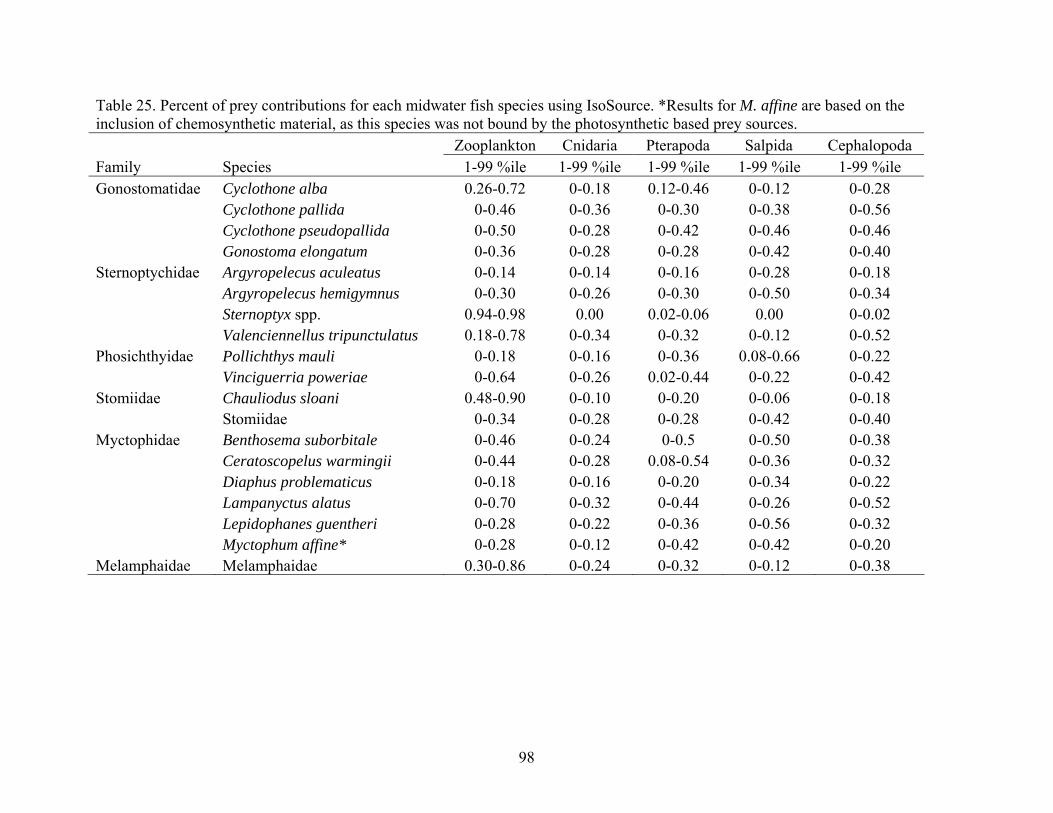
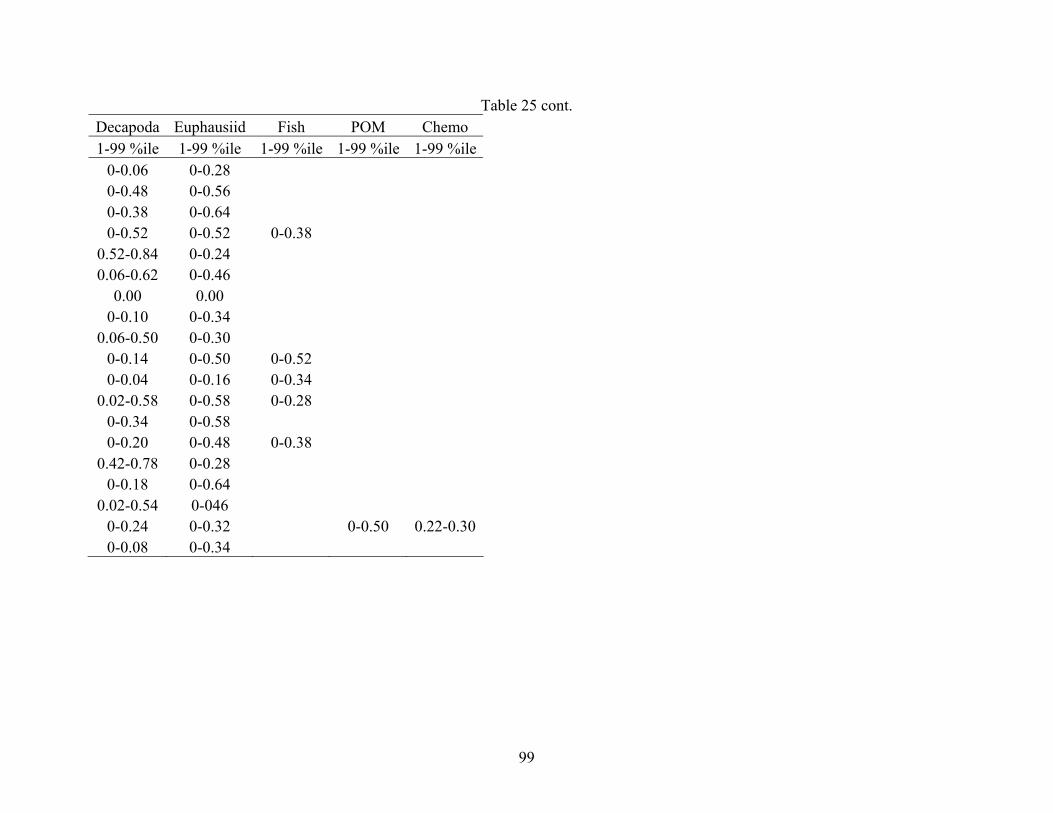
and carbon sources collected from each site (AC601, AT340, GC852) ...............95 25. Percent of prey contributions for each midwater fish species using
IsoSource ...............................................................................................................98 26. Mean trophic position (TP), one standard deviation (SD), range
(minimum – maximum) and number of fish (n) for each midwater fish species collected in the North-central Gulf of Mexico, using data from stable isotope and gut content analyses ......................................................100
x
LIST OF FIGURES
Figure Page 1. Sampling areas in the North-central Gulf of Mexico for midwater
fauna, 9-25 August 2007. The three cold seep sites (AT340, GC852, AC601) were located on the continental slope at depths > 1000 m. Each dot represents one station ...........................................................................101
2. Multidimensional scaling (MDS) plot documenting the differences
among the gut contents of midwater fishes. Data were based on the Bray-Curtis similarity matrix calculated from standardized, square root transformed, mean volumes of prey (12 general categories) .......................102
3. Relationships among stomach fullness, mean depth of capture and
time for midwater fishes. Data were compiled from all sites and excluded specimens lacking depth data...............................................................103
4. Plot of the average δ15N values against the average δ13C values
(± 1 standard error) for midwater fishes, invertebrates and primary producers collected in the Gulf of Mexico ..........................................................111
5. Relationship between δ15N and SL for all midwater fish species........................112
xi
INTRODUCTION
Midwater fishes constitute an important component of the pelagic food web due to their high
abundances, migratory behavior, and global distribution (Gjøsaeter and Kawaguchi 1980;
Cornejo and Koppelmann 2006). Many of these unique fishes inhabit the mesopelagic zone (200
to 1000 m), and although they are consumed by a variety of marine fauna, such as benthic
grenadiers (Laptikhovsky 2005), pelagic tuna (Potier et. al. 2007), and penguins (Adams et al.
2004), midwater fishes are also fierce predators. In the eastern Gulf of Mexico, midwater fishes
consume 5-10% of the daily zooplankton production in the epipelagic zone (< 200 m) (Hopkins
et al. 1996). The ability of midwater fishes to impact both surface and bottom communities
results from diel vertical migrations (DVMs), a unique behavior exhibited by many midwater
fish species. Species that undergo DVMs migrate from the mesopelagic zone to the epipelagic
zone at night, primarily at sunset, and return to the mesopelagic zone at sunrise. Through DVMs,
midwater fishes, particularly myctophids (Kinzer 1977; Hidaka et al. 2001; Cornejo and
Koppelmann 2006), contribute significantly to the vertical transport of organic matter from the
epipelagic zone to the mesopelagic zone (Ashjian et al. 2002; Brodeur and Yamamura 2005),
thus impacting the trophic structure of the water column. By tracking these trophic relationships,
a more thorough understanding of pelagic energy and material flow through the water column
can be established.
Previous dietary studies on midwater fishes (Hopkins and Baird 1985a,b; Lancraft et al.
1988; Hopkins et al. 1996; Butler et al. 2001; Pusch et al. 2004) utilized gut content analyses
(GCA) to determine trophic relationships. Generally, midwater fishes were divided into three
major feeding guilds: zooplanktivores, which consume planktonic organisms such as amphipods,
copepods and euphausiids; micronektonivores, which consume fishes and cephalopods; and
1
generalists, which consume a variety of unrelated taxa (Gartner et al. 1997). Unfortunately, as
GCA only represent short-term diet (< 24 hours) (Hadwen et al 2007), placement into these
feeding guilds could vary and may be inaccurate. Guild placement can be affected by dietary
shifts resulting from changes in prey abundance (Kawaguchi and Mauchline 1982), seasonality
(Kawaguchi and Mauchline 1982) and ontogeny (Kawaguchi and Mauchline 1982; Young and
Blaber 1986; Hopkins et al. 1996; Beamish et al. 1999; Williams et al. 2001; Butler et al. 2001).
Additionally, accurate guild placement is not possible for specimens with empty stomachs,
common in midwater fishes (Gartner et al. 1997). The trophic relationships relating midwater
fishes to their carbon sources are also limited with GCA, which may not always allow the
determination of which autotrophs contributed to a food web (Thomas and Cahoon 1993).
Therefore, despite providing detailed dietary data, GCA only documents a portion of the trophic
structure.
Issues related to GCA (noted above) may be addressed using stable isotope analyses (SIA).
Although SIA cannot provide detailed prey data (e.g., species level prey identifications), SIA
provides general information on the cumulative feeding habits of an organism (Fry 2006).
Trophic positions within a food web (Fry 1988; Van Dover 2000; Hobson 2002; Behringer and
Butler 2006; Fry 2006; Paradis et al. 2008) can be estimated from SIA due to the isotopic ratio of
nitrogen (15N/14N or δ15N), increasing an average of 3.4‰ per trophic level (Minagawa and
Wada 1984; Post 2002). In contrast to nitrogen, the isotopic ratio of carbon (13C/12C or δ13C), has
little fractionation between trophic levels, with an increase ≤ 1‰ per trophic level (Post 2002;
Lajtha and Michener 1994; Minagawa and Wada 1984). Despite this low fractionation, carbon is
useful for determining carbon sources as distinct ranges are documented for different autotrophs:
-22 to -16‰ for marine phytoplankton (Post 2002; Fry 2006), -18 to -15‰ for Sargassum spp.
2
(Rooker et al. 2006), -16 to -5‰ for turtlegrass (Hemminga and Mateo 1994), and -75 to -28‰
for chemosynthetic material (Kennicutt et al. 1992). Unfortunately, despite the added benefit of
combining SIA and GCA in dietary analyses, few studies (e.g. Vander Zanden et al. 1997;
Hadwen et al. 2007; Drazen et al. 2008; Rybczynski et al. 2008) have done so.
In the Gulf of Mexico (GOM), trophic structure may be affected by the complex bottom
topography and hydrography. The dominant current, the Loop Current, flows from the Caribbean
Sea through the Yucatan Channel, around the east-central portion of the GOM and flows out near
southern Florida (Hyun and Hogan 2008; Sturges et al. 2005). The oscillation of this current
often results in warm- and cold-core rings breaking off, which affect circulation (Schmitz et al.
2005), primary productivity and food web dynamics (Waite et al. 2007). Additionally, the
Mississippi River flows into the northern portion of the basin providing large amounts of
freshwater, sediments and nutrients, affecting both the water physics and chemistry and faunal
communities (Baguley et al. 2006; Jarosz and Murray 2005). The presence of benthic features,
like cold seeps or corals, can also affect trophic complexity, particularly as chemosynthetic
communities can be associated with cold seeps. Higher abundances of non-seep, benthic fauna
were occasionally observed in the vicinity of seeps (Levin 2005) and may consume
chemosynthetic material (MacAvoy et al. 2002; 2008). However, whether midwater fishes are
impacted by chemosynthetic energy pathways either in the water column above the seep areas or
by interactions with the associated benthic communities has not been examined.
Food web studies provide an effective means of tracking energy flow through an ecosystem.
The purpose of this study was to examine the trophic structure of midwater fishes over cold seep
areas (> 1000 m) in the north-central GOM. The presence of changing hydrography and prey
resources at the sites may affect trophic structure. This study used both GCA and SIA to
3
thoroughly document the trophic relationships of midwater fishes. The objectives were to: 1)
determine basic feeding patterns of the dominant midwater fish species collected, 2) document
feeding changes, if any, that occurred among species due to differences in size, time of day,
depth or location, 3) examine the relationship, if any, between feeding and DVM patterns in the
midwater fishes, 4) document the differences in short term (GCA) and long term (SIA) feeding,
and 5) examine the base of the midwater food web to determine whether the midwater
community utilized chemosynthetic energy sources from cold seeps.
METHODS
Study Area
Three cold seep sites in the GOM were selected for sampling based on data collected by TDI-
Brooks International, Inc: Atwater Valley Block 340 (AT340), Green Canyon Block 852
(GC852), and Alaminos Canyon Block 601 (AC601). These three sites are located on the middle
to lower continental slope in the north-central GOM, and each contained benthic chemosynthetic
communities (Fig. 1). Detailed bottom topography was documented for each site from previous
seismic profiles and surveys from a submersible and a remotely operated vehicle (Roberts et al.
2007). AT340 (2216 m) contained multiple mounds located on a topographic high. Submersible
surveys of the area documented extensive carbonate substrata, large mussel beds, clumps of
tubeworms and a few soft corals. GC852 (1450 m) was characterized by an elongated ridge
approximately 2 km long running north to south, with vast amounts of carbonate substrata and
numerous corals on the crest. Tubeworms and mussel beds were also documented at this site.
Additionally, oil slicks were present on the surface and bubble streams were reported on the
bottom, which may be potential mechanisms for transporting benthic material to the surface.
AC601 (2340 m) differed from the other two sites, having low topography and a large brine pool.
4
Some carbonate substrata and a few isolated aggregations of tubeworms were present, none of
which were near the brine pool. High methane concentrations in the water column were also
recorded in the water column over this site (Roberts et al. 2007).
Sample Collection
Intense sampling of the upper 1000 m of the water column was conducted during 24 hour
operations at all three sites from 9-25 August 2007; however, due to inclement weather, only
minimal night sampling was conducted at AC601. A total of 173 stations (45 day, 108 night, and
20 twilight) were sampled (Table 1). Multiple gear types were utilized to adequately sample the
fauna, including a Tucker trawl, Neuston net, and plankton nets, though discrete-depth Tucker
trawling was emphasized. Midwater fauna were collected using a Tucker trawl (2 x 2 m, 1.59
mm mesh, 505 μm cod end.) with a plankton net (0.5 m diameter, 335 μm mesh) attached inside
the Tucker trawl mouth to simultaneously sample the smaller components of the midwater fauna.
Trawls were equipped with a Sea-Bird SBE39 temperature-depth recorder (TDR) attached to the
upper frame bar to record time, depth, and temperature during deployment. The Tucker trawl
was deployed open, and it was assumed no significant fishing occurred during deployment due to
the rapid lowering, steep wire angle, and minimal forward movement of the vessel (Gartner et
al., 2008; Ross et al. 2010). Upon reaching the designated depth, the trawl fished for
approximately 30 min at a 2 knot (3.7 km/hr) ground speed and was triggered closed using a
double trip mechanism. Actual time and depth fished for each trawl was determined post-tow
using data from the TDR. TDR data were used throughout the cruise to adjust fishing strategies
to achieve desired sampling depths. The mean depth for each Tucker trawl tow was calculated by
averaging all depths recorded by the TDR from the start to the end of each tow. Tucker trawling
5
intensely sampled the upper 1000 m of the water column over the 24 hour time period at GC852
and AT340.
Zooplankton samples were collected from a 1.1 x 2.4-m Neuston net (6.4-mm mesh body and
3.2-mm tail bag) or plankton nets (0.5 m diameter, 335 μm and 1.0 m diameter, 505 μm mesh)
deployed at the surface and towed for 15-30 minutes (Table 8.1). Particulate organic material
(POM) was collected by filtering seawater through a 125-μm precombusted glass filter, and it
was assumed that the majority of POM was phytoplankton derived (Kling et al., 1992). POM and
zooplankton samples provided a food web base for SIA.
Fishes collected were preserved in 10% seawater-formalin solution and later transferred to
50% isopropyl for storage until dietary analyses. Invertebrates were preserved in 70% ethanol,
with the exception of jelly and salp specimens that were preserved in 10% seawater-formalin
solution. All specimens were sorted, identified to the lowest possible taxa and measured to the
nearest millimeter standard length (SL) (fishes) or total length (TL) (invertebrates). The life
history stage of fishes was also documented based on the presence or absence of gonads. Fish
specimens were classified as juvenile when either no gonads or immature gonads were present.
Dietary Analyses
Gut Content Analysis (GCA)
GCA was conducted for the five most abundant midwater families (31 species) using
methods outlined in Ross and Moser (1995). All abundant species collected (> 30 individuals,
with the exception of stomiids) were analyzed. In order to increase sample size for Stomiidae, all
stomiids, with the exception of C. sloani, were grouped together and were collectively referred to
as Stomiidae. Highly abundant fish species, Cyclothone alba (n = 614), C. braueri (n = 669), C.
pallida (n = 885), C. pseudopallida (n = 744), Valenciennellus tripunctulatus (n = 248), and
6
Notolychnus valdiviae (n = 1139), were randomly subsampled, with selected specimens spanning
the collected size range of the species, encompassing all depths sampled, and including day,
night and twilight samples. Selected fishes were dissected and the stomachs were removed.
Stomach fullness was estimated as 0%, 5%, 25%, 50%, 75% or 100%. Empty stomachs were
documented, though not included in most analyses, for day, night and twilight samples at all
sites. Stomach contents were placed on a Petri dish and identified to the lowest possible taxon.
Similar prey items were then piled together on a grid of 1 mm squares and flattened to a uniform
height, which was measured. This height multiplied by the number of squares occupied by the
food item yielded volume in mm3. The frequency of occurrence for a prey item equaled the
number of times a prey item occurred in the fish species examined divided by the total number of
stomachs analyzed.
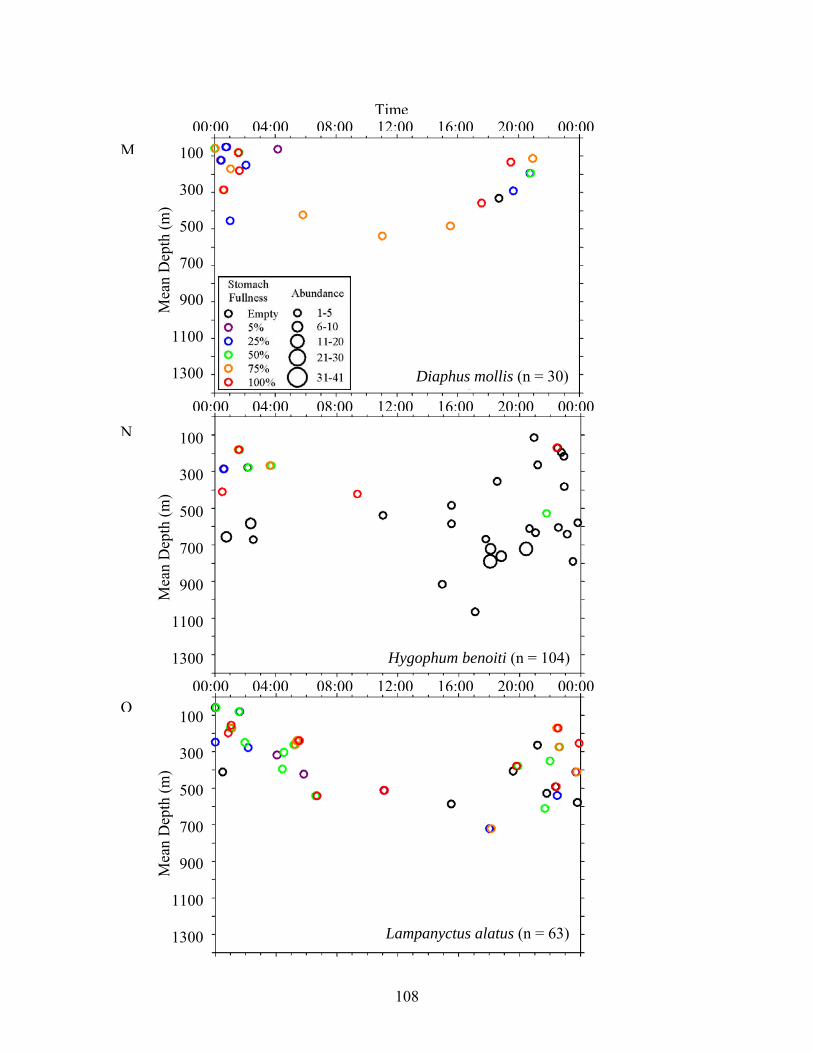
The relationship between DVMs and stomach fullness was examined by plotting stomach
fullness against time of day and mean sampling depth. Time of day was divided into three
categories: day (0730 to 1830 hr CDT), night (2030 to 0530 hr CDT), and twilight (0530 to 0730
hr CDT, one hour on either side of average sunrise, and 1830 to 2030 hr CDT, one hour on either
side of average sunset) and mean sampling depths were calculated based on Ross et al. (2010).
TT tows where no mean sampling depth was calculated were excluded.
Stable Isotope Analysis (SIA)
Prior to specimen preservation in formalin or ethanol, samples of white muscle tissue were
dissected from fishes and invertebrates and frozen. For consistency, tissue was removed from
similar body regions based on the type of specimen (i.e., muscle tissue removed from the dorsal
region of fishes, the caudal region of shrimps, the legs of crabs and the mantle of mollusks).
When specimens were too small to extract a tissue sample, the whole body was used. Minimal
contamination from other tissue types occurred as the head, scales, photophores, and entrails
7
were removed from specimens taken whole. For these specimens, species identification was
made either prior to tissue collection, or a replicate specimen was vouchered for future
identification. All collected isotope samples were dried and crushed into a powder. The majority
of samples were dried to a constant weight in an oven at 50-60˚C. Additional samples were
frozen at -80˚C for ≥ 24 hours and freeze dried in a VirTis Benchtop 3.3 Vac-Freeze. I assumed
there were no significant differences in isotopic ratios as a result of different drying techniques
(Bosley and Wainright 1999).
Tissue samples were analyzed for carbon and nitrogen isotope ratios. For each sample, 400-
600 μg were placed into a tin capsule and combusted in an Elemental Combustion System Model
4010 coupled to a Delta V Plus Isotope Ratio Mass Spectrometer (IRMS) via Conflo II interface
at the University of North Carolina Wilmington (UNCW). POM (provided by A. Demopoulos,
USGS), 49 fishes and 24 invertebrates were analyzed by IRMS at Washington State University
using a Costech (Valencia, USA) elemental analyzer interfaced to a GV instruments
(Manchester, UK) Isoprime IRMS. Precision of the IRMS at UNCW was verified by repeated
analysis of standards USGS 40 and USGS 41, which were incorporated into each sample run.
Raw delta values were corrected for linearity and normalized to known reference materials
USGS 40 and USGS 41. A similar procedure was utilized at Washington State University using
egg albumin powder calibrated against National Institute of Standards reference materials.
Reproducibility was monitored using several organic reference standards (Fry 2007). Isotope
ratios were expressed in the standard delta (δ) notation as parts per thousand (‰) according to
the following equation:
1000*R
)R - (R X
standard
standardsample=δ (1)
8
where X is 13C or 15N and R is the corresponding ratio 13C/12C or 15N/14N. The global standards
for δ13C and δ15N are Vienna PeeDee Belemnite and atmospheric nitrogen (air). A minimum of 5
samples were analyzed per fish species. Similar to GCA, the sample size of Stomiidae was
increased by combining all species, with the exception of C. sloani. Similarly, 3 melamphid
species (Melamphaes simus, M. typhlops, and Scopelogadus mizolepis) were grouped together to
increase sample size for analyses and were referred to as Melamphaidae. Diaphus spp. included
D. mollis and D. lucidus and Sternoptyx spp. included S. diaphana and S. pseudobscura.
Data were examined after SIA to determine whether inorganic carbon or lipids may have
significantly impacted the isotope results. According to Post et al. (2007) samples with C:N > 4
are likely affected by the presence of lipids, and inorganic carbon may be present when C:N >
3.5 or δ13C is highly enriched. Our results (all C:N < 4) indicated that neither lipids nor inorganic
carbon significantly impacted the isotope ratios of fishes; therefore, no lipid extraction or
acidification methods were utilized for fish isotope samples. In contrast, some invertebrates had
high C:N values that suggested the presence of inorganic carbon in the samples. As a result, an
acidification process was conducted on a subset of invertebrate samples, which included
amphipods, copepods, euphausiids, jellyfish, pterapods, salps, and zooplankton, to remove any
inorganic carbon. To acidify samples, 1.0 N hydrochloric acid was added one drop at a time to
dried, crushed tissue samples until bubbling no longer occurred. Acidified samples were air dried
for 8 hours before being re-dried in an oven at 50-60˚C for 24 hours. These samples were then
processed by the same method utilized for nonacidifed samples (see above). As acidification can
affect N values, acidified samples were reported with δ15N reflecting the ratio from the untreated
sample and δ13C reflecting the acidified sample (Jacob et al. 2005; Pinnegar and Polunin 1999).
9
Isotope Mixing Models
Isotope data were analyzed using IsoSource 3.5. IsoSource is a multisource mixing model
program that calculates all possible solutions for the contribution of each prey source to a
consumer’s diet based on the isotopic signatures of the prey and predator (Phillips and Gregg
2003; Benstead et al. 2006). For this study, the average carbon and nitrogen values for each prey
item and fish were entered into the mixing model to determine all feasible contributions. Prior to
analysis, nitrogen values for consumers were corrected for trophic fractionation, set at 2‰, based
on the trophic shift documented in my isotope data. It was assumed no trophic fractionation
occurred in carbon (Demopoulos et al. 2007; France and Peters 1997). Tolerance was set at 0.2%
with source increments set at 0.2%. Reported ranges represented the 1-99th percentile because
the resulting ranges (minimum to maximum) are sensitive to small numbers of observations at
the ends of the distribution and the 1-99th percentile range may be more robust to outliers
(Philips and Gregg 2003).
Trophic position analysis
Data collected during GCA and SIA were used to calculate the trophic position of each
individual fish based on the following two equations from Vander Zanden et al. (1997)
[ ]∑ += 1))(( iiGC TPVTP (2)
where TPGC is the trophic position of the fish based on gut content analysis, Vi is the percent
volume of a nth prey item and TPi is the trophic position of nth prey item based on data from
Rybczynski et al. (2008) and
2)(
)( 11515
+−
= °
fNN
TP consumerfishSIA
δδ (3)
10
where TPSIA is the trophic position of the fish based on stable isotope analysis and f is the trophic
fraction for one trophic level.
Statistical Analyses
Multivariate analyses were conducted on gut contents of each fish species to examine diet
differences based on four factors: size, time of day, depth and location. All analyses utilized the
software PRIMER-E version 6.1 (Clarke and Warwick 2001; Clarke and Gorley 2006). Factors
were divided into groups as follows: SL was divided into size classes based on 5 mm increments
(10-14 mm, 15-19 mm, 20-24 mm, 25-29 mm, 30-34 mm, 35-39 mm, 40-44 mm, 45-49 mm, 50-
54 mm, 55-59 mm, 60-64 mm, 65-69 mm, 70-74 mm, 75-79 mm, 80-84 mm, 85-89 mm, 90-94
mm, 95-99 mm, 100-104 mm, 105-109 mm, 110-114 mm, ≥115 mm); time of day was divided
into three categories, day (0730 to 1830 hr CDT), night (2030 to 0530 hr CDT), and twilight
(0530 to 0730 and 1830 to 2030 hr CTD); depth (based on mean sample depth) was divided into
ranges based on 50 m increments (surface-49 m, 50-99 m, 100-149 m, 150-199 m, 200-249 m,
250-299 m, 300-349 m, 350-399 m, 400-449 m, 450-499 m, 500-549 m, 550-599 m, 600-649 m,
650-699 m, 700-749 m, 850-899 m, 900-949 m, 950-999 m, 1000-1049 m, 1050-1099 m, 1100-
1149 m, 1150-1199 m); and location was divided into the three sites (AT340, GC852, AC601).
Organic material and animal parts (e.g., amphipod parts, copepod parts, decapod parts) were
excluded prior to analyses as these food items were ambiguous and may be pieces of prey items
identified to lower taxa. For each fish species, the prey item volumetric data were standardized
for each individual fish by dividing the volume of each prey item by the total volume of the
stomach in order to account for stomach fullness variability. Standardized volumes were then
square root transformed to down weight the contributions of abundant prey items. Next,
similarities among fish species were calculated using a Bray-Curtis similarity coefficient based
11
on each factor. The resulting similarity matrix was then subjected to a one way analysis of
similarities (ANOSIM) to determine if diets were significantly different for each factor, with
R>0.40 and p<0.05 used as the criteria for statistical significance. When significant differences
were found using ANOSIM, a similarity percentages routine (SIMPER) was utilized to
determine which prey items contributed to the dissimilarities. This process was repeated for each
fish species.
A similar multivariate procedure was implemented for diet comparisons among all 31 fish
species, disregarding the factors size, time of day, depth and location. After constructing a Bray-
Curtis similarity matrix, results were subjected to hierarchical clustering with group average
linkage and non-metric multidimensional scaling (MDS). With a large sample size (n = 1327), a
MDS plot can become cluttered with substantial “noise” in individual samples; therefore, data
were averaged by PRIMER based on species prior to standardization (see above for
standardization process) (Clarke and Gorley 2006). Additionally, to determine general feeding
guilds, all fish species were analyzed using general prey categories (Amphipoda, Annelida,
P.A. and S. Pesant. 2007. Food web structure in two counter-rotating eddies based on δ15N
and δ13C isotopic analyses. Deep-Sea Research II 54: 1055-1075
Williams, A., Koslow, J.A., Terauds, A. and K. Haskard. 2001. Feeding ecology of five fishes
from the mid-slope micronekton community off southern Tasmania, Australia. Marine
Biology 139: 1177-1192.
Woodward, G. and A.G. Hildrew. 2002. Food web structure in riverine landscapes. Freshwater
Biology 47: 777-798.
Young, J.W. and S.J.M. Blaber. 1986. Feeding ecology of three species of midwater fishes
associated with the continental slope of eastern Tasmania, Australia. Marine Biology 93:
147-156.
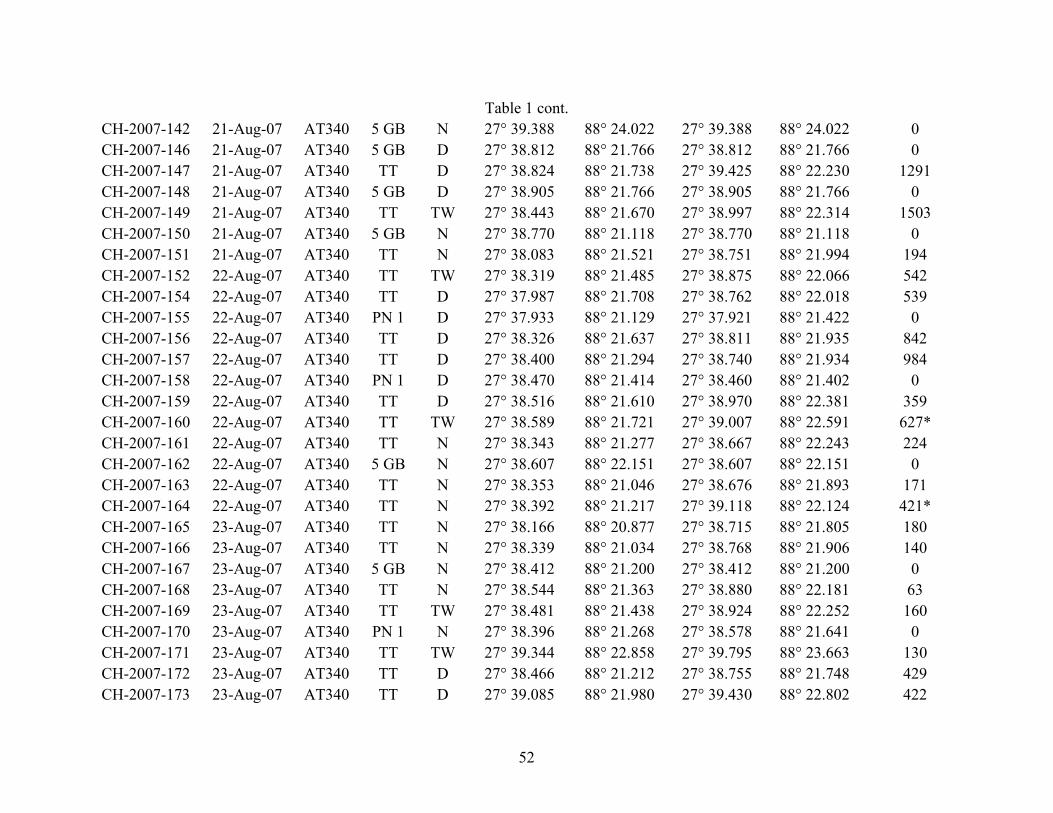
Table 1. Surface and midwater stations sampled over three cold seep sites (AT340, GC852, and AC601) (see Fig.1) in the Gulf of Mexico (9-25 August 2007). TT = Tucker trawl including plankton net inside Tucker trawl, PN 1 = 0.5 m dia. plankton net, PN 2 = 1 m dia. plankton net, NN = Neuston net, 5 GB = 5 gallon bucket for POM samples, D = day (0730 to 1830 hr CDT), N = night (2030 to 0530 hr CDT), TW = twilight (0530 to 0730 and 1830 to 2030 hr CDT). * = maximum depths sampled for non discrete tows (TT did not close and fished to surface). Blanks in depth columns indicated TDR did not record any data.
Table 2. The total number of all midwater fishes, invertebrates and autotrophs examined in dietary analyses from the North-central GOM. GCA = gut content analysis. SIA = stable isotope analysis, SL range = standard length size range for fish species (mm). Fish species marked with an * were grouped at family level for all analyses. Fish species marked with ^ were grouped at genera for stable isotope analyses. Species GCA SIA SL range FISH
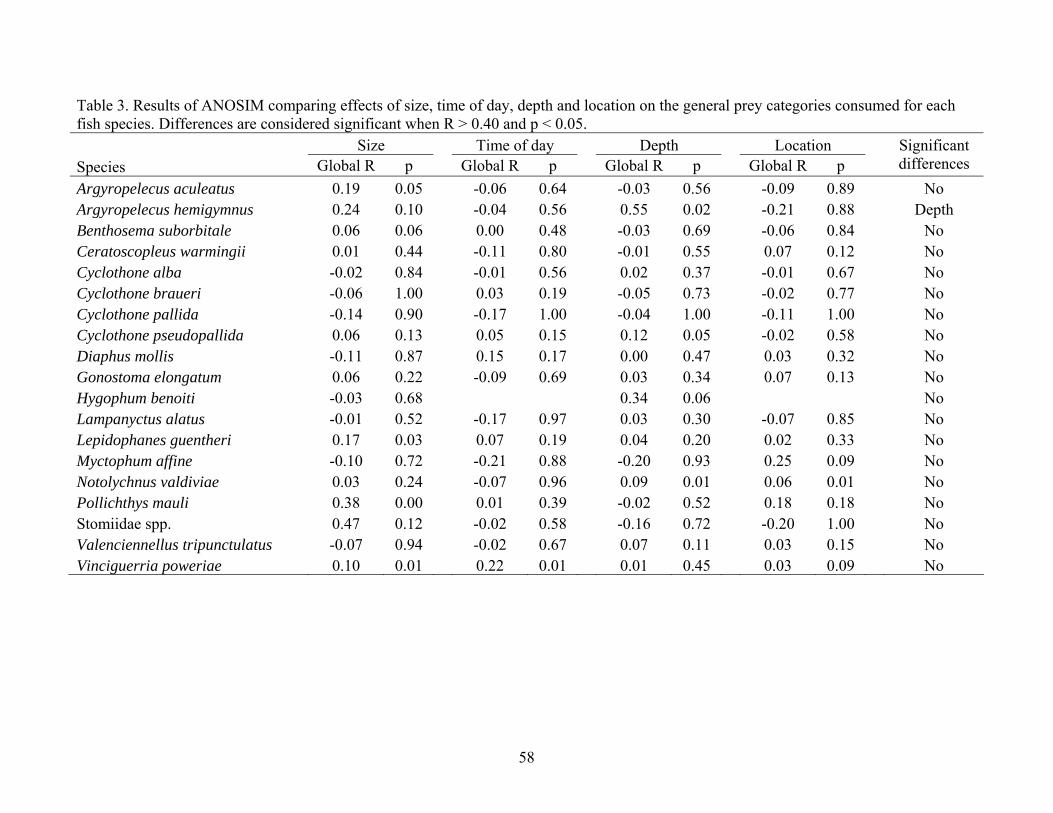
Table 3. Results of ANOSIM comparing effects of size, time of day, depth and location on the general prey categories consumed for each fish species. Differences are considered significant when R > 0.40 and p < 0.05. Size Time of day Depth Location Species Global R p Global R p Global R p Global R p
Table 4. Percent volume and frequency of prey items consumed by Chauliodus sloani collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 2 n = 8 n = 37 n = 2 n = 5 n = 7 n = 2 E = 2 E = 7 E = 30 E = 1 E = 5 E = 6 E = 1 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F FISH 100.0 100.0 86.5 28.6 100.0 100.0 100.0 100.0 Bregmaceros spp. 100.0 100.0 Myctophidae 67.5 14.3 Unidentified fish parts 19.0 14.3 100.0 100.0 100.0 100.0 CRUSTACEA <0.1 14.3 Unidentified crustacean parts <0.1 14.3 OTHER 13.5 71.4 100.0 100.0 Organic material 13.0 57.1 100.0 100.0 Unidentified animal parts 0.5 14.3
59
Table 5. Percent volume and frequency of prey items consumed by Gonostoma elongatum collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 0 n = 2 n = 28 n = 3 n = 3 n = 42 n = 10 E = 0 E = 0 E = 11 E = 0 E = 0 E = 10 E = 0 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 0.7 11.8 0.3 3.1 Amphipoda 0.7 11.8 Scina pusilla 0.3 3.1 CHAETOGNATHA 3.4 5.9 Heterokrohnia sp. 3.4 5.9 CNIDARIA 0.4 5.9 Cnidaria 0.4 5.9 18.0 20.0 COPEPODA 2.2 35.3 0.4 33.3 1.6 34.4 Aetideus acutus <0.1 5.9 Calanoida 0.5 17.6 0.1 3.1 Candacia longimana 14.4 10.0 Copepoda 0.1 11.8 0.3 9.4 3.6 10.0 Corycaeus furcifer <0.1 33.3 Corycaeus sp. 0.1 5.9 <0.1 3.1 Eucalanidae <0.1 3.1 Gaetanus pileatus 0.4 3.1 Haloptilus sp. 0.6 5.9 Pareucalanus attenuatus 0.1 3.1 Pleuromamma xiphias 0.7 11.8 0.4 3.1 Rhincalanus cornutus 0.4 33.3 Temora stylifera <0.1 5.9
Table 6. Percent volume and frequency of prey items consumed by Stomiidae collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 1 n = 5 n = 13 n = 3 n = 0 n = 8 n = 6 E = 1 E = 2 E = 9 E = 2 E = 0 E = 6 E = 5 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F COPEPODA 0.1 33.3 Oncaea sp. 0 33.3 Unidentified copepod parts 0.1 33.3 DECAPODA 61.4 33.3 8.3 25.0 Decapoda 8.3 25.0 Penaeidae 61.4 33.3 FISH 7.5 66.7 27.5 25.0 100.0 100.0 80.7 50.0 100.0 100.0 Diaphus mollis 100.0 100.0 Myctophidae 44.2 50.0 Unidentified fish parts 7.5 66.7 27.5 25.0 36.4 50.0 100.0 100.0OTHER 31.1 66.7 19.3 100.0 Invertebrate <0.1 33.3 Nematoda 3.3 33.3 64.2 50.0 Organic material 27.7 33.3 58.7 50.0 19.3 100.0 Unidentified animal parts 5.5 50.0
62
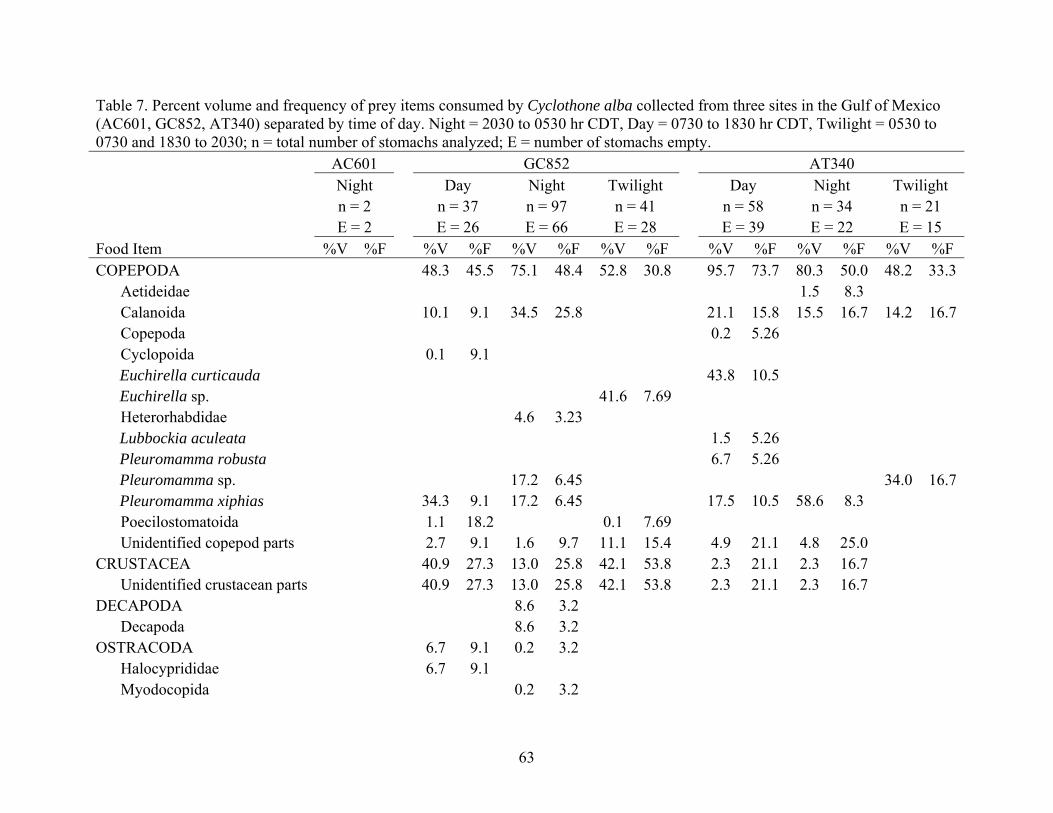
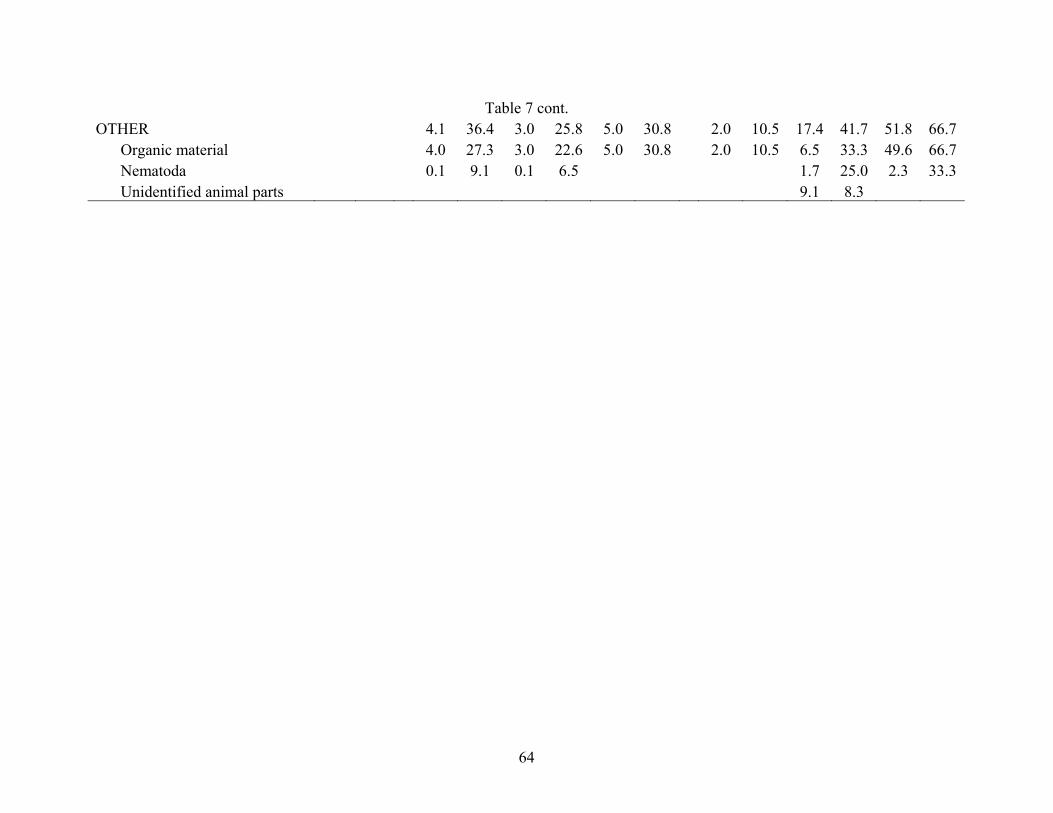
Table 7. Percent volume and frequency of prey items consumed by Cyclothone alba collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 2 n = 37 n = 97 n = 41 n = 58 n = 34 n = 21 E = 2 E = 26 E = 66 E = 28 E = 39 E = 22 E = 15 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F COPEPODA 48.3 45.5 75.1 48.4 52.8 30.8 95.7 73.7 80.3 50.0 48.2 33.3
Table 8. Percent volume and frequency of prey items consumed by Cyclothone braueri collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 2 n = 18 n = 100 n = 49 n = 55 n = 58 n = 37 E = 2 E = 14 E = 77 E = 34 E = 47 E = 41 E = 35 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F COPEPODA 100.0 100.0 64.3 39.1 53.0 66.7 42.4 87.5 69.2 52.9 7.0 50.0 Aegisthus mucronatus 57.2 25.0 19.7 13.3 16.2 25.0
OTHER 11.5 39.1 11.8 20.0 3.2 29.4 93.0 50.0 Organic material 11.5 39.1 7.6 13.3 3.2 29.4 93.0 50.0 Unidentified animal parts 4.2 6.7
65
Table 9. Percent volume and frequency of prey items consumed by Cyclothone pseudopallida collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 12 n = 90 n = 95 n = 34 n = 46 n = 34 n = 21 E = 11 E = 71 E = 70 E = 25 E = 37 E = 22 E = 20 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F COPEPODA 100.0 100.0 87.5 52.6 47.8 48.0 39.5 55.6 86.3 77.8 92.1 75.0 100.0 100.0 Aegisthus mucronatus 0.6 4.0 9.1 11.1
OTHER 3.0 36.8 29.0 32 1.6 22.2 5.0 22.2 <0.1 8.3 Animalia 2.7 11.1 Nematoda <0.1 5.3 0.1 4.0 Organic material 3.0 31.6 28.9 28.0 1.3 11.1 2.3 11.1 <0.1 8.3 Unidentified animal parts 0.3 11.1
67
Table 10. Percent volume and frequency of prey items consumed by Hygophum benoiti collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 6 n = 23 n = 37 n = 10 n = 7 n = 27 n = 1 E = 6 E = 23 E = 37 E = 10 E = 6 E = 11 E = 1 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 0.2 6.3
Unidentified copepod parts 25.4 68.8 CRUSTACEA 100.0 100.0 9.2 25.0
Unidentified crustacean parts 100.0 100.0 9.2 25.0 DECAPODA 3.8 6.3
Decapoda 3.8 6.3 MOLLUSCA 0.2 18.8
Bivalvia 0.2 18.8 OSTRACODA 0.6 12.5
Myodocopida 0.1 6.3 Ostracoda 0.5 6.3
OTHER 34.0 75.0 Organic material 34.0 75.0
68
Table 11. Percent volume and frequency of prey items consumed by Valenciennellus tripunctulatus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 1 n = 3 n = 37 n = 24 n = 9 n = 44 n = 29 E = 0 E = 0 E = 4 E = 3 E = 4 E = 12 E = 2 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 0.3 9.5
Amphipoda <0.1 4.8 Unidentified amphipod parts 0.3 4.8
Table 12. Percent volume and frequency of prey items consumed by Diaphus mollis collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 1 n = 0 n = 14 n = 3 n = 4 n = 10 n = 2 E = 0 E = 0 E = 0 E = 2 E = 0 E = 1 E = 0 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F CHAETOGNATHA 28.1 25.0 8.4 11.1
Sagittoidea 8.4 11.1 Unidentified chaetognath parts 28.1 25.0
Table 13. Percent volume and frequency of prey items consumed by Cyclothone pallida collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 18 n = 115 n = 95 n = 32 n = 58 n = 40 n = 4 E = 15 E = 109 E = 84 E = 32 E = 58 E = 39 E = 4 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 29.9 9.09
OTHER 8.5 33.3 9.5 33.3 42.1 63.6 100.0 100.0 Organic material 8.5 33.3 9.5 33.3 42.1 63.6 100.0 100.0
73
Table 14. Percent volume and frequency of prey items consumed by Vinciguerria poweriae collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 2 n = 6 n = 72 n = 19 n = 7 n = 45 n = 4 E = 0 E = 4 E = 21 E = 14 E = 3 E = 8 E = 2 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 14.3 17.6 12.8 25.0 1.7 5.4 Brachyscelus crusculum 4.7 2.0 Brachyscelus sp. 0.2 2.0 Eupronoe armata 1.6 2.0
Organic material 14.8 33.3 1.6 20.0 8.8 18.9 0.8 50.0Unidentified animal parts <0.1 7.8 <0.1 2.7
76
Table 15. Percent volume and frequency of prey items consumed by Myctophum affine collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 1 n = 10 n = 40 n = 6 n = 0 n = 4 n = 0 E = 0 E = 9 E = 21 E = 2 E = 0 E = 0 E = 0 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 48.2 15.8 26.0 25.0 26.0 25.0
OTHER 29.6 100.0 17.0 52.6 49.5 50.0 40.1 75.0 Animalia <0.1 5.3 Organic material 26.1 100.0 16.9 47.4 49.5 50.0 40.1 75.0 Phytoplankton 0.1 5.3 Unidentified animal parts 3.5 100.0 <0.1 5.3
78
Table 16. Percent volume and frequency of prey items consumed by Argyropelecus aculeatus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 3 n = 0 n = 22 n = 0 n = 4 n = 8 n = 3 E = 0 E = 0 E = 7 E = 0 E = 1 E = 2 E = 0 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 0.4 33.3 7.2 26.7 5.1 33.3 0.4 33.3 28.4 100.0
OTHER 24.7 33.3 31.5 60.0 6.8 33.3 26.9 83.3 18.4 66.7 Organic material 24.7 33.3 25.6 60.0 6.8 33.3 26.9 83.3 18.3 66.7 Unidentified animal parts 5.9 6.7 0.2 33.3
80
Table 17. Percent volume and frequency of prey items consumed by Argyropelecus hemigymnus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 1 n = 2 n = 8 n = 7 n = 3 n = 6 n = 3 E = 1 E = 0 E = 5 E = 4 E = 1 E = 5 E = 3 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %FAMPHIPODA 0.9 50.0
Table 18. Percent volume and frequency of prey items consumed by Pollichthys mauli collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 0 n = 0 n = 2 n = 0 n = 15 n = 21 n = 25 E = 0 E = 0 E = 0 E = 0 E = 6 E = 4 E = 5 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 63.5 50.0 16.0 5.0
Amphipoda 16.0 5.0 Unidentified amphipod parts 63.5 50.0
OTHER 2.2 11.1 4.2 17.6 3.2 20.0 Organic material 2.2 11.1 4.2 17.6 3.2 20.0
83
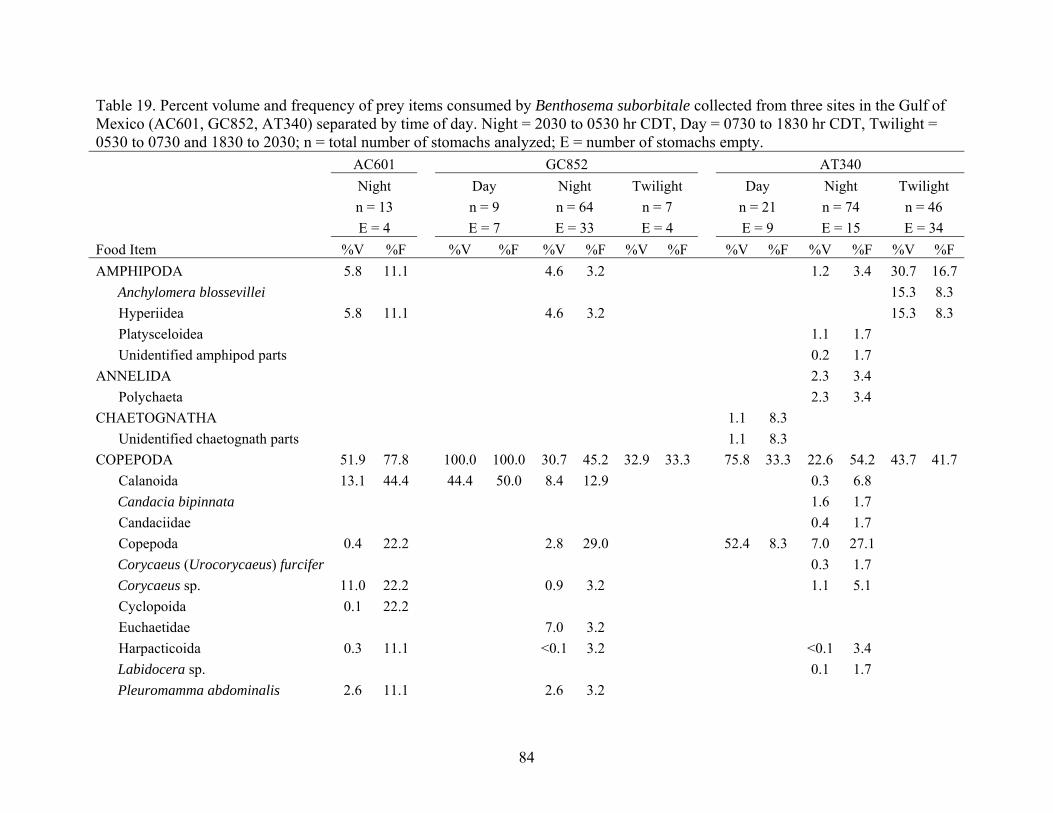
Table 19. Percent volume and frequency of prey items consumed by Benthosema suborbitale collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 13 n = 9 n = 64 n = 7 n = 21 n = 74 n = 46 E = 4 E = 7 E = 33 E = 4 E = 9 E = 15 E = 34 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 5.8 11.1 4.6 3.2 1.2 3.4 30.7 16.7 Anchylomera blossevillei 15.3 8.3
Table 20. Percent volume and frequency of prey items consumed by Lampanyctus alatus collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 6 n = 5 n = 25 n = 12 n = 3 n = 16 n = 5 E = 0 E = 3 E = 4 E = 4 E = 0 E = 4 E = 0 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 30.4 33.3 20.1 23.8 28.4 12.5
Table 21. Percent volume and frequency of prey items consumed by Lepidophanes guentheri collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 15 n = 4 n = 36 n = 7 n = 25 n = 57 n = 13 E = 0 E = 1 E = 11 E = 2 E = 8 E = 9 E = 7 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 39.3 8.0 10.9 5.9 0.2 2.1
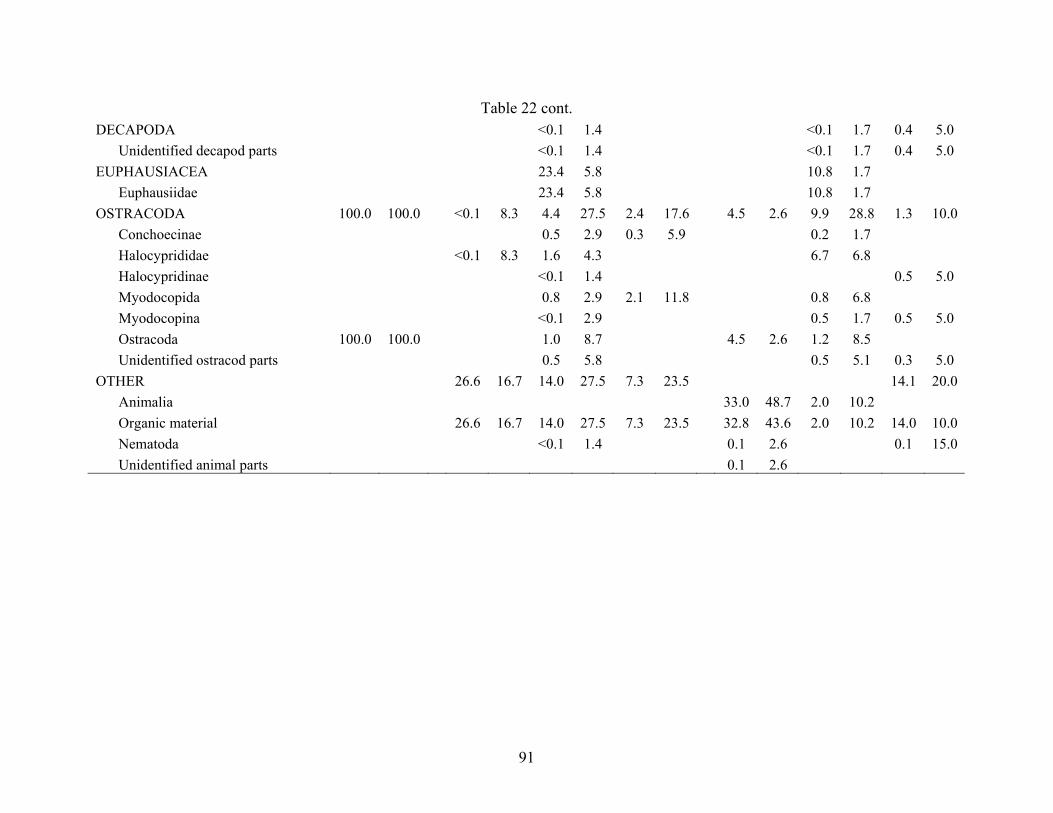
Table 22. Percent volume and frequency of prey items consumed by Notolychnus valdiviae collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 1 n = 14 n = 92 n = 30 n = 71 n = 94 n = 41 E = 0 E = 2 E = 23 E = 13 E = 32 E = 35 E = 21 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F CHAETOGNATHA <0.1 5.9
Table 23. Percent volume and frequency of prey items consumed by Ceratoscopelus warmingii collected from three sites in the Gulf of Mexico (AC601, GC852, AT340) separated by time of day. Night = 2030 to 0530 hr CDT, Day = 0730 to 1830 hr CDT, Twilight = 0530 to 0730 and 1830 to 2030; n = total number of stomachs analyzed; E = number of stomachs empty. AC601 GC852 AT340 Night Day Night Twilight Day Night Twilight n = 5 n = 6 n = 23 n = 5 n = 6 n = 39 n = 4 E = 1 E = 4 E = 4 E = 2 E = 3 E = 1 E = 1 Food Item %V %F %V %F %V %F %V %F %V %F %V %F %V %F AMPHIPODA 54.6 50.0 8.8 31.6 0.4 33.3 0.8 13.2
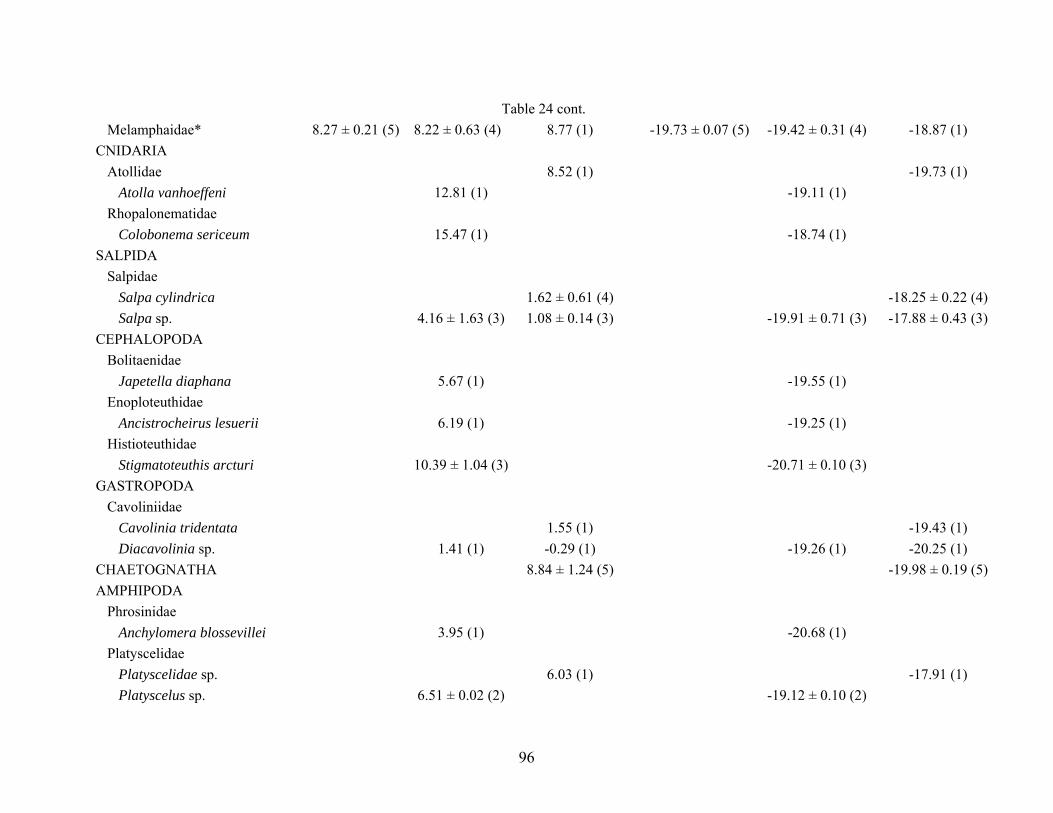
Table 24. Mean (± 1 standard error) δ13C and δ15N values for midwater fishes, invertebrates and carbon sources collected from each site (AC601, AT340, GC852). n = number in parentheses, * = multiple fish species grouped together δ15N δ13C Species AC601 GC852 AT340 AC601 GC852 AT340 FISH
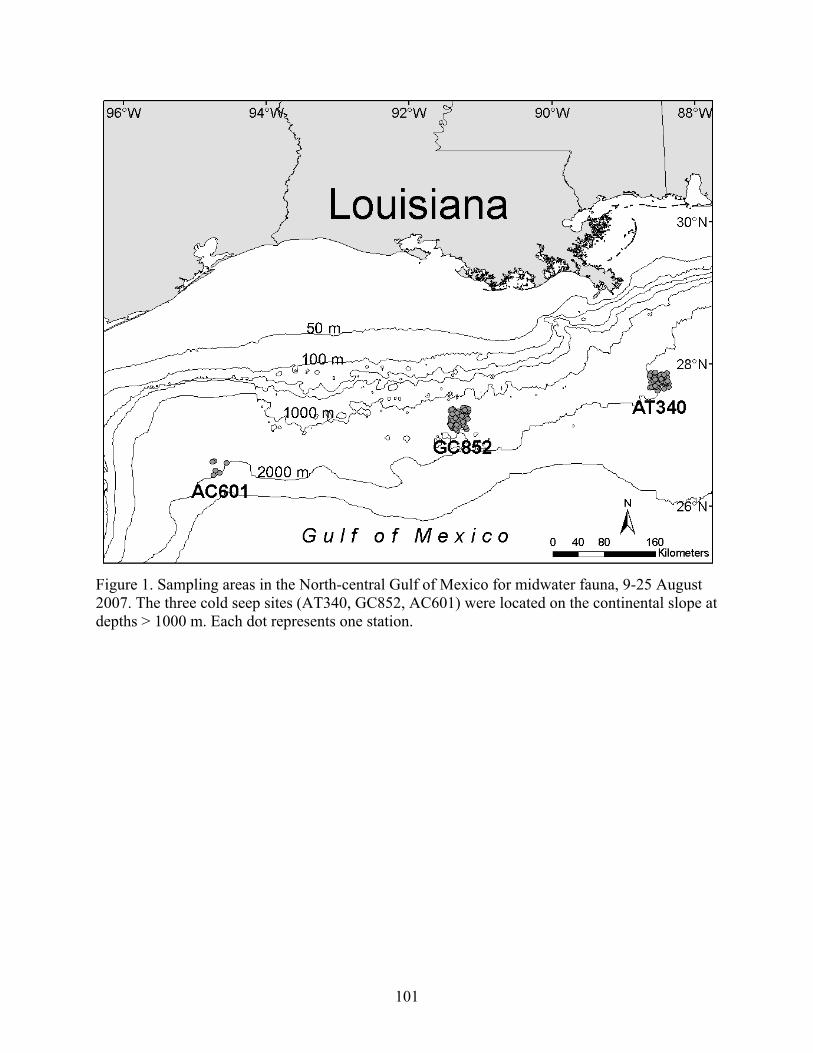
Figure 1. Sampling areas in the North-central Gulf of Mexico for midwater fauna, 9-25 August 2007. The three cold seep sites (AT340, GC852, AC601) were located on the continental slope at depths > 1000 m. Each dot represents one station.
101
Figure 2. Multidimensional scaling (MDS) plot documenting the differences among the gut contents of midwater fishes. Data were based on the Bray-Curtis similarity matrix calculated from standardized, square root transformed, mean volumes of prey (12 general categories). Colors represent the different fish families, red = Gonostomatidae, Orange = Sternoptychidae, Green = Phosichthyidae, Blue = Stomiidae, Purple = Myctophidae. Ca = Cyclothone alba, Cb = Cyclothone braueri, Cp = Cyclothone pallida, Cps = Cyclothone pseudopallida, Ge = Gonostoma elongatum, Aa = Argyropelecus aculeatus, Ah = Argyropelecus hemigymnus, Vt = Valenciennellus tripunctulatus, Pm = Pollichthys mauli, Vp = Vinciguerria poweriae, Cs = Chauliodus sloani, St = Stomiidae, Bs = Benthosema suborbitale, Cw = Ceratoscopelus warmingii, Dm = Diaphus mollis, Hb = Hygophum benoiti, La = Lampanyctus alatus, Lg = Lepidophanes guentheri, Ma = Myctophum affine, Nv = Notolychnus valdiviae. Clusters are defined at 30% (solid black line) and 60% (dashed black line) similarities.
102
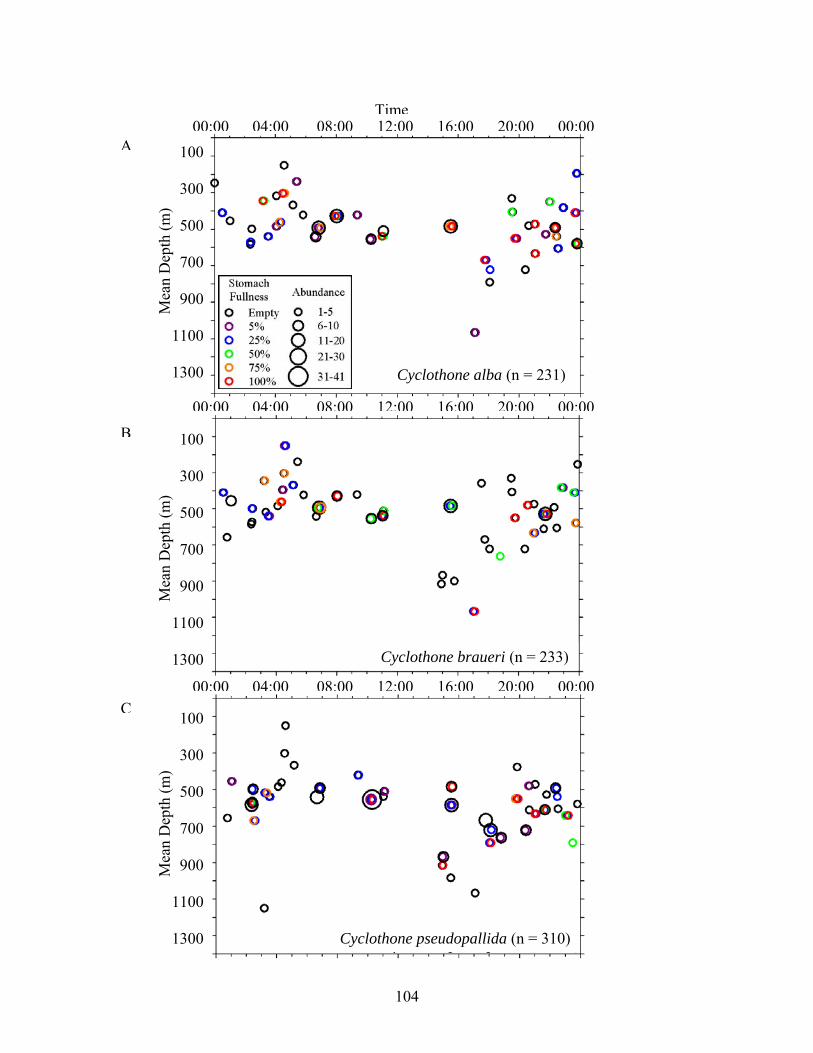
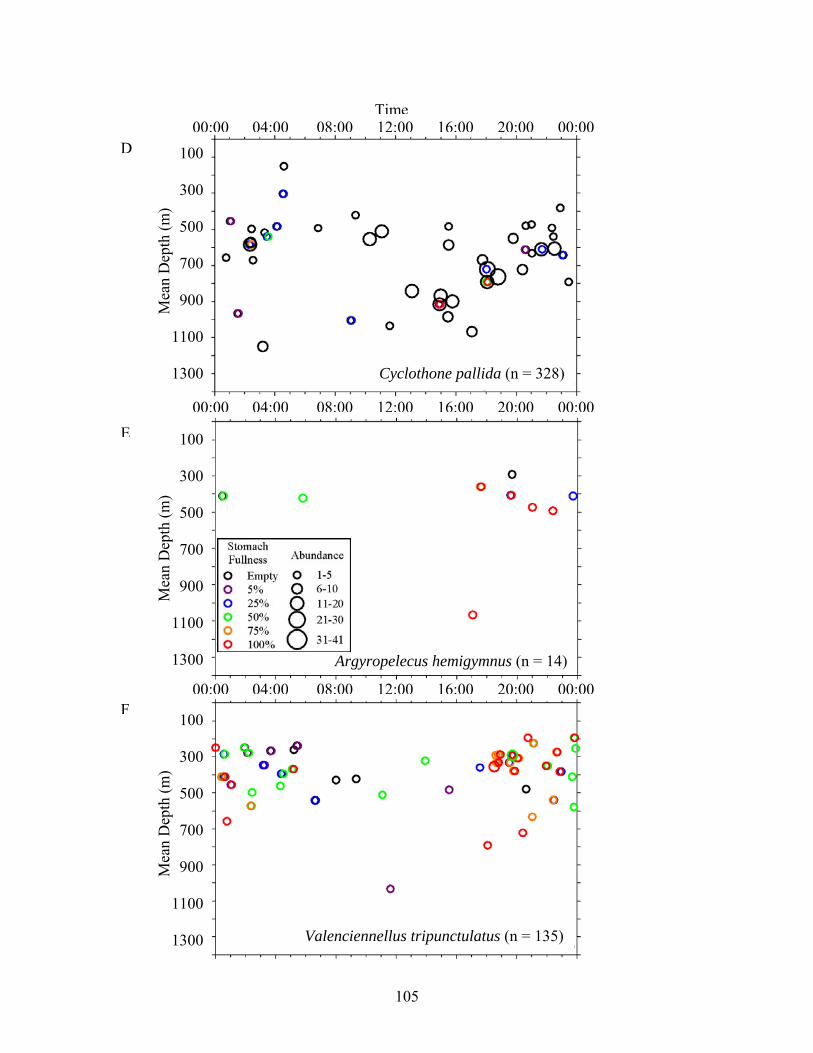
Figure 3. Relationships among stomach fullness, mean depth of capture and time for midwater fishes. Data were compiled from all sites and excluded specimens that lacked depth data. A) C. alba, B) C. braueri, C) C. pseudopallida, D) C. pallida, E) A. hemigymnus, F) V. tripunctulatus, G) G. elongatum, H) A. aculeatus, I) P. mauli, J) V. poweriae, K) B. suborbitale, L) C. warmingii, M) D. mollis, N) H. benoiti, O) L. alatus, P) L. guentheri, Q) N. valdiviae, R) M. affine, S) C. sloani, T) Stomiidae.
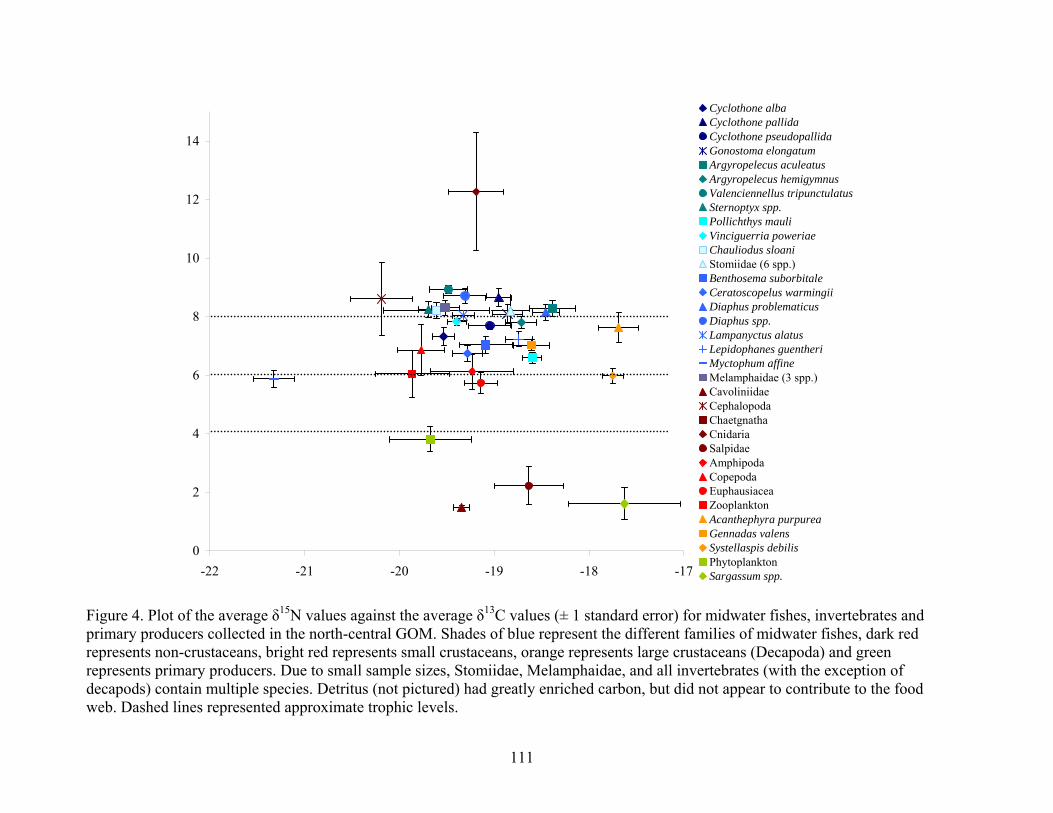
Figure 4. Plot of the average δ15N values against the average δ13C values (± 1 standard error) for midwater fishes, invertebrates and primary producers collected in the north-central GOM. Shades of blue represent the different families of midwater fishes, dark red represents non-crustaceans, bright red represents small crustaceans, orange represents large crustaceans (Decapoda) and green represents primary producers. Due to small sample sizes, Stomiidae, Melamphaidae, and all invertebrates (with the exception of decapods) contain multiple species. Detritus (not pictured) had greatly enriched carbon, but did not appear to contribute to the food web. Dashed lines represented approximate trophic levels.
A B
6.0
6.5
7.0
7.5
8.0
8.5
18 20 22 24 26 28 30 32 34 36
R2 = 0.223p = 0.646
R2 = 0.367p = 0.064
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
10.0
10.5
25 30 35 40 45 50
C D
R2 = 0.736p = 0.002
6.8
7.0
7.2
7.4
7.6
7.8
8.0
8.2
8.4
10 20 30 40 50
R2 = 0.618p = 0.002
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
10.0
0 50 100 150 200
E F
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
10.0
0 10 20 30 40 50 60
R2 = 0.400p = 0.068
6.0
6.5
7.0
7.5
8.0
8.5
9.0
0 10 20 30 40
R2 = 0.274p = 0.329
SL (mm)
δ15N
Figure 5. Relationship between δ15N and SL for midwater fish species. A) Cyclothone alba, B) Cyclothone pallida, C) Cyclothone pseudopallida, D) Gonostoma elongatum, E) Argyropelecus aculeatus, F) Argyropelecus hemigymnus, G) Sternoptyx spp., H) Valenciennellus tripunctulatus, I) Pollichthys mauli, J) Vinciguerria poweriae, K) Chauliodus sloani, L) Stomiidae, M) Benthosema suborbitale, N) Ceratoscopelus warmingii, O) Diaphus spp., P) Diaphus problematicus, Q) Lampanyctus alatus, R) Lepidophanes guentheri, S) Myctophum affine, T) Melamphaidae. The lines correspond to linear regressions.
112
G H
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
0 10 20 30 40 50 607.0
7.5
8.0
8.5
9.0
9.5
10.0
16 18 20 22 24 26 28
R2 = 0.123R2 = 0.509p = 0.399p = 0.128
I J
R2 = 0.331p = 0.082
5.0
5.5
6.0
6.5
7.0
7.5
8.0
24 29 34 39
R2 = 0.614p < 0.001
6.0
6.5
7.0
7.5
8.0
8.5
9.0
18 20 22 24 26 28 30 32
δ15N
K L
7.0
7.5
8.0
8.5
9.0
9.5
0 50 100 150 200 250
R2 = 0.852p = 0.009
R2 = 0.355p = 0.053
4.0
5.0
6.0
7.0
8.0
9.0
10.0
11.0
12.0
0 50 100 150 200 250 300 SL (mm)
Figure 5 cont. Relationship between δ15N and SL for all midwater fish species. A) Cyclothone alba, B) Cyclothone pallida, C) Cyclothone pseudopallida, D) Gonostoma elongatum, E) Argyropelecus aculeatus, F) Argyropelecus hemigymnus, G) Sternoptyx spp., H) Valenciennellus tripunctulatus, I) Pollichthys mauli, J) Vinciguerria poweriae, K) Chauliodus sloani, L) Stomiidae, M) Benthosema suborbitale, N) Ceratoscopelus warmingii, O) Diaphus spp., P) Diaphus problematicus, Q) Lampanyctus alatus, R) Lepidophanes guentheri, S) Myctophum affine, T) Melamphaidae. The lines correspond to linear regressions.
113
M N
6.0
6.5
7.0
7.5
8.0
18 19 20 21 22 23 24 25 26 27 28
R2 = 0.735p = 0.307
4.00
4.50
5.00
5.50
6.00
6.50
7.00
7.50
8.00
8.50
0 10 20 30 40 5
R2 = 0.356
0
p = 0.149
O P
R2 = 0.838p = 0.029
7.0
7.5
8.0
8.5
9.0
9.5
10.0
25 35 45 55 65 75
R2 = 0.738p = 0.013
7.0
7.5
8.0
8.5
9.0
9.5
40 45 50 55 60 65 70 75 80
δ15N
Q R
5.00
5.50
6.00
6.50
7.00
7.50
8.00
8.50
9.00
R2 = 0.113p = 0.220
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
25 30 35 40 45 50 55 35 40 45 50 55 60 65 70
R2 = 0.652p = 0.138
30 SL (mm) Figure 5 cont. Relationship between δ15N and SL for all midwater fish species. A) Cyclothone alba, B) Cyclothone pallida, C) Cyclothone pseudopallida, D) Gonostoma elongatum, E) Argyropelecus aculeatus, F) Argyropelecus hemigymnus, G) Sternoptyx spp., H) Valenciennellus tripunctulatus, I) Pollichthys mauli, J) Vinciguerria poweriae, K) Chauliodus sloani, L) Stomiidae, M) Benthosema suborbitale, N) Ceratoscopelus warmingii, O) Diaphus spp., P) Diaphus problematicus, Q) Lampanyctus alatus, R) Lepidophanes guentheri, S) Myctophum affine, T) Melamphaidae. The lines correspond to linear regressions.
114
S T
R2 = 0.408p = 0.047
4.0
4.5
5.0
5.5
6.0
6.5
7.0
7.5
13 14 15 16 17 18 19
R2 = 0.687p = 0.003
6.0
6.5
7.0
7.5
8.0
8.5
9.0
9.5
10.0
12 14 16 18 20 22 24 26 28
δ15N
SL (mm) Figure 5 cont. Relationship between δ15N and SL for all midwater fish species. A) Cyclothone alba, B) Cyclothone pallida, C) Cyclothone pseudopallida, D) Gonostoma elongatum, E) Argyropelecus aculeatus, F) Argyropelecus hemigymnus, G) Sternoptyx spp., H) Valenciennellus tripunctulatus, I) Pollichthys mauli, J) Vinciguerria poweriae, K) Chauliodus sloani, L) Stomiidae, M) Benthosema suborbitale, N) Ceratoscopelus warmingii, O) Diaphus spp., P) Diaphus problematicus, Q) Lampanyctus alatus, R) Lepidophanes guentheri, S) Myctophum affine, T) Melamphaidae. The lines correspond to linear regressions.