Page 1
ORIGINALARTICLE
Twentieth century demographic changesin cirio and cardon in Baja California,Mexico
Stephen H. Bullock1*, Nora E. Martijena1, Robert H. Webb2 and Raymond
M. Turner2
1Departamento de Biologıa de la Conservacion,
Centro de Investigacion Cientıfica y de
Educacion Superior de Ensenada, Baja
California, Mexico and 2US Geological Survey,
Tucson, AZ, USA
*Correspondence: Stephen H. Bullock,
Departamento de Biologıa de la Conservacion,
Centro de Investigacion Cientıfica y de
Educacion Superior de Ensenada, Apartado
Postal 2732, 22800 Ensenada, Baja California,
Mexico. E-mail: [email protected]
ABSTRACT
Aim Our purpose was to discern long-term large-scale patterns of survivorship
and recruitment of two dominant, charismatic and useful desert plants, cirio
[Fouquieriaceae; Fouquieria columnaris (Kell.) Kell. ex Curran] and cardon
[Cactaceae; Pachycereus pringlei (S.Watson) Britton & Rose], and to evaluate the
effects of physical and human factors.
Location The study included 77 sites distributed over c. 13,200 km2 in the
northern Vizcaıno Region of the Sonoran Desert, state of Baja California, Mexico.
Method Censuses used identified individuals (n ¼ 3780 cirio and 2246 cardon)
and were based on repeat photography. Time intervals between censuses ranged
from 29 to 96 years, ending in 1996–2002. Earlier repetition (1963) of old
photographs at 14 sites allowed analysis of temporal variation. The effect of
elapsed time was modelled with Weibull functions for survivorship and linear
functions for recruitment. To explain the residual variations, we tested categorical
and continuous variables related to substrate, geomorphology, climate and
geography. Human impacts were weighed with historical and quantitative
analysis.
Results Elapsed time accounted for 45% of the variation of survivorship in cirio
but only 35% in cardon. The fitted Weibull curves were used to estimate
longevity: all-size cohorts of 1000 individuals would die out in 388 (cirio) and
723 years (cardon). Recruitment in cirio was linearly related to time (r2 ¼ 0.71)
and averaged < 0.003 year)1 but was less than mortality at 90% of the sites. For
cardon, recruitment averaged > 0.005 year)1, was linearly related to time
(r2 ¼ 0.66) and was less than mortality at only 33% of the sites. Vital rates
were not strongly correlated between mid- and late-century but were mostly
similar to the long-term regional rates. Removing the factor of elapsed time, the
residual variations showed that survivorship was greater on sites protected from
winds for cirio and was less on very coarse soils for cardon. Recruitment increased
with latitude in cirio and was greater on southern exposures for both species. The
residual variations were not clearly affected, at a regional scale, by other factors
thought to be important, including elevation, distance to the Pacific Ocean,
geology, slope gradient, soil stability, older vs. young alluvial soils, and soil
Ca : Mg and Na : K ratios. Human impacts have been sporadic and
heterogeneous but locally strong; our quantitative indices of accessibility did
not show regionally significant effects. Blowdown by hurricanes is a sporadic
cause of major mortality for cirio but not necessarily for cardon.
Main conclusions At our scales, effects of time outweigh those of substrate, and
human impact was scant. Large patterns were pronounced: cirio experienced a
prolonged, widespread decline in the 1900s, while cardon fluctuated in different
directions and degree among local populations. Cirio was found to be inherently
Journal of Biogeography (J. Biogeogr.) (2005) 32, 127–143
ª 2005 Blackwell Publishing Ltd www.blackwellpublishing.com/jbi 127
Page 2
much shorter-lived than cardon. We also suggest that recruitment in cirio was
low due to long periods with relatively dry winters that affected the entire region,
while spatial heterogeneity of cardon recruitment seemed more related to the
variation in summer rains.
Keywords
Climatic variability, demography, Fouquieria, human impact, hurricanes, land-
scape, Metapopulation, Pachycereus, Sonoran Desert, substrate.
RESUMEN
Metas Nuestro proposito fue discernir, en escalas de largo plazo y
geograficamente amplias, patrones de sobrevivencia y reclutamiento de dos
plantas carismaticas, utiles y dominantes del desierto: el cirio [Fouquieriaceae;
Fouquieria columnaris (Kell.) Kell. ex Curran] y el cardon [Cactaceae; Pachycereus
pringlei (S.Watson) Britton & Rose]. A su vez, evaluar los efectos de factores
fısicos y humanos sobre estos patrones.
Localizacion El estudio incluyo 77 sitios distribuidos en c. 13,200 km2 en el
norte de la Region Vizcaıno del Desierto Sonorense, en Baja California, Mexico.
Metodo Con censos basados en la repeticion de fotografıas se identificaron los
individuos (n ¼ 3780 cirios y 2246 cardones). Los intervalos de tiempo entre los
censos variaron entre 29 y 96 anos, terminando en 1996–2002. La repeticion en
1963 de viejas fotografıas en 14 sitios, permitio el analisis de la variacion
temporal. El efecto del tiempo transcurrido fue modelado con funciones
Weibull para la sobrevivencia y con funciones lineares en el caso del
reclutamiento. Para explicar las variaciones residuales, se pusieron a prueba
variables categoricas y continuas relacionadas a substrato, geomorfologıa, clima y
geografıa. Los impactos humanos se consideraron mediante analisis historico y
cuantitativo.
Resultados El tiempo transcurrido explico el 45% de la variacion en la
sobrevivencia del cirio pero solamente el 35% en la del cardon. El ajuste a la
funcion Weibull permitio estimar la longevidad: las cohortes de todos los
tamanos de 1000 individuos morirıan en 388 (para el cirio) y 723 anos (para el
cardon). El reclutamiento del cirio estuvo linearmente relacionado al tiempo
(r2 ¼ 0.71) y promedio < 0.003 ano)1 pero fue menor que la mortalidad en el
90% de los sitios. Para el cardon, el reclutamiento promedio fue de > 0.005
ano)1, estuvo linearmente relacionado al tiempo (r2 ¼ 0.66) y fue menor que la
mortalidad en solamente el 33% de los sitios. Las tasas vitales no estuvieron
fuertemente correlacionadas entre mediados y finales del siglo pero fueron
principalmente similares a las tasas regionales de largo plazo. Removiendo el
factor del tiempo transcurrido, las variaciones residuales mostraron que la
sobrevivencia fue mayor en sitios protegidos del viento para el cirio y fue menor
en suelos muy gruesos para el cardon. El reclutamiento incremento con la latitud
para el cirio y fue mayor en las exposiciones sur para ambas especies. Las
variaciones residuales no fueron claramente afectadas, a una escala regional, por
otros factores considerados como importantes, incluyendo la elevacion, la
distancia al Oceano Pacıfico, la geologıa, el gradiente de pendiente, la estabilidad
del suelo, la caracterizacion de los suelos aluviales en jovenes vs. viejos o las
proporciones de Ca:Mg o Na:K en el suelo. Los impactos humanos han sido
esporadicos y heterogeneos pero localmente muy fuertes; nuestros ındices
cuantitativos de accesibilidad no mostraron efectos regionalmente significativos.
S. H. Bullock et al.
128 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 3
INTRODUCTION
In arid lands, moisture in the upper soil is only adequate for
the growth of most plants at sporadic intervals and then only
for short periods. Changes in individuals are often difficult to
perceive in these conditions, as are the trends of change in
populations. Some individual-based population studies have
extended to periods of several years to decades, but it remains
difficult to define both the spatio-temporal patterns of change
and their causes (Watson et al., 1997; Turner et al., 2003).
Large, succulent-stemmed species have been particularly well-
studied in Arizona, where populations of some of these and
other common or dominant species are far from static, at least
at the local level (Shreve, 1910; Niering et al., 1963; Tschirley &
Wagle, 1964; Steenbergh & Lowe, 1983; Goldberg & Turner,
1986; Parker, 1987,1988; Turner, 1990; Bowers et al., 1995;
Bowers, 1997; Pierson & Turner, 1998; Turner et al., 2003).
The dynamics of these populations and their ecosystems have
been affected strongly by human activity, directly and
indirectly (Niering et al., 1963; Bahre & Hutchinson, 1985;
Humphrey, 1987; Bahre & Shelton, 1993) with few exceptions
(Turner, 1990; Bowers et al., 1995). Climate variation also has
been a major factor (Brum, 1973; Turner, 1990; Pierson &
Turner, 1998; Turner et al., 2003), and there is evidence or
suggestions of other biotic or endogenous factors (Tschirley &
Wagle, 1964; Steenbergh & Lowe, 1983; McAuliffe, 1988).
Contrasts in population dynamics across substrates have
also been exemplified (Niering et al., 1963; Parker, 1987;
McAuliffe, 1994; Bowers, 1997; El-Keblawy et al., 1997;
Pierson & Turner, 1998). For a locality in the Vizcaıno region,
McAuliffe (1991) showed that the size structures differed
between adjacent alluvial terraces with different soil character-
istics, for populations of three dominant perennials: cirio
[Fouquieria columnaris (Kell.) Kell. ex Curran, Fouquieria-
ceae], cardon [Pachycereus pringlei (S. Watson) Britton & Rose,
Cactaceae] and gobernadora [Larrea tridentata (Moc. & Ses.)
Cav., Zygophyllaceae]. However, the interpretations may be
confounded by the problems of age estimation (Bullock et al.,
2004) and the probable instability of the populations on scales
of decades or centuries (see below). Substrate age can be
important, but a regional application could be confounded by
the diversity of geology, geomorphology and history.
In the context of the entire Sonoran Desert, or the world’s
deserts in general, the Vizcaıno Region of the Baja California
peninsula (Shreve, 1964) has been much less affected by
humans due to the scant physical and social infrastructure and
to the rough terrain and scarcity of water. Nonetheless, human
impacts cannot be neglected, nor can the persistent demands
to exploit some large perennial plants; technical assessments
are still needed to guide consideration of management options
(INE-SEMARNAP, 1997).
Cirio and cardon, woody succulents which often exceed 8 m
height (Fig. 1a), are two of the dominant and charismatic
plants of the Vizcaıno region. Both species have been subjected
to undocumented extraction for useful and decorative wood
and as ornamental plants. Both species are obliging subjects for
long-term measurements of growth, recruitment and mortality
due to their columnar form, but biological and population
data have been insufficient to evaluate their status or to
establish use or conservation plans. Current estimates of
longevity for cirio are 200–400 years (Bullock et al., 2004),
which is considerably less than previous estimates (Humphrey
& Humphrey, 1990; McAuliffe, 1991), while cardon reproduc-
tion may not start until age 100 years (M. Salazar & S.H.
Bullock, unpubl. data).
The present report explores long-term survivorship and
recruitment trends for these species. The study is unique for its
resolution, based on identified individuals, in combination
with a broad geographical and time scope, including 77 sites
scattered over c. 2� of latitude (Fig. 2), with census intervals of
29–96 years. It was possible to achieve this combination
through the use of repeat photography (Rogers et al., 1984;
Fig. 1a,b). We then evaluate several factors, which may affect
differences among sites in population dynamics, including
El derribamiento por huracanes fue una causa esporadica de mayor mortalidad en
el cirio pero no necesariamente en el cardon.
Principales conclusiones En nuestras escalas, los efectos del tiempo pesan mas
que los del substrato y el impacto humano es escaso. Los grandes patrones son
pronunciados: el cirio experimento una prolongada y amplia disminucion en los
1900s, mientras el cardon fluctuo en diferentes direcciones y grados entre
poblaciones locales. Se encontro que el cirio es inherentemente una planta de
muchamas corta vida que el cardon. Tambien sugerimos que el reclutamiento en el
cirio fue bajo debido a largos periodos con inviernos relativamente secos que
afectaron la region entera, mientras que la heterogeneidad espacial en el
reclutamiento del cardon parecio corresponder a la variacion en la lluvias de verano.
Palabras claves
Metapoblaciones, paisaje, demografıa, variabilidad climatica, huracanes,
impacto humano, substrato, Desierto Sonorense, Fouquieria, Pachycereus.
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 129
Page 4
time, the history of human impacts, geographical situation and
substrate conditions. We explicitly compared survivorship and
recruitment rates between the early vs. late twentieth century,
and between soils derived from granite vs. basalt and old vs.
new alluvium, because of their contrasting hydrology and
nutrient status (McAuliffe, 1991; Graham & Franco-Vizcaıno,
1992; Franco-Vizcaıno et al., 1993; Franco-Vizcaıno, 1994).
We also consider the impacts of hurricanes and of long-term
variations in precipitation.
MATERIALS AND METHODS
Species
Cirio has the general form of a tapered column; its greatest
reported height is 26.4 m (Humphrey, 1991). The column is
sustained by lignified tissue forming a fenestrated tube. Water-
storage tissue within the tube may develop large lacunae as a
result of drought. Orthotropic branches are usually absent or
sparse. Plagiotropic branches are non-succulent and have
xeromorphic xylem, in contrast to the trunk (Carlquist, 2001).
These lateral branches have leaf-bearing short-shoots, are
abundant and may be several decimetres long on small plants
but are typically extremely reduced after 2–3 m above ground
(Henrickson, 1975). Leaves can expand at any time of year, with
adequate moisture (Humphrey, 1974); stem water reserves may
contribute to maintaining leaf turgor (Nilsen et al., 1990).
Photosynthesis is all C3; stem photosynthesis may contribute to
maintenance respiration and the initial growth of new leaves
(Franco-Vizcaıno et al., 1990). The flowers are hermaphroditic,
open in summer and are visited by a variety of insects (Nabhan
et al., 1999). The seeds are dispersed by wind in late fall and
establishment is probably limited to the first winter or spring
(Humphrey, 1974).
Cardon is a columnar cactus that can reach 20 m height
(Turner et al., 1995). The column and the lesser or greater
abundance of large, orthotropic branches, are sustained by a
set of woody xylem strands that are separate or modestly
interconnected (Niklas et al., 1999). Variations in water
storage causes the stem to shrink or swell radially. Respiration
is sustained by CAM photosynthesis in the stem surface
(Franco-Vizcaıno et al., 1990). The reproductive biology of
cardon is complex and geographically variable. Samples from
two sites in our area were gynodioecious (Fleming et al., 1998;
pers. obs.), and experiments in a trioecious population in
Sonora showed hermaphrodites were self-compatible (Flem-
ing et al., 1994). Flowering is in spring; pollinators include
bats, birds and insects (Moran, 1962; Fleming et al., 1994).
Seeds are dispersed in summer by fruit-eating animals;
establishment may be restricted to summer and fall (Cancino
et al., 1993).
Our study area included about half the latitudinal range of
cirio and certainly the larger part of its total population. The
latitudinal range of cardon is considerably greater, extending
sparsely 1.5� north of our area, and almost 6� further south
(based on Turner et al., 1995). Both species also occur in
coastal Sonora; cirio occurs in an extremely limited area and
cardon is quite broadly distributed (Turner et al., 1995).
Geography and climate
The study area is within the Mexican state of Baja California
between 30.123 and 28.696�N latitude (Fig. 2) and includes a
large part of the federal protection area for flora and fauna
‘Valle de los Cirios’ (30–28�N; Diario Oficial de la Federacion,
2 June 1980, Primera Seccion, pp. 9–10). The great majority
(91%) of our sites are on the Pacific slope of the peninsula and,
on the north–south axis, are distributed from the San Vicente
and San Fernando watersheds to the San Borja watershed (see
Fig. 2 inset). This segment of the peninsula is c. 90 (to 120) km
wide, with most of the land surface above 200 m but only small
areas above 1000 m on the eastern and southern limits. The
elevation of our sites ranged from 55 to 780 m, with 58.4%
between 400 and 700 m.
Figure 1 Matched photographs of a site near San Fernando
mission, Baja California, featuring the columnar succulents cirio
[Fouquieria columnaris (Kell.) Kell. ex Curran] and cardon
[Pachycereus pringlei (S.Watson) Britton & Rose]. (a) 1905
(E. A. Goldman, courtesy of the Smithsonian Institution);
(b) 1998 (S. H. Bullock, CICESE).
S. H. Bullock et al.
130 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 5
The geological framework of the area consists of intrusive
felsic plutons (tonalites to granodiorites) and associated
metasedimentary rocks, extrusive basalts and intrusive ultra-
mafic rocks, marine sedimentary rocks, and Quaternary basin
fills (Gastil et al., 1975). Development of drainage networks is
chaotic, leading to a landscape patchwork of through-flowing
drainages and a few closed basins with playas. Relatively gentle
topography is of limited extent and diverse origin: a deposi-
tional plain SE of San Fernando; an erosional surface of the
batholith NW of Catavina; a rolling, basalt littered terrain NW
of Chapala; and a large valley with eroded marine sediments,
north and south of Punta Prieta.
Regional surveys of the soils include coarse scale mapping
with broad classification units and only scattered points of
verification (DETENAL, 1982). Fine-scale topographic vari-
ation hampers use of the soil maps for characterizing small
sites. However, the most widespread classes in our area are
lithic and rocky (INEGI, 2001). Soils are rarely sodic or
alkaline (DETENAL, 1982; Graham & Franco-Vizcaıno, 1992;
INEGI, 2001; E. Franco-Vizcaıno, pers. comm.). Caliche
(pedogenic carbonate) and thick petrocalic deposits are
prominent in some areas (INEGI, 2001) but have not been
well studied. Basalt-derived soils are notable for high clay
content, and granite-derived soils have high sand content. One
interesting feature in relation to the diversity and abundance of
succulent plants, is the variation in Ca:Mg ratio, although this
does not correspond to different parent rocks (Franco-
Vizcaıno et al., 1993), and so is not yet mappable.
The area is part of the Sonoran Desert as delimited according
to vegetation (Shreve, 1942) and climate (Schmidt, 1989) and
corresponds to the northern part of the Central Desert of the
Baja California peninsula as delimited by Aschmann (1959). In
terms of Shreve’s (1964) regions of the Sonoran Desert, the
study area corresponds to most of the northern Vizcaıno
region. Along with the Sonoran biogeographical affinity,
elements of a broad transition from/to California are common
(Shreve, 1936; Brown et al., 1992; Bullock, 1999).
Mean annual precipitation varies from115 to 153 mmamong
stations, but is non-normal in both seasonal and annual totals
(Bullock, 2003). Precipitation occurs predominately in the cool
season (61–91% in November–April) and for this season the
history of variation is highly correlated among stations in our
area (Bullock, 2003). Warm-season precipitation, of tropical or
low subtropical origin, can be extremely high but is usually very
local. The tropics also contribute occasional hurricanes (Court,
1980), which are perhaps the most erratic and unique feature of
this region’s climate; their high winds may be important to the
tall plants we studied. Available data showed no relation between
elevation and precipitation, but the contrast of Pacific and gulf
slope precipitation and the frequent incursion of coolmarine air
and even fog from the Pacific, produce an uneven gradient
inland from the west coast that strongly affects the peninsula’s
botanical characteristics (Shreve, 1964;Hastings&Turner, 1965;
Humphrey, 1974).
Reconstruction of cool-season precipitation from proxy
records indicated that exceptionally wet years were not only
Figure 2 Maps of the study area, with regional context, reference localities and census sites.
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 131
Page 6
rare but very unevenly distributed over the last 150 years: only
20 years between 1850 and 1997 had high precipitation
(January through March anomalies > 2.0 at San Diego), and
nine of these occurred after 1975 (Bullock, 2003). The recent
concentration of wet cool seasons raises the larger question of
whether the distribution of precipitation variability is station-
ary at the scale of > 50 years or if there have been larger shifts
or directional changes (Webb & Betancourt, 1992; Giorgi
et al., 1994; Meko et al., 1995; Turner et al., 2003). Attempts to
reconstruct the history of warm-season precipitation have not
been successful. Atmospheric and oceanic processes underlying
climatic patterns for the region have been discussed by various
authors (e.g. Garcıa & Mosino, 1968; Markham, 1972; Webb &
Betancourt, 1992).
Palaeoecology
The history of the biota of the Vizcaıno region is still difficult
to sketch. As recently as 10,000 years bp, pinyon–juniper
woodland occupied sites at Catavina and San Fernando
(Penalba & Van Devender, 1998; Wells, 2000; Sankey et al.,
2001) and a juniper–chaparral vegetation occupied a site south
of Sta. Gertrudis (Rhode, 2002); these sites are now dominated
by desert species. Pleistocene megafaunal fossils have been
found near Chapala (L. Davis, pers. com.) as well as further
south on the peninsula (Ferrusquıa-Villafranca & Torres-
Roldan, 1980); this fauna is unknown in the peninsula’s rock
art, some of which may predate 5000 years bp (Fullola et al.,
1993), although human occupation is evident from before
9000 years bp (Davis, 2003). In addition, repeated strong
changes of sea surface temperatures during at least the last
6000 years, which are evidenced by faunal changes in human
middens of marine molluscs (Moriarty, 1968), might have
affected the incursion of tropical storms, the oceanic moder-
ation of peninsular temperatures, and the occurrence of fog.
During this long period of human occupation, major climatic
fluctuations can be inferred from variations in lake levels of
now-dry playas (Arnold, 1957; Davis, 2003).
Human population and land-use
The pre-evangelization Cochimı population between the
territory of the San Ignacio mission (established 1728) and
San Fernando was estimated at 11,100 (Aschmann, 1959),
representing a density of c. 0.22 persons km)1. The economy
was entirely based on hunting and gathering of terrestrial and
marine resources. Concentration of the Cochimı at the
missions was impressive: San Borja had a maximum popula-
tion of c. 1800, and San Fernando reached c. 1400 (Aschmann,
1959, p. 182). However, the indigenous population declined
rapidly due to disease; by 1836, 15 years after Mexican
independence, the region’s mission population was less than
100 (Aschmann, 1959).
Secular occupation of the peninsula by people of mestizo or
European culture began at the end of the Jesuit period (1768),
unlike the early occupation (with livestock) of the eastern and
north-eastern areas of the Sonoran Desert (Turner et al.,
2003). Colonization was extremely scant North of Sta.
Gertrudis until the short-lived mining booms of the late
1800s, particularly the gold rush in the Calmallı district
starting in 1882 (Gabb, 1867; Deasy & Gerhard, 1944; Chaput
et al., 1992).
Many of the old ranches of the region are associated with
families that Martınez (1965) reported as immigrating to the
peninsula in the 1700s, but a historical account of the ranches
has yet to be made. Certainly the population was extremely
small until colonization efforts were promoted and subsidized
by the Mexican government in the 1970s and 1980s; the
network of dirt roads was vastly extended, and the density
(if not the dispersion) of ranches increased. In the period
1967–1979, the federal government established collective
landholdings (ejidos) that occupied almost the entire study
area (Fig. 3). The transpeninsular highway was first opened as
a dirt road in 1927, and was somewhat rerouted, graded and
paved in 1970–1973. A recent review estimated the population
of our area as roughly 572 individuals, with 50 permanent
settlements and 15 sporadically occupied localities, excluding
coastal fishing settlements that are more populous (Aramburo
et al., 2002). However, between the crash of the missions and
1930, censuses record only one settlement with > 100 inhab-
itants, the village at the onyx mine ‘El Marmol’ (c. 1930; Deasy
& Gerhard, 1944; Espinoza Arroyo, 1992). As of the year 2002,
Figure 3 Twentieth century historical events, including the
temporal distribution of old photographs (no. of sites recorded),
winter rainfall anomaly at San Diego [(observed ) mean)/SD],
territorial expansion of collective land holdings (ejidos), years with
tropical storms and hurricanes affecting the study area, years of
construction of the unpaved and paved transpeninsular highways,
years of ‘Baja 1000’ off-pavement vehicle races, and years of
exploitation of palmilla (Yucca schidigera Roezl ex Ortgies).
S. H. Bullock et al.
132 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 7
there were three settlements of > 100 inhabitants in our area
(Catavina, Punta Prieta and Rosarito).
Livestock were introduced more than 230 years ago (del
Barco, 1988, p. 304), but have remained extremely localized on
this arid landscape. After the establishment of the ejidos
(1967–79; Fig. 3), attempts at cattle ranching increased,
sustained by liberal subsidies from the federal government;
these in turn floated on the high price of petroleum. The rural
subsidy programmes did not fare well through the national
economic crises of the 1980s and 1990s. Goats have been
exceptionally rare in our area, although abundant and
devastating in the mountains south of Sta. Gertrudis. Feral
burros probably have been present but are not conspicuous
and their numbers have never been estimated. The lack of
surface water has limited the cattle population and restricted
most of its activity to moister arroyos and their immediate
vicinity, where impacts can be intense, but no careful study has
been made. Government range scientists classified the entire
region, except some major arroyos and the sierras SE of San
Borja, as unsuitable for any attempts at livestock ranching
(Paulın-Ramırez et al., 1981), but there have been many small,
marginal operations. In our area, the estimated number of
cattle in 1998 was < 7500 (Aramburo et al., 2002).
Census sites and methods
The sites for this study were chosen in an area relatively well
represented by old data, and that is relatively homogeneous in
climate and present and historical land use. Some potential
sites were excluded due to recent heavy disturbance, absence of
both cardon and cirio, poor quality of the old data or
uncertainties in the photointerpretation. In addition, an effort
was made to obtain a broad representation in terms of
geography and time.
The initial materials for this study were 107 old photographs
taken by 21 travellers and obtained from 15 archives,
individuals or publications (see Acknowledgments). The old
photographs dated from 1905 to 1967 such that the intervals
between censuses ranged from 29 to 95 years and were
> 55 years at half the sites (Fig. 3). At 14 sites, old photo-
graphs were repeated by J. R. Hastings and R. M. Turner in
1963, so that our work represented the third recording of those
sites (Fig. 2). In these cases, the earlier intervals were
25–40 years and the later interval was uniformly 33 years.
(Old documentary information is too vague for demography,
although occasionally useful for vegetation studies; Minnich &
Franco-Vizcaıno, 1998.)
The views were located in the field between 1996 and 2002.
Following standard methods (Rogers et al., 1984), the old
photographs were repeated from a camera position indistin-
guishable in the field from the old position. The old and new
photographs were printed to match. Censuses were then made
by detailed inspection, identifying and tabulating survivors,
disappearances (deaths) and appearances (recruits). In a few
cases where the number of individuals was small, the data were
combined from closely adjacent localities with similar substrate
and equivalent years of record. In addition, some photographs
were internally partitioned into two sites in order to separate
areas of highly contrasting substrates. The resulting 77 sites are
mapped in Fig. 2. We used the variation among sites to test for
factors affecting survivorship and recruitment, instead of
combining results among sites (Bowers et al., 1995).
We characterized several physical features of each site,
including latitude, elevation, distance from the Pacific coast
and precipitation. In order to approximate the variation of
precipitation, we grouped the sites into 10 physiographical
subareas, which could be characterized from the few weather
stations [nine reported by Bullock, 2003, plus Krutzio, on the
Pacific coast SSW of Punta Prieta (Adrian Aguirre Munoz,
pers. com.)]. Sites in eight subareas were assigned the mean
annual rainfall (1977+) of the included station. The ninth
subarea was assigned the mean of two adjacent stations, and
the 10th was given the mean of three stations.
We also developed several qualitative classifications of other
physical factors that might affect population dynamics:
landform (slope, pediment, plain, valley); geology (granite,
basalt, sedimentary, metasedimentary, mixed); topographic
protection from high winds (summit, flat, hills, valley); soil
texture (four classes); soil stability (eight classes); slope
gradient (four classes) and exposure (open, southerly, nor-
therly). Following the observations of McAuliffe (1991), we
compared vital rates between sites with old and new alluvial
soils. In addition, based on the observations of Franco-
Vizcaıno et al. (1993), we compared vital rates among the few
sites where detailed soil data were available.
Accessibility to people was described for each site in terms of
distances to road, ranch and town.
Demographic calculations
Survivorship was modelled by nonlinear regression with the
Weibull function
Nt ¼ N0 eð�t=BÞc
where N0 is 1000, t is time in years, and B and C are the fitted
constants. The observed number of survivors, of all sizes, at
each site was adjusted to an initial population of 1000 for this
analysis (Ebert, 1999); for either species, we excluded sites with
less than five individuals in the oldest photograph. The effects
of variables other than time were evaluated using the residuals
of survivorship (observed ) predicted from the Weibull
curves). This treatment differs substantially from Bowers et al.
(1995), who did not model a trend in survivorship but
calculated only cumulative mortality.
Recruitment was calculated as the ratio of new individuals to
the median between the initial number and the number of
survivors. Recruitment was modelled as a linear function of
time, and the residuals were used to test the effects of other
variables.
Residuals of the survivorship and recruitment regressions
were analysed with single factor analysis of variance or
regression, except the quantitative variables were also examined
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 133
Page 8
together in linear models. The qualitative variables presented
problems of unequal numbers of categories and non-uniform
distribution of cases among categories; parametric and nonpar-
ametric tests were applied, and for some analyses categories were
censored or combined. Sample sizes (number of sites) were
particularly small for the soil chemistry variables.
For these analyses, the sites with censuses from three
different years were represented by the data from the first and
last censuses only.
Census errors
Censuses of visible individuals are readily made for columnar
(and many other) species based on ground-based photogra-
phy. Nonetheless, it is important to consider the relative
accuracy of censuses from photogrammetric and standard
plots. Potential errors derive from problems of resolution due
to differences between camera lenses or lighting conditions at
the times of the original and repeat photographs. These
problems generally can be avoided by excluding doubtful parts
of the view in both photographs. Similarly, some parts of a
view may have to be excluded due to obscuring vegetation in
the old or new photographs. Other characteristics of repeat
photography are a variance in the intensity of sampling (in any
size of plot) because of geomorphic obstructions, a high
variance in ‘plot size’ among sites, and a strong bias against
inclusion of the smallest plants (particularly those < 30 cm
tall). The recruitment and mortality we report is of well-
established plants, probably more than 5 or 10 years old (pers.
obs.). Age is indeterminate, however. At least in the case of
cirio, studies of growth rates have shown that size is a very
poor indicator of age (Escoto-Rodrıguez & Bullock, 2002;
Ramırez Apud Lopez, 2002; Bullock et al., 2004).
In the case of cirio, the occasional occurrence of basal
branching may lead to overestimation of abundance, but no
consistent bias regarding vital rates. The geographical and
temporal variation of multiple trunks has not been assessed,
but basal branching has not been detected at all sites, and is not
common where it has been observed. Apparently, growth rates
of individual trunks do not differ appreciably between solitary
and multiple-trunk plants (Escoto-Rodrıguez, 1999).
RESULTS
Human impacts
For prehispanic times in the Vizcaıno region, destructive use of
live plants parts other than seeds and fruits, was only remarked
for Agave (Aschmann, 1959). In the mission decades, collecting
wood for fuel and construction may have had locally
significant effects. Cirio was used heavily for firewood at San
Borja although the missionaries considered it a remarkably
useless plant (del Barco, 1988, p. 93) and the Cochimı
considered it harmful (Linck, in Burrus, 1967, p. 46; see also
del Barco, loc. cit.). Cardon has long been used for its edible
fruits.
In the short-lived mining era, large amounts of wood were
required for structural uses and for fuel in steam-powered
stamp mills and smelters (Chaput et al., 1992), particularly in
the Calmallı district (Southworth, 1889, plates on pp. 87–88),
at San Fernando (Bose & Wittich, 1912–1913, plate 86), at
Julio Cesar (Bose & Wittich, 1912–1913, plate 80) and at San
Juan. Cirio and cardon often may have been spared the axe due
to their succulence, but as at mission San Borja, demand was
extreme and resulted in some notable examples of very local
depletion (Aschmann, 1972). The limitation of mine produc-
tion by scarcity of wood was occasionally remarked (e.g.
Engerrand & Paredes, 1912–1913) and is underscored by the
frequent practice of shipping raw ore to Europe for processing
(Chaput et al., 1992). The populous and long-lasting settle-
ment at El Marmol fuelled its kitchens with locally abundant
mesquite, although some propane may have been used since
the 1950s (P. Maclish, pers. com.).
The post-1960s increase of human population in the desert
brought greater demands for wood for fuel and construction,
more cattle, and more commerce in local products with distant
markets. Wood from cardon and cirio have been widely used,
the former principally to support roofing (Moran, 1968), and
the latter for corrals (Fig. 4) and various ornamental uses,
including wall coverings, for which cirio has had a market
outside the region. Extensive land uses probably have not had
obvious effects on cirio or cardon. Cattle ranching has
persisted in marginal operations since the early 1800s and
was heavily subsidized in the 1970s and 1980s, but impacts on
the vegetation are obvious only in some areas with permanent,
quality forage or water. The gathering of fruits of jojoba
(Simmondsia chinensis (Link) Schneid.), which lasted for a few
years in the 1970s and 1980s (Bullock, 1999), entailed well-
dispersed but selective and seasonal incursions. In addition,
harvesting of stems of palmilla (Yucca schidigera Roezl
ex Ortgies) in the Vizcaıno region, which dates from the 1970s,
is limited to scattered dense stands of a few thousand hectares
to the NW of Catavina (Carranza-Acevedo, 1997; Bullock,
1999); these stands lack sizeable cirio or cardon populations.
Vegetation has been destroyed incidental to road-building
and urbanization. Off-road vehicle use, which is known
elsewhere to be extremely damaging to desert soils and
vegetation (Webb & Wilshire, 1983), has been unrestricted
(and unstudied). However, all-terrain races have been run on
existing roads except, perhaps, for informal and/or poorly
documented events in the 1950s and 1960s (Edith Bernal,
SECTURE, unpubl. data; Thawley, 1968; Cortes, 1995).
Hurricanes
Baja California is occasionally affected by hurricanes that have
generated in low latitudes of the Eastern Pacific, as in
September 1959 and September 1997 (‘Nora’). Thorough data
on Eastern Pacific storms, including tracking and wind
intensity, is relatively recent (Court, 1980) so the frequency
and geography of impacts are not well known. Historical data
show our study area was crossed by 15 tropical storms or
S. H. Bullock et al.
134 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 9
hurricanes between 1920 and 2002 (Figs 3 & 5; Court, 1980;
Garcıa et al., 1980; http://weather.unisys.com/hurricane/
e_pacific). At least the 1959 and 1997 events traversed our
area at hurricane strength. Although the incursions between
1921 and 2002 give an average of 0.19 year)1, there were
periods of > 15 years without any identified incursion, but five
were reported in the 1920s (Garcıa et al., 1980). Of course,
hurricanes passing offshore or somewhat to the south of our
area also may have important impacts because the diameter of
the area of extreme winds may vary greatly (Hurd, 1929; Serra
Castelan, 1971).
Three surveys of Hurricane Nora’s impact were made in
November 1997. About 55 km ENE of Nora’s estimated
landfall (c. 93 km SSE of San Fernando) and 38 km north of
Punta Prieta, Clark & Ward (2000) observed 15% mortality in
cirio but none in cardon on a sandy 1.4 ha plot (n ¼ 31, 24,
respectively). In the same area, our own counts on more varied
soils along an 11.2 km transect showed c. 4.2% mortality in
cirio and 1.0% in cardon (considering plants > 1.5 m tall,
n ¼ 851, 796). We also made observations slightly further east
(c. 25 km NE of Punta Prieta) along a 12.6 km transect, and
found cirio was more affected than cardon (3.6% vs. 0.6%,
n ¼ 1452, 1470).
Photogrammetric plot data
We identified 3780 cirios and 2246 cardons at 77 sites, including
long-term survivors (2256 and 1508, respectively) and recruits
(404 and 491). Cirio occurred at more sites and was generally
more common than cardon. At the initial censuses (i.e. in the
older photographs), there were more than 50 individuals at
about one-third of the sites for cirio but at only c. 19%of the sites
for cardon. The maximum local sample sizes in the initial
censuses were 257 for cirio and 151 for cardon. There was a mild
positive relation between the abundances of the two species at
the chosen sites (Pearson’s r ¼ 0.339 in the old photos and 0.332
in the new, both P < 0.01; or, excluding four outliers, 0.500 and
0.514, P > 0.001).
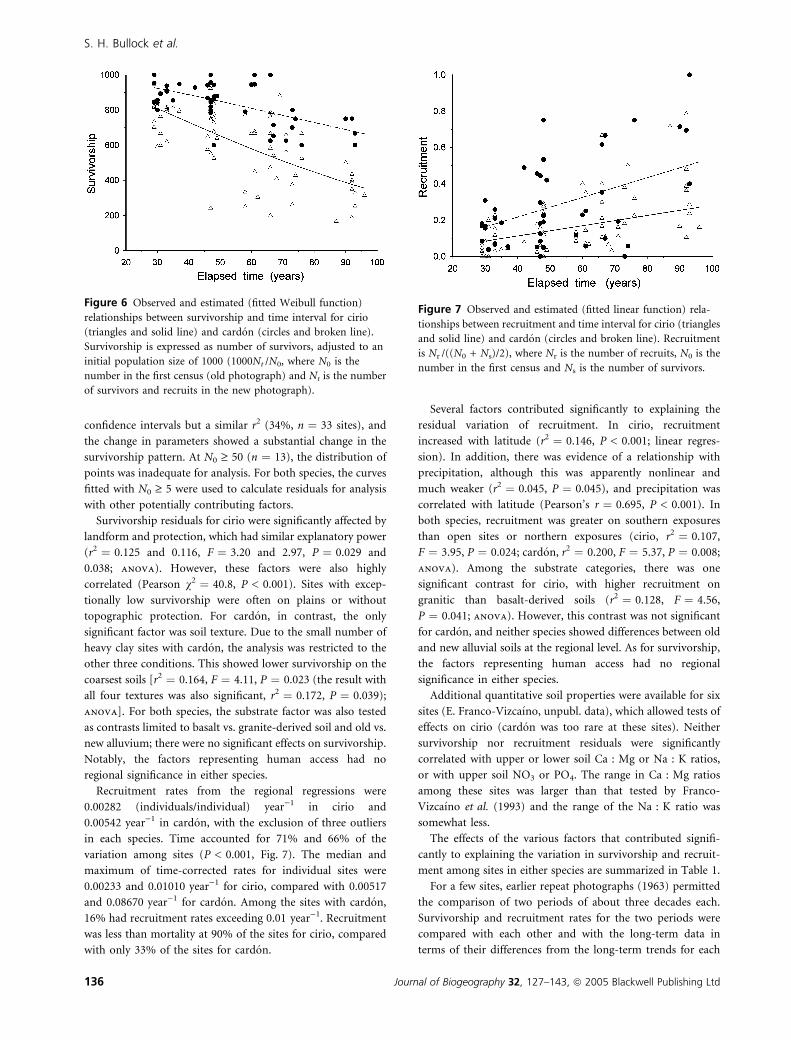
Survivorship was significantly related to the time interval
between photographs. The fitted Weibull curves of survivors
(adjusted to an initial N ¼ 1000) vs. time interval explained
45% of the variation for cirio and 35% for cardon for sites with
N0 ‡ 5 (both P < 0.001, n ¼ 72 and 48 sites, respectively;
Fig. 6). Because the curves are nonlinear, no single, overall
value of survivorship can be given. However, as an indication
of longevity, we calculated the estimated time until an all-size
cohort of 1000 would become extinct: 388 years for cirio and
723 years for cardon.
For cirio, by restricting the analyses to sites with ‡ 15 or
‡ 50 individuals (n ¼ 62 and 26), the explicative power of
elapsed time alone increased to 49% and 63% of the variance,
respectively. However, the confidence limits for the parameters
were very similar at all three N0, and were narrowest at N0 ‡ 5.
For cardon, limiting the analysis to N0 ‡ 15 gave poorerFigure 5 Recorded tracks of tropical storms and hurricanes that
crossed part of the study area, 1920–2002.
Figure 4 A 1996 view of a corral constructed of c. 113 cirio trunks, adjacent to ruins of the San Fernando mission; previous photographs of
this view showed no corral (1923) and a corral made entirely of mesquite (Prosopis sp.; 1963).
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 135
Page 10
confidence intervals but a similar r2 (34%, n ¼ 33 sites), and
the change in parameters showed a substantial change in the
survivorship pattern. At N0 ‡ 50 (n ¼ 13), the distribution of
points was inadequate for analysis. For both species, the curves
fitted with N0 ‡ 5 were used to calculate residuals for analysis
with other potentially contributing factors.
Survivorship residuals for cirio were significantly affected by
landform and protection, which had similar explanatory power
(r2 ¼ 0.125 and 0.116, F ¼ 3.20 and 2.97, P ¼ 0.029 and
0.038; anova). However, these factors were also highly
correlated (Pearson v2 ¼ 40.8, P < 0.001). Sites with excep-
tionally low survivorship were often on plains or without
topographic protection. For cardon, in contrast, the only
significant factor was soil texture. Due to the small number of
heavy clay sites with cardon, the analysis was restricted to the
other three conditions. This showed lower survivorship on the
coarsest soils [r2 ¼ 0.164, F ¼ 4.11, P ¼ 0.023 (the result with
all four textures was also significant, r2 ¼ 0.172, P ¼ 0.039);
anova]. For both species, the substrate factor was also tested
as contrasts limited to basalt vs. granite-derived soil and old vs.
new alluvium; there were no significant effects on survivorship.
Notably, the factors representing human access had no
regional significance in either species.
Recruitment rates from the regional regressions were
0.00282 (individuals/individual) year)1 in cirio and
0.00542 year)1 in cardon, with the exclusion of three outliers
in each species. Time accounted for 71% and 66% of the
variation among sites (P < 0.001, Fig. 7). The median and
maximum of time-corrected rates for individual sites were
0.00233 and 0.01010 year)1 for cirio, compared with 0.00517
and 0.08670 year)1 for cardon. Among the sites with cardon,
16% had recruitment rates exceeding 0.01 year)1. Recruitment
was less than mortality at 90% of the sites for cirio, compared
with only 33% of the sites for cardon.
Several factors contributed significantly to explaining the
residual variation of recruitment. In cirio, recruitment
increased with latitude (r2 ¼ 0.146, P < 0.001; linear regres-
sion). In addition, there was evidence of a relationship with
precipitation, although this was apparently nonlinear and
much weaker (r2 ¼ 0.045, P ¼ 0.045), and precipitation was
correlated with latitude (Pearson’s r ¼ 0.695, P < 0.001). In
both species, recruitment was greater on southern exposures
than open sites or northern exposures (cirio, r2 ¼ 0.107,
F ¼ 3.95, P ¼ 0.024; cardon, r2 ¼ 0.200, F ¼ 5.37, P ¼ 0.008;
anova). Among the substrate categories, there was one
significant contrast for cirio, with higher recruitment on
granitic than basalt-derived soils (r2 ¼ 0.128, F ¼ 4.56,
P ¼ 0.041; anova). However, this contrast was not significant
for cardon, and neither species showed differences between old
and new alluvial soils at the regional level. As for survivorship,
the factors representing human access had no regional
significance in either species.
Additional quantitative soil properties were available for six
sites (E. Franco-Vizcaıno, unpubl. data), which allowed tests of
effects on cirio (cardon was too rare at these sites). Neither
survivorship nor recruitment residuals were significantly
correlated with upper or lower soil Ca : Mg or Na : K ratios,
or with upper soil NO3 or PO4. The range in Ca : Mg ratios
among these sites was larger than that tested by Franco-
Vizcaıno et al. (1993) and the range of the Na : K ratio was
somewhat less.
The effects of the various factors that contributed signifi-
cantly to explaining the variation in survivorship and recruit-
ment among sites in either species are summarized in Table 1.
For a few sites, earlier repeat photographs (1963) permitted
the comparison of two periods of about three decades each.
Survivorship and recruitment rates for the two periods were
compared with each other and with the long-term data in
terms of their differences from the long-term trends for each
Figure 6 Observed and estimated (fitted Weibull function)
relationships between survivorship and time interval for cirio
(triangles and solid line) and cardon (circles and broken line).
Survivorship is expressed as number of survivors, adjusted to an
initial population size of 1000 (1000Nt /N0, where N0 is the
number in the first census (old photograph) and Nt is the number
of survivors and recruits in the new photograph).
Figure 7 Observed and estimated (fitted linear function) rela-
tionships between recruitment and time interval for cirio (triangles
and solid line) and cardon (circles and broken line). Recruitment
is Nr /((N0 + Ns)/2), where Nr is the number of recruits, N0 is the
number in the first census and Ns is the number of survivors.
S. H. Bullock et al.
136 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 11
species (n ¼ 12 sites for cirio, n ¼ 6 for cardon). Survivorship
and recruitment ‘residuals’ for the two periods were not closely
correlated in either species. By nonparametric comparison
with the long-term residuals, cardon showed no significant
differences in the overall variation of survivorship or recruit-
ment. Cirio showed no significant differences in recruitment,
but survivorship in both periods was exceptionally low
compared with the long-term, all-site trend (Mann–Whitney
test, P ¼ 0.032 for first period vs. long-term, P ¼ 0.024 for
second vs. long-term).
DISCUSSION
During the 1900s, mortality exceeded recruitment for cirio at
90% of sites in the region of its greatest abundance. This
species, a charismatic indicator of the Sonoran Desert in Baja
California, experienced a decline that was large in numbers,
geographically extensive and long-term. For cardon, on the
contrary, recruitment was higher than mortality at 67% of its
sites, indicating that the regional population of this species has
been either stable or slightly increasing, in accord with studies
in Sonora (Turner et al., 2003). Geographically extensive
explanations for the trends are as yet speculative and are
limited to ideas about the impacts of humans and the physical
environment, particularly climate. There has been no attention
to interactions with other organisms such as nurse plants, pests
or herbivores.
These trends are balances and may find explanation on the
side of survivorship or recruitment or both. These processes
can be examined separately because our method used identi-
fied individuals, not simply abundances. As expected, both
these processes were strongly time-dependent, to the extent of
explaining 45–63% of the variation in survivorship among sites
in cirio and 35% in cardon, as well as 71% and 66% for
recruitment. The residual variations were explained in as much
as 20% by some physical factor, but the factors differed
between species and between processes. Only site exposure
affected both species in regard to recruitment.
Time alone remains the major variable, which suggests merit
in further study of survivorship and recruitment, particularly
including estimates of age or stage (Pierson & Turner, 1998).
However, such estimates are difficult at best and subject to
large errors at worst (Bullock et al., 2004). The all-size cohorts
used for our survivorship curves may include plants differing
in age by tens or hundreds of years. This is probably the reason
that elapsed time explained much more variation for
recruitment than for survivorship. At the same time, further
study is needed of mortality factors, the conditions of
establishment, and of variations in the balance between these
processes in both species. Further study by other methods is
clearly needed to improve the representation of small juvenile
plants.
The net result of mortality and recruitment is depicted in
relation to elapsed time in Fig. 8. These results suggest a
tendency for cardon to increase and cirio to decrease. Lacking
any particular conceptual model, the trends were adequately
treated as linear, with an increase of 0.116% year)1 in cardon
and a much stronger and faster trend in cirio, decreasing at
0.446% year)1.
Survivorship
At the scale of one century, survivorship was best modelled as
decreasing with time in cirio, but models with constant or
decreasing survivorship were very similar in cardon. The
strength of the models is remarkable considering the disper-
Table 1 Significant explanatory factors for
variation among sites of survivorship and
recruitment in cirio and cardon Factors
Cirio Cardon
Survivorship Recruitment Survivorship Recruitment
Elapsed time (Ni > 5) r2 ¼ 0.454*** 0.708*** 0.347*** 0.664***
Latitude 0.146***
Landform 0.125*
Exposure (S vs. N) 0.107* 0.200**
Substrate (texture) 0.164*
Substrate (basalt vs. granite) 0.128*
*P < 0.05, **P < 0.01, ***P < 0.001.
Figure 8 Net change in cirio and cardon (recruitment ) mor-
tality), expressed as the percentage of the initially observed pop-
ulation at each site. For cirio (triangles), net change ¼ ) 0.446.
(elapsed time) (r2 ¼ 0.680, F ¼ 151.2, P > 0.001); for cardon
(circles), net change ¼ 0.116. (elapsed time) (r2 ¼ 0.178, F ¼ 9.8,
P < 0.001).
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 137
Page 12
sion and heterogeneity of the sites. Unfortunately, retrospec-
tive validation of the models cannot be extended significantly.
Our treatment of all sizes together, starting at an indefinite size
well beyond ‘seedling’, is a reasonable first approximation
because size apparently has a poor relation to age (Bullock
et al., 2004).
The models provide the first estimates of longevity in these
species and show a strong contrast between them. To estimate
local extinction time, the size of the initial cohort must be
specified; thus, for N0 ¼ 1000, extinction would be in less than
400 years for cirio but more than 700 years for cardon. The
times to halving of any N0 would be c. 71 and 140 years,
respectively. These estimates are not directly comparable with
other species for which long-term survivorship is not known
or has not been modelled numerically. Nonetheless, both cirio
and cardon are apparently much longer-lived than the large
saguaro cactus (Carnegiea gigantea (Englm.) Britton & Rose),
for which ages derived from growth rates suggest a ‘longevity’
of less than 200 years (Shreve, 1935; Turner et al., 2003), which
in turn is similar to the arborescent succulent kokerboom
(Aloe dichotoma Linn.f., Liliaceae) of south-western Africa
(Vogel, 1974), while falling short of the estimates of supra-
millenial longevity in gobernadora (McAuliffe, 1988).
Turner (1990) showed that mortality rates varied consider-
ably during the twentieth century for several large perennials in a
protected crater in the Pinacate region of Sonora, principally in
relation to drought. Those variations were important to long-
term demographic changes. In contrast, the saguaro population
at the same site did not show large variations in mortality, such
that its unstable age structure could be attributed to variations in
recruitment (Turner, 1990). However, lacking an analysis for
saguaro similar to that for cardon, we note again that time
accounted for only 35% of the variation in mortality of cardon,
so that the possibility of major dieback due to disease, for
example, remains of interest (Holguin et al., 1993).
Only two substrate variables contributed significantly to the
variation in survivorship, and these were not the same for cirio
and cardon. Cirio lost an exceptionally larger proportion of
individuals on sites offering no topographic protection from
high winds, while cardon mortality was higher on sites with
very coarse or very fine soils. At least in the context of our
design and data, several factors were not significant despite
suggestions by earlier authors, including the contrast of older
and newer alluvium, basalt and granite, some aspects of soil
chemistry, and slope exposition. McAuliffe (1991) found
contrasts in population structures of cirio and cardon between
alluvial surfaces of different ages at one site. The difference
with our negative result for surface age may be due to our
focus on observed population dynamics or problems with
McAuliffe’s assumptions about growth. However, our negative
result may be an artefact because our regional perspective
required that ‘new’ refer to very recent and ‘old’ include all
other surfaces, with a great variety of ages and histories. More
directly related to soil chemistry, our sparse data for effects
Ca : Mg, Na : K, NO3 and PO4, suggest no strong demogra-
phic effect that might correspond to the suggested effect on
succulent diversity (Franco-Vizcaıno et al., 1993). On the
contrary, the unexpected negative results for basalt vs. granite
corresponded to a negative result for soil texture in cirio. For
cardon, the parent rock contrast also was not significant, but
soil texture did explain some of the residual variation.
Like many columnar succulents, cirio and cardon can be
toppled by high winds. The impact of extreme north or east
winds has not been studied, but the passage of hurricanes over
the northern Vizcaıno is clearly a mortality factor of
importance (Clark & Ward, 2000; this study). The four to
six times greater impact of Hurricane Nora on cirio compared
with cardon may be largely a result of differences in the root
systems: cardon has both a near-surface, wide-spreading
system and a basal bayonet root (Niklas et al., 2002) but
cirio has only a shallow system with few major radials.
Although the long-term history of hurricane landfalls will
probably remain obscure, and many years may pass without a
landfall, the meteorology and inferred history of Eastern
Pacific hurricanes (Court, 1980) leaves no doubt of the
relevance of hurricanes to plant populations at the scale of
several decades and centuries.
Although cirio has been in demand for construction,
decoration and even fuel, observed mortality did not relate to
our indices of accessibility. Thus, we suggest that direct
exploitation has not had a simple or regionally significant
impact. Nonetheless, exploitation of the trunks is a locally
important factor even if the geography of impacts is complex.
Following the establishment of the ejidos in 1967–79, there
was undoubtedly increased use of cirio with the increases in
resident human population and cattle, although these remain
at low levels. No estimations of use have been attempted, but
cirio is still very common in homes, commercial establish-
ments and now-deteriorating corrals (Fig. 4). Moreover, just
as the ejidos flourished with the coming of the paved
highway, so did tourism and the consequent demand for cirio
wood for decoration, construction and export. Cardon ‘ribs’
are often used in construction, as small beams and posts in
roofs, walls and fencing, but apparently have no export
market.
With respect to the exploitation of wood of both species,
however, local people claim to use only naturally dead and
dried plants because unnatural death results in rotting; this has
not been independently verified. Because of its apparent
general decline, cirio populations should be protected from
excessive felling for local use or export.
Six of the sites were very near to missions, but none of these
showed an exceptional survivorship in terms of residuals of the
general time–survivorship relationship for cirio and cardon.
Photographs were also repeated at the San Fernando and Julio
Cesar copper mines, showing substantial increases in cover by
some shrubs, but the numbers of cirios and cardons distin-
guishable in the older photographs did not meet our minimum
criterion to include among the censuses. Recovery of cirio
around the San Fernando mine might require 200 years,
according to Aschmann’s (1972) interpretation of an informal
survey.
S. H. Bullock et al.
138 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 13
Recruitment
Elapsed time explained c. 70% of the variation in recruitment
in both cirio and cardon. For several reasons, there is no
necessary implication that establishment was spatially or
temporally uniform. Of course, in our methods recruitment
is not seedling establishment but appearance, which integrates
establishment over longer periods. The strength of the
relationship was undoubtedly enhanced by the long intervals
between censuses and by the interval of 29 years shared by all
sites (i.e. 1967–1996). In addition, excluding a few extreme
outliers in each species greatly improved the relationships.
Surprisingly, the median annual rate of recruitment was more
than twice as high in cardon (0.5%, compared with 0.2% in
cirio), and the maximum rate was more than eight times
higher in cardon (8.7% vs. 1.0%).
Some of the residual variation in cirio was explained by
latitude. This may be partly an effect of precipitation, but the
analysis suggested that some other gradients must also be
involved. Despite this positive relationship with latitude, cirio
also showed better recruitment on southerly exposures, which
was also true for cardon. A similar trend for exposure has been
noted for large cacti in the northern part of their distribution
(Parker, 1987,1993; Pierson & Turner, 1998). Temperature
regimes in the Vizcaıno region have not been studied
sufficiently to determine if the exposure effect could be
attributed to killing frosts. Of course, field experiments on
establishment would be particularly useful if active manage-
ment becomes a priority. Factors other than climate that merit
attention are nurse plant populations (McAuliffe, 1988) and the
relation of individual growth rates to demographic processes.
Cirio establishment is supposed to be favoured by wet
winters followed by cool summers (Humphrey, 1974). In the
region of the sites reported here, the history of winter rainfall is
spatially more uniform than summer rainfall (Bullock, 2003).
Thus, establishment should tend to be regionally synchronized.
However, opportunities for establishment may have been very
limited in the last 100 years. In the period of climatic records
(< 50 years), only c. 1 year in 10 had a cool season with 3 or
even 2 months of > 50 mm precipitation. A proxy-based
reconstruction for nearly 150 years (Bullock, 2003) suggests
opportunities for establishment have been not only infrequent
but also very unevenly distributed in time, as nearly half of the
exceptionally wet winters occurred after 1975. On the basis of
size structure of one population, Aschmann (1972) suggested
there were two or three recruitment events from c. 1900 to
c. 1950.
The long-term variation in precipitation suggests that cirio
may have had increased establishment in the late 1900s. These
plants would not be apparent in the repeat photographs due to
their diminutive size and the disguising of cirio’s conical trunk
by an abundance of lateral branches in the ‘juvenile’ stage. The
existence of such cohort awaits an independent test.
In contrast to cirio, cardon establishment may be favoured
by wet conditions in summer and fall, resulting from
incursions of tropical air masses, perhaps followed by mild
winters. The evidence in favour of this idea is that seed
dispersal occurs in summer (Moran, 1968), seed germination is
lower at temperatures below 40 �C (Cancino et al., 1993), and
the species is more widely distributed in the southern portion
of the peninsula where precipitation occurs predominantly in
summer (Hastings & Turner, 1965; Turner et al., 1995).
Appreciable summer precipitation is uncommon at any place
in the northern Vizcaıno region, and most rainfalls are local, as
shown by the poor spatial–temporal coherence of summer
precipitation records (Bullock, 2003). The corresponding
pattern of establishment for cardon should be sporadic events
that are asynchronous among localities. This is apparent to
some extent in the outlying values of recruitment, reaching
more than 15 times the median, and perhaps in the
diversity among sites of population increases and decreases.
Nonetheless, it is remarkable that, overall, greater recruitment
and less mortality has occurred in the species which is more
responsive to summer rains.
In cardon, both the diversity among sites and the overall
majority of increasing populations is remarkably similar to
results from its coastal populations in Sonora (Turner et al.,
2003). Thus, cardon contrasts with saguaro, the other giant
columnar cactus of the Sonoran Desert, which appears to be in
a widespread, long-term decline (Turner et al., 2003). The
natural history of saguaro bears little resemblance to cirio, so
the causes of their declines probably differ. The potential
difficulty in understanding cirio and cardon is suggested by the
history and variety of hypotheses regarding changes in saguaro
(e.g. Niering et al., 1963), most recently including global
climate change (Thompson et al., 1998). Compared with
declines of arborescent succulents in south-western Africa
(Midgley et al., 1997), the decline of cirio is apparently on a
different time scale and is not attributable to obvious non-
climatic factors.
Populations of desert perennials may be typically unstable
(Turner, 1990; Van Devender, 1990; McAuliffe, 1994), and the
patterns and causes of their changes may be heterogeneous
among species at any site or among sites for any species
(Parker, 1987; Turner, 1990; McAuliffe, 1994; Nabhan &
Suzan, 1994; El-Keblawy et al., 1997; Turner et al., 2003). Cirio
and cardon show very different overall patterns at the same
sites across a substantial region. The decades-long decline of
cirio may be a consequence of regional pulses in establishment
during the past 400 years, with establishment in the twentieth
century failing to match the mortality of earlier cohorts. This is
analogous to interpretations of the fluctuations of other
perennials of various life forms, in more continental but
biogeographically related regions (Jordan & Nobel, 1981;
Goldberg & Turner, 1986; Parker, 1993; Turner, 1990; Bowers
et al., 1995; Pierson & Turner, 1998; Turner et al., 2003). As
with cirio and cardon, many of these species have shown net
changes that are similar although numerically disparate
between the early and late twentieth century (Bowers et al.,
1995; Turner et al., 2003). Episodic establishment and con-
tinuous mortality could provide a conceptual framework for
the population dynamics of cirio and cardon, but we cannot
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 139
Page 14
yet rule out other combinations of continuous and episodic
processes (Watson et al., 1997). In addition, a regionally
coherent hypothesis like direct effects of climatic variability
ought to be subject to other tests (Jordan & Nobel, 1981;
El-Keblawy et al., 1997; Watson et al., 1997; Pierson & Turner,
1998), and need not apply at any particular site, considering
other idiosyncratic features.
Our methods permit contrasts of overall population trends,
geographical variation and factors affecting survivorship and
recruitment, based on known individuals at many sites over a
large region, observed at intervals of several decades to a
century. This study provides the first synchronic view of
vegetation processes in the coastal, subtropical desert of the
Vizcaıno region, showing time scales that contrast between two
dominant plant species and with shorter-lived dominants of
other regions. The apparent paucity of human impacts is
remarkable, but much variation remains to be explained in
terms other than our characterizations of the populations and
their physical environment.
ACKNOWLEDGEMENTS
We are grateful to the individuals whose photographs have
become a legacy of growing importance: C.G. Abbott, B.A.
Arnold, H. Aschmann, E. Bose, H. Bravo-Hollis, A. Bridge,
H.W. Crosby, N.H. Darton, J. Engerrand, E.A. Goldman, H.E.
Gulick, J.R. Hastings, L.M. Huey, G.E. Lindsay, T.D. Mallery,
M. McDonald, T. Paredes, E.S. Ross, F. Shreve, R.M. Turner,
and I.L. Wiggins. We also thank several individuals as well as
institutions and their personnel for facilitating the use of old
photographs: Brigham Arnold, Louise Aschmann, Helia Bravo-
Hollis, Arthur Bridge and Edward Ross; US Geological Survey
Desert Laboratory, Smithsonian Institution Archives, Special
Collections of the University of Arizona Libraries, Mandeville
Special Collections of the University of California, San Diego,
US Geological Survey Photographic Library (Denver), Library
of the San Diego Natural History Museum and the US
National Archives.
This project was funded by CICESE, the National Geo-
graphic Society (5576-95) and USGS, and was assisted by
sabbatical support from CONACYT (990293) and San Diego
State University. We are grateful for comments on the
manuscript by Paul Zedler, Paul Dayton, Kathleen Parker
and anonymous reviewers, and for technical support by Mario
Salazar, and by Alejandro Hinojosa, Cesar Almeda, Hugo
Riemann, Fernando Escoto and Sean Connin. Loans of
equipment and facilities were kindly made by USGS-Tucson,
Glenn Northcutt, Sergio Ramos, CICESE’s Department of
Optics, and L.G. Alvarez. Adrian Aguirre kindly provided
precipitation data from Krutzio. We thank the people of the
region for their tolerance and help.
REFERENCES
Aramburo, G., Espejel, I., Leyva, C. & Martınez, R. (2002)
Programa de manejo del area de proteccion de flora y fauna
valle de los Cirios, Baja California. Informe Tecnico Final.
Comision Nacional de Areas Protegidas, Secretarıa de Eco-
logıa Medio Ambiente y Recursos Naturales, Mexico.
Arnold, B.A. (1957) Late Pleistocene and recent changes in
land forms, climate and archaeology in Central Baja Cali-
fornia. University of California Publications in Geography, 10,
201–318.
Aschmann, H. (1959) The Central Desert of Baja California:
demography and ecology. Ibero-americana, 42, 1–315.
Aschmann, H. (1972) Recovery of desert vegetation. Interna-
tional geography, 1972, Vol. 1 (ed. by W.P. Adams and F.M.
Helleiner), pp. 631–633. University of Toronto Press, Tor-
onto.
Bahre, C.J. & Hutchinson, C.F. (1985) The impact of historical
fuelwood cutting on semidesert woodlands of southeastern
Arizona. Journal of Forest History, 29, 175–186.
Bahre, C.J. & Shelton, M.L. (1993) Historic vegetation change,
mesquite increases, and climate in southeastern Arizona.
Journal of Biogeography, 20, 489–504.
Bose, E. & Wittich, E. (1912–13) Informe relativo a la
exploracion de la region Norte de la costa occidental de la
Baja California. Parergones del Instituto Geologico de Mexico,
4, 307–533 (plates LXXXVIII–CXII).
Bowers, J.E. (1997) Demographic patterns of Ferocactus cyl-
indraceus in relation to substrate age and grazing history.
Plant Ecology, 133, 37–48.
Bowers, J.E., Webb, R.H. & Rondeau, R.J. (1995) Longevity,
recruitment and mortality of desert plants in Grand
Canyon, Arizona, USA. Journal of Vegetation Science, 6,
551–564.
Brown, J.W., Real, H.G. & Faulkner, D.K. (1992) Butterflies of
Baja California: faunal survey, natural history, conservation
biology. Lepidoptera Research Foundation, Beverly Hills, CA.
Brum, G.D. (1973) Ecology of the saguaro (Carnegiea gigan-
tea): phenology and establishment in marginal populations.
Madrono, 22, 195–204.
Bullock, S.H. (1999) La vegetacion del noroeste de Baja Cali-
fornia en el contexto de la inestabilidad ambiental. Revista
Chilena de Historia Natural, 72, 501–516.
Bullock, S.H. (2003) Seasonality, spatial coherence and history
of precipitation in a desert region of the Baja California
peninsula. Journal of Arid Environments, 53, 169–182.
Bullock, S.H., Turner, R.M., Hastings, J.R., Escoto-Rodrıguez,
M., Ramırez Apud Lopez, Z. & Rodrıguez-Navarro, J.L.
(2004) Variance of size-age curves: bootstrapping with
covariance. Ecology, 85, 2114–2117.
Burrus, E.J. (1967) Wenceslaus Linck’s reports and letters 1762–
1778. Dawson’s Book Shop, Los Angeles, CA.
Cancino, J., Leon de la Luz, J.L., Coria, R. & Romero, H.
(1993) Effect of heat treatment on germination of seeds of
cardon [Pachycereus pringlei (S. Wats.) Britt. & Rose, Cac-
taceae]. Journal of the Arizona-Nevada Academy of Science,
27, 47–54.
Carlquist, S. (2001) Wood anatomy of Fouquieriaceae in
relation to habit, ecology, and systematics; nature of meri-
stems in wood and bark. Aliso, 19, 137–163.
S. H. Bullock et al.
140 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 15
Carranza-Acevedo, R.A. (1997) Caracterizacion ecologico-silvi-
cola de poblaciones de Yucca schidigera con potencial de
aprovechamiento. Master’s Thesis, Facultad de Ciencias,
Universidad Autonoma de Baja California, Ensenada.
Chaput, D., Mason, W.H. & Loperena, D.Z. (1992) Modest
fortunes: mining in northern Baja California. Natural History
Museum of Los Angeles County, Los Angeles.
Clark, W.H. & Ward, D.M. Jr (2000) Hurricane impacts in the
Central Desert of Baja California Norte, Mexico. Haseltonia,
7, 81–85.
Cortes, J.A. (1995) Bajo el desafio Baja 1000. Reforma,
November, 29, 1995, 3.
Court, A. (1980) Tropical cyclone effects on California. NOAA
Technical Memorandum NWS WR, 159, 1–45.
Davis, L.G. (2003) Geoarchaeology and geochronology of
pluvial Lake Chapala, Baja California, Mexico. Geoarchae-
ology, 18, 205–223.
Deasy, G.F. & Gerhard, P. (1944) Settlements in Baja Cali-
fornia: 1768–1930. Geographical Review, 34, 574–586.
del Barco, M. (1988) Historia natural y cronica de la antigua
California [adiciones y correciones a la noticia de Miguel
Venegas] (ed. by M. Leon-Portilla), 2nd edn. Universidad
Nacional Autonoma de Mexico, Mexico.
DETENAL (1982) Cartas Edafologicas 1 : 250000. Direccion
General de Geografıa del Territorio Nacional, Mexico.
Ebert, T.A. (1999) Plant and animal populations: methods in
demography. Academic Press, San Diego, CA.
El-Keblawy, A., Shaltout, K.H., Lovett-Doust, J. & Ramadan, A.
(1997) Population dynamics of an Egyptian desert shrub,
Thymelaeahirsuta.CanadianJournalofBotany,75,2027–2037.
Engerrand, J. & Paredes, T. (1912–13) Informe relativo a la
parte occidental de la region Norte de la Baja California.
Parergones del Instituto Geologico de Mexico, 4, 278–306
(plates LXVI–LXXXVII).
Escoto-Rodrıguez, M. (1999) Variaciones temporales y espaci-
ales en la tasa de crecimiento del cirio (Fouquieria col-
umnaris). Master’s Thesis, Facultad de Ciencias,
Universidad Autonoma de Baja California, Ensenada.
Escoto-Rodrıguez, M. & Bullock, S.H. (2002) Long-term
growth rates of cirio (Fouquieria columnaris), a giant suc-
culent of the Sonoran Desert in Baja California. Journal of
Arid Environments, 50, 593–611.
Espinoza Arroyo, A. (1992) Los Rosarenos: memorias del naci-
miento y vida de un pueblo bajacaliforniano, 1774–1992.
Sector Pesquero de El Rosario & Museo de Historia de
Ensenada, Ensenada.
Ferrusquıa-Villafranca, I. & Torres-Roldan, V. (1980) El
registro de mamıferos terrestres del Mesozoico y Cenozoico
de Baja California. Universidad Autonoma de Mexico, In-
stituto de Geologıa, Revista, 4, 56–62.
Fleming, T.H., Maurice, S., Buchmann, S.L. & Tuttle, M.D.
(1994) Reproductive biology and relative male and female
fitness in a trioecious cactus, Pachycereus pringlei (Cacta-
ceae). American Journal of Botany, 81, 854–867.
Fleming, T.H., Maurice, S. & Hamrick, J.L. (1998) Geographic
variation in the breeding system and the evolutionary sta-
bility of trioecy in Pachycereus pringlei (Cactaceae). Evolu-
tionary Ecology, 12, 279–289.
Franco-Vizcaıno, E. (1994) Water regime in soils and plants
along an aridity gradient in central Baja California, Mexico.
Journal of Arid Environments, 27, 309–323.
Franco-Vizcaıno, E., Goldstein, G. & Ting, I.P. (1990) Com-
parative gas exchange of leaves and bark in three stem
succulents of Baja California. American Journal of Botany,
77, 1272–1278.
Franco-Vizcaıno, E., Graham, R.C. & Alexander, E.B. (1993)
Plant species diversity and chemical properties of soils in the
central desert of Baja California, Mexico. Soil Science, 155,
406–416.
Fullola, J.M., Petit, M.A., Rubio, A., del Castillo, V. & Bergada,
M.M. (1993) Esquema crono-cultural del poblamiento
prehistorico de las sierras centrales de la penınsula de Baja
California, Mexico. Arqueologıa, 9/10, 3–15.
Gabb, W.M. (1867) Exploration of Lower California.
Resources of the Pacific Slope (ed. by J.R. Browne) [In
appendix A sketch of the settlement and exploration
of Lower California], pp. 82–112. D. Appleton & Co,
New York.
Garcıa, E. & Mosino, P. (1968) Los climas de la Baja Cali-
fornia. Comite Mexicano para el Decenio Hidrologico
Internacional. Memoria 1966–67, pp. 29–56. Instituto de
Geofısica, Universidad Nacional Autonoma de Mexico,
Mexico.
Garcıa, J.A., Castelan, S.S. & Betancourt, A.E. (1980) Trayec-
torias de ciclones tropicales. Centro de Ciencias de la
Atmosfera, Mexico. Universidad Nacional Autonoma de
Mexico, Mexico.
Gastil, R.G., Phillips, R.P. & Allison, E.C. (1975)
Reconnaissance geology of the state of Baja California.
Geological Society of America Memoir, 140, 1–170.
Giorgi, F., Shields Brodeur, C. & Bates, G.T. (1994) Regional
climate change scenarios over the United States produced
with a nested regional climate model. Journal of Climate, 7,
375–399.
Goldberg, D.E. & Turner, R.M. (1986) Vegetation change and
plant demography in permanent plots in the Sonoran
Desert. Ecology, 67, 695–712.
Graham, R.C. & Franco-Vizcaıno, E. (1992) Soils on igneous
and metavolcanic rocks in the Sonoran Desert of Baja
California. Geoderma, 54, 1–21.
Hastings, J.R. & Turner, R.M. (1965) Seasonal precipitation
regimes in Baja California, Mexico. Geografiska Annaler,
47A, 204–222.
Henrickson, J. (1975) Vegetative morphology of the cirio
(Fouquieria columnaris). Cactus and Succulent Journal,
(Suppl.), 33–46.
Holguin, G., Bowers, R. & Bashan, Y. (1993) The degeneration
of cardon populations in Baja California Sur, Mexico.
Cactus and Succulent Journal, 65, 64–67.
Humphrey, R.R. (1974) The boojum and its home: Idria col-
umnaris Kellogg and its ecological niche. University of Ari-
zona Press, Tucson, AZ.
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 141
Page 16
Humphrey, R.R. (1987) 90 years and 535 miles: vegetation
changes along the Mexican border. University of New Mexico
Press, Albuquerque.
Humphrey, R.R. (1991) Montevideo Valley and its tallest
recorded cirio. Cactus and Succulent Journal, 63, 239–
240.
Humphrey, R.R. & Humphrey, A.B. (1990) Idria columnaris:
age as determined by growth rate. Desert Plants, 10,
51–54.
Hurd, W.E. (1929) Tropical cyclones of the eastern North
Pacific Ocean. Monthly Weather Review, 57, 43–49.
INEGI (2001) Sıntesis de informacion geografica del estado de
Baja California. Instituto Nacional de Estadıstica, Geografıa
e Informatica, Aguascalientes.
INE-SEMARNAP (1997) Programa de conservacion de la vida
silvestre y diversificacion productiva en el sector rural 1997–
2000. Instituto Nacional de Ecologıa, Secretarıa del Medio
Ambiente, Recursos Naturales y de Pesca, Mexico.
Jordan, P.W. & Nobel, P.S. (1981) Seedling establishment of
Ferocactus acanthodes in relation to drought. Ecology, 62,
901–906.
McAuliffe, J.R. (1988) Markovian dynamics of simple and
complex desert plant communities. American Naturalist,
131, 459–490.
McAuliffe, J.R. (1991) Demographic shifts and plant succes-
sion along a late Holocene soil chronosequence in the
Sonoran Desert of Baja California. Journal of Arid Environ-
ments, 20, 165–178.
McAuliffe, J.R. (1994) Landscape evolution, soil formation,
and ecological patterns and processes in Sonoran Desert
bajadas. Ecological Monographs, 64, 111–148.
Markham, C.G. (1972) Baja California’s climate. Weatherwise,
25, 64–76.
Martınez, P.L. (1965) Guia familiar de Baja California 1700–
1900. Editorial Baja California, Mexico.
Meko, D., Stockton, C.W. & Boggess, W.R. (1995) The tree-
ring record of severe sustained drought. Water Resources
Bulletin, 31, 789–801.
Midgley, J.J., Cowling, R.M., Hendricks, H., Desmet, P.G.,
Esler, K. & Rundel, P. (1997) Population ecology of tree
succulents (Aloe and Pachypodium) in the arid western
Cape: decline of keystone species. Biodiversity and Con-
servation, 6, 869–876.
Minnich, R.A. & Franco-Vizcaıno, E. (1998) Land of chamise
and pines: historical accounts and current status of northern
Baja California’s vegetation. University of California Publi-
cations in Botany, 80, 1–166.
Moran, R. (1962) Visitors to the flowers of Pachycereus
pringlei. National Cactus and Succulent Journal, 17, 21.
Moran, R. (1968) Cardon. Pacific Discovery, 21, 2–9.
Moriarty, J.R. (1968) Climatologic, ecologic and temporal
inferences from radiocarbon dates on archaeological sites,
Baja California, Mexico. Pacific Coast Archaeological Society
Quarterly, 4, 11–38.
Nabhan, G.P., Donovan, J., Buchmann, S.L. & Burquez, A.
(1999) Riqueza y estabilidad de los polinizadores de los
cirios (Fouquieria columnaris) del desierto sonorense: un
analisis comparativo. Boletın de la Sociedad Botanica de
Mexico, 65, 39–50.
Nabhan, G.P. & Suzan, H. (1994) Boundary effects on en-
dangered cacti and their nurse plants in and near a Sonoran
Desert biosphere reserve. Ironwood: an ecological and cul-
tural keystone of the Sonoran Desert (ed. by G.P. Nabhan and
J.L. Carr), pp. 55–67. Conservation International, Wash-
ington, DC.
Niering, W., Whittaker, R.H. & Lowe, C.H. (1963) The
saguaro: a population in relation to its environment. Science,
142, 15–23.
Niklas, K.J., Molina-Freaner, F. & Tinoco-Ojanguren, C.
(1999) Biomechanics of the columnar cactus Pachycereus
pringlei. American Journal of Botany, 86, 767–775.
Niklas, K.J., Molina-Freaner, F., Tinoco-Ojanguren, C. &
Paolillo, D.J., (2002) The biomechanics of Pachycereus
pringlei root systems. American Journal of Botany, 89, 12–21.
Nilsen, E.T., Sharifi, M.R., Rundel, P.W., Forseth, I.N. &
Ehleringer, J.R. (1990) Water relations of stem succu-
lent trees in north-central Baja California. Oecologia, 82,
299–303.
Parker, K.C. (1987) Site-related demographic patterns of organ
pipe cactus populations in southern Arizona. Bulletin of the
Torrey Botanical Club, 114, 149–155.
Parker, K.C. (1988) Growth rates of Stenocereus thurberi and
Lophocereus schottii in southern Arizona. Botanical Gazette,
149, 335–346.
Parker, K.C. (1993) Climatic effects on regeneration trends for
2 columnar cacti in the northern Sonoran Desert. Annals of
the Association of American Geographers, 83, 452–474.
Paulın-Ramırez, O., Navarro-Cordova, A., Morales-Guiza,
V.F., Rosinol-Monges, J.R., Preciado-Torres, A., Aguirre-
Wallace, C.A., Peralta-Castro, G. & Camacho-Camacho, J.A.
(1981) La determinacion regional de los coeficientes de agos-
tadero para el estado de Baja California. Secretaria de Agri-
cultura y Recursos Hidraulicos, Ensenada.
Penalba, M.C. & Van Devender, T.R. (1998) Cambios de
vegetacion y clima en Baja California, Mexico, durante los
ultimos 20,000 anos. Geologıa del Noroeste, 2, 21–23.
Pierson, E.A. & Turner, R.M. (1998) An 85-year study of
saguaro (Carnegiea gigantea) demography. Ecology, 79,
2676–2693.
Ramırez Apud Lopez, Z. (2002) Modelos de crecimiento para la
estimacion de la edad con aplicacion en el cirio (Fouquieria
columnaris). Master’s Thesis, Facultad de Ciencias, Univer-
sidad Autonoma de Baja California, Ensenada.
Rhode, D. (2002) Early Holocene juniper woodland and cha-
parral taxa in the central Baja California peninsula, Mexico.
Quaternary Research, 57, 102–108.
Rogers, G.F., Malde, H.E. & Turner, R.M. (1984) Bibliography
of repeat photography for evaluating landscape change.
University of Utah Press, Salt Lake City, UT.
Sankey, J.T., Van Devender, T.R. & Clark, W.H. (2001) Late
Holocene plants, Catavina, Baja California, Mexico. South-
western Naturalist, 46, 1–7.
S. H. Bullock et al.
142 Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd
Page 17
Schmidt, R.H. Jr (1989) The arid zones of Mexico: Climatic
extremes and conceptualization of the Sonoran Desert.
Journal of Arid Environments, 16, 241–256.
Serra Castelan, S. (1971) Hurricanes and tropical storms of the
west coast of Mexico. Monthly Weather Review, 99, 302–308.
Shreve, F. (1910) Rate of establishment of the giant cactus.
Plant World, 13, 235–240.
Shreve, F. (1935) The longevity of cacti. Cactus and Succulent
Journal, 7, 66–68.
Shreve, F. (1936) The transition from desert to chaparral in
Baja California. Madrono, 3, 257–264.
Shreve, F. (1942) The deserts of North America. Botanical
Review, 8, 195–246.
Shreve, F. (1964) Vegetation of the Sonoran Desert. Vegetation
and flora of the Sonoran Desert (ed. by F. Shreve and I.L.
Wiggins), pp. 9–186. Stanford University Press, Stanford, CA.
Southworth, J.R. (1889) El territorıo de la Baja California
Mexico ilustrada: Su agricultura, comercio, minerıa e indus-
trias en Ingles y Espanol. Reprint, 1989, Government of Baja
California Sur, La Paz.
Steenbergh, W.F. & Lowe, C.H. (1983) Ecology of the saguaro:
III. Growth and demography. National Park Service Scientific
Monograph Series 17, Washington, DC.
Thawley, J. (1968) Big bad Baja: the first annual Mexican 1000
rally. Hot Rod, 21, 46–47.
Thompson, R.S., Hostetler, S.W., Bartlein, P.J. & Anderson,
K.H. (1998) A strategy for assessing potential future changes
in climate, hydrology, and vegetation in the western United
States. US Geological Survey Circular, 1153, 1–20.
Tschirley, F.H. & Wagle, R.F. (1964) Growth rate and
population dynamics of jumping cholla (Opuntia fulgida
Engelm.). Journal of the Arizona Academy of Science, 3,
67–71.
Turner, R.M. (1990) Long-term vegetation change at a fully
protected Sonoran desert site. Ecology, 71, 464–477.
Turner, R.M., Bowers, J.E. & Burgess, T.L. (1995) Sonoran
Desert plants: an ecological atlas. University of Arizona Press,
Tucson, AZ.
Turner, R.M., Webb, R.H., Bowers, J.E. & Hastings, J.R. (2003)
The changing mile revisited. University of Arizona Press,
Tucson, AZ.
Van Devender, T.R. (1990) Late Quaternary vegetation and
climate of the Sonoran Desert, United States and Mexico.
Packrat middens: the last 40,000 years of biotic change (ed. by
J.L. Betancourt, T.R. Van Devender and P.S. Martin), pp.
134–163. University of Arizona Press, Tucson, AZ.
Vogel, J.C. (1974) The life span of the kokerboom. Aloe, 12,
66–68.
Watson, I.W., Westoby, M. & Holm, A.M. (1997) Continuous
and episodic components of demographic change in arid
zone shrubs: Models of two Eremophila species from Wes-
tern Australia compared with published data on other spe-
cies. Journal of Ecology, 85, 833–846.
Webb, R.H. & Betancourt, J.L. (1992) Climatic variability and
flood frequency of the Santa Cruz River, Pima County,
Arizona. US Geological Survey Water Supply Paper, 2379,
40 pp.
Webb, R.H. & Wilshire, H.G. (eds) (1983) Environmental ef-
fects of off-road vehicles. Springer-Verlag, New York.
Wells, P.V. (2000) Pleistocene macrofossil records of four-
needled Pinyon or Juniper encinal in the northern Vizcaıno
desert, Baja California del Norte. Madrono, 47, 189–194.
BIOSKETCHES
Stephen Bullock studies plant demography, phenology and
reproductive biology in arid and semi-arid ecosystems,
particularly on the Pacific slope of Mexico.
Nora Martijena is interested in relating population and
community dynamics of forests and arid scrublands to biotic
and abiotic factors, using survey, experimental and dendro-
chronological techniques.
Robert Webb studies landscape change in the Sonoran,
Mojave and Great Basin Deserts, emphasizing historical
changes in upland vegetation, vegetation recovery and the
effects of landscape changes on fluvial systems.
Raymond Turner works on phytogeography, plant ecology
and ecosystem management; his early interest in vegetation
dynamics prompted establishment of long-term vegetation
study plots as well as the use of repeat photography for docu-
menting landscape change in the Sonoran Desert and Kenya.
Demographic changes in cirio and cardon
Journal of Biogeography 32, 127–143, ª 2005 Blackwell Publishing Ltd 143