ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 226, No. 1, October 1, pp. 190-197, 1933 Uricase from Soybean Root Nodules: Purification, Properties, and Comparison with the Enzyme from Cowpea’ KAREN LUCAS, MICHAEL J. BOLAND, AND KAREL R. SCHUBERT Department of Biochemistry, Michisan State University, East Lansing, Michigan 48824 Received February 24.1933, and in revised form May 13,1933 A 45-fold purification of uricase (urate:08 oxidoreductase, EC 1.7.3.3) from soybean root nodules by ammonium sulfate fractionation, gel filtration, and affinity chroma- tography is described. Electrophoresis on nondenaturing gels using an activity stain or on sodium dodecyl sulfate (SDS) gels demonstrated that the enzyme obtained was nearly homogeneous. The subunit molecular weight of uricase estimated from SDS gels was 32,000 -t 3000. Gel-filtration studies indicated that the native enzyme is a monomer at pH 7.5 which associates to form a dimer at pH 8.8. Enzyme activity was stabilized by the addition of dithiothreitol. The pH dependence of the enzyme showed an optimum of 9.5. Initial rate kinetics showed Km values of 10 and 31 PM for uric acid and oxygen, respectively, with an intersecting pattern of substrate dependence. Uricase activity was inhibited strongly by xanthine, which was competitive with respect to uric acid (Ki = 10 PM). No significant inhibition was observed in the presence of a variety of amino acids, ammonium, adenine, or allopurinol, in contrast with results reported for the cowpea enzyme. Gel-filtration chromatography and SDS-gel electrophoresis of uricase purified by the same method from cowpea nodules indicated that the native enzyme exists as a monomer of M, 50,000 at pH 7.5. Those legumes which transport fixed ni- trogen from the nodules in the form of ureides (allantoin and allantoic acid) syn- thesize the ureides by means of purine synthesis followed by oxidative degrada- tion (l-4). The degradative pathway con- sists of the three enzymes, xanthine de- hydrogenase, uricase (urate:Oz oxidore- 1 This research was supported by a grant to K.R.S. from the United States Department of Agriculture Science and Education Administration Competitive Research Grants Office (Grant 5901-0410-9-0948-O). M.J.B. was supported in part by a New Zealand Public Service Study Award and was on leave from the Ap- plied Biochemistry Division, D.S.I.R., Palmerston North, New Zealand. This is journal article No. 10169 of the Michigan Agricultural Experiment Station. ductase, EC 1.7.3.3), and allantoinase (3,5, 6). The purification and properties of xan- thine dehydrogenase from navy bean (Phmeolus vulgaris) (7) and soybean (Gig- tine m) (8) have recently been described. Uricase has been partially purified from cowpea (fiw ungu&&ta) (9). We report here the purification and properties of uri- case from soybean nodules, an enzyme which shows some similarities and a num- ber of important differences from the en- zymes from cowpea (9). A partial purifi- cation and some properties of the soybean nodule enzyme have been reported previ- ously (10). MATERIALS AND METHODS s To whom correspondence should be addressed: Growth ofplants Soybean seeds (G. - IL.1 Merr. Plant Sciences Division-OBE, Monsanto Agricultural cv. Amsoy 71) were inoculated with Rhizobium ja- Products Co., 800 N. Lindbergh Boulevard, St. Louis, ponicum strain USDA 311b 110. Cowpea seeds (V. MO. 63166. unguiculata [L.] Walp. cv. California Blackeye) were 0003-9861/83 $3.00 Copyright Q 1983 by Academic Press. Inc. All rights of reproductionin any form reserved. 190
Transcript
ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS Vol. 226, No. 1, October 1, pp. 190-197, 1933
Uricase from Soybean Root Nodules: Purification, Properties, and Comparison with the Enzyme from Cowpea’
KAREN LUCAS, MICHAEL J. BOLAND, AND KAREL R. SCHUBERT
Department of Biochemistry, Michisan State University, East Lansing, Michigan 48824
Received February 24.1933, and in revised form May 13,1933
A 45-fold purification of uricase (urate:08 oxidoreductase, EC 1.7.3.3) from soybean root nodules by ammonium sulfate fractionation, gel filtration, and affinity chroma- tography is described. Electrophoresis on nondenaturing gels using an activity stain or on sodium dodecyl sulfate (SDS) gels demonstrated that the enzyme obtained was nearly homogeneous. The subunit molecular weight of uricase estimated from SDS gels was 32,000 -t 3000. Gel-filtration studies indicated that the native enzyme is a monomer at pH 7.5 which associates to form a dimer at pH 8.8. Enzyme activity was stabilized by the addition of dithiothreitol. The pH dependence of the enzyme showed an optimum of 9.5. Initial rate kinetics showed Km values of 10 and 31 PM for uric acid and oxygen, respectively, with an intersecting pattern of substrate dependence. Uricase activity was inhibited strongly by xanthine, which was competitive with respect to uric acid (Ki = 10 PM). No significant inhibition was observed in the presence of a variety of amino acids, ammonium, adenine, or allopurinol, in contrast with results reported for the cowpea enzyme. Gel-filtration chromatography and SDS-gel electrophoresis of uricase purified by the same method from cowpea nodules indicated that the native enzyme exists as a monomer of M, 50,000 at pH 7.5.
Those legumes which transport fixed ni- trogen from the nodules in the form of ureides (allantoin and allantoic acid) syn- thesize the ureides by means of purine synthesis followed by oxidative degrada- tion (l-4). The degradative pathway con- sists of the three enzymes, xanthine de- hydrogenase, uricase (urate:Oz oxidore-
1 This research was supported by a grant to K.R.S. from the United States Department of Agriculture Science and Education Administration Competitive Research Grants Office (Grant 5901-0410-9-0948-O). M.J.B. was supported in part by a New Zealand Public Service Study Award and was on leave from the Ap- plied Biochemistry Division, D.S.I.R., Palmerston North, New Zealand. This is journal article No. 10169 of the Michigan Agricultural Experiment Station.
ductase, EC 1.7.3.3), and allantoinase (3,5, 6). The purification and properties of xan- thine dehydrogenase from navy bean (Phmeolus vulgaris) (7) and soybean (Gig- tine m) (8) have recently been described. Uricase has been partially purified from cowpea (fiw ungu&&ta) (9). We report here the purification and properties of uri- case from soybean nodules, an enzyme which shows some similarities and a num- ber of important differences from the en- zymes from cowpea (9). A partial purifi- cation and some properties of the soybean nodule enzyme have been reported previ- ously (10).
MATERIALS AND METHODS
s To whom correspondence should be addressed: Growth ofplants Soybean seeds (G. - IL.1 Merr. Plant Sciences Division-OBE, Monsanto Agricultural cv. Amsoy 71) were inoculated with Rhizobium ja- Products Co., 800 N. Lindbergh Boulevard, St. Louis, ponicum strain USDA 311b 110. Cowpea seeds (V. MO. 63166. unguiculata [L.] Walp. cv. California Blackeye) were
0003-9861/83 $3.00 Copyright Q 1983 by Academic Press. Inc. All rights of reproduction in any form reserved.
190
URICASE FROM SOYBEAN NODULES 191
inoculated with Rhizobium cowpea strain 32Hl. Plants were grown in Perlite and watered daily with a ni- trogen-free nutrient solution (11). Nodules were har- vested from plants 5 to 8 weeks old.
Uricase assay. Uricase activity was assayed at room temperature (22°C) by following the disappearance of uric acid at 292.5 nm (3), using E = 12.2 mM-' cm-’ (10). A unit of activity is defined as 1 pmol uric acid oxidized min-‘. Assay solutions contained 0.1 M 2(N- cyclohexylamino)ethanesulfonic acid (Ches)S-KOH, pH 9.5, saturated with air, 0.1 rnhf uric acid, and lo- 100 ~1 of enzyme solution in a total volume of 1.5 ml. Because of a lag phase of l-2 min after addition of uric acid, assays were run for several minutes to allow maximum velocity to be reached.
Kinetic studies. The pH dependence was measured using the standard assay method described above in different buffers. Buffers used were 0.1 M N- Tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid (Tes)-KOH for pH 6.5-8.4 and 0.1 M Ches-KOH for pH 8.4-11.1. Substrate-dependence experiments were run in mixtures of air-sparged and degassed, nitrogen-sparged buffer in anaerobic cuvettes with varying amounts of uric acid. Air-saturated buffer was used with varying concentrations of uric acid for xanthine inhibition studies. For other inhibitor stud- ies, 18 pM uric acid in air-saturated buffer was used.
Native gel electrophureeis. Native gels (9% total monomer concentration) were prepared in 7.5 mm X 13-cm glass tubes according to Davis (12) without a stacking gel and prerun for 30 min at 2 mA per gel before loading on the protein samples. Samples were subjected to electrophoresis at 2 mA per gel for 5 h. Gels were either stained for protein with 1.25g Coomassie blue R per liter 25% isopropanol:lO% acetic acid (v/v) or for enzyme activity. The activity stain contained (per 100 ml) 40 mg nitroblue tetrazolium, 0.8 mg phenazine methosulfate, and 20 mg uric acid in 50 mrd Tris-glycine, pH 8.3. Gels were incubated until activity hands were visible and then stored in 5% (v/v) acetic acid.
SDS-gel electrophoresis. SDS-gel electrophoresis was carried out on 10% polyacrylamide slab gels (13). A l-ml aliquot of enzyme from the blue-Sepharose column was lyophilized and taken up in 0.2 ml 0.1 M
Tris-HCl buffer, pH 6.8. Samples of enzymes that had not been concentrated and molecular weight standards were also used. All samples were boiled for 2 min with 2% SDS and 10% /3-mercaptoethanol before loading onto the gel. Bromphenol blue was used as
tracking dye and gels were electrophoresed at 2 mA cm-‘. Gels were stained with Coomassie blue.
Protein &ternzination Protein was estimated by a modification of the method of Lowry et al as described by Bensadoun and Weinstein (14).
Enzyme p-urificntia. All operations were carried out at 0-4’C. Nodules (5-50 g) were harvested and crushed in 2 ml of 25 mM Tes-KOH, pH 7.5, containing 5 mM dithiothreitol, 5% acid-washed insoluble poly- vinylpyrrolidone (PVP), and 5.5% sorbitol per gram of nodules. PVP was added to prevent protein de- naturation by phenolics (15). The brei was centrifuged for 10 min at 121g and the supernatant fluid was recentrifuged for 15 min at 27,OOOg. The supernatant fluid was taken to 35% saturation with ammonium sulfate (202 g liter-‘) and let stand for 30 min. The precipitate was removed by centrifugation at 27,OOOg for 20 min and redissolved in a minimum volume (4- 16 ml) of 10 mM Tes-KOH, 2.5 mM dithiothreitol, pH 7.5, and loaded onto a column (2.5 X 75 cm) of Se- pharose 6B. The column was developed with the same buffer at a flow rate of 48 ml h-l, and 8-ml fractions were collected. Fractions showing peak activity were pooled. If further purification was required, uricase was applied to a 0.25 X 5-cm column of reactive blue- Sepharose CL-6B which was then washed with 20 ml Tes-dithiothreitol buffer and developed with the same buffer saturated with uric acid. Fractions showing appreciable activity were pooled.
Catakwe assay. The disappearance of HzOz was fol- lowed at 240 nm (16). Reaction mixtures contained 20 mM H202 in 0.1 M phosphate, pH 7.5.
RESULTS AND DISCUSSION
Enzyme pur$catim Results of a typical enzyme purification are presented in Table I. The final purification was 45fold, with a recovery of 25%. Losses during purifi- cation were mostly due to the sacrifice of fractions showing less than maximum spe- cific activity, particularly after gel-filtra- tion chromatography. Dithiothreitol was required to maintain activity, and replace- ment with /3-mercaptoethanol caused a rapid and irreversible loss of activity. The enzyme was not stable upon freezing at -20 or -80°C but was relatively stable at 0-4°C (less than 10% loss of activity after storage for 3 months). The final specific activity was 6.8 pmol min-’ mg protein-’ at 22”C, fourfold higher than that obtained in an earlier preparation (10). The uv spec- trum of the purified protein exhibited a maximum at 2’7’7 nm and a 280/260 ratio
192 LUCAS, BOLAND, AND SCHUBERT
TABLE I
PURIFICATION OF URICASE FROM SOYBEAN NODULES
Volume Activity Protein Specific activity Percentage (ml) (pm01 min-‘) (mg) (U mg protein-‘) recove@
a The activity of the crude extract is defined as 100%; other values are corrected for samples withheld at each step.
of 1.15, which suggest that the enzyme was not contaminated with phenolics.
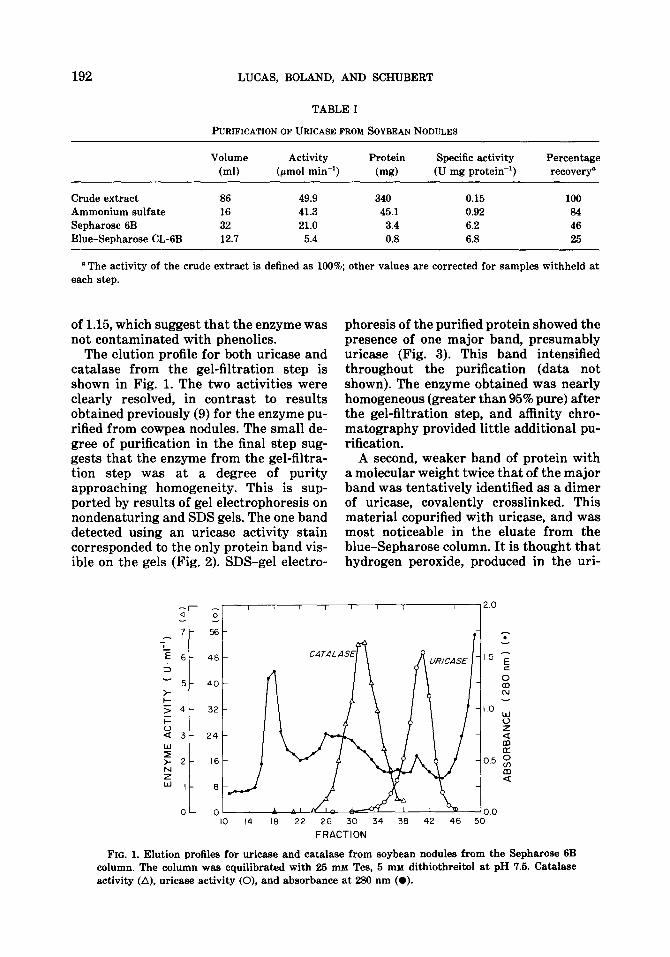
The elution profile for both uricase and catalase from the gel-filtration step is shown in Fig. 1. The two activities were clearly resolved, in contrast to results obtained previously (9) for the enzyme pu- rified from cowpea nodules. The small de- gree of purification in the final step sug- gests that the enzyme from the gel-filtra- tion step was at a degree of purity approaching homogeneity. This is sup- ported by results of gel electrophoresis on nondenaturing and SDS gels. The one band detected using an uricase activity stain corresponded to the only protein band vis- ible on the gels (Fig. 2). SDS-gel electro-
phoresis of the purified protein showed the presence of one major band, presumably uricase (Fig. 3). This band intensified throughout the purification (data not shown). The enzyme obtained was nearly homogeneous (greater than 95% pure) after the gel-filtration step, and affinity chro- matography provided little additional pu- rification.
A second, weaker band of protein with a molecular weight twice that of the major band was tentatively identified as a dimer of uricase, covalently crosslinked. This material copurified with uricase, and was most noticeable in the eluate from the blue-Sepharose column. It is thought that hydrogen peroxide, produced in the uri-
24
IO 14 18 22 26 30 34 38 42 46 50
FRACTION
FIG. 1. Elution profiles for uricase and catalase from soybean nodules from the Sepharose 6B column. The column was equilibrated with 25 rnM Tes, 5 rnrd dithiothreitol at pH 7.5. Catalase activity (A), uricase activity (0), and absorbance at 280 nm (0).
URICASE FROM SOYBEAN NODULES 193
FIG. 2. Gel electrophoresis of uricase under non- denaturing conditions. Purified uricase (20 fig of pro- tein from the pooled peak fractions off of a Sepharose 6B column) was subjected to electrophoresis for 5 h at 2 mA/gel. Gels were stained as follows: (A) activity stain minus uric acid, (B) activity stain with 0.2 mg/ ml uric acid, and (C) Coomassie blue.
case-catalyzed reaction, could cause oxi- dative crosslinking of enzyme molecules. This reaction would be most favorable when the reaction with uric acid occurs at high concentrations of enzyme and in the absence of catalase. Such conditions were present during the elution and in the eluate of the blue-Sepharose column. A third, very faint band which may be an impurity was sometimes visible at heavy loadings of protein.
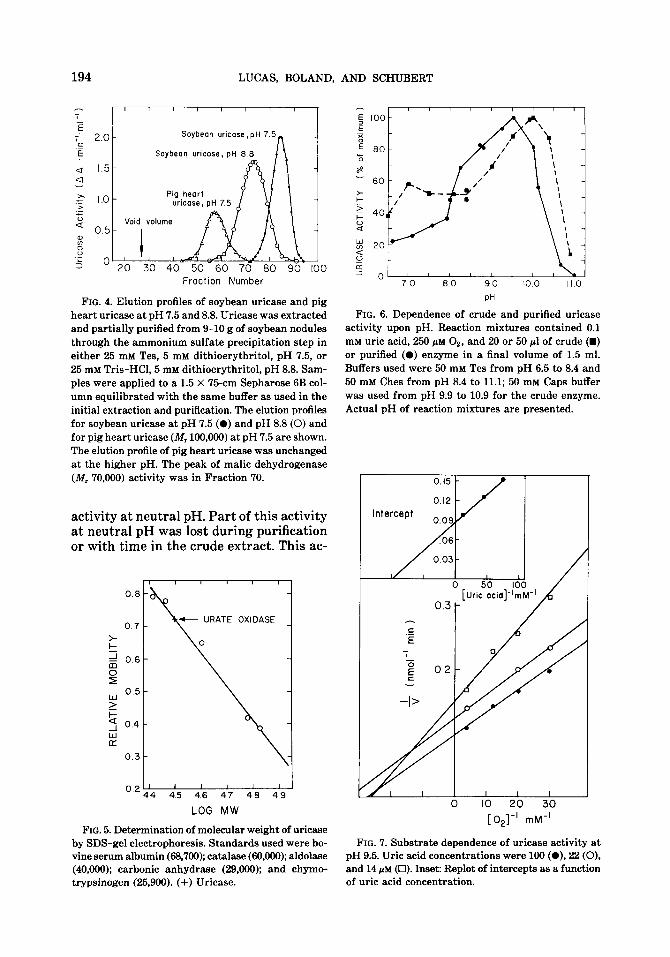
Molecular weight. The molecular weight of the enzyme was estimated by gel-filtra- tion chromatography. At pH 7.5, the ap- parent molecular weight of uricase was 35,000 f 5000. The estimated molecular weight was twice that for the enzyme ex- tracted and purified at pH 8.8 (Fig. 4). Based on the relative mobility after SDS- gel electrophoresis, a subunit molecular
weight of 32,000 f 3000 was obtained (Fig. 5). A value of 68,000 was obtained for the second, weaker band. These results suggest that the native enzyme at pH 7.5 is a monomer with a molecular weight of about 32,000 to 35,000. At pH 8.8, the enzyme as- sociates to form a dimer of about 68,000. This is an exceptionally low molecular weight for uricase. The enzyme from most other sources has a molecular weight of about 100,000 (17).
pH dependence. The pH dependence of purified uricase from soybean nodules can be seen in Fig. 6. The maximum activity at pH 9.5 is in accord with a pH optimum of between 9 and 10 observed for the pu- rified cowpea enzyme (9) and an early re- port on the soybean enzyme (10). Studies on the pH dependence of uricase in crude extracts showed a significant shoulder of
- Origin
-Dimer ?
- Urate oxidase monomer
-Dye +
FIG. 3. SDS-gel electrophoresis of purified uricase from soybean nodules plus molecular weight markers. (A) 1.8 pg uricase after blue-Sepharose affinity chro- matography; (B) molecular weight standards, BSA, aldolase, carbonic anhydrase; (C) 45 pg of purified uricase.
194 LUCAS, BOLAND, AND SCHUBERT
T 2
; 2.0 Soybean uricose, pH 7
.E E Soybean uricase, pH 8.8
u 1.5 a
a I.0 .z .> t u 0.5 % z .- 5 ’ 20 30 40 50 60 70 80 90 100
Fraction Number
FIG. 4. Elution profiles of soybean uricase and pig heart uricase at pH 7.5 and 8.8. Uricase was extracted and partially purified from 9-10 g of soybean nodules through the ammonium sulfate precipitation step in either 25 mM Tes, 5 mM dithioerythritol, pH 7.5, or 25 mM Tris-HCl, 5 mM dithioerythritol, pH 8.8. Sam- ples were applied to a 1.5 X ‘75-cm Sepharose 6B col- umn equilibrated with the same buffer as used in the initial extraction and purification. The elution profiles for soybean uricase at pH 7.5 (0) and pH 8.8 (0) and for pig heart uricase (IV, 100,000) at pH 7.5 are shown. The elution profile of pig heart uricase was unchanged at the higher pH. The peak of malic dehydrogenase (M, 70,000) activity was in Fraction 70.
activity at neutral pH. Part of this activity at neutral pH was lost during purification or with time in the crude extract. This ac-
0.8
Y
0.5
F 4 0.4
kt 0.3
0.2 i 44 4.5 4.6 4.7 4.6 4.9
LOG MW
FIG. 5. Determination of molecular weight of uricase by SDS-gel electrophoresis. Standards used were bo- vine serum albumin (68,700); catalase (60,000); aldolase (40,000); carbonic anhydrase (29,000); and chymo- trypsinogen (25,900). (+) Uricase.
PH
FIG. 6. Dependence of crude and purified uricase activity upon pH. Reaction mixtures contained 0.1 mM uric acid, 250 @M Oz, and 20 or 50 gl of crude (m) or purified (0) enzyme in a final volume of 1.5 ml. Buffers used were 50 mM Tes from pH 6.5 to 8.4 and 50 mM Ches from pH 8.4 to 11.1; 50 mM Caps buffer was used from pH 9.9 to 10.9 for the crude enzyme. Actual pH of reaction mixtures are presented.
I I I
IO 20 30
[ 02]-’ mM-’
FIG. 7. Substrate dependence of uricase activity at pH 9.5. Uric acid concentrations were 100 (O), 22 (0), and 14 PM (Cl). Inset: Replot of intercepts as a function of uric acid concentration.
URICASE FROM SOYBEAN NODULES 195
0 IO 20 30 [xanthine] mM
I I I I I I
0 IO 20 30 40 50 60
[Uric acid]-’ mM-’
FIG. 8. Inhibition of uricase activity by xanthine at pH 9.5 in the presence of 250 qd oxygen. Concen- trations of xanthine used were 0 (O), 10 (O), 20 (A), and 30 (0) pM. Inset: Replot of slopes as a function of xanthine concentrations.
tivity may be due to an unstable isozyme of uricase, such as that described from radicles of developing soybean seedlings (10). Even the purified enzyme exhibited a shoulder of activity at neutral pH. This may be due to the reduced catalytic activity of monomeric uricase. Alternatively, small amounts of the dimer may be in equilib- rium with the monomer at neutral pH and account for the activity.
Substrate dependent. The dependence of the reaction rate on substrate concentra- tion is given in Fig. 7, which indicates an intersecting pattern of lines, i.e., the re- action involves a ternary complex. Lines were fitted using a weighted least-squares method (18). Each point is the mean of three replicates, and the degree of scatter is mostly due to difficulty in maintaining the required oxygen concentrations. The intercept replot gives a K, for uric acid of 10 f 1 PM. When these results were plotted as a function of uric acid concentration (not shown), a K, for oxygen of 31 & 5 PM was obtained from the intercept replot.
The K, values found here do not differ markedly from values of 18 PM for uric acid and 29 PM for oxygen reported for the cowpea enzyme (9). A Km for uric acid of 8.1 PM has been reported for the soybean
TABLE II
COMPARISON BETWEEN PURIFIED SOYBEAN AND COWPEA URICASE
Property Soybean Cowpea”
pH optimum Km (Uric acid) Km (0,) Specific activityb Molecular weight
Catalase activity Xanthine inhibition Inhibition by NH:, glutamine,
adenine, allantoin
9.5 10 PM 31 &LM
10.0 gmol-’ mg-’ 32,000 (pH 7.5) 68,000 (pH 8.8)
No Strong
Weak, if at all
9-10 18 /.LM 29 pM
5.7 pm01 min-’ mg-’ 50,000 (pH7.5)
190,000 (pH 8.8) Yesd
Strong
Strong
“Data from Ref. (9). * Corrected for temperature of assays in (9) (30°C). ‘Results obtained in this work. d Not confirmed in this work.
196 LUCAS, BOLAND, AND SCHUBERT
I I
40 FRACTION
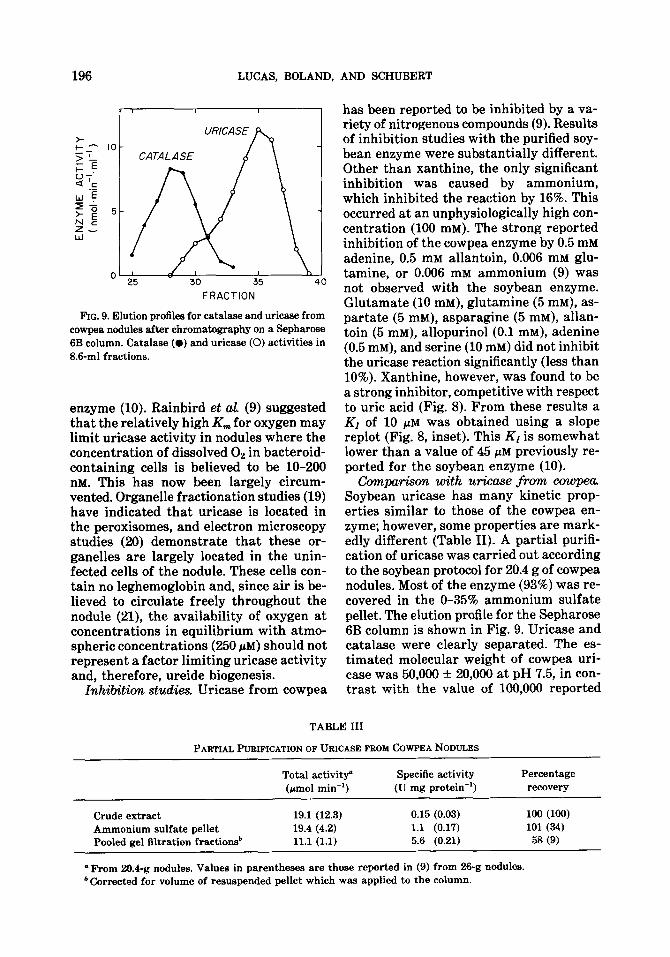
FIG. 9. Elution profiles for catalase and uricase from cowpea nodules after chromatography on a Sepharose 6B column. Catalase (0) and uricase (0) activities in 8.6-ml fractions.
enzyme (10). Rainbird et al. (9) suggested that the relatively high Km for oxygen may limit uricase activity in nodules where the concentration of dissolved Oz in bacteroid- containing cells is believed to be lo-200 nM. This has now been largely circum- vented. Organelle fractionation studies (19) have indicated that uricase is located in the peroxisomes, and electron microscopy studies (20) demonstrate that these or- ganelles are largely located in the unin- fected cells of the nodule. These cells con- tain no leghemoglobin and, since air is be- lieved to circulate freely throughout the nodule (21), the availability of oxygen at concentrations in equilibrium with atmo- spheric concentrations (250 PM) should not represent a factor limiting uricase activity and, therefore, ureide biogenesis.
Inhibition studies. Uricase from cowpea
has been reported to be inhibited by a va- riety of nitrogenous compounds (9). Results of inhibition studies with the purified soy- bean enzyme were substantially different. Other than xanthine, the only significant inhibition was caused by ammonium, which inhibited the reaction by 16%. This occurred at an unphysiologically high con- centration (100 mM). The strong reported inhibition of the cowpea enzyme by 0.5 mM adenine, 0.5 mM allantoin, 0.006 mM glu- tamine, or 0.006 mM ammonium (9) was not observed with the soybean enzyme. Glutamate (10 mM), glutamine (5 mM), as- partate (5 mM), asparagine (5 InM), allan- toin (5 mM), allopurinol (0.1 mM), adenine (0.5 IrIM), and serine (10 mM) did not inhibit the uricase reaction significantly (less than 10%). Xanthine, however, was found to be a strong inhibitor, competitive with respect to uric acid (Fig. 8). From these results a K1 of 10 I.~M was obtained using a slope replot (Fig. 8, inset). This KI is somewhat lower than a value of 45 PM previously re- ported for the soybean enzyme (10).
Com~+m with urkase from cowpea. Soybean uricase has many kinetic prop- erties similar to those of the cowpea en- zyme; however, some properties are mark- edly different (Table II). A partial purifi- cation of uricase was carried out according to the soybean protocol for 20.4 g of cowpea nodules. Most of the enzyme (93%) was re- covered in the O-35% ammonium sulfate pellet. The elution profile for the Sepharose 6B column is shown in Fig. 9. Uricase and catalase were clearly separated. The es- timated molecular weight of cowpea uri- case was 50,000 + 20,006 at pH 7.5, in con- trast with the value of 100,000 reported
TABLE III
PARTIAL PURIFICATION OF URICASE FROM COWPEA NODULES
Total activity” Specific activity Percentage (pm01 min-‘) (U mg protein-‘) recovery
o From 20.4-g nodules. Values in parentheses are those reported in (9) from 26-g nodules. b Corrected for volume of resuspended pellet which was applied to the column.
URICASE FROM SOYBEAN NODULES 197
previously for the enzyme at pH 8.8 (9). The estimated subunit molecular weight was 50,000 (data not shown). The formation of a dimer at pH 8.8, as observed for the soybean enzyme, would account for this apparent anomaly. Our purification is summarized in Table III. Results reported previously are given in parentheses. The yields and specific activity for cowpea uri- case purified according to the procedure developed for soybeans were several-fold higher than those reported previously (9). Three significant differences exist between the two methods of purification: (i) the in- clusion of PVP in the extraction medium which helps prevent denaturation of pro- teins by phenolic compounds in the nodule tissue; (ii) the use of dithiothreitol in buff- ers throughout purification which stabi- lizes enzyme activity; and (iii) the differ- ence in pH used during the isolation and purification of uricase.
ACKNOWLEDGMENTS
We would like to thank Dr. N. E. Tolbert and Dr. C. Sueiter for their review of this manuscript.
REFERENCES
1. ATKINS, C. A., RAINBIRD, R. M., AND PATE, J. S. (1980) 2. PJEanzenphysioL 97,249-260.
2. FUJIHARA, S., AND YAMAGUCHI, V. (1978) Plant PhysioL 62, 495-498.
3. SCHUBERT, K. R. (1981) Plant PhysioL 68, 1115- 1122.
4. BOLAND, M. J., AND SCHUBERT, K. R. (1981) Arch. Biochem Biophys. 209,524-533.
5. TRIPLEXT, E. W., BLEVINS, D. E., AND RANDALL, D. D. (1980) Plant PhysioL 65, 1203-1206.
6. ATKINS, C. A. (1981) FEBS ktt. 125, 89-93. 7. BOLAND, M. J. (1981) Biochem. Int. 2,567-574. 8. TRIPLETT, E. W., BLEVINS, D. G., AND RANDALL,
D. D. (1981) Plant PhysioL 675, 27. 9. RAINBIRD, R. M., AND ATKINS, C. A. (1981) B&him
Biophys. Actu 659, 132-140. 10. TAJIMA, S., AND YAMAMOTO, Y. (1975) Plant Cell
PhysioL 16,271-282. 11. FISHBECK, K., EVANS, H. J., AND BOERSMA, L. L.
(1973) Apm J. 65,429-433. 12. DAVIS, B. J. (1964) Ann N. Y. Acud. Sci 121,404-
427. 13. LAEMMLI, U. K., ANDFAVRE, M. (1973)J. MoL BioL
80,575-599. 14. BENSADOLJN, A., AND WEINSTEIN, D. (1976) Anal
Biochem. 70,241-250. 15. LOOMIS, W. D. (1974) in Methods in Enzymology
(Fleischer, S., and Packer, L., eds.), Vol. 31, pp. 528-544, Academic Press, New York.
16. LUCK, H. (1965) in Methods of Enzymatic Analysis (Bergmeyer, H. U., ed.), 2nd ed., pp. 885-894, Academic Press, London/New York.
17. VOGELS, G. D., AND VAN DER DRIFT, C. (1976) Bac- terioL Rev. 40,403-468.
18. WILKINSON, G. N. (1969) Biodwm. J. 80,324. 19. HANKS, J. F., TOLBERT, N. E., AND SCHUBERT,
K. R. (1981) Plant PhysioL 68, 65-69. 20. NEWCOMB, E. H., ANDTANDON, S. I. (1981) Science
212, 1394-1396. 21. BERGERSEN, F. J., AND GOODCHILD, D. J. (1973)