Abstract. Mesoclemmys dahli and M. zuliae are two endangered, little-known toad-headed turtles with small distributionranges in Colombia and Venezuela, respectively. Using the mitochondrial cytochrome b gene as a marker, we investigatetheir phylogeographic differentiation. Furthermore, based on 2341 bp of mtDNA and 2109 bp of nDNA of M. dahli, M.zuliae and allied chelid turtles, we infer their divergence time using a fossil-calibrated relaxed molecular clock approach.Mesoclemmys dahli and M. zuliae are closely related species, with an estimated mean divergence time of 10.6 million years.This estimate correlates with the uplift of the Serranía de Perijá, an Andean mountain chain separating their distributionranges, suggesting that this event could have caused the evolution of the two species. Haplotype and nucleotide diversitiesof M. dahli are markedly higher than in Podocnemis lewyana, another endemic turtle species of Colombia. This pronounceddissimilarity may reflect differences in the phylogeographies and demographic histories of the two species, but also differenthabitat preferences.
While the phylogeographies of many WesternPalaearctic and Nearctic turtles and tortoisesare well-known (e.g. Walker et al., 1998; Lenket al., 1999; Weisrock and Janzen, 2000; Karland Wilson, 2001; Fritz et al., 2006, 2007,2008, 2009; Rosenbaum, Robertson and Za-mudio, 2007; Amato, Brooks and Fu, 2008;McGaugh, Eckerman and Janzen, 2008; Butleret al., 2011; Pedall et al., 2011; Ureña-Arandaand Espinosa de los Monteros, 2012), relativelyfew studies have focused on species from Cen-tral and South America so far (Souza et al.,2003; Pearse et al., 2006; Vargas-Ramírez et
1 - Museum of Zoology (Museum für Tierkunde), Sencken-berg Dresden, A. B. Meyer Building, D-01109 Dresden,Germany
2 - Department of Functional Morphology and Biome-chanics, Institute of Zoology, Christian-Albrechts-Universität zu Kiel, Am Botanischen Garten 1-9, D-24118 Kiel, Germany
3 - Instituto de Ciencias Naturales, Universidad Nacionalde Colombia, Apartado 7495, Bogotá, Colombia∗Corresponding author; e-mail:[email protected]
al., 2007, 2010a, 2012; González-Porter et al.,2011; Fritz et al., 2012a, 2012b). Among theSouth American species, members of the familyChelidae are the least studied ones, with just onepublication on the phylogeography of Hydrome-dusa maximiliani (Souza et al., 2003). Chelidae,with 23 species, is the most species-rich of theseven families of turtles and tortoises in con-tinental South America, and the most speciosegroup within this family are the ‘toad-headedturtles’ of the genus Mesoclemmys. Currently,ten Mesoclemmys species are recognized, mostof which occur in central South America (Bourand Zaher, 2005; Fritz and Havaš, 2007; vanDijk et al., 2011). Mesoclemmys dahli (Zangerland Medem, 1958) and M. zuliae (Pritchard andTrebbau, 1984) represent little-known, isolatednorthern species with relict character (Bour andZaher, 2005).
Dahl’s toad-headed turtle (M. dahli), endemicto Colombia, is the only chelid species occur-ring west of the Andes mountain range (Me-dem, 1966). It is known from the western partof the Caribbean region of Colombia, includ-ing the departments of Córdoba, Bolivar, Su-
dahli is listed in the threat category “Critically
Endangered” by the IUCN Red List of Threat-
ened Species (IUCN, 2011). The closely re-
lated Zulia toad-headed turtle (M. zuliae), en-
demic to Venezuela, is confined to the Zulia
river and its tributaries in the south-western part
of the Maracaibo basin (Pritchard and Trebbau,
1984; Rueda-Almonacid et al., 2007; fig. 1).
Acknowledging its small distribution range, M.
zuliae is listed in the category “Vulnerable”
by the IUCN Red List of Threatened Species
(IUCN, 2011).
To date, some studies on the ecology and nat-
ural history of the two species have been per-
formed (M. dahli: Medem, 1966; de la Ossa-
Velasquez, 1998; Castaño-Mora and Medem,
2002; Rueda-Almonacid et al., 2007; Forero-
Medina, Cárdenas-Arevalo and Castaño-Mora,
2011; M. zuliae: Rueda-Almonacid et al., 2007;
Rojas-Runjaic, 2009). Furthermore, the two
species were included in a study that investi-
gated the patterns of geographical distribution
of South American chelid turtles (Souza, 2005).
The two species have similar habitat prefer-
ences (Medem, 1966; Pritchard and Trebbau,
1984; Rueda-Almonacid et al., 2007) and are
morphologically very similar (see images and
descriptions in Pritchard and Trebbau, 1984;
Rueda-Almonacid et al., 2007). Yet, they have
never been studied and compared using molec-
ular markers. The present study aims to fill this
gap by analyzing their kinship at a molecular
genetic level and by applying a relaxed molec-
ular clock to elucidate their divergence and bio-
geography.
Materials and methods
Sampling and laboratory procedures
Twenty-five saliva samples of Mesoclemmys dahli and fourtissue samples of M. zuliae were studied (fig. 1; Appendix).The samples from M. dahli were collected in both regionswhere the species is known to occur; 18 samples came fromthe department of Córdoba and seven samples from the de-partment of Cesar. The samples from M. zuliae were col-lected at the type locality and two additional sites, coveringlargely its distribution range in north-south direction. Forinferring phylogeography, sequences of the mitochondrialcytochrome b gene (cyt b) were generated for all samples.For molecular clock calculations, the mitochondrial 12SrRNA and NADH dehydrogenase subunit 4 (ND4) genesand three nuclear loci, the oocyte maturation factor Mosgene (C-mos), the intron 1 of the RNA fingerprint pro-tein 35 gene (R35) and the recombination-activating gene2 (Rag 2), of one sample each of M. dahli and M. zuliaewere sequenced. Sequences of the same mitochondrial andnuclear DNA fragments were also produced for M. gibba,Phrynops geoffroanus and P. hilarii, as far as these were notavailable from GenBank (see Appendix).
Genomic DNA was extracted using a Qiagen DNA bloodextraction kit (Qiagen Benelux B.V., Venlo, The Nether-lands), following the manufacturer’s instructions. The 12SrRNA gene was amplified and sequenced using the primersL1091 and H1478 (Kocher et al., 1989), and for the par-tial ND4 gene plus adjacent DNA coding for tRNAs, theprimers L-ND4 and H-Leu (Stuart and Parham, 2004) wereapplied. The cyt b gene was amplified and sequenced intwo fragments overlapping by approximately 300 bp us-ing the primer pairs mt-a-neu3 + mt-E-Rev2 and mt-c-For2 + mt-f-na (Fritz et al., 2006; Praschag et al., 2007).For the nuclear loci the following primer pairs were used C-mos: G136 + G137 (Georges et al., 1998), R35: R35Ex1 +R35Ex2 (Fujita et al., 2004), and Rag 2: F2-1 + R2-1 (Le etal., 2006). PCRs were carried out in a total volume of 50 μlcontaining 1 unit Taq polymerase (Bioron, Ludwigshafen,Germany), 1 × buffer (as recommended by the supplier),0.5 μM of each primer, and 0.2 mM of each dNTP (Fer-mentas, St. Leon-Rot, Germany). PCR products were pu-rified using the ExoSAP-IT enzymatic cleanup (USB Eu-rope GmbH, Staufen, Germany; modified protocol: 30 minat 37°C, 15 min at 80°C) and sequenced on an ABI 3130xlGenetic Analyzer (Applied Biosystems, Foster City, CA,USA) using the BigDye Terminator v3.1 Cycle SequencingKit (Applied Biosystems).
Phylogeographic analyses
DNA sequences were edited and aligned using CHROMAS1.51 (http://www.technelysium.com.au/chromas.html) andBIOEDIT 7.0.5.2 (Hall, 1999). For the 1067-bp-long cyt bsequences of Mesoclemmys dahli, M. gibba and M. zuliae,uncorrected p distances were calculated in MEGA 5.05(Tamura et al., 2011). Evolutionary relationships betweensequences of M. dahli and M. zuliae were inferred usingTCS 1.21 (Clement et al., 2000). For M. dahli, haplotype
Weak genetic divergence between turtles Mesoclemmys dahli and M. zuliae (Testudines: Pleurodira: Chelidae) 375
Figure 1. Distribution of Mesoclemmys dahli and M. zuliae (based on Rueda-Almonacid et al., 2007) and sampling sites(colour-coded). For M. dahli, the Córdoba population is indicated in green and the Cesar population in orange; sites of M.zuliae, blue. The Serranía de Perijá is highlighted in red. Inset: juvenile Mesoclemmys dahli from the Cesar population (photo:Guido Medina).
diversity (h) and nucleotide diversity (π ) were calculatedusing DNASP 5.0 (Librado and Rozas, 2009). The influ-ence of geography on genetic divergence within M. dahliand between M. dahli and M. zuliae was examined usingMantel tests as implemented in the software IBD (Isolation-by-Distance; Bohonak, 2002). A first Mantel test was basedon uncorrected p distances and geographical distances (km)between the two populations of M. dahli and M. zuliae. Inanother Mantel test, the influence of the Serranía de Pe-rijá, an Andean mountain chain separating the distributionranges of M. dahli and M. zuliae, was examined by combin-ing a categorical matrix with genetic distances. In this ma-trix, the distance between the two populations of M. dahliwas coded with 0 (corresponding to the absence of a moun-tain barrier), while the distance between each population ofM. dahli and M. zuliae was coded with 1 (acknowledgingthe presence of the mountain barrier).
Relaxed molecular clock
The phylogeny of Mesoclemmys dahli, M. zuliae and otherchelid turtle species was inferred to obtain a backbone formolecular clock calculations. For this purpose, the 12SrRNA, cyt b, ND4, C-mos, R35 and Rag 2 sequences ofM. dahli and M. zuliae were aligned with homologous se-quence data of M. gibba, Chelus fimbriatus, Chelodina ru-
gosa, Phrynops geoffroanus, P. hilarii, Pelomedusa sub-rufa, Podocnemis expansa, and Indotestudo elongata (foraccession numbers, see Appendix). Sequences for eachtaxon were concatenated. The resulting supermatrix was of4450 bp length (including gaps), corresponding to 2341 bpof mtDNA and 2109 bp of nDNA. The partial 12S rRNAgene contributed 493 bp; the nearly complete cyt b gene,1067 bp; the partial ND4 gene plus adjacent DNA codingfor tRNAs, 781 bp; C-mos, 390 bp; R35, 1025 bp; andRag 2, 694 bp. Based on this supermatrix, phylogenetictrees were calculated using Maximum Likelihood (ML) andBayesian Inference (BI) analyses, applying the followingpartition schemes: (1) unpartitioned, (2) by gene, i.e., eachgene corresponds to a distinct partition, and (3) maximumpartitioning, i.e., using each codon of each protein-codinggene as distinct partition plus each non-protein-coding geneor DNA block as distinct partition. Indotestudo elongata(family Testudinidae, suborder Cryptodira) served for tree-rooting. ML analyses were run with RAxML 7.2.6 (Sta-matakis, 2006) using the graphical user interface raxml-GUI 0.93 (Silvestro and Michalak, 2011) and the GTR +G model across every partition. To explore the robustnessof the branching patterns, five independent ML searcheswere performed using the fast bootstrap algorithm. Sub-sequently, 1000 thorough bootstrap replicates were calcu-lated and plotted against the tree with the highest likeli-hood value. The BI analyses were run in MrBAYES 3.1
376 M. Vargas-Ramírez et al.
(Ronquist and Huelsenbeck, 2003) using four incrementallyheated Markov chains; posterior probabilities were obtainedfrom the 50% majority rule consensus tree. For each inde-pendent run, the variation in likelihood scores was exam-ined by plotting − ln L scores against the number of gener-ations, and the burn-in was set to sample only the plateau ofthe most likely trees. In a conservative approach, 40% of allsampled trees were discarded, although the plateau of like-lihood values had been reached before. The best-fit modelof nucleotide substitution was established for each partitionusing the Akaike information criterion of MrMODELTEST(Nylander, 2002) and incorporated into a single tree search(mixed model partition approach; Nylander et al., 2004).The following models were suggested for the partitioningby gene: 12S rRNA – GTR + G, cyt b – TVM + I + G,ND4 – GTR + G, t-RNAs – TVM + G, C-mos – K80 + G,R35 – TVM + G, and Rag 2 – HKY; and for the codons ofprotein-coding genes in the maximum partitioning scheme:cyt b 1st codon position – GTR + G, cyt b 2nd codon posi-tion – GTR + I + G, cyt b 3rd codon position – TIM + G,ND4 1st codon position – TVM + G, ND4 2nd codon po-sition – TVM + G, ND4 3rd codon position – HKY + I +G, C-mos 1st codon position – HKY + I, C-mos 2nd codonposition – TrNef + G, C-mos 3rd codon position – K81uf +G, Rag 2 1st codon position – TrN + G, Rag 2 2nd codonposition – GTR + G, and Rag 2 3rd codon position – HKY.
The divergence time of M. dahli and M. zuliae wasestimated by a Bayesian relaxed molecular clock ap-proach (MULTIDISTRIBUTE package; Thorne, Kishinoand Painter, 1998; Thorne and Kishino, 2002). Fossil evi-dence was used for constraining the minimum ages of twonodes within the obtained phylogeny. The split betweenChelodina rugosa and the South American chelids was cal-ibrated with the range of 125.0-99.6 million years (ma),based on the Lower Cretaceous record of Prochelidella cer-robarcinae (Cerro Barcino formation, Aptian-Albian?), theoldest South American chelid (de la Fuente et al., 2011;node A in fig. 4). Furthermore, the split between Pelome-dusidae (represented by Pelomedusa subrufa) and Podo-cnemididae (represented by Podocnemis expansa) was set tothe lower and upper boundaries of the Valanginian (140.4-136.2 ma), the Early Cretaceous stage from which the earli-est podocnemidoid turtle is known (Cadena, 2011; node B infig. 4). Chelodina rugosa is a representative of Australasianchelids that constitute the sister group of South Americanchelids (Georges et al., 1998), and Prochelidella is thoughtto be allied to the extant genus Acanthochelys of SouthAmerica (de la Fuente et al., 2011). Pelomedusa subrufaand Podocnemis expansa are representatives of the familiesPelomedusidae and Podocnemididae, respectively. Amongextant turtles, Pelomedusidae and Podocnemididae togetherare the sister group of Chelidae (Gaffney et al., 2006).
In order to determine the appropriate nucleotide substi-tution model parameters, the data set was analyzed usingthe program PAML 3.13 (Yang, 1997). Subsequently, thebranch lengths and their variance-covariance matrix wereestimated with the program ESTBRANCHES. Using the ap-plication MULTIDIVTIME, Markov chains were run threetimes with the settings numsamps = 1 000 000, sampfreq =
100 and a burn-in of 10 000 and compared to test the stabil-ity of the results. According to Cadena (2011), the prior forthe mean of the ingroup root age (rttm) was set to a min-imum of 161.2 ma (with 2 ma SD), corresponding to thesplit between (Pelomedusidae + Podocnemididae) and Che-lidae. Mean and standard deviation of the rate of molecularevolution at the ingroup root node (rtrate and rtratesd) were0.0004341 substitutions per site and million years with 1time unit = 1 ma (calculated with the mean of the branchlengths from ESTBRANCHES). Mean and standard devi-ation of the Brownian motion constant (brownmean andbrownsd) were set to 0.006203 and bigtime to 220 ma ac-cording to the age of the oldest known chelonian, Odon-tochelys semitestacea (Li et al., 2008).
Results
Phylogeography
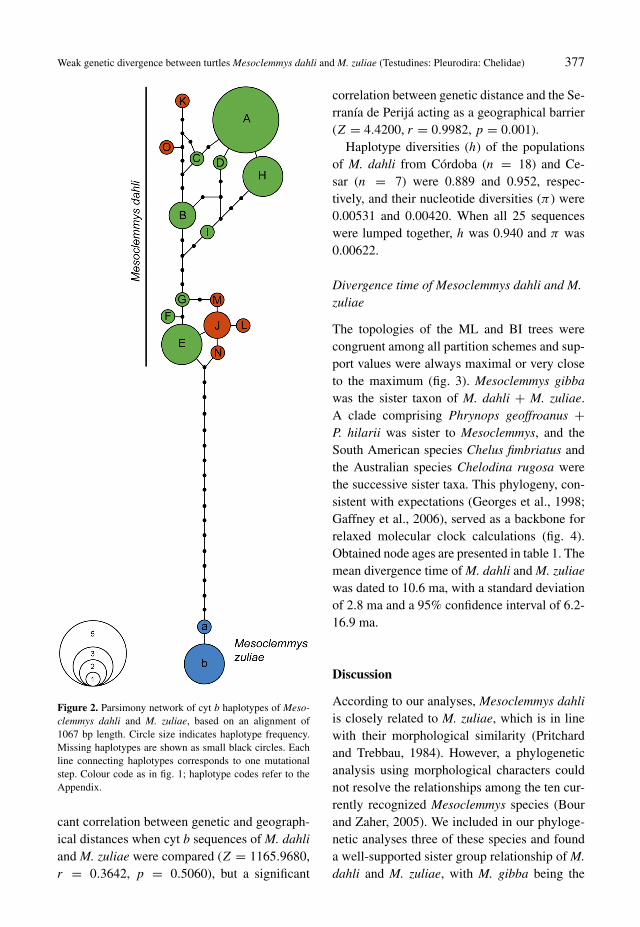
The 25 cyt b sequences of Mesoclemmys dahliand the four cyt b sequences of M. zuliae wereassigned to two clearly distinct haplotype clus-ters in the parsimony network (fig. 2), differ-ing by a minimum of 18 mutational steps. Se-quences of M. dahli corresponded to 15 hap-lotypes; sequences of M. zuliae, to two hap-lotypes. Among haplotypes of M. dahli, someloops occurred. The maximum number of muta-tional steps within M. dahli resembles the min-imum divergence between M. dahli and M. zu-liae. The two haplotypes of M. zuliae differedby only one mutation. For the two populationsof M. dahli, no shared haplotypes were ob-served. However, this could be a bias due tosmall sample size because the haplotypes ofeach population did not form a distinct cluster,but were rather randomly distributed in the net-work (fig. 2).
The mean uncorrected p distance between se-quences of M. dahli and M. zuliae was 2.22%;mean divergences within each species were0.58% and 0.04%, respectively. The two popu-lations of M. dahli differed by 0.77%. Withinthe Córdoba population of M. dahli a sequencedivergence of 0.41% occurred; within the Cesarpopulation the divergence was 0.33%. The se-quence of M. gibba differed by 14.16% fromthat of M. dahli and by 13.27% from that ofM. zuliae. The Mantel tests revealed no signifi-
Weak genetic divergence between turtles Mesoclemmys dahli and M. zuliae (Testudines: Pleurodira: Chelidae) 377
Figure 2. Parsimony network of cyt b haplotypes of Meso-clemmys dahli and M. zuliae, based on an alignment of1067 bp length. Circle size indicates haplotype frequency.Missing haplotypes are shown as small black circles. Eachline connecting haplotypes corresponds to one mutationalstep. Colour code as in fig. 1; haplotype codes refer to theAppendix.
cant correlation between genetic and geograph-ical distances when cyt b sequences of M. dahliand M. zuliae were compared (Z = 1165.9680,r = 0.3642, p = 0.5060), but a significant
correlation between genetic distance and the Se-rranía de Perijá acting as a geographical barrier(Z = 4.4200, r = 0.9982, p = 0.001).
Haplotype diversities (h) of the populationsof M. dahli from Córdoba (n = 18) and Ce-sar (n = 7) were 0.889 and 0.952, respec-tively, and their nucleotide diversities (π) were0.00531 and 0.00420. When all 25 sequenceswere lumped together, h was 0.940 and π was0.00622.
Divergence time of Mesoclemmys dahli and M.zuliae
The topologies of the ML and BI trees werecongruent among all partition schemes and sup-port values were always maximal or very closeto the maximum (fig. 3). Mesoclemmys gibbawas the sister taxon of M. dahli + M. zuliae.A clade comprising Phrynops geoffroanus +P. hilarii was sister to Mesoclemmys, and theSouth American species Chelus fimbriatus andthe Australian species Chelodina rugosa werethe successive sister taxa. This phylogeny, con-sistent with expectations (Georges et al., 1998;Gaffney et al., 2006), served as a backbone forrelaxed molecular clock calculations (fig. 4).Obtained node ages are presented in table 1. Themean divergence time of M. dahli and M. zuliaewas dated to 10.6 ma, with a standard deviationof 2.8 ma and a 95% confidence interval of 6.2-16.9 ma.
Discussion
According to our analyses, Mesoclemmys dahliis closely related to M. zuliae, which is in linewith their morphological similarity (Pritchardand Trebbau, 1984). However, a phylogeneticanalysis using morphological characters couldnot resolve the relationships among the ten cur-rently recognized Mesoclemmys species (Bourand Zaher, 2005). We included in our phyloge-netic analyses three of these species and founda well-supported sister group relationship of M.dahli and M. zuliae, with M. gibba being the
378 M. Vargas-Ramírez et al.
Figure 3. Bayesian tree based on 2341 bp of mitochondrial DNA (12 S rRNA, ND4, cyt b) and 2109 bp of nuclear DNAsequences (C-mos, R35, Rag 2; partitioned by gene). Support values are Bayesian posterior probabilities (top) and MLbootstrap values (bottom); asterisks indicate maximum support under both methods. This tree was used for the RelaxedMolecular Clock calculations.
successive sister taxon (fig. 3). Compared to M.gibba, M. dahli and M. zuliae seem to be weaklydifferentiated. The number of mutational stepsamong haplotypes of M. dahli resembles the ob-served divergence between M. dahli and M. zu-liae (fig. 2), and this pattern is also reflected bylow between-species divergences with respectto uncorrected p distances of the cyt b gene.The sequence divergence between M. dahli andM. zuliae amounts only to 2.22%, while thesetwo species differ from M. gibba by 14.16% and13.27%, respectively. With respect to cyt b se-quences, the value of 2.22% is close to the lowerdivergence limit for any distinct congeneric tur-tle and tortoise species. Yet, for some emy-did turtles (Emys, Graptemys, Trachemys) evenlower species divergence values were reported
(Lamb et al., 1994; Fritz et al., 2005, 2012b). Inanalogy to the widely used barcoding approach(Hebert, Ratnasingham and de Waard, 2003) re-lying on divergences of the COI gene as a yard-stick, uncorrected p distances of the cyt b genehave been frequently used as a tool for speciesdelineation of chelonians (e.g. Vargas-Ramírezet al., 2010b; Praschag et al., 2011; Stuckasand Fritz, 2011; Fritz et al., 2012a, 2012b;Kindler et al., 2012). However, these studiespointed out that no universal thresholds shouldbe used because the critical divergence valuediffers among various chelonian groups. There-fore, additional Mesoclemmys species need tobe studied before the status of M. dahli andM. zuliae as distinct species should be chal-lenged.
Weak genetic divergence between turtles Mesoclemmys dahli and M. zuliae (Testudines: Pleurodira: Chelidae) 379
Figure 4. Divergence time estimates using the MULTIDISTRIBUTE package. Dark grey bars at nodes represent 95%confidence intervals. The light grey column on the right shows the time of the main uplift of the Serranía de Perijá (LateMiocene-Pliocene; Kellogg, 1984). Letters indicate nodes calibrated with fossil evidence (see table 1). PP, Plio-Pleistocene.
Table 1. Results of the relaxed molecular clock analysis using the MULTIDISTRIBUTE package. SD = Standard deviation,CI = 95% confidence interval. All dates are given in million years (ma).
∗ Node A (fig. 4): Fossil constraint: 125.0-99.6 ma (Prochelidella cerrobarcinae; de la Fuente et al., 2011).∗∗ Node B (fig. 4): Fossil constraint: 140.4-136.2 ma (earliest podocnemidoid turtle; Cadena, 2011).
Haplotype and nucleotide diversities of M.dahli are much higher than in another en-demic turtle species of Colombia, Podocnemislewyana. Based on the rapidly evolving D-loop,haplotype diversity (h) and nucleotide diversity(π) of 119 samples of P. lewyana were 0.292and 0.00006, respectively (Vargas-Ramírez etal., 2012), contrary to h = 0.940 and π =
0.00622 in the 25 samples of M. dahli analyzedin the present study for the more slowly evolv-ing cyt b gene. This pronounced dissimilaritymay reflect differences in the phylogeographiesand demographic histories of the two species,but also different habitat preferences. While P.lewyana is a true river turtle, M. dahli is typi-cally found in shallow and quiet ponds or small
380 M. Vargas-Ramírez et al.
brooks in seasonally dry tropical forest (Me-dem, 1966; Rueda-Almonacid et al., 2007), sug-gesting that the patchy structures in such habitatmay favour more genetic diversity than the oc-currence along a river course.
Souza (2005) proposed that the distributionof South American chelids is mainly corre-lated with major river or drainage basins. Ac-cording to this hypothesis, the distribution ofM. dahli and M. zuliae is associated with theMagdalena and Orinoco basins, respectively.Notwithstanding that M. dahli occurs close tothe Magdalena river, its typical habitats are nei-ther directly linked to this river nor to its ma-jor tributaries. Mesoclemmys zuliae lives in sim-ilar habitats as M. dahli. Moreover, the rangeof M. zuliae is far away from the Orinoco(Rueda-Almonacid et al., 2007), and the riversand streams of the Maracaibo basin are notconnected to the Orinoco basin. Unlike trueriver turtles, as Podocnemis species, M. dahliand M. zuliae seem therefore to be not tied tothe ecosystems of the Magdalena and Orinocorivers. Consequently, it is unlikely that the ori-gin of M. dahli and M. zuliae is directly re-lated to the hydrographic history of these rivers.However, the estimated mean divergence timeof M. dahli and M. zuliae correlates quite wellwith the uplift of the Serranía de Perijá (seeabove) in the Late Miocene and Pliocene (Kel-logg, 1984; fig. 4), suggesting that this oroge-netic process could be directly responsible forthe evolution of the two species. Also in an-other South American chelid species, Hydrome-dusa maximiliani from eastern Brazil, phylo-geographic structure is shaped by mountainchains, and the estimated divergence time of themajor phylogeographic groups within H. maxi-miliani fits the uplift of the respective mountainsduring the Pliocene and Miocene (Souza et al.,2003).
Acknowledgements. We thank the Grupo de Conservacióny Biodiversidad of the Instituto de Ciencias Naturales dela Universidad Nacional de Colombia, Fundación Biodi-versa Colombia and Instituto de Biología Tropical Roberto
Franco (IBTRF) de la Universidad Nacional de Colombiafor institutional and logistical support. Carl J. Franklin (Am-phibian and Reptile Research Center, University of Texasat Arlington) and Ingo Pauler provided samples of Meso-clemmys zuliae. Christian Kehlmaier, Edgar Lehr, AnkeMüller, and Anja Rauh assisted in the lab. Ylenia Chiari andJoão Lourenço shared sequences of Phrynops hilarii withus. Massimo Delfino, Marcelo Sánchez-Villagra, MarkusWilmsen and Juliana Sterli helped with literature of fos-sil chelids. Thanks to Guido Medina for the photograph ofM. dahli. Mario Vargas-Ramírez’ research in Germany isfunded by the Humboldt Foundation (Georg Forster fellow-ship).
References
Amato, M.L., Brooks, R.J., Fu, J. (2008): A phylogeo-graphic analysis of populations of the wood turtle(Glyptemys insculpta) throughout its range. Mol. Ecol.17: 570-581.
Bohonak, A.J. (2002): IBD (Isolation-by-Distance): a pro-gram for analyses of isolation by distance. J. Hered. 93:153-154.
Bour, R., Zaher, H. (2005): A new species of Mesoclemmys,from the open formations of Northeastern Brazil (Che-lonii, Chelidae). Pap. Avul. Zool. 45: 295-311.
Butler, J.M., Dodd, C.K., Aresco, M., Austin, J.D. (2011):Morphological and molecular evidence indicates that theGulf Coast box turtle (Terrapene carolina major) is nota distinct evolutionary lineage in the Florida Panhandle.Biol. J. Linn. Soc. 102: 889-901.
Cadena, E.A. (2011): Potential earliest record of podocne-midoid turtles from the Early Cretaceous (Valanginian)of Colombia. J. Paleontol. 85: 877-881.
Castaño-Mora, O.V., Medem, F. (2002): Batrachemys dahli.In: Libro rojo de reptiles de Colombia. Libros rojos deespecies amenazadas de Colombia, p. 77-79. Castaño-Mora, O.V., Ed., Instituto de Ciencias Naturales (Univer-sidad Nacional de Colombia), Ministerio del Medio Am-biente and Conservación Internacional, Bogotá, Colom-bia.
Clement, M., Posada, D., Crandall, K.A. (2000): TCS:a computer program to estimate gene genealogies. Mol.Ecol. 9: 1657-1660.
de la Fuente, M., Umazano, A.M., Sterli, J., Carballido, J.L.(2011): New chelid turtles of the lower section of theCerro Barcino formation (Aptian-Albian?), Patagonia,Argentina. Cretaceous Res. 32: 527-537.
de la Ossa-Velasquez, J.L. (1998): Phrynops dahli: a littleknown turtle endemic to the Caribbean coast of Colom-bia. Reptilia 3: 47-50.
Forero-Medina, G., Cárdenas-Arevalo, G., Castaño-Mora,O.V. (2011): Abundance, home range, and movementpatterns of the endemic species Dahl’s toad-headed tur-tle (Mesoclemmys dahli) in Cesar, Colombia. Chelon.Conserv. Biol. 10: 228-236.
Fritz, U., Havaš, P. (2007): Checklist of chelonians of theworld. Vertebr. Zool. 57: 149-368.
Weak genetic divergence between turtles Mesoclemmys dahli and M. zuliae (Testudines: Pleurodira: Chelidae) 381
Fritz, U., Fattizzo, T., Guicking, D., Tripepi, S., Pennisi,M.G., Lenk, P., Joger, U., Wink, M. (2005): A newcryptic species of pond turtle from southern Italy, thehottest spot in the range of the genus Emys. Zool. Scr.34: 351-371.
Fritz, U., Auer, M., Bertolero, A., Cheylan, M., Fattizzo, T.,Hundsdörfer, A.K., Martín Sampayo, M., Pretus, J.L.,Široký, P., Wink, M. (2006): A rangewide phylogeog-raphy of Hermann’s tortoise, Testudo hermanni (Rep-tilia: Testudines: Testudinidae): implications for taxon-omy. Zool. Scr. 35: 531-543.
Fritz, U., Guicking, D., Kami, H., Arakelyan, M., Auer, M.,Ayaz, D., Ayres Fernández, C., Bakiev, A.G., Celani, A.,Džukic, G., Fahd, S., Havaš, P., Joger, U., Khabibullin,V.F., Mazanaeva, L.F., Široký, P., Tripepi, S., ValdeónVélez, A., Velo Antón, G., Wink, M. (2007): Mitochon-drial phylogeography of European pond turtles (Emysorbicularis, Emys trinacris) – an update. Amphib.-Reptil. 28: 418-426.
Fritz, U., Ayaz, D., Buschbom, J., Kami, H.G., Mazanaeva,L.F., Aloufi, A.A., Auer, M., Rifai, L., Šilic, T., Hunds-dörfer, A.K. (2008): Go east: phylogeographies of Mau-remys caspica and M. rivulata – discordance of mor-phology, mitochondrial and nuclear genomic markersand rare hybridization. J. Evolution. Biol. 21: 527-540.
Fritz, U., Ayaz, D., Hundsdörfer, A.K., Kotenko, T., Guick-ing, D., Wink, M., Tok, C.V., Çiçek, K., Buschbom,J. (2009): Mitochondrial diversity of European pondturtles (Emys orbicularis) in Anatolia and the Ponto-Caspian Region: multiple old refuges, hotspot of extantdiversification and critically endangered endemics. Org.Divers. Evol. 9: 100-114.
Fritz, U., Alcalde, L., Vargas-Ramírez, M., Goode, E.V.,Fabius-Turoblin, D.U., Praschag, P. (2012a): Northerngenetic richness and southern purity, but just one speciesin the Chelonoidis chilensis complex. Zool. Scr. 41: 220-232.
Fritz, U., Stuckas, H., Vargas-Ramírez, M., Hundsdörfer,A.K., Maran, J., Päckert, M. (2012b): Molecular phy-logeny of Central and South American slider turtles:implications for biogeography and systematics (Tes-tudines: Emydidae: Trachemys). J. Zool. Syst. Evol. Res.50: 125-136.
Fujita, M.F., Engstrom, T.N., Starkey, D.E., Shaffer, H.B.(2004): Turtle phylogeny: insights from a novel nuclearintron. Mol. Phylogenet. Evol. 31: 1031-1040.
Gaffney, E.S., Tong, H., Meylan, P.A. (2006): Evolutionof the side-necked turtles: the families Bothremydidae,Euraxemydidae, and Araripemydidae. Bull. Am. Mus.Nat. Hist. 300: 1-700.
Georges, A., Birrel, J., Saint, K.M., McCord, W.P., Don-nellan, S.C. (1998): A phylogeny for side-necked tur-tles (Chelonia: Pleurodira) based on mitochondrial andnuclear gene sequence variation. Biol. J. Linn. Soc. 67:213-246.
González-Porter, G.P., Hailer, F., Flores-Villela, O., García-Anleu, R., Maldonado, J.E. (2011): Patterns of geneticdiversity in the critically endangered Central Americanriver turtle: human influence since the Mayan age? Con-serv. Genet. 12: 1229-1242.
Hall, T.A. (1999): BIOEDIT: a user-friendly biological se-quence alignment editor and analysis program for Win-dows 95/98/NT. Nucl. Acids Symp. Ser. 4: 95-98.
Hebert, P.D.N., Ratnasingham, S., de Waard, J.R. (2003):Barcoding animal life: cytochrome c oxidase subunit 1divergences among closely related species. Proc. R. Soc.Lond. B 270: S96-S99.
IUCN [International Union for the Conservation of Na-ture and Natural Resources] (2011): IUCN Red List ofThreatened Species 2011.2. http://www.iucnredlist.org[accessed 22 February 2012].
Karl, S.A., Wilson, D.S. (2001): Phylogeography and sys-tematics of the mud turtle, Kinosternon baurii. Copeia2001: 797-801.
Kellogg, J.N. (1984): Cenozoic tectonic history of the Sierrade Perijá, Venezuela-Colombia, and adjacent basins.Geol. Soc. Am. Mem. 162: 239-261.
Kindler, C., Branch, W.R., Hofmeyr, M.D., Maran, J.,Široký, P., Vences, M., Harvey, J., Hauswaldt, J.S.,Schleicher, A., Stuckas, H., Fritz, U. (2012): Molecu-lar phylogeny of African hinge-back tortoises (Kinixys):implications for phylogeography and taxonomy (Tes-tudines: Testudinidae). J. Zool. Syst. Evol. Res. 50: 192-201.
Kocher, T.D., Thomas, W.K., Meyer, A., Edwards, S.V.,Pääbo, S., Villablanca, F.X., Wilson, A.C. (1989): Dy-namics of mitochondrial DNA evolution in mammals:amplification and sequencing with conserved primers.PNAS 86: 6196-6200.
Lamb, T., Lydeard, C., Walker, R.B., Gibbons, J.W. (1994):Molecular systematics of map turtles (Graptemys):a comparison of mitochondrial restriction site versus se-quence data. Syst. Biol. 43: 543-559.
Le, M., Raxworthy, C.J., McCord, W.P., Mertz, L. (2006):A molecular phylogeny of tortoises (Testudines: Testu-dinidae) based on mitochondrial and nuclear genes. Mol.Phylogenet. Evol. 40: 517-531.
Lenk, P., Fritz, U., Joger, U., Wink, M. (1999): Mitochon-drial phylogeography of the European pond turtle, Emysorbicularis (Linnaeus 1758). Mol. Ecol. 8: 1911-1922.
Li, C., Wu, X.-C., Rieppel, O., Wang, L.-T., Zhao, L.-J.(2008): An ancestral turtle from the Late Triassic ofsouthwestern China. Nature 456: 497-501.
Librado, P., Rozas, J. (2009): DNASP v5: a software forcomprehensive analysis of DNA polymorphism data.Bioinformatics 25: 1451-1452.
Medem, F. (1966): Contribuciones al conocimiento sobre laecología y distribución geográfica de Phrynops (Batra-chemys) dahli (Testudinata, Pleurodira, Chelidae). Cal-dasia 9: 467-489.
Pearse, D.E., Arndt, A.D., Valenzuela, N., Miller, B.A.,Cantarelli, V., Sites, J.R. (2006): Estimating populationstructure under nonequilibrium conditions in a conserva-tion context: continent-wide population genetics of thegiant Amazon River turtle, Podocnemis expansa (Chelo-nia; Podocnemididae). Mol. Ecol. 15: 985-1006.
Pedall, I., Fritz, U., Stuckas, H., Valdéon, A., Wink, M.(2011): Gene flow across secondary contact zones of theEmys orbicularis complex in the Western Mediterraneanand evidence for extinction and re-introduction of pondturtles on Corsica and Sardinia (Testudines: Emydidae).J. Zool. Syst. Evol. Res. 49: 44-57.
Praschag, P., Hundsdörfer, A.K., Fritz, U. (2007): Phy-logeny and taxonomy of endangered South and South-east Asian freshwater turtles elucidated by mtDNA se-quence variation (Testudines: Geoemydidae: Batagur,Callagur, Hardella, Kachuga, Pangshura). Zool. Scr.36: 429-442.
Praschag, P., Stuckas, H., Päckert, M., Maran, J., Fritz, U.(2011): Mitochondrial DNA sequences suggest a revisedtaxonomy of Asian flapshell turtles (Lissemys Smith,1931) and the validity of previously unrecognized taxa(Testudines: Trionychidae). Vertebr. Zool. 61: 147-160.
Pritchard, P.C.H., Trebbau, P. (1984): The Turtles ofVenezuela. Contributions to Herpetology, 2. Society forthe Study of Amphibians and Reptiles, Oxford, Ohio.
Rojas-Runjaic, F.J.M. (2009): Mesoclemmys zuliae, el enig-mático galápago “cabezón” del sur del lago Maracaibo.Reptilia 80: 31-38.
Rosenbaum, P.A., Robertson, J.A., Zamudio, K.R. (2007):Unexpectedly low genetic divergences among popula-tions of the threatened bog turtle (Glyptemys muhlen-bergii). Conserv. Genet. 8: 331-342.
Rueda-Almonacid, J.V., Carr, J.L., Mittermeier, R.A.,Rodríguez-Mahecha, J.V., Mast, R.B., Vogt, R.C.,Rhodin, A.G.J., de la Ossa-Velásquez, J., Rueda, J.N.,Goettsch-Mittermeier, C. (2007): Las Tortugas y loscocodrilianos de los países andinos del trópico. Bogotá,Conservación Internacional (Serie de guías tropicales decampo).
Silvestro, D., Michalak, I. (2011): raxmlGUI: a graphicalfrontend for RAxML. Org. Divers. Evol., DOI:10.1007/s13127-011-0056-0.
Souza, F.L. (2005): Geographical distribution patterns ofSouth American side-necked turtles (Chelidae), withemphasis on Brazilian species. Rev. Esp. Herpetol. 19:33-46.
Souza, F.L., Cunha, A.F., Oliveira, M.A., Pereira, G.A.G.,dos Reis, S.F. (2003): Preliminary phylogeographicanalysis of the Neotropical freshwater turtle Hydrome-dusa maximiliani (Chelidae). J. Herpetol. 37: 427-433.
Stamatakis, A. (2006): RAxML-VI-HPC: maximumlikelihood-based phylogenetic analyses with thou-sands of taxa and mixed models. Bioinformatics 22:2688-2690.
Stuckas, H., Fritz, U. (2011): Identity of Pelodiscus sinensisrevealed by DNA sequences of an approximately 180-year-old type specimen and a taxonomic reappraisal ofPelodiscus species (Testudines: Trionychidae). J. Zool.Syst. Evol. Res. 49: 335-339.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei,M., Kumar, S. (2011): MEGA 5: Molecular Evolution-ary Genetics Analysis using maximum likelihood, evo-lutionary distance, and maximum parsimony methods.Mol. Biol. Evol. 28: 2731-2739.
Thorne, J.L., Kishino, H. (2002): Divergence time andevolutionary rate estimation with multilocus data. Syst.Biol. 51: 689-702.
Thorne, J.L., Kishino, H., Painter, I.S. (1998): Estimatingthe rate of evolution of the rate of molecular evolution.Mol. Biol. Evol. 15: 1647-1657.
Ureña-Aranda, C.A., Espinosa de los Monteros, A. (2012):The genetic crisis of the Mexican Bolson tortoise (Go-pherus flavomarginatus: Testudinidae). Amphib.-Reptil.33: 45-53.
van Dijk, P.P., Iverson, J.B., Shaffer, H.B., Bour, R., Rhodin,A.G.J. (2011): Turtles of the world, 2011 update: anno-tated checklist of taxonomy, synonymy, distribution, andconservation status. Chelon. Res. Monogr. 5: 000.165-000.242.
Vargas-Ramírez, M., Maran, J., Fritz, U. (2010a): Red-and yellow-footed tortoises (Chelonoidis carbonaria, C.denticulata) in South American savannahs and forests:do their phylogeographies reflect distinct habitats? Org.Divers. Evol. 10: 161-172.
Vargas-Ramírez, M., Vences, M., Branch, W.R., Daniels,S.R., Glaw, F., Hofmeyr, M.D., Kuchling, G., Maran,J., Papenfuss, T.J., Široký, P., Vieites, D.R., Fritz, U.(2010b): Deep genealogical lineages in the widely dis-tributed African helmeted terrapin: evidence from mi-tochondrial and nuclear DNA (Testudines: Pelomedusi-dae: Pelomedusa subrufa). Mol. Phylogenet. Evol. 56:428-440.
Vargas-Ramírez, M., Stuckas, H., Castaño-Mora, O.V.,Fritz, U. (2012): Extremely low genetic diversityand weak population differentiation in the endangeredColombian river turtle Podocnemis lewyana (Testudines:Podocnemididae). Conserv. Genet. 13: 65-77.
Walker, D., Moler, P.E., Buhlmann, K.A., Avise, J.C.(1998): Phylogeographic uniformity in mitochondrialDNA of the snapping turtle (Chelydra serpentina).Anim. Conserv. 1: 55-60.
Weisrock, D.W., Janzen, F.J. (2000): Comparative molecu-lar phylogeography of North American softshell turtles(Apalone): implications for regional and wide-scale his-torical evolutionary forces. Mol. Phylogenet. Evol. 14:152-164.
Weak genetic divergence between turtles Mesoclemmys dahli and M. zuliae (Testudines: Pleurodira: Chelidae) 383
Yang, Z. (1997): PAML: a program package for phyloge-netic analysis by maximum likelihood. Comput. Appl.Biosci. 13: 555-556.
Zangerl, R., Medem, F. (1958): A new species of chelidturtle, Phrynops (Batrachemys) dahli, from Colombia.Bull. Mus. Comp. Zool. 119: 373-390.
Submitted: February 28, 2012. Final revision received: July25, 2012. Accepted: July 30, 2012.Associated Editor: Sylvain Ursenbacher.
384 M. Vargas-Ramírez et al.A
ppen
dix
Tabl
eA
.1.M
esoc
lem
mys
sam
ples
and
outg
roup
sus
edin
the
pres
ent
stud
y.M
TD
Tnu
mbe
rsre
fer
tosa
liva
ortis
sue
sam
ples
inth
eco
llect
ion
ofth
eM
useu
mof
Zoo
logy
,Se
ncke
nber
gD
resd
en.
Spec
ies
Loc
ality
mtD
NA
hapl
otyp
eG
enB
ank
acce
ssio
nnu
mbe
rsM
TD
T
cytb
12S
rRN
AN
D4
C-m
osR
35R
ag2
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)A
JX13
9039
––
––
–46
94M
esoc
lem
mys
dahl
iA
rach
e,C
him
á,C
órdo
ba,C
olom
bia
(9°4
′ 32.
80′′ N
75°3
8′37
.25′′
W)
AJX
1390
40–
––
––
4695
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)A
JX13
9041
––
––
–47
12M
esoc
lem
mys
dahl
iA
rach
e,C
him
á,C
órdo
ba,C
olom
bia
(9°4
′ 32.
80′′ N
75°3
8′37
.25′′
W)
AJX
1390
42–
––
––
4714
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)A
JX13
9043
––
––
–47
11M
esoc
lem
mys
dahl
iM
omíl,
Cór
doba
,Col
ombi
a(9
°14′
53.3
7′′N
75°4
0′26
.33′′
W)
BJX
1390
44–
––
––
4696
Mes
ocle
mm
ysda
hli
Serr
ader
o,Pu
risi
ma,
Cór
doba
,B
JX13
9045
––
––
–46
98C
olom
bia
(9°1
4′33
.17′′
N75
°43′
0.27
′′ W)
Mes
ocle
mm
ysda
hli
La
Con
fianz
a,L
oric
a,C
órdo
ba,
CJX
1390
46–
––
––
4697
Col
ombi
a(9
°14′
14.2
9′′N
75°4
8′30
.13′′
W)
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)D
JX13
9047
JX13
9086
JX13
9071
JX13
9082
JX13
9078
JX13
9073
4699
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)E
JX13
9048
––
––
–47
00M
esoc
lem
mys
dahl
iA
rach
e,C
him
á,C
órdo
ba,C
olom
bia
(9°4
′ 32.
80′′ N
75°3
8′37
.25′′
W)
EJX
1390
49–
––
––
4703
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)E
JX13
9050
––
––
–47
10M
esoc
lem
mys
dahl
iA
rach
e,C
him
á,C
órdo
ba,C
olom
bia
(9°4
′ 32.
80′′ N
75°3
8′37
.25′′
W)
FJX
1390
51–
––
––
4701
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)G
JX13
9052
––
––
–47
05M
esoc
lem
mys
dahl
iA
rach
e,C
him
á,C
órdo
ba,C
olom
bia
(9°4
′ 32.
80′′ N
75°3
8′37
.25′′
W)
HJX
1390
53–
––
––
4706
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)H
JX13
9054
––
––
–47
09M
esoc
lem
mys
dahl
iA
rach
e,C
him
á,C
órdo
ba,C
olom
bia
(9°4
′ 32.
80′′ N
75°3
8′37
.25′′
W)
HJX
1390
55–
––
––
4715
Mes
ocle
mm
ysda
hli
Ara
che,
Chi
má,
Cór
doba
,Col
ombi
a(9
°4′ 3
2.80
′′ N75
°38′
37.2
5′′W
)I
JX13
9056
––
––
–47
07M
esoc
lem
mys
dahl
iC
him
icha
gua,
Cie
naga
deZ
apat
oza,
Ces
ar,
JJX
1390
57–
––
––
9147
Col
ombi
a(9
°17′
18.6
0′′N
73°4
7′35
.80′′
W)
Mes
ocle
mm
ysda
hli
Chi
mic
hagu
a,C
iena
gade
Zap
atoz
a,C
esar
,J
JX13
9058
––
––
–91
48C
olom
bia
(9°1
7′18
.60′′
N73
°47′
35.8
0′′W
)M
esoc
lem
mys
dahl
iC
him
icha
gua,
Cie
naga
deZ
apat
oza,
Ces
ar,
KJX
1390
59–
––
––
9149
Col
ombi
a(9
°17′
18.6
0′′N
73°4
7′35
.80′′
W)
Mes
ocle
mm
ysda
hli
Chi
mic
hagu
a,C
iena
gade
Zap
atoz
a,C
esar
,L
JX13
9060
––
––
–91
50C
olom
bia
(9°1
7′18
.60′′
N73
°47′
35.8
0′′W
)M
esoc
lem
mys
dahl
iC
him
icha
gua,
Cie
naga
deZ
apat
oza,
Ces
ar,
MJX
1390
61–
––
––
9151
Col
ombi
a(9
°17′
18.6
0′′N
73°4
7′35
.80′′
W)
Weak genetic divergence between turtles Mesoclemmys dahli and M. zuliae (Testudines: Pleurodira: Chelidae) 385