Page 1
A Review Molecular Typing Methods for Aspergillus flavusIsolates
Ines Hadrich • Fattouma Makni • Sourour Neji •
Fatma Cheikhrouhou • Hayet Sellami •
Ali Ayadi
Received: 25 February 2010 / Accepted: 16 February 2011 / Published online: 3 March 2011
� Springer Science+Business Media B.V. 2011
Abstract Aspergillus flavus is the second most
important Aspergillus species causing human infec-
tions. The importance of this fungus increases in
regions with a dry and hot climate. Small phylogenetic
studies in Aspergillus flavus indicate that the morpho-
logical species contains several genetically isolated
species. Different genotyping methods have been
developed and employed in order to better understand
the genetic and epidemiological relationships between
environmental and clinical isolates. Understanding
pathogen distribution and relatedness is essential for
determining the epidemiology of nosocomial infec-
tions and aiding in the design of rational pathogen
control methods. Typing techniques can also give us a
deeper understanding of the colonization pattern in
patients. Most of these studies focused on Aspergillus
fumigatus because it is medically the most isolated
species. To date, there has not been any publication
exclusively reviewing the molecular typing techniques
for Aspergillus flavus in the literature. This article
reviews all these different available methods for this
organism.
Keywords Molecular typing � RFLP �Microsatellites
Introduction
Human infections due to members of the genus
Aspergillus are recognized with increasing frequency
in immunocompromised patients. The majority
(approximately 80%) of invasive Aspergillus infec-
tions is caused by Aspergillus fumigatus. The second
most frequent pathogenic species is Aspergillus
flavus and to a lesser extent, Aspergillus niger and
Aspergillus terreus [1].
Aspergillus flavus is an ubiquitous mold. This
name is now used to describe a species as well as a
group of closely related species. Climate and geo-
graphical factors are important determinants of the
local prevalence of A. flavus infections. In countries
with semi-arid and arid dry weather conditions like
Saudi Arabia Sudan and Tunisia (Sfax), A. flavus is
the main etiological agent of invasive aspergillosis
[2–5]. It is also able to cause a number of other
diseases in immunocompetent patients such as
chronic indolent invasive sinonasal infection, kerati-
tis, otitis, and onychomycosis [6, 7]. Incrimination of
I. Hadrich (&) � F. Makni � S. Neji � F. Cheikhrouhou �H. Sellami � A. Ayadi (&)
Faculte de medecine, Laboratoire de biologie moleculaire
parasitaire et fongique, 3029 Sfax, Tunisie
e-mail: [email protected]
A. Ayadi
Fungal and parasitic molecular biology laboratory,
School of Medicine Sfax, Sfax, Tunisia
e-mail: [email protected]
123
Mycopathologia (2011) 172:83–93
DOI 10.1007/s11046-011-9406-x
Page 2
different environmental sources of Aspergillus infec-
tion has always been discussed [8–10].
The identification of A. flavus is not straightfor-
ward due to similarities with closely related species
(e.g., A. parasiticus and A. nomius). Aspergillus
subgenus Circumdati section Flavi, also referred to as
the Aspergillus flavus group, has attracted worldwide
attention for its industrial use and toxigenic potential.
Section Flavi is divided in two groups of species. One
includes the aflatoxigenic species A. flavus, A. para-
siticus, A. parvisclerotigenus, A. minisclerotigenes,
A. arachidicola, and A. nomius, which cause serious
problems worldwide in agricultural commodities, and
the other includes the non-aflatoxigenic species
A. oryzae, A. sojae, and A. tamarii, traditionally used
for the production of fermented foods [11]. Recently,
multiple molecular typing methods have been devel-
oped to trace the spread of particular subspecific
strains. Various DNA fingerprinting systems have
been described, for example, multilocus enzyme
electrophoresis [12], restriction fragment length
polymorphisms of total DNA [13–15], hybridization
of endonuclease digested DNA with DNA probes
[8, 16], interrepeat polymerase chain reaction [17],
and random amplification of polymorphic DNA
[18, 19]. Polymorphic microsatellite marker analyses
have been developed to delineate strains of Asper-
gillus species [20, 21]. In this review, we will discuss
the molecular tools for genotyping analyses of
A. flavus isolates to understand the epidemiology of
this opportunistic pathogenic fungus.
Random Amplification of Polymorphic DNA
(RAPD)
Using random primers of approximately 10 bases,
amplicons throughout the genome are targeted and
amplified. Amplified products are subsequently sep-
arated on an agarose gel and stained with ethidium
bromide. The genetic variation analysis based on
RAPD allows proper genetic diversity due to its
capacity to generate random markers from the entire
genome.
Diaz-Guerra TM et al. [22] have used RAPD with
three primers (R-108, R-151 and AP12 h) for typing
11 A. flavus isolates and they have found a satisfac-
tory discriminatory power. In one case, a genetic
similarity was noted between strains isolated from
patients and strains from the grilles of a dual-
reservoir cooler–heater used in the operating room
confirming the nosocomial origin of contamination
(Table 1).
Myoken Y et al. [23] have used RAPD with three
different PCR primers for typing 6 A. flavus isolates
obtained from leukemic patients with invasive
Aspergillus stomatitis. The molecular analysis
revealed that three isolates of A. flavus obtained from
three patients have the same type (Table 1).
A case of A. flavus nosocomial infection after
sternal surgery has been reported. RAPD with two
different primers (ERIC- 1, BG-2) to type A. flavus
strains from patients and from the hospital environ-
ment was used. All these strains showed the same
genotype, proving the clonal single-source of the
environmental contamination and the intraoperative
acquisition of A. flavus in the sternal surgical-site
infections outbreak (Table 1) [24].
The use of RAPD has allowed to differentiate
species of the A. flavus complex. This approves the
role of taxonomic studies. Batista PP et al. have used
six random primers giving a RAPD profile with very
different products for each A. flavus strain providing
evidence of its high genetic diversity. The primer
OPW-04 revealed low intraspecific variability and
high interspecific variability [21]. Among some strains
previously classified as A. flavus, one was reclassified
as A. oryzae, one as A. parasiticus, and two as
A. tamarii. But also one strain previously identified as
A. parasiticus was reclassified as A. flavus [21]. This
was similar to the report of Yuan et al. [25] who used
RAPD to differentiate two morphologically similar
species A parasiticus and A. sojae and who has
reclassified one strain as A. flavus.
The major problem in RAPD typing is a lack of
reproducibility of patterns (variation of number, size,
and intensity of bands) [15, 26] and the interlabora-
tory reproducibility.
Restriction Fragment Length Polymorphism
(RFLP)
The basic technique for detecting RFLPs involves
fragmenting a sample of DNA by a restriction
enzyme that can recognize and cut DNA wherever
a specific short sequence occurs in a process known
as a restriction digest. The resulting DNA fragments
84 Mycopathologia (2011) 172:83–93
123
Page 3
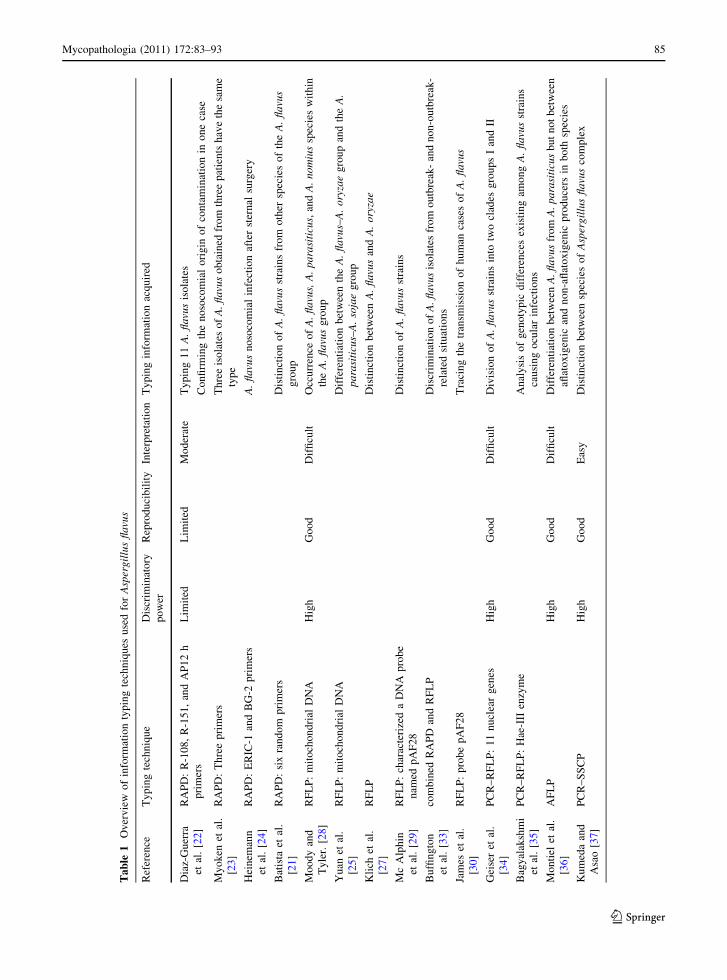
Ta
ble
1O
ver
vie
wo
fin
form
atio
nty
pin
gte
chn
iqu
esu
sed
for
Asp
erg
illu
sfl
avu
s
Ref
eren
ceT
yp
ing
tech
niq
ue
Dis
crim
inat
ory
po
wer
Rep
rod
uci
bil
ity
Inte
rpre
tati
on
Ty
pin
gin
form
atio
nac
qu
ired
Dia
z-G
uer
ra
etal
.[2
2]
RA
PD
:R
-10
8,
R-1
51
,an
dA
P1
2h
pri
mer
s
Lim
ited
Lim
ited
Mo
der
ate
Ty
pin
g1
1A
.fl
avu
sis
ola
tes
Co
nfi
rmin
gth
en
oso
com
ial
ori
gin
of
con
tam
inat
ion
ino
ne
case
My
ok
enet
al.
[23
]
RA
PD
:T
hre
ep
rim
ers
Th
ree
iso
late
so
fA
.fl
avu
so
bta
ined
fro
mth
ree
pat
ien
tsh
ave
the
sam
e
typ
e
Hei
nem
ann
etal
.[2
4]
RA
PD
:E
RIC
-1an
dB
G-2
pri
mer
sA
.fl
avu
sn
oso
com
ial
infe
ctio
naf
ter
ster
nal
surg
ery
Bat
ista
etal
.
[21
]
RA
PD
:si
xra
nd
om
pri
mer
sD
isti
nct
ion
of
A.
fla
vus
stra
ins
fro
mo
ther
spec
ies
of
the
A.
fla
vus
gro
up
Mo
od
yan
d
Ty
ler.
[28]
RF
LP
:m
ito
cho
nd
rial
DN
AH
igh
Go
od
Dif
ficu
ltO
ccu
rren
ceo
fA
.fl
avu
s,A
.p
ara
siti
cus,
and
A.
no
miu
ssp
ecie
sw
ith
in
the
A.
fla
vus
gro
up
Yu
anet
al.
[25
]
RF
LP
:m
ito
cho
nd
rial
DN
AD
iffe
ren
tiat
ion
bet
wee
nth
eA
.fl
avu
s–A
.o
ryza
eg
rou
pan
dth
eA
.p
ara
siti
cus–
A.
soja
eg
rou
p
Kli
chet
al.
[27
]
RF
LP
Dis
tin
ctio
nb
etw
een
A.
fla
vus
and
A.
ory
zae
Mc
Alp
hin
etal
.[2
9]
RF
LP
:ch
arac
teri
zed
aD
NA
pro
be
nam
edp
AF
28
Dis
tin
ctio
no
fA
.fl
avu
sst
rain
s
Bu
ffin
gto
n
etal
.[3
3]
com
bin
edR
AP
Dan
dR
FL
PD
iscr
imin
atio
no
fA
.fl
avu
sis
ola
tes
fro
mo
utb
reak
-an
dn
on
-ou
tbre
ak-
rela
ted
situ
atio
ns
Jam
eset
al.
[30
]
RF
LP
:p
rob
ep
AF
28
Tra
cin
gth
etr
ansm
issi
on
of
hu
man
case
so
fA
.fl
avu
s
Gei
ser
etal
.
[34
]
PC
R–
RF
LP
:1
1n
ucl
ear
gen
esH
igh
Go
od
Dif
ficu
ltD
ivis
ion
of
A.
fla
vus
stra
ins
into
two
clad
esg
rou
ps
Ian
dII
Bag
yal
aksh
mi
etal
.[3
5]
PC
R–
RF
LP
:H
ae-I
IIen
zym
eA
nal
ysi
so
fg
eno
typ
icd
iffe
ren
ces
exis
tin
gam
on
gA
.fl
avu
sst
rain
s
cau
sin
go
cula
rin
fect
ion
s
Mo
nti
elet
al.
[36
]
AF
LP
Hig
hG
oo
dD
iffi
cult
Dif
fere
nti
atio
nb
etw
een
A.
fla
vus
fro
mA
.p
ara
siti
cus
bu
tn
ot
bet
wee
n
aflat
ox
igen
ican
dn
on
-afl
ato
xig
enic
pro
du
cers
inb
oth
spec
ies
Ku
med
aan
d
Asa
o[3
7]
PC
R–
SS
CP
Hig
hG
oo
dE
asy
Dis
tin
ctio
nb
etw
een
spec
ies
of
Asp
erg
illu
sfl
avu
sco
mp
lex
Mycopathologia (2011) 172:83–93 85
123
Page 4
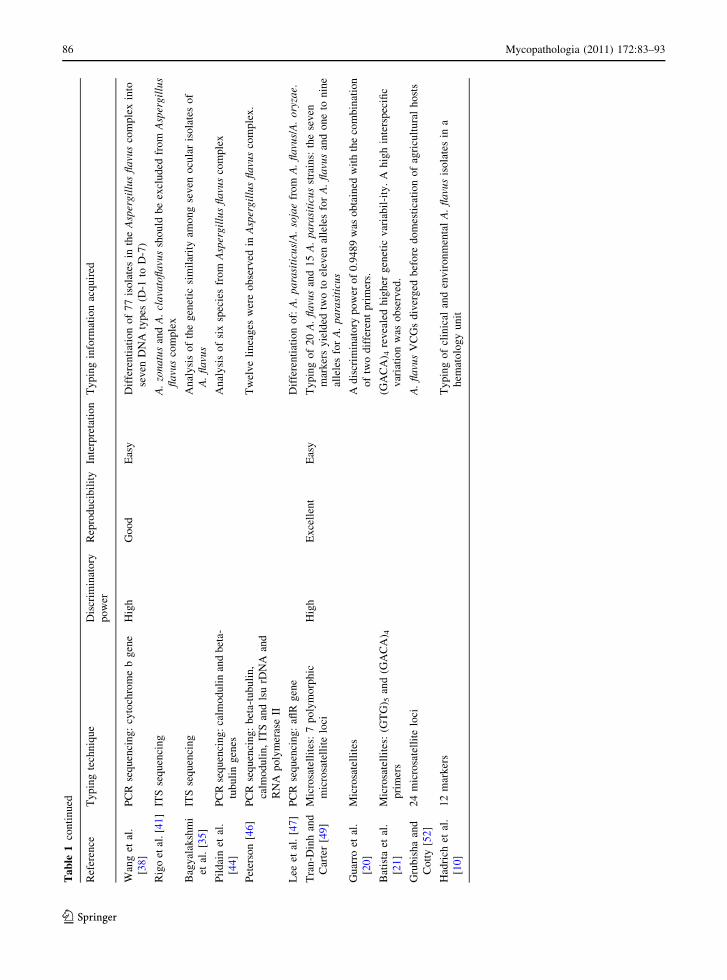
Ta
ble
1co
nti
nu
ed
Ref
eren
ceT
yp
ing
tech
niq
ue
Dis
crim
inat
ory
po
wer
Rep
rod
uci
bil
ity
Inte
rpre
tati
on
Ty
pin
gin
form
atio
nac
qu
ired
Wan
get
al.
[38
]
PC
Rse
qu
enci
ng
:cy
toch
rom
eb
gen
eH
igh
Go
od
Eas
yD
iffe
ren
tiat
ion
of
77
iso
late
sin
the
Asp
erg
illu
sfl
avu
sco
mp
lex
into
sev
enD
NA
typ
es(D
-1to
D-7
)
Rig
oet
al.[4
1]
ITS
seq
uen
cin
gA
.zo
na
tus
and
A.
cla
vato
fla
vus
sho
uld
be
excl
ud
edfr
om
Asp
erg
illu
sfl
avu
sco
mp
lex
Bag
yal
aksh
mi
etal
.[3
5]
ITS
seq
uen
cin
gA
nal
ysi
so
fth
eg
enet
icsi
mil
arit
yam
on
gse
ven
ocu
lar
iso
late
so
f
A.
fla
vus
Pil
dai
net
al.
[44
]
PC
Rse
qu
enci
ng
:ca
lmo
du
lin
and
bet
a-
tub
uli
ng
enes
An
aly
sis
of
six
spec
ies
fro
mA
sper
gil
lus
fla
vus
com
ple
x
Pet
erso
n[4
6]
PC
Rse
qu
enci
ng
:b
eta-
tub
uli
n,
calm
od
uli
n,
ITS
and
lsu
rDN
Aan
d
RN
Ap
oly
mer
ase
II
Tw
elv
eli
nea
ges
wer
eo
bse
rved
inA
sper
gil
lus
fla
vus
com
ple
x.
Lee
etal
.[4
7]
PC
Rse
qu
enci
ng
:afl
Rg
ene
Dif
fere
nti
atio
no
f:A
.p
ara
siti
cus/
A.
soja
efr
om
A.
fla
vus/
A.
ory
zae.
Tra
n-D
inh
and
Car
ter
[49
]
Mic
rosa
tell
ites
:7
po
lym
orp
hic
mic
rosa
tell
ite
loci
Hig
hE
xce
llen
tE
asy
Ty
pin
go
f2
0A
.fl
avu
san
d1
5A
.p
ara
siti
cus
stra
ins:
the
sev
en
mar
ker
sy
ield
edtw
oto
elev
enal
lele
sfo
rA
.fl
avu
san
do
ne
ton
ine
alle
les
for
A.
pa
rasi
ticu
s
Gu
arro
etal
.
[20
]
Mic
rosa
tell
ites
Ad
iscr
imin
ato
ryp
ow
ero
f0
.94
89
was
ob
tain
edw
ith
the
com
bin
atio
n
of
two
dif
fere
nt
pri
mer
s.
Bat
ista
etal
.
[21
]
Mic
rosa
tell
ites
:(G
TG
) 5an
d(G
AC
A) 4
pri
mer
s
(GA
CA
) 4re
vea
led
hig
her
gen
etic
var
iab
il-i
ty.
Ah
igh
inte
rsp
ecifi
c
var
iati
on
was
ob
serv
ed.
Gru
bis
ha
and
Co
tty
[52
]
24
mic
rosa
tell
ite
loci
A.
fla
vus
VC
Gs
div
erg
edb
efo
red
om
esti
cati
on
of
agri
cult
ura
lh
ost
s
Had
rich
etal
.
[10
]
12
mar
ker
sT
yp
ing
of
clin
ical
and
env
iro
nm
enta
lA
.fl
avu
sis
ola
tes
ina
hem
ato
log
yu
nit
86 Mycopathologia (2011) 172:83–93
123
Page 5
are then separated by length by agarose gel electro-
phoresis and transferred to a membrane via the
Southern blot procedure. Hybridization of the mem-
brane to a labeled DNA probe determines the length
of the fragments that are complementary to the probe.
A RFLP occurs when the length of a detected
fragment varies between individuals. Each profile is
considered an allele and can be used in genetic
analysis.
RFLP has been used to distinguish between two
related species A. flavus and A. oryzae and to analyze
their phylogenetic relationships [27].
The characterization of mitochondrial DNA is a
useful adjunct to the standard morphological and
physiological characteristics to determine the taxo-
nomic status of isolates in Aspergillus flavus com-
plex. Moody et al. [28] identified mitochondrial
DNA RFLP and used them to propose the occurrence
of species within the A. flavus group: A. flavus,
A. parasiticus, and A. nomius (Table 1).
Mc Alphin et al. [29] constructed and character-
ized a DNA probe named pAF28 for distinguishing
strains of A. flavus (Table 1). This probe has been
used to type clinical and environmental strains
isolated from a neonatal intensive care unit [30].
James M et al. [30] demonstrated that DNA finger-
printing with the pAF28 repetitive probe is a highly
reproducible and discriminatory method for tracing
the transmission of human cases of A. flavus infection
expanding its utility beyond the agricultural purpose
for which it was developed (Table 1). These results
are consistent with the successful findings obtained
with the AfutI repetitive DNA sequence probe in
investigations of the nosocomial transmission of
A. fumigatus infection [31, 32].
Buffington J et al. [33] used RFLP analysis of Sma
I-digested DNA to discriminate A. flavus isolates
from outbreak- and non-outbreak-related situations
(Table 1). They combined the products from RAPD
and RFLP analysis of a tester strain of A. flavus to
produce a DNA probe for Southern blot analysis. So,
a high degree of discrimination among strain types
was achieved.
Although, complex RFLP patterns can be difficult
to interpret, RFLP remains a reproducible and high
discriminatory system.
Geiser et al. [34] have conducted PCR–RFLP
analysis of 11 nuclear genes and concluded that a
collection of A. flavus, A. parasiticus, and A. oryzae
strains could be divided into two clades (groups I and
II) with group I comprising isolates of A. oryzae
(Table 1).
The PCR–RFLP with Hae-III enzyme has been
used to analyze the genotypic differences existing
among A. flavus strains causing ocular infections.
This method showed the same profile of bands for all
isolates [35]. This approach is generally suitable for
discriminating among different species, but has
insufficient discriminatory power to distinguish
among unrelated isolates within a species.
Amplified Fragment Length Polymorphism
(AFLP)
This method combines the principle of the RFLP
analysis with highly specific PCR amplification.
Genomic DNA is usually cut with two restriction
enzymes, one with an average cutting frequency and
a second one with a higher cutting frequency, and
double-stranded adapters are ligated to the ends of the
DNA fragments to generate template DNA for
amplification. The sequence of the adapters and the
adjacent restriction site serve as primer binding sites
for subsequent amplification of the restriction frag-
ments. Selective nucleotides are included at the 30
ends of the PCR primers, which therefore can only
prime DNA synthesis from a subset of the restriction
sites. Only restriction fragments in which the nucle-
otides flanking the restriction site match the selective
nucleotides will be amplified. Variations between
different isolates originate by differences in the
number and the location of restriction enzyme
recognition sites in the genome.
AFLPs were used to explore genetic diversity
among twenty-four isolates of Aspergillus flavus
complex using twelve selective primer combinations.
This study demonstrated that AFLP can be an
excellent typing method to differentiate between
isolates of Aspergillus flavus complex [36]. It should
be generally useful in distinguishing between closely
related species or strains. It revealed a clear separa-
tion of A. flavus from A. parasiticus, and no
genotypic differences between aflatoxigenic and
non-aflatoxigenic producers could be detected
(Table 1) [36].
Mycopathologia (2011) 172:83–93 87
123
Page 6
PCR–Single-Strand Conformation Polymorphism
(PCR–SSCP)
DNA material is denatured to single-stranded DNA
which is subjected to polyacrylamide gel electropho-
resis. The mobility of single-stranded DNA in the gel
is dependent on its secondary structure as determined
by the nucleotide sequence.
Kumeda Y and Asao T [37] optimized a PCR with
universal primers (ITS1 and ITS4) and a SSCP
conditions. PCR–SSCP analysis offers a reliable
method to distinguish between species of Aspergillus
flavus complex and is more simple and rapid than
some other methods based on DNA hybridization.
Non-radiolabeled PCR–SSCP analysis as well as
PCR–RFLP analysis is practical to perform without
any special apparatus or skill and should assist in
fungal morphological identification (Table 1) [37].
PCR Sequencing
PCR was a major breakthrough for molecular marker
research; any genomic region could be amplified and
analyzed in many individuals without the require-
ment for cloning and isolating large amounts of
ultrapure genomic DNA. PCR sequencing involves
the determination of the nucleotide sequence within a
DNA fragment amplified by the PCR, using primers
specific for a particular genomic site. Many genes
were studied for typing A. flavus isolates such as
cytochrome b gene, the beta-tubulin (BenA), cal-
modulin (CF), RNA polymerase II (RPB2), and aflR
genes.
DNA Sequencing of Cytochrome b Gene
The mitochondrial DNA cytochrome b sequence
analyses provided a reliable tool for classifying some
of the closely related species in Aspergillus flavus
complex [38].
Wang et al. described the use of partial sequences
of the mitochondrial cytochrome b gene (402 bp) to
differentiate 77 isolates in the Aspergillus flavus
complex into seven DNA types (D-1 to D-7): A. sojae
was defined as D-1, A. parasiticus as D-2, A. flavus
and A. oryzae were grouped together as D-4, A.
tamarii was defined as D-5, and A. nomius as D-7.
Furthermore, D-3 was found to be closely related to
A. parasiticus (D-2), also including one strain that
had been deposited as A. flavus var flavus. DNA
type D-6 included one strain that was identified
as A. flavus and was closely related to A. tamarii
(Table 1) [38].
PCR Sequencing of Internal Transcribed Spacer
(ITS) Region
The internal transcribed spacer (ITS) region is
located between the 18S and 28S rRNA genes and
offer distinct advantages over other molecular targets
including high sensitivity of detection due to the
existence of multiple copies per genome. The
sequence variation of ITS region has led to their
use in phylogenetic studies of different fungal
organisms [39]. Single-strand conformation polymor-
phism of ITS regions has also been used as a genetic
approach to differentiate species in the A. flavus
complex [37, 40].
The polymorphism of ITS regions confirmed that
the A. flavus complex seems to comprise distinct
clades [41]. The three main clades (P. alliaceus,
A. flavus, and A. tamarii) could also be distinguished
based on colony color and their ubiquinone system.
Based on ITS sequences, A. robustus, A. caelatus,
A. lanosus, A. albertensis, A. coremiiformis, A. flavo-
furcatis, A. toxicarius, A. terricola var indica,
and A. terricola were all located in Aspergillus
flavus complex. In addition, A. pseudotamarii and
A. bombycis were found to be closely related to
A. caelatus and A. nomius, respectively (Table 1) [41].
Rigo et al. [41] suggested that A. zonatus and A.
clavatoflavus should be excluded from Aspergillus
flavus complex (Table 1). This suggestion was pre-
viously made by Kozakiewicz and based on scanning
electron microscopic studies [42]. Recently, Frisvad
et al. found that A. toxicarius resembles A. parasit-
icus but differs in at least three nucleotide differences
in the ITS regions. Usually, the presence of three or
more sequence differences in ITS regions is an
indication of a different species [43]. A. zhaoqingen-
sis was considered the same as A. nomius in this study
[43].
Bagyalakshmi et al. used a PCR sequencing of ITS
regions to analyze the genetic similarity among seven
ocular isolates of A. flavus. The alignment of
contiguous fungal sequences demonstrated that both
single-nucleotide differences and short lengths of
88 Mycopathologia (2011) 172:83–93
123
Page 7
sequence diversity due to insertion or deletion existed
in the ITS regions among the pathogenic A. flavus
strains (Table 1) [35].
Combined Gene Sequencing
Pildain MB et al. analyzed six species from Asper-
gillus flavus complex isolated from Argentinean
peanuts: A. caelatus, A. flavus, A. tamari, A. parasit-
icus, A. parvisclerotigenus, and A. parasiticus. For
the molecular analysis, two regions of the genome
were analyzed, namely parts of the calmodulin and
beta-tubulin genes [44]. Most of the sequenced
Argentinean isolates fell into one of two main clades,
represented by A. flavus and A. parasiticus. Six
Argentinean isolates from peanuts seeds formed
a well-defined clade related to A. flavus and
A. parvisclerotigenus on trees based on beta-tubulin
and calmodulin sequence data (Table 1). Further-
more, these Argentinean isolates belong to the same
vegetative compatibility groups (VCG) as described
by Pildain et al. [44]. This clade also includes four
isolates assigned to A. flavus group II by Geiser et al.
[34, 45] and three isolates producing small sclerotia
collected from soils from Australia. The calmodu-
lin and beta-tubulin sequence data indicate that
A. oryzae, A.thomii, A. kambarensis, A. asciculatus,
and A. subolivaceus are very closely related or
synonymous with A. flavus.
A total of 460 Aspergillus isolates were analyzed
by DNA sequencing of 4 loci: the beta-tubulin
(BenA), calmodulin (CF), ITS, and lsu rDNA and
RNA polymerase II (RPB2). Twelve lineages were
observed in Aspergillus flavus complex. The A. flavus
lineage included the ex-type cultures of A. oryzae,
A. flavus var columnaris, and A. thomii. One species
often held to be synonymous with A. flavus,
A. subolivaceus, was supported as a separate lineage.
The A. parasiticus lineage included the type strain of
A. terricola var. americana and an isolate of A. sojae.
The A. tamarii lineage included the type isolates of
A. flavofurcatis and A. terricola (Table 1) [46].
PCR Sequencing of the aflR Gene
Lee CZ et al. examined 34 strains of Aspergillus
flavus complex. The sequenced aflR genes from the
23 positive strains had greater than 96.6% similarity.
It was particularly conserved in the zinc-finger
DNA-binding domain. The aflR gene of A. sojae
has two obvious characteristics: an extra CTCATG
sequence fragment and a C to T transition that causes
premature termination of AFLR protein synthesis.
Differences between A. parasiticus/A. sojae and
A. flavus/A. oryzae aflR genes were also identified [47].
A detailed comparison of the aflR gene sequences
demonstrated that certain base variations can be used
to differentiate A. parasiticus/A. sojae from A. flavus/
A. oryzae. These differences comprise ten transitions,
three transversions, and one deletion (Table 1) [47].
Microsatellites
Microsatellites or short tandem repeats (STRs) are
short repetitive sequences that are abundantly present
in the genomes of most of the higher organisms and
to a lesser extent in several prokaryotic genomes [48].
Different isolates can be distinguished from each
other based on differences in repeat numbers.
Microsatellite markers are easily amplified by PCR
using primers based on their flanking sequences. If
one of the primers is fluorescently labeled they can be
sized very accurately using high-resolution electro-
phoresis platforms. The number of repeats in each
marker can be deduced from the sizes of the
fragments. All repeat numbers of the analyzed
markers form a genotype for each individual isolate.
These genotypes are easily compared to each other.
Tran-Dinh N and D Carter tested seven polymor-
phic microsatellite loci for 20 A. flavus and 15
A. parasiticus strains. Searches for microsatellite
motifs were performed on genomic sequences of
Aspergillus flavus complex. These seven markers
yielded two to eleven alleles for A. flavus and one to
nine alleles for A. parasiticus (Table 1) [49].
Guarro et al. used random amplified microsatel-
lites (RAMS) to type isolates of A. fumigatus and A.
flavus obtained from a supposed outbreak RAMS
combines microsatellite and RAPD analysis. A
discriminatory power of 0.9489 was obtained with
the combination of two different primers. A full
understanding of population of A. flavus and the
discriminatory power of these and other typing
systems awaits a full population genetics study
(Table 1) [20].
Batista et al. have used (GTG)5 and (GACA)4
primers that produced differential amplification
Mycopathologia (2011) 172:83–93 89
123
Page 8
products varying both in size and band intensity.
(GACA)4 revealed higher genetic variability. The
number and size of (GTG)5 bands were in a charac-
teristic pattern in several strains of A. flavus. A high
interspecific variation was observed. Furthermore, four
strains initially classified as A. flavus displayed differ-
ential banding patterns that prompted to review their
taxonomic identification (Table 1) [21]. Microsatel-
lite-based approaches appear to be species specific and
it is able to recognize instantaneously mixed genotypes
by the presence of multiple peaks of different lengths of
the marker analyzed [50, 51].
Grubisha LC and Cotty PJ. investigated the distri-
bution of genetic variation in 243 samples of cotton
from three common vegetative compatibility groups
(VCGs) in Arizona and Texas. They used 24 micro-
satellite loci and the mating type locus to assess
population structure among A. flavus VCGs in sym-
patric populations. They found high levels of genetic
differentiation and no evidence of gene flow between
VCGs, including VCGs of opposite mating type. These
results suggest that these VCGs diverged before
domestication of agricultural hosts (Table 1) [52].
Hadrich et al. identified and selected suitable
microsatellite markers for A. flavus typing. 63
A. flavus isolates (48 from Sfax, Tunisia and 15 from
Marseille, France) were analyzed. The combination
of all 12 markers yielded 35 different haplotypes with
a 0.97 D value. A 5 markers combination (AFLA1,
AFLA3, AFLA7, AFM3, and AFM7) yielded 27
different alleles with a 0.952 D value. Isolates from
Tunisia and Marseille displayed distinct haplotypes,
indicating a highly significant geographical structur-
ing in A. flavus. The typing of clinical and environ-
mental A. flavus isolates in a hematology unit
provided insights into its hospital epidemiology.
From a heterogeneous genetic background, a cluster
indicative of a clonal propagation episode within the
unit could be identified. In two patients with invasive
aspergillosis, the same genotype was found in clinical
and environmental isolates, indicating hospital-
acquired colonization and infection (Table 1) [10].
Interpretation
The performance of typing techniques can be com-
pared to each other in respect of their practical
feasibility.
Specific advantages and disadvantages are evalu-
ated in terms of applicability, the ease of use, the
exchangeability, and the reproducibility within a
laboratory and between laboratories. The discrimina-
tory power of the typing methods will be also
evaluated. Table 1 gives an overview of the different
aspects for all typing techniques discussed before.
All techniques mentioned in this review can
roughly be divided into pattern-based approaches
and exact techniques. The patterns obtained with
band-based techniques can be extremely complex,
because they are composed of both strong and faint
bands, which make the interpretation of these meth-
ods difficult [15, 26]. In contrast, exact fingerprinting
techniques are much easier to interpret [10, 20].
RFLP analysis with Southern blotting may be
tedious and labor intensive [30]. RAPD analysis is
the most frequently applied method, although lack of
reproducibility is a well-known limitation of this
technique [15, 26]. Given the extensive polymor-
phism of microsatellites, they have proved to be
epidemiologically useful for typing A. flavus [10, 20].
Reproducibility refers to the ability of a technique to
yield the same result when a particular isolate is
repeatedly tested. For large scale and longitudinal
epidemiological studies, stable fingerprinting tech-
niques are required. The main source of ambiguity in
band-based approaches like RAPD, RFLP, SSDP, and
AFLP concerning reproducibility is the variable inten-
sity of bands, which is probably on account of small
variations in the various steps of the procedures that may
affect the final peak intensity [15, 26, 36, 37]. In contrast,
exact techniques, like microsatellites, are potentially
100% reproducible [10, 20]. Another important aspect
in evaluating typing techniques is the ability to exchange
data between different laboratories. Therefore, exact
techniques seem to be most suitable, because the data
generated with these techniques can be fully expressed
in a simple, digital format.
A clear advantage of microsatellite-based methods
compared to band-based approaches is its ability to
identify mixtures of isolates. Mixed genotypes in
microsatellite-based methods are recognized instan-
taneously by the presence of multiple peaks of
different lengths of the marker analyzed. In band-
based approaches, mixed genotypes are very difficult,
if not impossible, to recognize.
Another problem among the analysis of general
typing data is the subjective interpretation of a
90 Mycopathologia (2011) 172:83–93
123
Page 9
genotype. The assignment of a genotype is user
dependent when using band-based approaches,
whereas the genotype resulting from exact techniques
is unambiguous.
Apart from the advantages and disadvantages of
practical feasibility and the interpretation of the
different techniques, another important factor, the
discriminatory power must be evaluated.
Buffington et al. combined the products from
RAPD analysis and RFLP analysis of a tester strain of
A. flavus to produce a DNA probe for Southern blot
analysis [33]. Although a high degree of discrimina-
tion among strain types was achieved, the probe and
target sequences remain undisclosed.
A discriminatory power of 0.9489 was obtained
with the combination of two different primers [20].
Grubisha LC and Cotty PJ used 24 microsatellite
loci. They found high levels of genetic differentiation
(ID = 0.86–1.0) [52]. The combination of all 12
markers yielded 35 different haplotypes with a 0.97 D
value [10].
Conclusion
The molecular typing of clinical and environmental
isolates of A. flavus will provide insights into
important epidemiological and public health issues
including tracing sources and routes of transmission,
identification of pathogenic or drug-resistant strains,
and the genetic relatedness of isolates. Assessing the
relatedness of strains isolated from patients and their
environment is instrumental in understanding the
epidemiology of this mold and documenting the
source of preventable health care-associated life-
threatening human infections. RAPD analysis may
not be appropriate as a tool for epidemiologic
tracking of isolates or for surveying the genetic
variation in natural populations because of many
artefactual variations [15]. Restriction endonuclease
analysis of total cellular DNA has not proved to be a
suitable method for the discrimination of strains of A.
flavus. RFLP analysis of A. flavus nuclear DNA
probed with recombinant DNA clones from
A. nidulans and Neurospora crassa supported the
results obtained with mitochondrial DNA but
revealed limited geographic correlations among
A. flavus strains. In recent years, it has been a
growing tendency to use exact typing methods for
discrimination between isolates. These methods are
advantageous over conventional methods in the
production of reproducible, portable, and exchange-
able typing data. The exact typing methods to date
are multilocus sequence typing (MLST) and micro-
satellite-based typing. A MLST scheme has been
developed for A. fumigatus typing (http://pubmlstorg/
afumigatus/) [53], but not for A. flavus. Most of the
techniques are developed to analyze A. fumigatus
isolates which is the most common species. The
sequencing of total genome of A. flavus may provide
more exact high-resolution fingerprinting techniques
in the future.
References
1. Krishnan S, Manavathu EK, Chandrasekar PH. Aspergillusflavus: an emerging non-fumigatus Aspergillus species of
significance. Mycoses. 2009;52:206–22.
2. Khairallah SH, Byrne KA, Tabbara KF. Fungal keratitis in
Saudi Arabia. Doc Ophthalmol. 1992;79:269–76.
3. Kameswaran M, al-Wadei A, Khurana P, Okafor BC.
Rhinocerebral aspergillosis. J Laryngol Otol. 1992;
106:981–5.
4. Mahgoub ES, el-Hassan AM. Pulmonary aspergillosis
caused by Aspergillus flavus. Thorax. 1972;27:33–7.
5. Hadrich I, Makni F, Sellami H, et al. Invasive aspergillosis:
epidemiology and environmental study in haematology
patients (Sfax, Tunisia). Mycoses 2009.
6. Choudhary SV, Koley S, Mallick S, Bose S, Basak S.
Proximal subungual onychomycosis caused by Aspergillusflavus in a HIV-positive patient. Indian J Dermatol Vene-
reol Leprol. 2009;75:410–2.
7. Gordon G, Giddings NA. Invasive otitis externa due to
Aspergillus species: case report and review. Clin Infect
Dis. 1994;19:866–70.
8. Debeaupuis JP, Sarfati J, Chazalet V, Latge JP. Genetic
diversity among clinical and environmental isolates of
Aspergillus fumigatus. Infect Immun. 1997;65:3080–5.
9. Chazalet V, Debeaupuis JP, Sarfati J, et al. Molecular
typing of environmental and patient isolates of Aspergillusfumigatus from various hospital settings. J Clin Microbiol.
1998;36:1494–500.
10. Hadrich I, Makni F, Ayadi A, Ranque S. Microsatellite
typing to trace Aspergillus flavus infections in a hematol-
ogy unit. J Clin Microbiol. 2010;48:2396–401.
11. Chang PK, Ehrlich KC. What does genetic diversity of
Aspergillus flavus tell us about Aspergillus oryzae? Int J
Food Microbiol. 2010;138:189–99.
12. Rodriguez E, Symoens F, Mondon P, et al. Combination of
three typing methods for the molecular epidemiology of
Aspergillus fumigatus infections. European Research
Group on Biotype and Genotype of Aspergillus. J Med
Microbiol. 1999;48:181–94.
Mycopathologia (2011) 172:83–93 91
123
Page 10
13. Birch M, Nolard N, Shankland GS, Denning DW. DNA
typing of epidemiologically-related isolates of Aspergillusfumigatus. Epidemiol Infect. 1995;114:161–8.
14. Denning DW, Clemons KV, Hanson LH, Stevens DA.
Restriction endonuclease analysis of total cellular DNA of
Aspergillus fumigatus isolates of geographically and epide-
miologically diverse origin. J Infect Dis. 1990;162:1151–8.
15. Lin D, Lehmann PF, Hamory BH, et al. Comparison of
three typing methods for clinical and environmental iso-
lates of Aspergillus fumigatus. J Clin Microbiol.
1995;33:1596–601.
16. Anderson MJ, Gull K, Denning DW. Molecular typing by
random amplification of polymorphic DNA and M13
southern hybridization of related paired isolates of Asper-gillus fumigatus. J Clin Microbiol. 1996;34:87–93.
17. van Belkum A, Quint WG, de Pauw BE, Melchers WJ,
Meis JF. Typing of Aspergillus species and Aspergillusfumigatus isolates by interrepeat polymerase chain reac-
tion. J Clin Microbiol. 1993;31:2502–5.
18. Aufauvre-Brown A, Cohen J, Holden DW. Use of ran-
domly amplified polymorphic DNA markers to distinguish
isolates of Aspergillus fumigatus. J Clin Microbiol.
1992;30:2991–3.
19. Rinyu E, Varga J, Ferenczy L. Phenotypic and genotypic
analysis of variability in Aspergillus fumigatus. J Clin
Microbiol. 1995;33:2567–75.
20. Guarro J, Sole M, Castany R, et al. Use of random
amplified microsatellites to type isolates from an outbreak
of nosocomial aspergillosis in a general medical ward. Med
Mycol. 2005;43:365–71.
21. Batista PP, Santos JF, Oliveira NT, Pires AP, Motta CM,
Luna-Alves Lima EA. Genetic characterization of Brazil-
ian strains of Aspergillus flavus using DNA markers. Genet
Mol Res. 2008;7:706–17.
22. Diaz-Guerra TM, Mellado E, Cuenca-Estrella M, Gaz-
telurrutia L, Navarro JI, Tudela JL. Genetic similarity
among one Aspergillus flavus strain isolated from a patient
who underwent heart surgery and two environmental
strains obtained from the operating room. J Clin Microbiol.
2000;38:2419–22.
23. Myoken Y, Sugata T, Fujita Y, et al. Molecular epidemi-
ology of invasive stomatitis due to Aspergillus flavus in
patients with acute leukemia. J Oral Pathol Med.
2003;32:215–8.
24. Heinemann S, Symoens F, Gordts B, Jannes H, Nolard N.
Environmental investigations and molecular typing of
Aspergillus flavus during an outbreak of postoperative
infections. J Hosp Infect. 2004;57:149–55.
25. Yuan GF, Liu CS, Chen CC. Differentiation of Aspergillusparasiticus from Aspergillus sojae by random amplifica-
tion of polymorphic DNA. Appl Environ Microbiol.
1995;61:2384–7.
26. Meunier JR, Grimont PA. Factors affecting reproducibility
of random amplified polymorphic DNA fingerprinting. Res
Microbiol. 1993;144:373–9.
27. Klich MA, Yu J, Chang PK, Mullaney EJ, Bhatnagar D,
Cleveland TE. Hybridization of genes involved in aflatoxin
biosynthesis to DNA of aflatoxigenic and non-aflatoxigenic
aspergilli. Appl Microbiol Biotechnol. 1995;44:439–43.
28. Moody SF, Tyler BM. Restriction enzyme analysis of
mitochondrial DNA of the Aspergillus flavus group:
A. flavus, A. parasiticus, and A. nomius. Appl Environ
Microbiol. 1990;56:2441–52.
29. McAlpin CE, Mannarelli B. Construction and character-
ization of a DNA probe for distinguishing strains of
Aspergillus flavus. Appl Environ Microbiol. 1995;61:
1068–72.
30. James MJ, Lasker BA, McNeil MM, Shelton M, Warnock
DW, Reiss E. Use of a repetitive DNA probe to type
clinical and environmental isolates of Aspergillus flavusfrom a cluster of cutaneous infections in a neonatal
intensive care unit. J Clin Microbiol. 2000;38:3612–8.
31. Lasker BA. Evaluation of performance of four genotypic
methods for studying the genetic epidemiology of Asper-gillus fumigatus isolates. J Clin Microbiol. 2002;40:
2886–92.
32. Semighini CP, Delmas G, Park S, Amstrong D, Perlin D,
Goldman GH. New restriction fragment length polymor-
phism (RFLP) markers for Aspergillus fumigatus. FEMS
Immunol Med Microbiol. 2001;31:15–9.
33. Buffington J, Reporter R, Lasker BA, et al. Investigation of
an epidemic of invasive aspergillosis: utility of molecular
typing with the use of random amplified polymorphic DNA
probes. Pediatr Infect Dis J. 1994;13:386–93.
34. Geiser DM, Dorner JW, Horn BW, Taylor JW. The phy-
logenetics of mycotoxin and sclerotium production in
Aspergillus flavus and Aspergillus oryzae. Fungal Genet
Biol. 2000;31:169–79.
35. Bagyalakshmi R, Therese KL, Madhavan HN. Nucleotide
polymorphisms associated with Internal Transcribed
Spacer (ITS) regions of ocular isolates of Aspergillus fla-vus. J Microbiol Methods. 2007;68:1–10.
36. Montiel D, Dickinson MJ, Lee HA, et al. Genetic differ-
entiation of the Aspergillus section Flavi complex using
AFLP fingerprints. Mycol Res. 2003;107:1427–34.
37. Kumeda Y, Asao T. Single-strand conformation polymor-
phism analysis of PCR-amplified ribosomal DNA internal
transcribed spacers to differentiate species of Aspergillussection Flavi. Appl Environ Microbiol. 1996;62:2947–52.
38. Wang L, Yokoyama K, Takahasi H, et al. Identification of
species in Aspergillus section Flavi based on sequencing of
the mitochondrial cytochrome b gene. Int J Food Micro-
biol. 2001;71:75–86.
39. Guarro J. GeneJ, Stchigel AM. Developments in fungal
taxonomy. Clin Microbiol Rev. 1999;12:454–500.
40. Kumeda Y, Asao T. Heteroduplex panel analysis, a novel
method for genetic identification of Aspergillus Section
Flavi strains. Appl Environ Microbiol. 2001;67:4084–90.
41. Rigo K, Varga J, Toth B, Teren J, Mesterhazy A, Ko-
zakiewicz Z. Evolutionary relationships within Aspergillussection Flavi based on sequences of the intergenic tran-
scribed spacer regions and the 5.8S rRNA gene. J Gen
Appl Microbiol. 2002;48:9–16.
42. Kozakiewicz Z. Aspergillus species on stored products.
Mycol Pap. 1989;161:1–88.
43. Frisvad JC, Skouboe P, Samson RA. Taxonomic compar-
ison of three different groups of aflatoxin producers and a
new efficient producer of aflatoxin B1, sterigmatocystin
and 3-O-methylsterigmatocystin, Aspergillus rambellii sp.
nov. Syst Appl Microbiol. 2005;28:442–53.
44. Pildain MB, Vaamonde G, Cabral D. Analysis of popula-
tion structure of Aspergillus flavus from peanut based on
92 Mycopathologia (2011) 172:83–93
123
Page 11
vegetative compatibility, geographic origin, mycotoxin and
sclerotia production. Int J Food Microbiol. 2004;93:31–40.
45. Geiser DM, Pitt JI, Taylor JW. Cryptic speciation and
recombination in the aflatoxin-producing fungus Asper-gillus flavus. Proc Natl Acad Sci USA. 1998;95:388–93.
46. Peterson SW. Phylogenetic analysis of Aspergillus species
using DNA sequences from four loci. Mycologia.
2008;100:205–26.
47. Lee CZ, Liou GY, Yuan GF. Comparison of the aflR gene
sequences of strains in Aspergillus section Flavi. Micro-
biology. 2006;152:161–70.
48. Puers C, Hammond HA, Jin L, Caskey CT, Schumm JW.
Identification of repeat sequence heterogeneity at the
polymorphic short tandem repeat locus HUM-
TH01[AATG]n and reassignment of alleles in population
analysis by using a locus-specific allelic ladder. Am J Hum
Genet. 1993;53:953–8.
49. Tran-Dinh N, Carter D. Characterization of microsatellite
loci in the aflatoxigenic fungi Aspergillus flavus and
Aspergillus parasiticus. Mol Ecol. 2000;9:2170–2.
50. Bart-Delabesse E, Humbert JF, Delabesse E, Bretagne S.
Microsatellite markers for typing Aspergillus fumigatusisolates. J Clin Microbiol. 1998;36:2413–8.
51. de Valk HA, Meis JF, Curfs IM, Muehlethaler K, Mouton
JW, Klaassen CH. Use of a novel panel of nine short
tandem repeats for exact and high-resolution fingerprinting
of Aspergillus fumigatus isolates. J Clin Microbiol. 2005;
43:4112–20.
52. Grubisha LC, Cotty PJ. Genetic isolation among sympatric
vegetative compatibility groups of the aflatoxin-producing
fungus Aspergillus flavus. Mol Ecol. 2010;19:269–80.
53. Bain JM, Tavanti A, Davidson AD, et al. Multilocus
sequence typing of the pathogenic fungus Aspergillusfumigatus. J Clin Microbiol. 2007;45:1469–77.
Mycopathologia (2011) 172:83–93 93
123