Gap Dynamics in a Seagrass Landscape Susan S. Bell,* Bradley D. Robbins, and Susan L. Jensen Department of Biology, University of South Florida, Tampa, Florida 33620-5150, USA ABSTRACT We investigated gap dynamics within a shallow subtidal landscape characterized by seagrass vegeta- tion and examined the relationship between gap formation and selected physical factors. The study was conducted over 2 y by using a biannual map- ping of seagrass and water depth across an 48,800-m 2 area in Tampa Bay, Florida. In addition, monthly sediment deposition or erosion was recorded at 96 locations within the landscape. Gaps represented from 2.4% to 5.7% of the seagrass landscape, and all were within monospecific stands of Halodule wrightii. Gaps ranged in size from 10 to 305 m 2 and most frequently decreased in size over time. Most gaps were small and short lived (less than 6-mo duration), but the second age group most frequently recorded was at least 1.5 y old. No new species of seagrass invaded the gaps with Halodule replacing itself 100% of the time. Gaps were recorded over the entire range of water depths within the land- scape. Neither gap area nor persistence of gaps was related to water depth. However gap area was associated positively with the number of extreme sedimentation events. Gaps originated not only from removal of interior vegetation (similar to classic gaps) but also from differential growth of the seagrass margin (similar to edaphic gaps). Distinct seasonal components to the mode of formation were detected with interior-produced gaps originat- ing primarily in the winter and margin gaps most commonly during summer. These results combine to illustrate the importance of large-scale studies with fine-scale resolution for deciphering unique features of seagrass landscape dynamics. Our histori- cal information suggests that a static enumeration of gaps may not provide an accurate assessment of disturbance intensity in this system, and the sea- grass mosaic probably is explained best by a combi- nation of disturbance regimes and edaphic factors, such as sediment stability. Moreover, we suggest that even in areas characterized by monospecific stands of vegetation and over short or moderate time periods, gaps indirectly may influence commu- nity structure and ecosystem function via modifica- tion of habitat arrangement. Key words: gaps; landscape; patch dynamics; sea- grasses; sedimentation; Florida; Tampa Bay; Halod- ule wrightii. INTRODUCTION Gaps, commonly defined as openings created in structured environments by disturbance events (for example, Whitmore 1989), have received consider- able attention in terrestrial settings (Brokaw 1987; Lawton and Putz 1988; Canham 1989; Kubo and others 1996) but less so in aquatic ecosystems (Kay and Keough 1981; Karlson 1983; Sousa 1984). Whereas most information on gaps comes from investigations on vegetation, some data on gaps in benthic faunal assemblages are also available (for example, Paine and Levin 1981). Studies of gap dynamics have provided insight into comparative life history features of both the organisms that colonize gaps early in succession and those more typical of later stages (for example, Dayton 1971; Shumway and Bertness 1994). The contribution of gaps to the maintenance of species diversity (see Petraitis and others 1989) is a common theme among the investiga- tions on this topic. Received 17 September 1998; accepted 26 April 1999. *Corresponding author: e-mail: [email protected]Ecosystems (1999) 2: 493–504 ECOSYSTEMS r 1999 Springer-Verlag 493
Transcript
Gap Dynamics in a SeagrassLandscape
Susan S. Bell,* Bradley D. Robbins, and Susan L. Jensen
Department of Biology, University of South Florida, Tampa, Florida 33620-5150, USA
ABSTRACTWe investigated gap dynamics within a shallowsubtidal landscape characterized by seagrass vegeta-tion and examined the relationship between gapformation and selected physical factors. The studywas conducted over 2 y by using a biannual map-ping of seagrass and water depth across an 48,800-m2
area in Tampa Bay, Florida. In addition, monthlysediment deposition or erosion was recorded at 96locations within the landscape. Gaps representedfrom 2.4% to 5.7% of the seagrass landscape, andall were within monospecific stands of Halodulewrightii. Gaps ranged in size from 10 to 305 m2 andmost frequently decreased in size over time. Mostgaps were small and short lived (less than 6-moduration), but the second age group most frequentlyrecorded was at least 1.5 y old. No new species ofseagrass invaded the gaps with Halodule replacingitself 100% of the time. Gaps were recorded overthe entire range of water depths within the land-scape. Neither gap area nor persistence of gaps wasrelated to water depth. However gap area wasassociated positively with the number of extremesedimentation events. Gaps originated not onlyfrom removal of interior vegetation (similar to
classic gaps) but also from differential growth of theseagrass margin (similar to edaphic gaps). Distinctseasonal components to the mode of formationwere detected with interior-produced gaps originat-ing primarily in the winter and margin gaps mostcommonly during summer. These results combineto illustrate the importance of large-scale studieswith fine-scale resolution for deciphering uniquefeatures of seagrass landscape dynamics. Our histori-cal information suggests that a static enumeration ofgaps may not provide an accurate assessment ofdisturbance intensity in this system, and the sea-grass mosaic probably is explained best by a combi-nation of disturbance regimes and edaphic factors,such as sediment stability. Moreover, we suggestthat even in areas characterized by monospecificstands of vegetation and over short or moderatetime periods, gaps indirectly may influence commu-nity structure and ecosystem function via modifica-tion of habitat arrangement.
Gaps, commonly defined as openings created instructured environments by disturbance events (forexample, Whitmore 1989), have received consider-able attention in terrestrial settings (Brokaw 1987;Lawton and Putz 1988; Canham 1989; Kubo andothers 1996) but less so in aquatic ecosystems (Kayand Keough 1981; Karlson 1983; Sousa 1984).
Whereas most information on gaps comes frominvestigations on vegetation, some data on gaps inbenthic faunal assemblages are also available (forexample, Paine and Levin 1981). Studies of gapdynamics have provided insight into comparativelife history features of both the organisms thatcolonize gaps early in succession and those more typicalof later stages (for example, Dayton 1971; Shumwayand Bertness 1994). The contribution of gaps to themaintenance of species diversity (see Petraitis andothers 1989) is a common theme among the investiga-tions on this topic.
Received 17 September 1998; accepted 26 April 1999.*Corresponding author: e-mail: [email protected]
From an ecosystem perspective, information isbeginning to accumulate to suggest that some eco-system functions are altered by gap formation orwithin gap areas. Smith and others (1994) foundthat mangrove tree mortality after a hurricane wasreduced for trees that previously had invaded smallgaps created by lightning and listed different soilbiogeochemical traits in gap areas relative to undis-turbed sites. Vitousek and Denslow (1986) investi-gated treefall gaps with no reported change in Nmineralization but slight increases in P concentra-tion in gap areas. More recently, Zhang and Zak(1995) revealed that ecosystem functions, such asmicrobial activity and rates of litter decomposition,may be altered in gap areas. In such studies, it iscritical to assess the frequency and areal extent ofgap formation so that potential impacts of gaps canbe evaluated.
As ecological investigations expand to larger scales,the role of gaps as modifiers of landscape continuityhas begun to be appreciated (Denslow 1985). Gapshave been incorporated into spatially explicit descrip-tions of large-scale patterns of habitat (Coffin andLauenroth 1989; Wu and Levin 1994) and in dy-namic habitat models including lattice structuremodels or those of cellular automata (see Kubo andothers 1996). Landscape-level studies conductedover large spatial scales thus provide the templatefor examining gap characteristics that may impactecosystem level processes.
If gap formation is intense and persistent, a spatialmosaic characterized by large areas devoid of land-scape elements may result (for example, Sprugel1976). Alteration in habitat arrangement by re-moval of structure may act to either decrease thecontinuity of the landscape or increase the availabil-ity of edges. The impact of landscape modificationby gaps may be appreciable if the behavior oforganisms that use habitat structure as physical/biotic refugia or as feeding/nesting sites also ismodulated by the arrangement of the landscapeelements (With and Crist 1995). Viewed in thisperspective, gap size may be a critical feature forthose organisms whose behavior is linked to habitatarrangement. Likewise size of gap may impact ratesof nutrient exchange especially if removal of vegeta-tion alters physical conditions.
Herein we extend the study of gap dynamics to asubtidal seagrass landscape characterized by a mono-specific stand of Halodule wrightii, extending overthousands of meters. Seagrasses are broadly distrib-uted in both temperate and tropical coastal settings(den Hartog 1970; Iverson and Bittaker 1986) andprovide structural complexity. Within the oftenlarge expanses of submerged aquatic vegetation,
gaps are recognizable and can originate by eitherbiotic or abiotic sources. Known gap producersinclude disturbance agents, such as sea turtles(Thayer and others 1984) rays (Orth 1975), crusta-ceans (Suchanek 1983), and persistent algal mats(Cowper 1978). Anthropogenic effects, such as boatpropeller scars (Zieman 1976), also may removeseagrass thereby producing a spatial mosaic of unveg-etated and vegetated areas. Water depth and sedi-mentation also may exhibit relationships to gapdynamics or characteristics, although their role isless well understood. Water depth might be impor-tant because shallow areas may be more susceptibleto prolonged exposure or biological disturbanceagents might be limited to particular depths. Sedi-mentation may have a role in gap formation if stormevents lead to severe deposition that can buryseagrasses (Preen and others 1995) or cause large-scale rippling or erosion of sediments (Patriquin1975). Harlin and Thorne-Miller (1982) suggestedthat seagrass distribution in a flood tidal delta inRhode Island was controlled by sediment move-ment, which in turn was regulated by wind-generated water movement. Thus the process of gapformation may logically be tied to the physicalsetting.
The impacts of either physical or biological distur-bance agents on seagrass vegetation may vary sea-sonally with respect to intensity, frequency, orduration as has been discussed previously in anumber of other habitats (Pickett and White 1985).Strong seasonal trends in gap dynamics may exist inseagrass systems. For example, seagrass growth isminimized commonly in winter and is most prolificduring spring and summer in subtropical climates(Dunton 1990). Such seasonal patterns may be aresult of available light, temperature, and/or aerialexposure in the higher tidal levels. Therefore boththe origin and persistence of gaps, coupled with theresponse by vegetation to gap formation, may havea distinct seasonal signature that imparts temporalas well as spatial heterogeneity of seagrass coverageto the landscape.
Most studies on macrophyte spatial dynamicshave been conducted on relatively small scales, andmany focus on describing the patterns of vegetationpatch growth exclusively (for example, Brouns1987; Williams 1990; Sand Jensen and Madsen1992; Oleson and Sand-Jensen 1994; Ruckelshaus1994). Our study provides an unprecedented viewof the persistence of large-scale spatial organizationof submerged macrophytes in the marine subtidal ata level of resolution previously unreported. Themain objectives of our investigation were to (a) useinformation collected from a large-scale mapping of
494 S. S. Bell and others
seagrass beds to investigate patterns of gap dynam-ics within a seagrass landscape; (b) compare sea-sonal variation in gap formation and closure; and (c)examine relationships between gap characteristicsand physical setting.
METHODS AND MATERIALS
Defining Gaps
Gaps have been identified in a variety of ways. Theclassic definition would focus on disturbance-generated openings in either floral or faunal cover(Connell 1978). However, some reports do notaddress presence/absence of vegetation but insteadcompare background canopies with those of re-duced height (gaps; Lawton and Putz 1988; Tanakaand Nakashizuka 1997) after a disturbance event.Shumway and Bertness (1994) referred to openareas in a salt marsh as patches, but the majority ofstudies appears to consider patches as areas withvegetation. Moreover, Lertzman and others (1996)present information on gap formation in forests thatfocuses on two types of gaps: (a) classic gaps thathave openings resulting from vegetation mortality;and (b) edaphic gaps that represent canopy open-ings associated with topographical or edaphic fea-tures, such as soil conditions. The latter gap neednot be linked with a gap maker. Therefore modifica-tions of the classic definition of gaps have occurredsuch that the designation of openings in a landscapemay not reflect areas completely devoid of vegeta-tion, and the formation of openings need not neces-sarily be linked to disturbance events. An overallconclusion here is that terminology associated withthe study of gaps is highly variable and semanticproblems can easily arise.
In spatial models, designation of locations aseither gap or no gap is of primary interest (see Kuboand others 1996), and we adopt this approach.Because we do not know precisely the source ofgaps in our seagrass system, we follow the precedentestablished by studies on dynamic spatial models,and in our first level of analyses, we limit ourselvesto identifying gap and no gap areas within theseagrass landscape. Gaps were operationally definedin our study as areas greater than 10 m2 lackingvegetation (that is, sand) surrounded by a largercontiguous area of seagrass vegetation. We do notinclude smaller size gaps because our mappingsindicated that smaller gaps closed rapidly and werenot best represented by our temporal scale of sam-pling. Specifically, a survey of gaps smaller than 10m2 (not presented here) on each date revealed onlytwo out of a possible 56 instances of their expanding
to greater than 10 m2 on any date (S. S. Bell,unpublished).
The second approach that we use in this studyparallels that of Lertzman and others (1996). Weanalyzed gaps in the landscape that are aptly catego-rized as those fitting the classic definition of gaps.We also used the results of our sequential mappingsof gaps (see below) to identify openings in theseagrass vegetation that might not fit the classicdefinition of gaps but rather are more appropriatelyaligned with edaphic conditions in shallow subtidalareas, but remembering that types of edaphic condi-tions that may be important in an aquatic settingmay differ markedly from that encountered interrestrial habitats.
Field SamplingTo obtain a spatially explicit representation of theseagrass landscape, we conducted a field-based map-ping in a subtidal area approximately 122 m 3 400m on a shallow sand shoal. The mapped site wassituated at the mouth of Tampa Bay, Florida(27°478N, 82°378W) with annual salinity and tem-perature ranges typically 30–32o/oo and 15–34°C,respectively (S. S. Bell, unpublished). Three sea-grass species are present on the shoal with thelandward edge dominated by contiguous Thalassiatestudinum meadows, the deeper bay edge domi-nated by a mixture of T. testudinum and Syringodiumfiliforme, and the upper shoal dominated by a com-posite of large and small Halodule wrightii beds. Ourmapping was conducted almost exclusively withinHalodule beds. The shoal has a mean depth of 40 cmat low tide, although seasonally extreme tidal eventsin this wind-dominated estuary may result in expo-sure of the shoal for extended periods of time.
The site was mapped biannually during the spring(S) and fall (F) of 1994 and 1995 (S94, F94, S95, andF95, respectively) when water clarity and tidalconditions, which are major limitations for collect-ing large spatial representations of seagrasses, wereoptimal. Each seagrass species was identified, andpresence/absence was recorded at 1-m intervals ateach of 48,800 points by systematically transversingthe shoal. This in situ methodology follows fromtechniques used by Fonseca and Bell (1998) inprevious work conducted in smaller seagrass beds inNorth Carolina.
Because we did not know from where new gapswould form over the course of our study, wemeasured both depth and sedimentation synopti-cally across the entire landscape. On each of the fourmapping dates, water depths to the nearest 5 cmwere recorded at intervals of 3 m (every thirdlocation) with the aid of a 2-m pole calibrated into
Seagrass Gaps 495
5-cm intervals. The time of each water depth mea-surement also was recorded so that all depths couldbe standardized to mean low water (MLW). Sedi-mentation within the seagrass landscape also wasmeasured and here refers to the deposition orerosion of sediment in seagrass beds, often related tothe passage of sand ripples through the seagrass bed.As sand ripples pass through a seagrass bed, anyspecific location may experience alternating inter-vals of sediment deposition and erosion, and areaswith substantial sediment movement should exhibitsuch fluxes in sediment accumulation. Thereforewe monitored changes in sediment depth monthlyfrom May 1994 to October 1995 by using a modifica-tion of the buried sediment plate method outlinedby Emerson (1991). Sediment depth was measuredusing thin plexiglass plates (6 cm length 3 6 cmwidth 3 1 cm height) to which a 250-cm greenribbon was fastened in the center. The distance fromthe sediment surface to a buried plate was measuredmonthly with the aid of a thin rod, which wasinserted into the sediment until it contacted thesediment plate, and marked at the height of thesediment level; the rod then was removed from thesediment, and the length of the rod covered bysediment was measured using a ruler (mm). Wechose the plate size as a compromise in keepingplates sufficiently small so that they would haveminimum impact on plants and sediment processes(for example, they were smaller than clams found insediments), yet large enough that the thin rod usedto measure sediment depth had a good probabilityof contacting the sediment plate. Plates were buriedby inserting the plate sideways into the sedimentand then sliding them to a horizontal position belowthe rhizomes of the seagrasses with the ribbonextending through the rhizome mat to the sedimentsurface. The belowground insertion of sedimentplates allowed us to avoid problems with placementof structures on sediment surfaces, and the greenribbon, which mimicked seagrass blades, served asthe plate marker. Sediment plates were placedwithin the seagrass beds at least 1 m from theseagrass margin in 96 locations throughout thestudy area with the number of plates representingthe upper limit that could be visibly relocatedunderwater and sampled on a monthly basis, espe-cially during spring–summer months when waterclarity was extremely reduced.
We focused on the extreme sediment events,arbitrarily defined in this study as any monthlyincrease or decrease in sedimentation greater thanor equal to 6 cm. Both depositional and erosionalprocesses were visually noted at this site. We estab-lished this definition after examining the range of
sedimentation values for all sites over all dates. Wereasoned that if gaps were linked to either sedimenterosion or deposition, then evaluation of extremeconditions should provide the most probable indica-tion of a relationship. For each gap within thelandscape, we located the four closest sedimenta-tion plates at north, south, east, and west directions.We then recorded the frequency of extreme sedi-mentation at each of these four plates monthly overthe time interval (approximately 6 mo) immedi-ately preceding the detection of new gaps.
Data AnalysesSpatial representations of the area, by season, weregenerated from binary data files representing thepresence/absence of Halodule across the sampledarea, by using a raster-based GIS program (GRASS;Geographic Resources Analysis Support System;version 4.1) at a resolution of 1 m2. Gaps wereenumerated by season and size, and location (inte-rior vs edge) was determined. Using GIS, we com-pared maps between dates revealing the location andsize of gap birth (origin) and extinction (closure).
Depth data were standardized across the land-scape by using field depth adjusted for tidal shift andmapping time. These data were incorporated intoGRASS and interpolated using an inverse weighteddistance algorithm of the nearest points (r.surf.idw;Shapiro 1993). Using this procedure, we couldlocate gaps on the GIS layer of water depth for aparticular season and the corresponding water depthnearest to the center of each gap extracted.
Relationships between gap size (m2), number ofgaps, and persistence of gaps (whose age was pre-cisely known) over the study interval versus waterdepth were evaluated with Spearman rank ordercorrelation. The number of gaps versus the fre-quency of extreme sedimentation events was exam-ined using data combined from all dates exceptspring 1994 because we had no preceding sedimen-tation data for this set of values. Additionally weexamined the relationship between the size of gapsformed and the number of extreme sedimentationevents over the mapping interval during which thegaps were first detected with Spearman rank ordercorrelation. The median size of gaps that weregenerated by different modes of formation (see below)was compared using a Mann-Whitney test. Simi-larly, the median size of gaps that went extinct ver-sus persisting over consecutive sampling dates wascompared with a Mann-Whitney test. A null hypoth-esis that the mode of gap formation (disturbance vsedaphic) was independent of the type of extremesediment event [depositional (positive) vs erosional(negative)] was tested using a Fisher exact test.
496 S. S. Bell and others
RESULTS
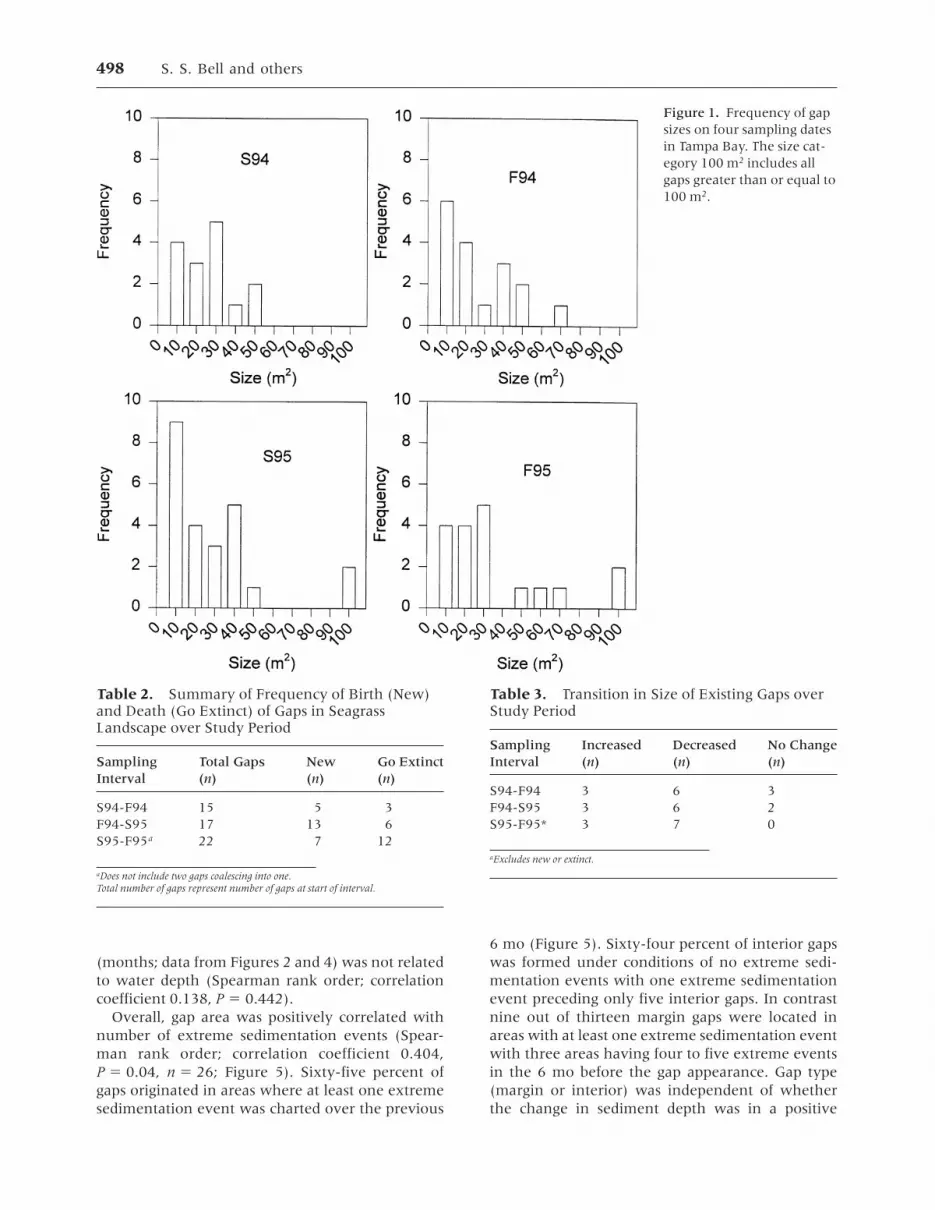
Gap DynamicsA total of 41 different gaps, greater than or equal to10 m2 in size, were identified over the study inter-val, and all were within Halodule beds. Of these, 15were present at the start of the study, and 26 werenewly generated over remaining sampling dates.Over the four dates examined, gaps greater than orequal to 10 m2 occupied from 2.4% to 5.7% of thetotal seagrass landscape (Table 1). Gaps ranged insize from 10 to 305 m2 on any one sampling datewith gaps greater than 80 m2 appearing only in S95and F95 (Figure 1). The largest number of new gapsappeared in the interval between F94 and S95(Table 2). Only in S95-F95 was there a higherincidence of gap extinction than birth. The rate ofgap extinction (extinctions/total number gaps/sampling interval) ranged from 20% to 54% perinterval; the rate of gap origin ranged from 32% to76% per interval. By the second time interval(F94-S95), the total number of newly formed gapssurpassed the number of gaps existing at the start ofthe study (Table 2).
Of those gaps that persisted from one samplingdate to another, the most frequent type of changewas a decrease in areal extent (Table 3). On all dates,the number of gaps that increased in size was 50%of those that decreased areally. Spatial dimensionsof five gaps remained static over one samplinginterval (Table 3), but no gap remained of constantdimension over two intervals. Gaps greater than 45m2 always decreased in size from one age interval tothe next. The median size of gaps going extinct bythe next sampling interval was significantly smallerthan that for gaps remaining to the next date (15.0vs 32.0 m2, respectively) (Mann-Whitney rank sumtest, T 5 496.5, P 5 0.008).
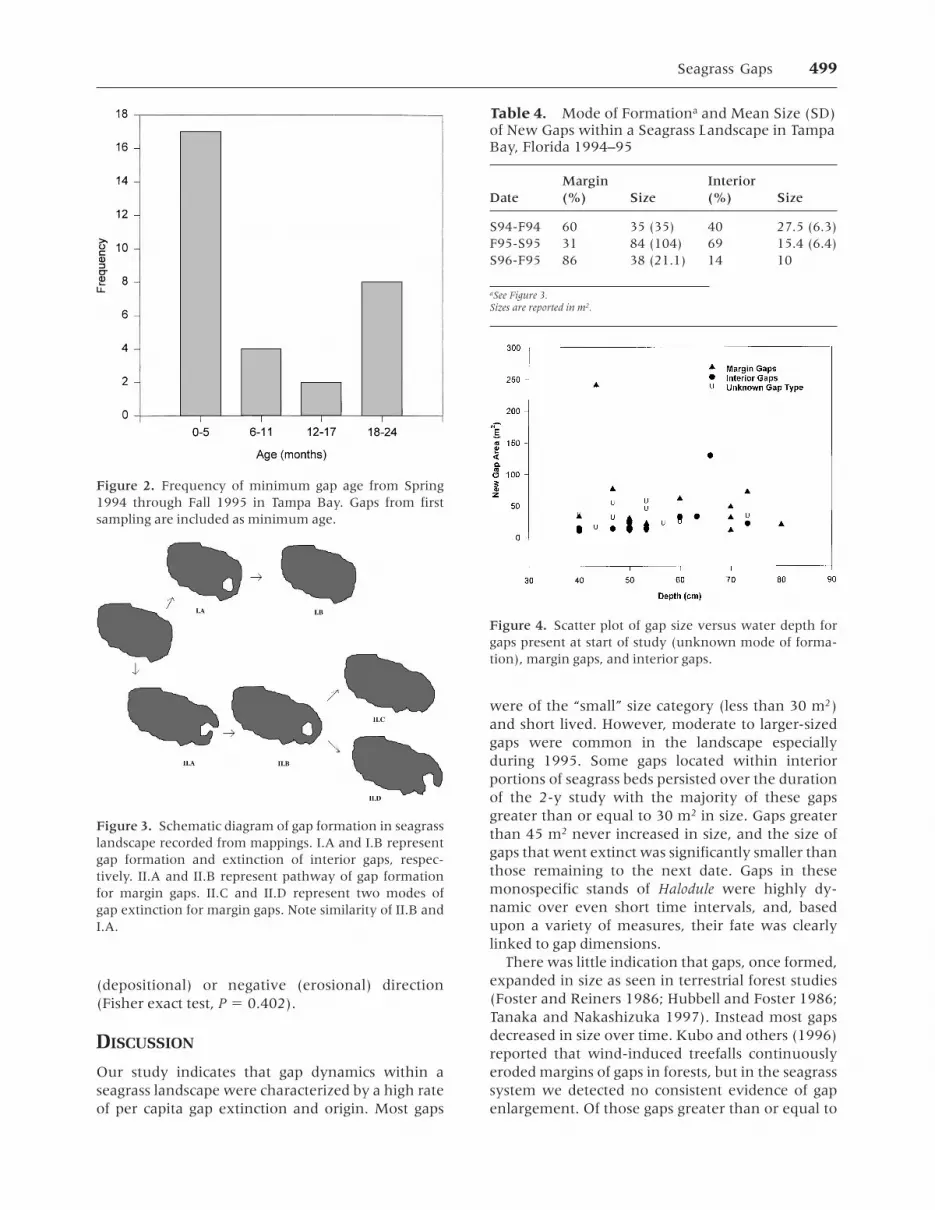
Scrutiny of the age structure of gaps indicatedthat the greatest number of gaps comprised the agecategory less than 6 mo (that is, most gaps persisted
for less than the duration between sample intervals;Figure 2). The second most frequently recorded agewas the oldest age group of at least 1.5 y (Figure 2).Of the 15 original gaps present at the start of thestudy, eight were present at the end. Eighty-twopercent of the newly formed gaps in F94-S95 wentextinct by the following 6 mo. No gaps greater thanor equal to 10 m2 and less than 15 m2 persisted formore than 1 y.
For those gaps that persisted over more than onesampling interval, no transitional constancy (that is,consistent direction of change) was recorded. Eightgaps were maintained over three sampling inter-vals, but only two displayed a consistent decrease insize. This was true for two of four gaps that re-mained over two sampling intervals. Other gapsdisplayed combinations of decrease, increase, or nochange. Importantly, in all cases, no new species ofseagrass or rhizophytic algae invaded gaps; Halodulewrightii replaced itself 100% of the time.
Visual inspection of maps indicated that new gapsin the landscape originated by either of two pro-cesses: (1) removal of vegetation from within exist-ing seagrass beds (classic definition of gaps) (inte-rior); or (2) the disproportional growth of seagrassat the margins of beds that expanded and envelopedunvegetated sediments (margin; Figure 3). Themost common mode of gap formation was marginenclosure in S94 to F94 and S95 to F95, whereasremoval of interior vegetation was the dominantprocess over the interval F94 to S95 (Table 4). Of theeight original gaps persisting to the end of the study,six were located in interior areas of the seagrassbeds. Mean size of margin-generated gaps wasgreater than that for interior openings (Mann-Whitney rank sum test, T 5 103.5, P 5 0.032). Notethat extinction of margin-produced gaps occurrednot only by total coverage of a previous opening (asin interior gaps) but also by the opening in an outerrim of vegetation (Figure 3). The former scenariowas the most common, whereas the latter occurredonly twice.
Relationship to Physical FactorsGaps were found at all water depths within thelandscape with both shallow and deep locationscontaining the smallest gaps (Figure 4). Gap areadisplayed no significant relationship to water depth(Spearman rank order; correlation coefficient 0.192,P 5 0.229; Figure 4). When examined by gap type,size of interior gaps was significantly positivelycorrelated with depth (Spearman rank order; corre-lation coefficient 0.671, P 5 0.001), but margin gapsize was not (Spearman rank order; correlationcoefficient 20.136, P 5 0.643). Gap persistence
Table 1. Spatial Extent (m2) of SeagrassVegetation and Gaps over Four Sampling Dates1994–95 in Tampa Bay, Florida
(months; data from Figures 2 and 4) was not relatedto water depth (Spearman rank order; correlationcoefficient 0.138, P 5 0.442).
Overall, gap area was positively correlated withnumber of extreme sedimentation events (Spear-man rank order; correlation coefficient 0.404,P 5 0.04, n 5 26; Figure 5). Sixty-five percent ofgaps originated in areas where at least one extremesedimentation event was charted over the previous
6 mo (Figure 5). Sixty-four percent of interior gapswas formed under conditions of no extreme sedi-mentation events with one extreme sedimentationevent preceding only five interior gaps. In contrastnine out of thirteen margin gaps were located inareas with at least one extreme sedimentation eventwith three areas having four to five extreme eventsin the 6 mo before the gap appearance. Gap type(margin or interior) was independent of whetherthe change in sediment depth was in a positive
Figure 1. Frequency of gapsizes on four sampling datesin Tampa Bay. The size cat-egory 100 m2 includes allgaps greater than or equal to100 m2.
Table 2. Summary of Frequency of Birth (New)and Death (Go Extinct) of Gaps in SeagrassLandscape over Study Period
SamplingInterval
Total Gaps(n)
New(n)
Go Extinct(n)
S94-F94 15 5 3F94-S95 17 13 6S95-F95a 22 7 12
aDoes not include two gaps coalescing into one.Total number of gaps represent number of gaps at start of interval.
Table 3. Transition in Size of Existing Gaps overStudy Period
SamplingInterval
Increased(n)
Decreased(n)
No Change(n)
S94-F94 3 6 3F94-S95 3 6 2S95-F95* 3 7 0
aExcludes new or extinct.
498 S. S. Bell and others
(depositional) or negative (erosional) direction(Fisher exact test, P 5 0.402).
DISCUSSION
Our study indicates that gap dynamics within aseagrass landscape were characterized by a high rateof per capita gap extinction and origin. Most gaps
were of the ‘‘small’’ size category (less than 30 m2)and short lived. However, moderate to larger-sizedgaps were common in the landscape especiallyduring 1995. Some gaps located within interiorportions of seagrass beds persisted over the durationof the 2-y study with the majority of these gapsgreater than or equal to 30 m2 in size. Gaps greaterthan 45 m2 never increased in size, and the size ofgaps that went extinct was significantly smaller thanthose remaining to the next date. Gaps in thesemonospecific stands of Halodule were highly dy-namic over even short time intervals, and, basedupon a variety of measures, their fate was clearlylinked to gap dimensions.
There was little indication that gaps, once formed,expanded in size as seen in terrestrial forest studies(Foster and Reiners 1986; Hubbell and Foster 1986;Tanaka and Nakashizuka 1997). Instead most gapsdecreased in size over time. Kubo and others (1996)reported that wind-induced treefalls continuouslyeroded margins of gaps in forests, but in the seagrasssystem we detected no consistent evidence of gapenlargement. Of those gaps greater than or equal to
Figure 2. Frequency of minimum gap age from Spring1994 through Fall 1995 in Tampa Bay. Gaps from firstsampling are included as minimum age.
Figure 3. Schematic diagram of gap formation in seagrasslandscape recorded from mappings. I.A and I.B representgap formation and extinction of interior gaps, respec-tively. II.A and II.B represent pathway of gap formationfor margin gaps. II.C and II.D represent two modes ofgap extinction for margin gaps. Note similarity of II.B andI.A.
Table 4. Mode of Formationa and Mean Size (SD)of New Gaps within a Seagrass Landscape in TampaBay, Florida 1994–95
Figure 4. Scatter plot of gap size versus water depth forgaps present at start of study (unknown mode of forma-tion), margin gaps, and interior gaps.
Seagrass Gaps 499
10 m2, only 27% increased in size between mappingdates with the remainder either decreasing or re-maining of the same area. Physical or biologicalfactors that would facilitate removal of seagrass atmargins of gaps, such as crab activity (Valentine andothers 1994), do not seem to be important inmaintaining the spatial patterns of the seagrasslandscape examined here.
One of the intriguing findings from our study wasthat gaps not only originated from removal ofinterior vegetation but also from differential growthat the seagrass bed margin (see Figure 3). In thelatter case, most gaps were of the largest size class,and given that at least one extreme sedimentationevent was noted at 77% of margin beds, expansionof the beds may be a response to sediment alter-ation. Because margin growth represents the appear-ance of new shoots in areas where they werepreviously absent, these gaps, although indistin-guishable from interior gaps in our spatial represen-tation (see Figure 3), do not conform to the classicview of disturbance-generated openings (for ex-ample, Sousa 1985). Thus, as discussed by Lertzmanand others (1996), gaps that arise other than byclassic disturbance events contributed substantiallyto the overall pattern of vegetation openings in theseagrass beds. These findings also warn that unlikeother systems that have focused upon gap dynamics(see Introduction), a static enumeration of gaps maynot provide an accurate assessment of disturbanceintensity in this seagrass system. A more accurateview would be that the seagrass mosaic is best
explained by a combination of disturbance regimesand edaphic factors possibly linked to sedimentstability. In some seagrass beds, light attenuationthat increases with water depth might be a majoredaphic feature of this subtidal environment, butour results do not support water depth as beingconsistently important in this study.
Gap size frequencies from our seagrass study, as insome terrestrial systems (Brokaw 1985; Kubo andothers 1996), were skewed to the smallest sizeclasses. Such size distributions are logically linked tothe source of disturbances and their frequency ofoccurrence. However, unlike reports from previ-ously studied areas, there was a considerable num-ber of larger-sized gaps present in the seagrasssystem as well. This last feature is mainly a result ofthe margin-produced gaps in seagrass vegetation,which apparently either have not been detectedpreviously or are unimportant in the other habitats.Other reports on seagrasses either are at such a largescale that they do not have the resolution to detectmargin gaps (for example, Robbins 1997), such asseen here, or are of sufficient resolution, but thespatial dimensions of the sampling site may berelatively small (for example, Ruckelshaus 1994).Our study demonstrates that the accessibility of atemporal record of large spatial representations withfine-scale resolution is critical to discerning some ofthe unique features of seagrass dynamics, in particu-lar, and possibly submerged macrophytes, in gen-eral. The need for fine-scale resolution of large-scalestudies is further discussed by Tanaka and Nakashi-zuka (1997) for forest systems.
Distinct seasonal components to the mode offormation of new gaps were recorded with marginformation being most common during summermonths, whereas interior formation originated pri-marily in winter. The causes of differential margingrowth are poorly understood but may be in re-sponse to smaller variations in depth (not withinour level of resolution) or nutrient availabilityassociated with sediment movement/deposition. Ifso, then expansion of margins is expected to occurmost often during periods of rapid seagrass growthin spring to fall but should be curtailed in wintermonths, as our data suggests (Table 4). In two of thethree sampling intervals, the number of gaps pro-duced by margin growth exceeded those originatingfrom vegetation removal suggesting that the modeof gap generation may play an unexplained role ingap dynamics. Strong seasonal patterns in interiorgap formation may reflect the combined effects ofgreater tidal amplitudes and aerial exposure as aresult of extremely low tides and winds typicallypresent in winter months in the wind-dominated
Figure 5. Scatter plot of gap size versus number ofextreme sedimentation events. Extreme sedimentation isdefined here as sediment depth increase or decrease ofgreater than or equal to 6 cm per month.
500 S. S. Bell and others
Tampa Bay estuary. Such seasonal patterns in theintensity of disturbance is commonly reported inmarine benthic systems (see Sousa 1984).
We documented that change in sediment depth inour sampling site may be substantial (66 cm/mo) inareas near where many margin gaps were locatedsuggesting that burial of plants may be one mecha-nism of gap origin in our system. Marba and others(1994) have discussed how one species of seagrass,Cymodocea nodosa, can respond to the passage of sandwaves through seagrass beds by growing upwardfrom buried rhizomes, thus illustrating that some‘‘apparent gaps’’ simply may be buried shoots. IfHalodule also is able to respond to sediment deposi-tion by regrowth, then this may help to explain highturnover and rapid extinction of gaps recorded here.Sedimentation events as generators of landscapepatterns have been reported in terrestrial grasslands(Loucks and others 1985) and may be analogous inour system. Whereas it would be desirable to havemeasures of sediment changes at the precise loca-tions where gaps are formed, this would be difficultbecause future gap sites are not predictable and thenumber of sediment plates required would exceedlogistical capabilities.
Unlike disturbance agents that are readily observ-able in the field and/or leave distinctive signatures,the agents of gap formation in this seagrass systemremain poorly understood. Not knowing the ‘‘gapmakers’’ is not uncommon especially in marinehabitats (Paine and Levin 1981). We did documentthat the agents of disturbance were not limited to aparticular depth because gaps of all sizes were foundspanning the range of depths in this area. Forinterior-produced gaps, we do not know the precisesource of gap formation because we did not observeactual removal of vegetation. In a few sites, blow-outs (sensu Patriquin 1975) were visually obviousin 20% of gaps because of exposed rhizomes andsediment depressions; in other areas, surface sedi-ments in gaps were not visually distinguishablefrom sediments with seagrass cover. Whereas gap-producing organisms (for example, rays and horse-shoe crabs) are abundant at the site, we have notdirectly observed evidence of such destructive activi-ties. Anthropogenic effects, such as propeller scars,may cause barren areas within seagrass beds, butgap shapes from this study did not display the longlinear swaths usually associated with propeller scars,and we discount their importance as gap makers atour site. Macroalgal accumulations within seagrassbeds were not monitored in this study but canproduce barren areas if the trapped patches aresubstantial in size and persistent (Holmquist 1992).However, information on macroalgae from this site
suggests that algal mats do not accumulate for longperiods of time within the seagrass beds at Ft.Desoto in lower Tampa Bay, FL (Bell and Hall 1997).These results combine to suggest that, unlike terres-trial systems where treefalls are reportedly linked towind-driven disturbance, gap formation in sea-grasses may be due to multiple causes that probablyrepresent complex interactions between the physi-cal setting and biological agents of disturbance.
Although precise information on the origin ofgaps within seagrass beds is not available, our largescale and spatially explicit study provides uniqueinsight into the mechanisms of gap closure. Wedocumented that gap persistence was not related towater depth and that gaps often went extinct withina 6-mo period. Such closure from revegetation mayoccur in a variety of ways. Seagrasses are largelyclonal and vegetation coverage may expand via seedrecruitment and/or by shoot production from under-ground rhizomatic growth. The importance of thesemethods varies among species and locations (seeOrth and others 1994; Ruckelshaus 1994). In areaswith reduced flowering and seed production, below-ground growth patterns rather than seedling recruit-ment dynamics may play a major role in the closureof gaps generated by a variety of disturbances.Growth by rhizomatic extension into unvegetatedareas and the subsequent production of short shootsmay be the most direct manner and is probably ofmost importance in our system. Estimates of rhi-zome elongation for Halodule are at least 1 m/y(Jensen 1998), and thus closure of most gaps can beexplained by this mechanism. Additionally, seedrecruitment to unvegetated areas may occur but,unlike Zostera for which seeds are abundant andrepopulation of sediments by seed has been noted(Orth and others 1996), is probably unimportant inthis area. Evidence of some seed reserves of Halodulehave been reported from other sites in Texas andFlorida (MacMillan 1981, 1983), but we have notencountered any flowering Halodule in monthlyseagrass samples nor seen any seeds in Tampa Bayfrom over 1750 sediment cores taken from our area(S. S. Bell, unpublished observations). A third mech-anism may be regrowth of short shoots from buriedrhizomes. Because mapping of seagrass landscapesis based upon the presence/absence of abovegroundvegetation, it is possible that gap closure could occurif sediments shift sufficiently to allow regrowth frompreviously buried belowground components. It re-mains unknown as to how long rhizomes of Halod-ule can be covered by sediment and subsequentlyproduce new short shoots. Answers to these ques-tions would require extensive and destructive map-
Seagrass Gaps 501
ping of the underground components of the sea-grass landscape.
It is difficult to compare the findings of our studywith other seagrass systems because of the paucityof large-scale studies addressing gap dynamics. Thisis most probably a result of the difficulty of obtain-ing sufficient information on spatial distributions ofsubmerged plants. However, existing informationon dispersal and recruitment in other seagrass sys-tems allows us to speculate on possible similarities.For example, the high rates of sexual recruitmentand low dispersal distances of Zostera marina on thewest coast of North America (Alberte and others1994; Ruckleshaus 1994) and in Denmark (Olesenand Sand-Jensen 1994) indicate that seed dispersaland germination may be the most frequent mecha-nism of large-scale gap closure, relative to closurevia underground rhizomatic growth for this taxa.Conversely, seagrasses that exhibit predominantlyvegetative growth could fill in gaps more quickly byrhizome growth than seedling recruitment. Cymodo-cea nodosa populations in the Mediterranean pre-dominantly recruit through vegetative growth (Du-arte and Sand-Jensen 1990), and Syringodiumfiliforme can display lateral vegetative growth of upto 8 m y21 (Brouns 1987). In the Caribbean, Wil-liams (1990) found all recolonization of seagrassesHalodule wrightii, Syringodium filiforme, and Thalassiatestudinum into cleared plots to be solely throughvegetative means. Recolonization and filling-in ofgaps may be location and/or species specific, wherelocal climate influences reproduction and recruit-ment success of seagrass populations. The methodsused in our study could be used to expand investiga-tions on seagrass gap dynamics and modes of gapformation into other geographic areas.
Information from this study provides uniquecomparative information on gaps in marine subtidalseagrass beds versus those reported from terrestrialgrasslands. Links between sedimentation and gapformation have been reported in both seagrass andgrassland habitats. Animal reworking of sedimentsin serpentine grasslands can change soil featuresleading to gap formation (for example, Bonis andothers 1997), and disturbance by sediment moundscreated by gophers are a common component of theserpentine grasslands (Wu and Levin 1994). Simi-larly, our study suggests that sediment deposition ina shallow seagrass bed may influence gap formation.However, differences between gaps in seagrass andgrasslands also exist with respect to gap size andrecruitment into openings. Gaps reported in theseagrass system here were relatively large (greaterthan 10 m2), which appears to be uncommon interrestrial grassland communities where gaps are
often less than 1 m in diameter (White 1979; Hookand others 1994). No new seagrass species invadedgaps via rhizome elongation because all gaps werebordered by Halodule, and thus the beds remainedmonospecific. In fact only in 6% of locations withinthe total landscape was Halodule replaced by otherseagrass species over a 2-y period (see Robbins 1998and discussion within). In contrast, in terrestrialgrasslands, gaps serve as sites for seed recruitmentand offer areas for invasion by additional plantspecies (Grubb 1977).
ImplicationsThe importance of gaps and disturbance in maintain-ing diversity has been discussed repeatedly in avariety of systems based upon arguments that inter-mediate disturbance (removal of landscape ele-ments) may reset the successional scenario and leadto a higher diversity of structural elements (mostoften plants; Connell 1978). But if modification ofvegetation cover via intermediate disturbance altersthe ability of other organisms to successfully use thelandscape, the impact on ‘‘overall’’ diversity (bothstructure and associated organisms) may not neces-sarily be a positive one, especially when gaps arelarge. It remains to be determined whether interme-diate disturbance, which arguably enhances struc-tural diversity, maintains connectivity of the land-scape for those members of the ecosystem that relyon the structural features of the spatial mosaic. Inthe seagrass system, information on the movementof epibenthos and nekton across landscapes withdiffering densities and arrangements of gaps wouldbe necessary to evaluate whether openings pro-vided by gaps are analogous to fragmentation. Theeffects of gaps on diversity may extend to theattached epifaunal (for example, mussels) and infau-nal components of the seagrass ecosystem as well,given that some taxa show strong affiliation withseagrass presence (for example, Virnstein 1987; Belland others 1993; Reusch and Chapman 1995), andthe removal of vegetation is expected to impactthose fauna intimately related to seagrass. Recentdata demonstrate that both sediment organic con-tent and abundance of small infaunal benthos arealtered in gaps generated by boat propellers (Love-joy 1997).
Although margin gaps may look like interior gaps,do they function in a similar way? For example,Orth (1977) reported that infaunal diversity insediments within seagrass beds was higher thanunvegetated areas adjacent to seagrass. Thus, onemight expect that gap areas, especially those formedat margins, also may have lower infaunal diversitythan areas with intact vegetation. In the Tampa Bay
502 S. S. Bell and others
seagrass system, the expected reduction in infaunaldiversity may be short lived for many of the gapsthat close rapidly; the effects of the older and largergaps on infaunal diversity may be more persistent.Some of these differences in gap areas may be linkedto a reduction in the amount of fine sediments thatare typically trapped by subtidal plants. As arguedby Robbins and Bell (1994), information gatheredfrom a landscape approach, such as presented here,provides a baseline for evaluating the impact of gapson ecosystem structure and function that are stronglyaffected by the arrangement of structural features ofthe habitat.
ACKNOWLEDGMENTS
P. Stiling, M. Ruckelshaus, and two anonymousreviewers made many helpful comments on anearlier version of the manuscript. R. Brooks, K.Madley, C. Rose, P. Rodriquez, M. Platz, J. Clarke,and K. Overholtzer assisted in field mappings. Thiswork was funded in part by National Science Foun-dation grant (DEB 93–18130) to S. S. Bell.
REFERENCES
Alberte RS, Suba GK, Procaccini G, Zimmerman RC, Fain SR.1994. Assessment of genetic diversity of seagrass populationsusing DNA fingerprinting: implications for population stabilityand management. Proc Nat Acad Sci 91:1049–53.
Bell SS, Hall MO. 1997. Drift macroalgal abundance in seagrassbeds: investigating large-scale associations with physical andbiotic attributes. Mar Ecol Prog Ser 147:277–83.
Bell SS, Clements LAJ, Kurdziel J. 1993. Production in naturaland restored seagrasses: case study of a macrobenthic poly-chaete. Ecol Appl 3:610–21.
Bonis A. 1997. Requirements of gap-demanding species in chalkgrassland: reduction of root competition versus nutrient enrich-ment by animals. J Ecol 85:625–33.
Brokaw NVL. 1985. Treefalls, regrowth and community structurein tropical forests. In: Pickett ST, White PS, editors. The ecologyof natural disturbance and patch dynamics. New York: Aca-demic Press. p 53–70.
Brokaw NVL. 1987. Gap-phase regeneration of three pioneer treespecies in a tropical forest. J Ecol 75:9–19.
Brouns JJWM. 1987. Growth patterns in some Indo-West Pacificseagrasses. Aquat Bot 28:39–61.
Canham CD. 1989. Different responses to gaps among shadetolerant tree species. Ecology 70:548–9.
Coffin DP, Lauenroth WK. 1989. Disturbances and gap dynamicsin a semiarid grassland: a landscape-level approach. Ecology70:19–27.
Connell JH. 1978. Diversity in tropical rain forests and coral reefs.Science 199:1302–10.
Cowper SW. 1977. The drift algal community of seagrass beds inRedfish Bay, Texas. Contrib Mar Sci 31:125–32.
Dayton PK. 1971. Competition, disturbance, and communityorganization: the provision and subsequent utilization of spacein a rocky intertidal community. Ecol Monogr 41:351–89.
Denslow JS. 1985. Disturbance-mediated coexistence of species.In: Pickett ST, White PS, editors. The ecology of naturaldisturbance and patch dynamics. New York: Academic Press. p307–24.
den Hartog C. 1970. The seagrasses of the world. London: NorthHolland Publishing Company.
Duarte CM, Sand-Jensen K. 1990. Seagrass colonization: biomassdevelopment and shoot demography in Cymodocea nodosapatches. Mar Ecol Prog Ser 67:97–103.
Dunton KH. 1990. Production ecology of Ruppia maritima L.s. 1.and Halodule wrightii Aschers in two tropical estuaries. J ExpMar Biol Ecol 143:147–64.
Emerson CW. 1991. A method for the measurement of bedloadsediment transport and passive faunal transport on intertidalsand flats. Estuaries 14:361–71.
Fonseca MS, Bell SS. 1998. The influence of wave exposure andtidal currents on seagrass landscapes. Mar Ecol Prog Ser171:109–21.
Foster JR, Reiners WA. 1986. Size distribution and expansion ofcanopy gaps in a northern Appalachian spruce-fir forest.Vegetatio 68:109–14.
Grubb PJ. 1977. The maintenance of species richness in plantcommunities: the importance of the regeneration niche. BiolRev 52:107–45.
Harlin MM, Thorne-Miller B. 1982. Seagrass-sediment dynamicsof a flood tidal delta in Rhode Island (USA). Aquat Bot14:127–38.
Holmquist JG. 1992. Disturbance, dispersal and patch insularityin a marine benthic assemblage: influence of a mobile habitaton seagrasses and associated fauna [dissertation]. Tallahassee,FL: Florida State University.
Hook PB, Laurenroth WK, Burke IC. 1994. Spatial patterns ofroots in a semiarid grassland: abundance of canopy openingsand regeneration gaps. J Ecol 82:485–4.
Hubbell SP, Foster RB. 1986. Canopy gaps and the dynamics of aneotropical forest. In: Crawley MS, editor. Plant ecology.Oxford UK: Blackwell Scientific. p. 77–96.
Iverson RI, Bittaker HF. 1986. Seagrass distribution and abun-dance in eastern Gulf of Mexico coastal waters. Estuar CoastShelf Sci 221:577–602.
Jensen SL. 1998. Below-ground morphological plasticity of theseagrass, Halodule wrightii, in relation to sediment resources[dissertation]. Tampa: University of South Florida.
Karlson RH. 1983. Disturbance and monopolization of a spatialresource by Zooanthus sociatus (Coelenterata: Anthozoa). BullMar Sci 33:118–131.
Kay AM, Keough MJ. 1981. Occupation of patches in theepifaunal communities on pier piling and the bivalve Pinnabicolor at Edithburg, South Australia. Oecologia 48:123–30.
Kubo T, Iwasa Y, Furumoto N. 1996. Forest spatial dynamics withgap expansion: total gap area and gap size distribution. J TheorBiol 180:229–46.
Lawton OR, Putz FE. 1988. Natural disturbance and gap-phaseregeneration in a wind exposed tropical cloud forest. Ecology69:764–77.
Lertzman KP, Sutherland GD, Inselberg A, Saunders AC. 1996.Canopy gaps and the landscape mosaic in a coastal temperaterainforest. Ecology 77:1254–70.
Loucks OL, Plumb-Mentjes ML, Rogers D. 1985. Gap processesand large-scale disturbances in sand prairies. In: Pickett ST,White PS, editors. The ecology of natural disturbance andpatch dynamics. New York: Academic Press. p 71–83.
Seagrass Gaps 503
Lovejoy S. 1997. Assessment of the ecological effects of propellerdamage from motor boats on a Thalassia testudinum (Banks exKoenig) bed in Charlotte Harbor, Florida, USA. Masters thesis.UK: University of North Wales. 51 p.
MacMillan C. 1981. Seed reserves and seed germination for twoseagrasses Halodule wrightii and Syringodium filiforme from thewestern Atlantic. Aquat Bot 11:279–96.
MacMillan C. 1983. Seed germination in Halodule wrightii andSyringodium filiforme from Texas and the US Virgin Islands.Aquat Bot 15:217–20.
Marba N, Cerbian J, Enriques S, Duarte CM. 1994. Migration oflarge-scale subaqueous bed forms measured with seagrasses(Cymodocea nodosa) as tracers. Limnol Oceanogr 39:126–33.
Olesen B, Sand-Jensen K. 1994. Demography of shallow eelgrass(Zostera marina) populations-shoot dynamics and biomass de-velopment. J Ecol 82:379–90.
Orth RJ. 1975. Destruction of eelgrass Zostera marina by thecownose ray, Rhinoptera bonasus in the Chesapeake Bay, Vir-ginia, USA. Chesapeake Sci 16:205–8.
Orth RJ. 1977. The importance of sediment stability in seagrasscommunities. In: Coull BC, editor. The ecology of marinebenthos. Columbia: University of South Carolina Press p281–300.
Orth RJ, Luckenbach M, Moore KA. 1996. Seed dispersal in amarine macrophyte: implications for colonization and restora-tion. Ecology 75:1927–39.
Paine RT, Levin SA. 1981. Intertidal landscapes: disturbance andthe dynamics of pattern. Ecol Monogr 51:145–78.
Patriquin DG. 1975. Migration of blowouts in seagrass beds atBarbados and Carriacou, West Indies and its ecological andgeological implications. Aquatic Bot 1:163–89.
Petraitis PS, Latham RE, Niesenbaum RA. 1989. The mainte-nance of species diversity by disturbance. Q Rev Biol 64:393–418.
Pickett STA, White PS. 1985. Patch dynamics: a synthesis. In:Pickett ST, White PS, editors. The ecology of natural distur-bance and patch dynamics. New York: Academic Press. p.371–84.
Preen AR, Lee Long WJ, Coles RG. 1995. Flood and cyclonerelated loss and partial recovery of more than 1000 km2 ofseagrass in Hervey Bay, Queensland, Australia. Aquat Bot52:3–17.
Reusch TBH, Chapman ARO. 1995. Storm effects on eelgrass(Zostera marina L.) and blue mussel (Mytilus edulis L.) beds. JExp Mar Biol Ecol 192:257–71.
Robbins BD. 1997. Quantifying temporal change in seagrass arealcoverage: the use of GIS and low resolution aerial photogra-phy. Aquat Bot 58:259–68.
Robbins BD. 1998. Landscape dynamics: an example using asubtidal seagrass-dominated landscape [dissertation]. Tampa:University of South Florida.
Robbins BD, Bell SS. 1994. Seagrass landscapes: a terrestrialapproach to the marine subtidal environment. Trends EcolEvol 9:301–4.
Ruckelshaus MH. 1994. Ecological and genetic factors affectingpopulation structure in the marine angiosperm Zostera marinaL. (eelgrass) [dissertation]. Seattle: University of Washington.
Sand Jensen K, Madsen TV. 1992. Patch dynamics of the streammacrophyte, Callitriche cophocarpa. Freshwater Biol 27:277–82.
Shapiro M. 1993. r.surf.idw2-surface generation program. GRASS4.1 USA CERL, Champagne, Illinois.
Smith TJ III, Robblee MB, Wanless HR, Doyle TW. 1994. Man-groves, hurricanes and lightning strikes. Bioscience 44:256–62.
Sousa WP. 1984. Intertidal mosaics: patch size, propagule avail-ability and spatially variable patterns of succession. Ecology65:1918–35.
Sousa WP. 1985. Disturbance and patch dynamics on rockyintertidal shores. In: Pickett ST, White PS, editors. The ecologyof natural disturbance and patch dynamics. New York: Aca-demic Press. p 101–24.
Sprugel DG. 1976. Dynamic structure of wave generated Abiesbalsamea forests in the northeastern United States. J Ecol64:889–911.
Suchanek TH. 1983. Control of seagrass communities and sedi-ment distribution by Callianassa (Crustacea: Thalassinidea)bioturbation. J Mar Res 41:281–98.
Tanaka H, Nakashizuka T. 1997. Fifteen years of canopy dynam-ics analyzed by aerial photography in a temperate deciduousforest, Japan. Ecology 78:612–20.
Thayer GW, Bjorndal KA. Ogden JC, Williams SL, Zieman JL.1984. Role of larger herbivores in seagrass communities.Estuaires 7:351–76.
Valentine JF, Heck Jr KL, Harper P, Beck M. 1994. Effects ofbioturbation in controlling turtlegrass (Thalassia testudinumbanks ex Konig) abundance: evidence from field enclosuresand observations in the northern Gulf of Mexico. J Exp MarBiol Ecol 178:181–92.
Virnstein RW. 1987. Seagrass-associated invertebrate communi-ties of the southeastern USA: a review. In: Durako MJ, PhillipsRC, Lewis III RR, editors. Proceedings of the symposium onsubtropical-tropical seagrasses of the southeastern United States.Saint Petersburg, FL: Publication number 42. Florida Depart-ment of Natural Resources. p 89–116
Vitousek PM, Denslow JS. 1986. Nitrogen and phosphorusavailability in treefall gaps of a lowland tropical rainforest. JEcol 74:1167–78.
White PS. 1979. Pattern, process and natural disturbance invegetation. Bot Rev 45:229–99.
Whitmore TC. 1989. Canopy gaps and the two major groups offorest trees. Ecology 7:536–7.
Williams SL. 1990. Experimental studies of Caribbean seagrassbed development. Ecol Monogr 60:449–69.
With KA, Crist TO. 1995. Critical thresholds in species—responses to landscape structure. Ecology 76:2446–60.
Wu J, Levin SA. 1994. A spatial dynamics modeling approach topattern and process in annual grassland. Ecol Monogr 64:447–64.
Zhang Q, Zak JC. 1995. Effects of gap size on litter decompositionand microbial activity in a subtropical forest. Ecology 76:2196–204.
Zieman JC. 1976. The ecological effects of physical damage frommotor boats on turtlegrass in southern Florida. Aquat Bot2:127–39.