Blackwell Science, LtdOxford, UKBOJBotanical Journal of the Linnean Society0024-4074The Linnean Society of London, 2004? 20041464469481Original Article
INTERCONTINENTAL DISTRIBUTION OF PLAGIOCHILA CORRUGATAJ. HEINRICHS ET AL.
(Plagiochilaceae, Hepaticae) inferred from nrDNA ITS sequences and morphology
JOCHEN HEINRICHS
1
*, HENK GROTH
1
, MELANIE LINDNER
1
, CARSTEN RENKER
2
, TAMÁS PÓCS
3
and THOMAS PRÖSCHOLD
4
1
Albrecht-von-Haller-Institut für Pflanzenwissenschaften, Abteilung Systematische Botanik, Universität Göttingen, Untere Karspüle 2, D-37073 Göttingen, Germany
2
Insitut für Botanik, Abteilung für Terrestrische Ökologie, Johannisallee 21, D-04103 Leipzig, Germany
3
Research Group for Bryology, Hungarian Academy of Sciences, Department of Botany, Eszterházy College, H-3301, Eger, PO Box 222, Hungary
4
Botanisches Institut I, Universität zu Köln, Gyrhofstraße 15, D-50931 Köln, Germany
Received February 2003; accepted for publication June 2004
Plagiochila
sect.
Vagae
is a large pantropical clade that is characterized morphologically by frequent terminalbranching, vegetative distribution by propagules on the ventral surface of the leaves and a capsule wall with thick-enings in all layers.
Plagiochila corrugata
from Brazil is characterized by strongly undulate, toothed leaf marginsand represents the only known neotropical species of sect.
Vagae
with unispiral elaters.
Plagiochila cambuena
fromMadagascar is distinguished by the same features. Maximum likelihood and parsimony analyses of 38 nrDNA ITSsequences of
Plagiochila
reveal
P. corrugata
and
P. cambuena
in a weakly (ML) to well (MP) supported monophyleticlineage within
P.
sect.
Vagae
. As an outcome of the morphological and molecular investigation,
P. cambuena
is rel-egated to the synonymy of
P. corrugata. Plagiochila corrugata
is placed in a
Vagae
-subclade with 11 further Amer-ican species. The range of
P. corrugata
can be ascribed to long-range dispersal from the Neotropics rather than aGondwanan distribution. Species from tropical Asia and Africa are placed at the base of the
Africa – biogeography – Madagascar – maximum likelihood analysis – Neotropics
– parsimony analysis –
Plagiochila
sect.
Vagae
.
INTRODUCTION
Plagiochila
(Dumort.) Dumort. with more than 1600published binomials (Inoue, 1989) and roughly 400–450 morphospecies (So & Grolle, 2000) is the largestgenus of leafy liverworts. This cosmopolitan genus iswell defined by its dioecism, the laterally com-pressed perianth, alternate foliation and succubousleaves. In contrast, taxonomy at species and sectionlevel is hampered by lack of stable gametophyticcharacters.
Many authors (e.g. Gottsche, 1863–7; Spruce, 1884–5) tried to cope with the huge variability of
Plagiochila
gametophytes by describing numerous species thatwere usually assigned to small ranges only. However,the application of a narrow species concept is problem-atic because of the presence of numerous intermedi-ates. Modern revisions (e.g. Inoue, 1984) have thusadopted a wider species concept, resulting in binominawith larger geographical ranges and numerous syn-onyms.
Several authors have tried to subdivide
Plagiochila
into natural species groups. The last comprehensiveapproach to structure the genus was published by Carl(1931), who gave attention to
Plagiochila
worldwidewith the exception of Africa. Carl accepted 50 sectionsthat were usually restricted to a single continent or flo-ristic kingdom. He spread Neotropical
cies with frequent terminal branching and propaguleson the leaf surface over several sections, e.g.
Crispatae
Carl,
Parallelae
Carl, and
Contiguae
Carl. However,Groth, Lindner & Heinrichs (2004) demonstrated thatrepresentatives of these sections form a robust mono-phyletic lineage and that a pantropical clade is at hand;the oldest available name is
P.
sect.
Vagae
Lindenb.(Heinrichs, Sauer & Grolle, 2002b). Intercontinentalranges at species level are still exceptional within
P.
sect.
Vagae
(Heinrichs
et al
., 2002a). Neotropical
Vagae
are provided with bispiral elaters (e.g. Heinrichs& Gradstein, 2000), whereas several representatives of
P.
sect.
Vagae
from tropical Africa have unispiral elat-
ers (Jones, 1962; Vanden Berghen, 1981). Recently wediscovered unispiral elaters in Brazilian
P. corrugata
(Nees) Mont. & Nees. Here we present the resultsof a morphological and molecular comparison of
P. corrugata
with some African species.
MATERIAL AND METHODS
DNA
EXTRACTION
, PCR
AMPLIFICATION
AND
SEQUENCING
Upper parts of a few shoots of herbarium specimenswere isolated (Table 1). DNA was extracted using an
Table 1.
Geographical origins, voucher numbers and GenBank/EMBL accession numbers (bold for new sequences) of thetaxa investigated. Vouchers are deposited at GOET unless indicated otherwise
Taxon Origin Voucher Accession number
P. aerea
Taylor Costa Rica Heinrichs
et al
. 4321 AJ422028
P. arbuscula
(Lehm. & Lindenb.) Lindenb. Japan Yamaguchi 18280 (HIRO) AY550131
Invisorb Spin Plant Mini Kit (Invitek, Berlin, Ger-many). PCR-amplification followed the protocols ofHeinrichs et al. (2002a; P. corrugata, Brazil) or Hein-richs, Lindner & Pócs (2004; all others). Sequencingwas carried out on an ABI 3100 capillary sequencerusing the BigDye Terminator Cycle Sequencing v.2.0kit (PE Biosystems).
TAXON SAMPLING AND PHYLOGENETIC ANALYSES
The 13 new nrITS sequences were compared withGenBank sequences using the BLASTN program andput into a large alignment of Plagiochilaceae ITSsequences. In all cases sequences of Plagiochila sect.Vagae were identified as most similar to the newsequences (data not shown). Based on these results,representatives of Plagiochila sect. Vagae and theclosely related Plagiochila sects Cucullatae Schiffn.,Plagiochila, and Zonatae (e.g. Groth & Heinrichs,2003; Groth et al., 2003) were sampled. Plagiochilaaerea Taylor and P. fuscolutea Taylor (P. sect.Fuscoluteae Carl) were chosen as outgroup, accordingto the results of Groth et al. (2003).
25 ITS1-, 5.8S-, and ITS2 rDNA sequences fromHeinrichs (2002), Heinrichs et al. (2002a), Groth et al.(2003), Groth et al. (2004), Renker et al. (2002) and the13 new sequences were aligned manually in BIOEDITv.5.0.9 (Hall, 1999), resulting in an alignment includ-ing 806 putatively homologous sites (alignment avail-able from JH).
Phylogenetic trees were inferred using maximumlikelihood (ML) and maximum parsimony (MP) crite-ria as implemented in PAUP* v.4.0b10 (Swofford,2003).
Maximum likelihood analysesIn order to decide upon the nucleotide substitutionmodel with the smallest number of parameters thatbest fitted the data, the program MODELTEST v.3.06(Posada & Crandall, 1998) was used. Based on theresults of the tests, the model selected by the hierar-chical LRT was the TrN model (Tamura & Nei, 1993)with gamma shape parameter (G) for among site vari-ation calculated from the data set (TrN + G). A MLanalysis (with the TrN + G model) was implementedas heuristic search with ten random-additionsequence replicates. The confidence of branching wasassessed using 100 bootstrap resamplings in ML-analysis (Felsenstein, 1985).
MP analysesTwo datasets were analysed: (1) the above ITS datasetand (2) a dataset including only the P. sect. Vagaesequences. The analyses were performed with the fol-lowing options implemented: heuristic search modewith 1000 random-addition-sequence replicates, treebisection-reconnection branch swapping (TBR), MUL-
Trees option on, and collapse zero-length branches off.All characters were treated as equally weighted andunordered. Gaps were coded as unknown characters.Clade support was estimated from 1000 bootstrap rep-licates using heuristic searches with TBR branchswapping.
MORPHOLOGICAL INVESTIGATION
The morphological investigation was based on numer-ous specimens of Plagiochila sect. Vagae from the her-baria BM, EGR, F, FH, FLAS, G, GOET, JE, INB, LPB,MO, NY, PC, RB, S, STR, U, W, as well as identifiedand unidentified African Plagiochila vouchers fromthe private herbaria of Eberhard Fischer, AndreasSolga and Volker Buchbender (Koblenz).
RESULTS
MORPHOLOGICAL COMPARISON OF PLAGIOCHILA CORRUGATA AND AFRICAN VAGAE SPECIES WITH
UNISPIRAL ELATERS
Neotropical Plagiochila corrugata (Fig. 4) is charac-terized by terminal branching, presence of copiousunderleaves, dense foliation, leaves with stronglyundulate and crisped, toothed ventral and basal mar-gins, and predominantly unispiral elaters. Unispiralelaters are well known from several African Vagaebinomials (P. cambuena Steph., P. lastii Mitt.,P. pinniflora Steph., P. squamulosa Mitt.; Jones, 1962;Vanden Berghen, 1981). Of these, only P. cambuenafrom Madagascar has strongly undulate leaves(Fig. 5). The only weak morphological differencebetween P. cambuena and P. corrugata is found in thedentation of the ventral leaf base. The ventral leafbase of P. corrugata is almost always beset withbroadly to elongate triangular teeth. Ventral leafbases of P. cambuena are more often entire; however,at least some ventral leaf bases are provided with pre-dominantly broadly triangular teeth.
SEQUENCE VARIATION
The length of ITS 1 within P. sect. Vagae variesbetween 359 bp (P. arbuscula) and 386 bp (P. disticha).The ITS 1 sequence of P. corrugata is 381 bp long, thatof P. cambuena 369 bp. ITS 1 within the other inves-tigated sections have lengths between 341 bp(P. orbicularis) and 363 bp (P. subplana, Peru). The5.8S sector is nearly unvariable at 161 ± 2 bp. ITS 2could not always be determined completely. SectionVagae sequences vary between 257 bp (P. subtropica)and 303 bp in P. corrugata. ITS 2 of P. cambuena hasa length of 282 bp. ITS 2 length variation withinthe other investigated sections is between 250 bp(P. fuscolutea) and 278 bp (P. subplana Peru). The
aligned P. corrugata and P. cambuena ITS 1 sequencesdiffer in 18 base positions as well as 14 indels. ITS 2 ofboth taxa has 14 different base positions and 35 indelpositions.
PHYLOGENETIC ANALYSES OF THE ITS REGION
Maximum likelihoodThe maximum likelihood analysis produced a singletree (Fig. 1) in which the 36 ingroup sequences areplaced in four robust sectional clades: Cucullatae, Pla-
giochila, Zonatae, Vagae. Plagiochila sect. Cucullataeis sister to the remainder of the genus but this topol-ogy is not well supported. The relationships betweenP. sects. Vagae, Zonatae and Plagiochila are notresolved. Plagiochila sect. Vagae is placed sister tosect. Zonatae albeit without support. Plagiochila sect.Vagae species from the Palaeotropics form a paraphyl-etic grade at the base of the Vagae clade. Foursequences of P. squamulosa from Zaire and Kenyaform a robust monophyletic lineage. The subdivision ofP. squamulosa into varieties is not supported in the
Figure 1. Molecular phylogeny of Plagiochila species based on ITS1-5.8S-ITS2 rDNA sequence comparisons using 806aligned positions. The rooted tree shown resulted from a ML analysis of 38 sequences using the model of Tamura & Nei(1993) with estimated gamma shape (G = 0.488), calculated as the best model by MODELTEST v.3.06 (Posada & Crandall,1998). Bootstrap percentage values (>50%) at branches.
INTERCONTINENTAL DISTRIBUTION OF PLAGIOCHILA CORRUGATA 473
ML topology. A robust polytomous clade with 11 Neo-tropical species and P. virginica from Tenerife isplaced sister to P. squamulosa. Plagiochila corrugatafrom Brazil forms a weakly supported monophyleticlineage with P. cambuena from Madagascar.Plagiochila fastigiata Lindenb. & Gottsche fromMexico is sister to P. corrugata/cambuena withoutsupport.
Maximum parsimonyComplete dataset: Of the 806 investigated charac-ters, 182 were parsimony informative, 134 autapo-
morphic and 490 constant. The heuristic searchrecovered 402 equally most parsimonious trees (notshown) with a length of 593 steps, a consistencyindex (CI) of 0.64, a CI excluding uninformativecharacters of 0.52, a retention index (RI) of 0.76 anda rescaled consistency index (RC) of 0.49. The strictconsensus of these trees (Fig. 2) is largely congru-ent to the ML tree shown in Figure 1. Differencesare found only in weakly or unsupported topologies.Contrary to the ML topology, P. sect. Zonatae isplaced sister to P. sect. Plagiochila in a weakly sup-ported sister relationship. The sister relationship of
Figure 2. Rooted strict consensus of 402 equally parsimonious trees recovered during 1000 random taxon additionheuristic searches of the ITS data set. Bootstrap support (> 50%) is indicated at branches.
P. corrugata and P. cambuena is well supported(bootstrap support of 77). The two P. raddianasequences are placed in a polytomous clade withseveral other Neotropical species. However, theirsister relationship achieves a bootstrap support of76.
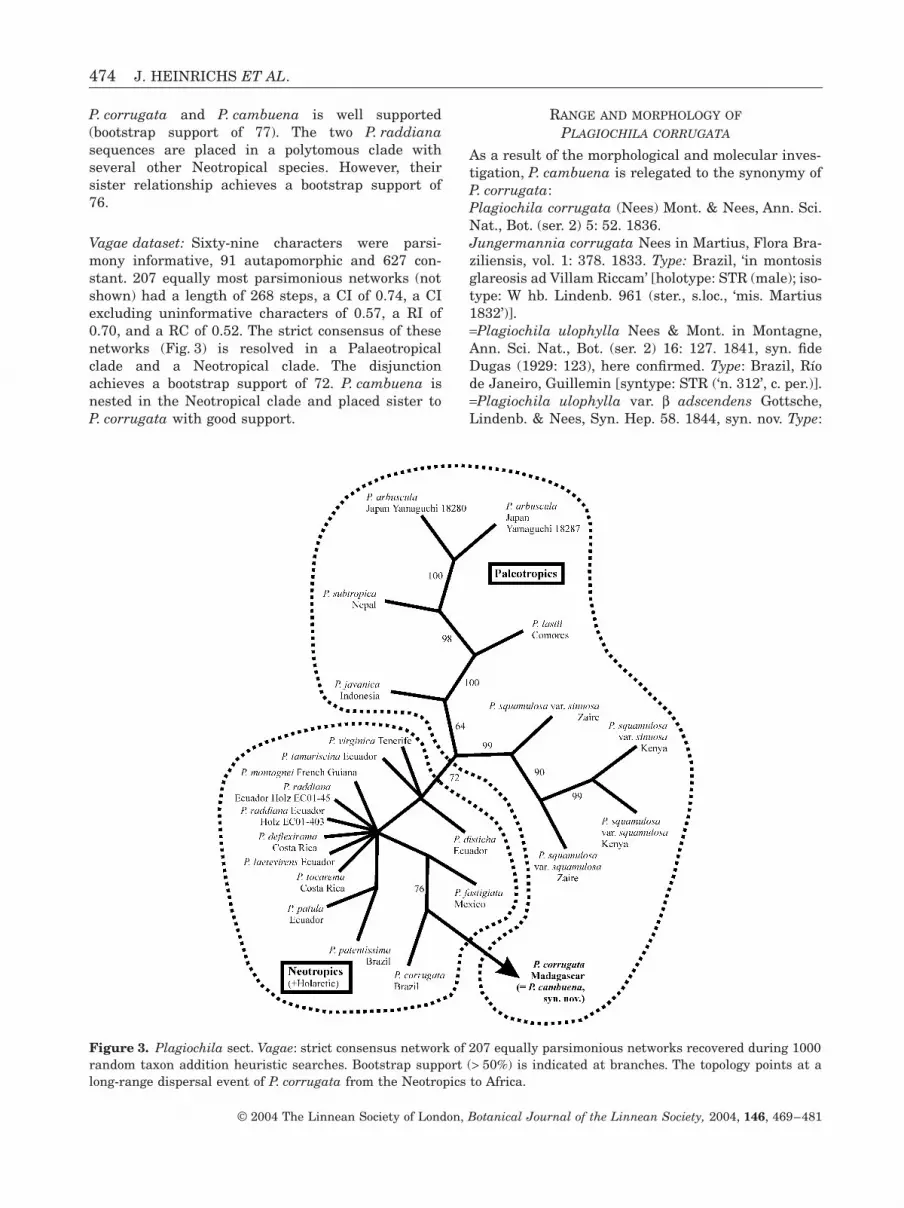
Vagae dataset: Sixty-nine characters were parsi-mony informative, 91 autapomorphic and 627 con-stant. 207 equally most parsimonious networks (notshown) had a length of 268 steps, a CI of 0.74, a CIexcluding uninformative characters of 0.57, a RI of0.70, and a RC of 0.52. The strict consensus of thesenetworks (Fig. 3) is resolved in a Palaeotropicalclade and a Neotropical clade. The disjunctionachieves a bootstrap support of 72. P. cambuena isnested in the Neotropical clade and placed sister toP. corrugata with good support.
RANGE AND MORPHOLOGY OF PLAGIOCHILA CORRUGATA
As a result of the morphological and molecular inves-tigation, P. cambuena is relegated to the synonymy ofP. corrugata:Plagiochila corrugata (Nees) Mont. & Nees, Ann. Sci.Nat., Bot. (ser. 2) 5: 52. 1836. Jungermannia corrugata Nees in Martius, Flora Bra-ziliensis, vol. 1: 378. 1833. Type: Brazil, ‘in montosisglareosis ad Villam Riccam’ [holotype: STR (male); iso-type: W hb. Lindenb. 961 (ster., s.loc., ‘mis. Martius1832’)].=Plagiochila ulophylla Nees & Mont. in Montagne,Ann. Sci. Nat., Bot. (ser. 2) 16: 127. 1841, syn. fideDugas (1929: 123), here confirmed. Type: Brazil, Ríode Janeiro, Guillemin [syntype: STR (‘n. 312’, c. per.)].=Plagiochila ulophylla var. b adscendens Gottsche,Lindenb. & Nees, Syn. Hep. 58. 1844, syn. nov. Type:
Figure 3. Plagiochila sect. Vagae: strict consensus network of 207 equally parsimonious networks recovered during 1000random taxon addition heuristic searches. Bootstrap support (> 50%) is indicated at branches. The topology points at along-range dispersal event of P. corrugata from the Neotropics to Africa.
INTERCONTINENTAL DISTRIBUTION OF PLAGIOCHILA CORRUGATA 475
Figure 4. Plagiochila corrugata (Nees) Mont. & Nees: A, spores; B, part of elater; C, top of female shoot with sporophyte,dorsal view; D, part of male plant with androecium, dorsal view; E, recurved part of ventral leaf base; F, cross section ofcapsule wall; G, innermost layer of capsule wall, surface view; H, epidermal layer of capsule wall, surface view; I, leafcells; J, leaves; K, underleaves [A–D, F–H from Grossmann 4 (GOET, Bryoph. Exs. General Plag. 88), E, I–K from isotypeof P. corrugata (W)].
Berghen (1981: 76) and here confirmed. Type:Madagascar, Hildebrandt s.n. [G 20896 (c.per.)].=Plagiochila corrugata (Nees) Mont. & Nees var. sub-integerrima Herzog, Arch. Bot. São Paulo (Brazil) 1:36, syn. nov. Type: Brazil, São Paulo, Butantan, 1921Gehrt 8133/447 [holotype: JE (male); Inoue (1989:144) cites Zikan as collector, however, no collector ismentioned in the protologue].
=Plagiochila corrugata (Nees) Mont. & Nees f. tenuiorHerzog, Feddes Rep. Spec. Nov Regni Veg. 55: 7. 1952‘1953’, nom. inval. [Art. 32.1 (c) ICBN], syn. nov. Orig-inal material: Argentina, Salta, Orán, El Gomal, RíoBermejo, 1933, Hosseus 34 A [JE (male)].
Description: Plants medium in size, c. (1–)2.5–7 cmlong and 2.5–5 mm broad when moistened, yellowish
Figure 5. Plagiochila corrugata (Nees) Mont. & Nees: A, cross section of capsule wall; B, innermost layer of capsule wall,surface view; C, capsule wall epidermis, surface view; D, spore; E, marginal teeth of ventral leaf base; F, underleaf; G,central part of elater; H, cells from leaf centre; I, J, leaves [A–I from Madagascar, Besson 304 (G), J from holotype ofP. cambuena (G)].
INTERCONTINENTAL DISTRIBUTION OF PLAGIOCHILA CORRUGATA 477
green, green or brownish green, in diffuse patches,with differentiation into creeping stoloniform shootsthat give rise to prostrate or ascending leafy stems.STEMS dark brown to brown, in upper part sometimesgreenish brown or greenish, densely covered by leavesboth dorsally and ventrally, main stems dorsoven-trally flattened, c. 300–450 ¥ 180–330 mm in diameter,in cross section c. 17–28 cells across, the corticalcells in (3–)4–6(-7) layers, thick-walled, c. 8–20 ¥14–25 mm, the medullary cells thin- to moderatelythick-walled, c. 14–24 ¥ 18–35 mm, often with small- tomedium-sized, triangular to subnodulose trigones.BRANCHES moderate to frequent in number (in weakerplants occasionally lacking), branching off from themore or less straight main stem (or the more or lessstraight primary branches), sometimes, especially inupper parts of the plants, bipinnate, giving the plant asomewhat pseudodichotomous habit; branches pre-dominantly terminal, of the Frullania-type, in lowerand medium parts plant sometimes with a few addi-tional lateral-intercalary branches.
Creeping stoloniform shoots often with short leaf-lets, aerial shoots with moderately to densely imbri-cate leaves. LEAVES wide spreading when moist; in drycondition usually somewhat postically secund or lat-erally appressed to stem; ventral margin moderatelyto very long decurrent, decurrent strip often broad,weakly to distinctly undulate, reflexed with tendencyto form an indistinct water-sack; dorsal marginmoderately to very long decurrent. Leaves ovate-triangular, ovate-oblong or ovate, ventral base ampli-ate, leaves c. (0.9–)1.1–2.1(-2.6) ¥ (0.9–)1.1–1.7(-2.1)mm, c. 0.8–1.3(-1.6) times as long as wide; leaf apexrounded to truncate; ventral margin straight orweakly curved outwards, usually strongly undulateand crispate; dorsal margin curved inwards orstraight, proximal part sometimes weakly curved out-wards, dorsal recurved or ± flat. Leaves toothed butdorsal margin often entire, occasionally only ventralleaf base with teeth or a few leaves completely entire;dorsal leaf margin with 0–8 forward pointed, ± trian-gular teeth, teeth 1–2(-3) cells broad at base, 1–2(-4)cells long, other teeth variable in shape, straight orcurved, pointing in various directions, lobe-like, trian-gular to elongate triangular, occasionally broad basedand narrowed to ciliate apical part (1–)2–8(-12) cellsbroad at base and (1–)2–5(-8) cells long, leaf apex with2–7 teeth, ventral leaf margin with (0–)2–23 teeth,ventral leaf base with 0–9 teeth. Leaf cell pattern reg-ular to irregular, cells in upper half of leaf slightlyshorter than broad to moderately elongate, c. 16–45 ¥ 14–36 mm and 0.8–1.5 times as long as wide,above stem in all more elongate, up to 65 mm long and2.6 times as long as wide, cells thin-walled, at leafmargins and ventral leaf base slightly to moderatelythickened, trigones (small to) medium to large in size,
(triangular) subnodulose or nodulose, intermediatethickenings occasionally present on long walls, trigo-nes often subconfluent, on long cell walls of decurrentventral leaf strip often subconfluent to confluent orintegrated in the ± regularly thickened cell walls;cuticle smooth. Oil bodies c. 4–7(-12) per medianleaf cell, ovoid to (linear-) ellipsoidal, colourless,coarsely few-segmented by rather large globules topapillose, 2–4 ¥ 3–8(-10) mm. Underleaves usuallypresent, large, c. 0.15–0.6(-1.1) mm long and 0.2–0.5 mm broad, usually divided into 2 lanceolate lobes,each with several cilia, or variously plurilaciniate.Asexual reproduction rarely by means of irregularlybranched, multicellular propagules developing intoplantlets on the ventral surface of the leaves.
Male plants as large as female plants. Androeciasimple, becoming intercalary on leafy branches as wellas on main stems, compact; bracts in (2–)4–9(-14)pairs, imbricate, opposite bracts overlapping on dorsalside of stem; basal part of bracts strongly inflated,mainly composed of hyaline and ± inflated cells withthin to moderately thickened walls without trigones orwith small triangular ones; distal part of bractsobliquely spreading, composed of cells similar to thoseof leaves; ventral margin often moderately to longdecurrent and somewhat reflexed; apex rounded totruncate or broadly and asymmetrically triangular,apex and ventral margin undulate/crispate, dentationsimilar to leaves. Antheridia 2–3 per bract, round tobroadly ellipsoidal, c. 250–330 mm in diameter, on along stalk.
Gynoecia terminal on branches, subgynoecial inno-vations frequent in absence of fertilization, usually1(-2) in number, bracts somewhat larger than theleaves below, adjacent to perianth and covering itnearly completely, ovate-triangular to oblong ovatewith a rounded to subtruncate apex, ventral and dor-sal margin short to moderately decurrent, dorsal mar-gin strongly recurved, ventral margin stronglyundulate, margins toothed, distal part of dorsalmargin often entire. Perianths broadly cylindrical tocampanulate in lateral view, inflated basally, often ±distinctly narrowed towards the mouth in dorsal view,c. 1.7–2.6 mm long and 1.5–2.4 mm broad, dorsallykeeled but keel usually unwinged; perianth mouthweakly arched upwards or truncate, with triangular toelongate triangular teeth.
CAPSULE short-exserted, globose, valves c. 1.3–1.6 ¥0.5–0.8 mm, brown, not twisted; epidermal cells insurface view shorter than broad to moderately elon-gate, towards base of valve more elongate, with 1–2large nodulose, occasionally somewhat cone-like thick-enings on many walls; innermost cells in surface viewelongate, irregular, with thickenings on the longitudi-nal and few transverse walls, opposite thickenings oflongitudinal walls connected by ± slender coalescences
on the radial walls, occasionally thickenings in partsof the valves nodulose to cone-like, discrete; valves incross section c. 40–70 mm thick, 4–5-stratose, epider-mal cells slightly larger than inner ones, c. 14–24 mmthick, inner cells c. 8–18 mm thick. SPORES c. 15–38 ¥18–52 mm, subglobose to short cylindrical, 1–4 cellularat time of release, sporoderm baculate. ELATERS
c. 8–12 mm thick, unispiral, in the middle rarelybispiral, covered by granulae.
BUCO: Caruaru, Brejo dos Cavalos, 1987, Pôrto21617 (GOET); RIO DE JANEIRO: Petropolis, 1924,Baudeira 343 (JE); RIO GRANDE DO SUL: Nova Würt-temberg, Elsenau, 450 m, 1904, Bornmüller s.n.,Bryoth. Levier 4627 (JE); SAO PAULO: Serra Negra,1927, Hoehne s.n. & 838 (JE). BOLIVIA. CHU-
QUISACA: Hernando Siles, 8 km west of Monteagudo,1190 m, 1984, Lewis 84–0920 (LPB); SANTA CRUZ:O’Connor, Río Tambo south-east of Narvaez, 2190 m,1984, Lewis 84–2633 (LPB), PARAGUAY. CAAGUAZÙ:Coronel Oviedo, Barrio Azucena, 1985, Bordas 237(GOET, NY). ARGENTINA. MISIONES: San Javier,San Isidro, Río Uruguay, 1933, Hosseus 509 & 551(JE).
AFRICA : COMORO ARCHIPELAGO. NGAZIDJA
ISLAND: Karthala volcano, 1992, Pócs et al. 9269/AV(EGR). MADAGASCAR. s.loc., Besson 304 (G);ANTANANARIVO: Angavo, between Ambatolaona andMandraka, 1350–1400 m a.s.l., 1998, Pócs & Szabó9868 (EGR, GOET), Andringitra Mts, 1000–1270 m,1994, Pócs 9473/AV (EGR, GOET). TANZANIA.MUFINDI ESCARPMENT: Lupembe Tea Estate, 1750–1800 m, 1970, Pócs & Jones 2073 (EGR).
Notes: Plagiochila corrugata is widespread in lowlandand lower montane regions of south-eastern Brazil,northern Argentina, Bolivia and Paraguay. Further-more the species occurs in Madagascar, the Comorosand in Tanzania (Fig. 6). In the Andes, P. corrugata isfound up to c. 2200 m a.s.l. Usually the species growsepiphytically.
DISCUSSION
Plagiochila sect. Vagae in the circumscription of Hein-richs et al. (2002b) is a large pantropical clade that ischaracterized morphologically by dominating termi-nal branching, vegetative reproduction by propagulesand plantlets from the leaf surface, a capsule wall withthickenings in all layers, and simple, intercalaryandroecia with bracts overlapping dorsally. Severaldozen species are accepted in Vagae (e.g. Jones, 1962;Inoue, 1984; So, 2000, 2001), being presumably thelargest section of Plagiochila. Groth et al. (2003, 2004)have demonstrated that the Vagae are nested in alarge clade with Plagiochila sections from tropical
Figure 6. Geographical distribution of Plagiochila corrugata (Nees) Mont. & Nees.
INTERCONTINENTAL DISTRIBUTION OF PLAGIOCHILA CORRUGATA 479
Asia and the Holarctic. Accordingly, the pantropicalVagae most likely descended from Asian ancestors andreached Africa and America later. The topologies inFigures 1 and 2 also point to a palaeotropical origin ofP. sect. Vagae.
Twelve Vagae species from the Neotropics and theHolarctic form a well supported cluster in theunrooted tree in Figure 3. Nearly no resolution wasattained at species level, although the complete Neo-tropical Vagae phenotype spectrum with severalextreme morphotypes was included in the ITSsequence analyses (Figs 1–3). Obviously a rapid radi-ation is at hand, in this case possibly forced by the fre-quent vegetative distribution. Similar observationshave been made within an Andean species complex ofP. sect. Hylacoetes including the former genera Steere-ochila Inoue and Szweykowskia Gradst. & M.Reiner(Heinrichs et al., 2003) or within P. sect. Arrectae Carl(e.g. Heinrichs et al., 2004).
The tropical American-African Plagiochila corru-gata is nested within the Neotropical species of sect.Vagae (Figs 1–3). The low ITS sequence variationwithin this group may be explained by the disjunctrange of P. corrugata being the result of long rangedispersal rather than a Gondwana distribution. Judg-ing from the analysed taxon set, dispersal originatedfrom the Neotropics. Similar ranges are not unknownin Plagiochila and have been documented forP. boryana Steph., the only African representative ofthe Neotropical P. sect. Hylacoetes (Heinrichs, 2002;Groth et al., 2003), which occurs in Bolivia, Brazil,Uganda, Tanzania and on the Mascarenes. The distri-bution of P. corrugata is not unique among the Afro-American disjuncts. Widespread in South Americaand on Madagascar Island, the species occurs onlysporadically in the crystalline Eastern Arc mountainsof Tanzania: Usambara Mountains and MufindiEscarpment (Pócs, 1985). The moss Adelotheciumbogotense (Hampe) Mitt. is also widespread in theNeotropics, has three localities in Madagascar and onein the Uluguru Mountains of the Tanzanian EasternArc (Ochyra et al., 1992 and T. Pócs, unpubl. data).The link between the Eastern Arc and Madagascar iswell known and supported by a high number of speciesrestricted to these areas (Pócs, 1975, 1999a) Anotherinteresting type of distribution both at generic and atspecies level is a disjunction between the Neotropicsand Madagascar without stands in continental Africa(e.g. Bryopteris (Nees) Lindenb., Symbyezidium Tre-vis, Xylolejeunea X.-L. He & Grolle, PhyllogoniumBrid.; Gradstein, Pócs & Vána, 1983; Pócs, 1999b;Sass-Gyarmati & Pócs, 2002). The distribution ofP. corrugata and of Adelothecium bogotense seems tobe a combination of the two types.
Disjunct Africa-tropical America ranges of bryo-phytes have been postulated as a result of morpholog-
ical analyses but have rarely been checked bysequence comparisons (Symphogyna brasiliensis Nees& Mont., S. podophylla (Thunb.) Mont. & Nees;Schaumann et al., 2003). Shaw (2001) points out thatmorphological uniformity of bryophytes may belie acomplex genetic structure. However, the results of ourmolecular analyses are in good accordance with mor-phological species and section concepts. Monophyleticlineages for morphologically inseparable units are nota general phenomenon in bryophytes. For instance,morphospecies of Fontinalis Hedw. are non-monophyl-etic in nr and in cpDNA sequence trees (Shaw & Allen,2000).
The level of ITS sequence variation in P. corrugatais not dissimilar to that found in other morphologicallyand phylogenetically defined species of Plagiochila(e.g. P. bifaria (Sw.) Lindenb.; Heinrichs et al. 2004;P. carringtonii (Balf.) Grolle; Renker et al. 2002) or ofmosses (Shaw, Werner & Ros, 2003). Infraspecificlength variation of ITS sequences as documented herefor P. corrugata is a common phenomenon in Plagio-chila (e.g. Groth et al., 2003; Heinrichs et al., 2003)and often the result of indel events in variable loopregions.
The considerable ITS variation within the Vagaeclade leads only to a poor resolution, most likely a con-sequence of the diffuse dispensation of variable posi-tions and the high degree of autapomorphiccharacters. However, the largely unresolved Neotropi-cal Vagae clade with several extreme morphotypespermits the establishment of a wide species conceptfor this group in the forthcoming Flora Neotropicatreatment (J. Heinrichs, unpubl. data). Herzog (1932)has already indicated that nearly unmanageablegametophytic variation is found in the group and thatthe determinations found in the herbaria cause confu-sion rather than help with understanding the species.Obviously leaf shape and dentation have limited tax-onomic value in Plagiochila, implying that most Neo-tropical Vagae binomials should be placed in thesynonymy of a few broadly defined species. The resultsconcerning the African P. squamulosa s.l. point in thesame direction.
The unispiral elaters of P. corrugata justify separa-tion at species level from the morphologically closelyrelated P. raddiana Lindenb. with bispiral elaters(Heinrichs & Gradstein, 2000). The latter otherwisediffers from P. corrugata by the somewhat less undu-late leaf margins and the often entire ventral leafbases. Morphologically closely related is also a poorlyunderstood Central and North American species com-plex including P. undata Sull., P. crispata Gottsche,P. fastigiata, P. sancta Gottsche (cf., e.g. Gottsche,1863–7; Schuster, 1980). These binomials differ mostlyin the entire ventral leaf bases and are placed sister toP. corrugata in the molecular analyses (P. fastigiata).
Sporophytes have not yet been available for study.However, Gottsche (1863–7) points out that P. sanctahas bispiral and partly unispiral elaters, which indi-cates that the above binomina could be included inP. corrugata at the variety or subspecies level at somefuture point.
ACKNOWLEDGEMENTS
We would like to thank the curators of the herbariacited in the text for the loan of material as well asMichael Lüth (Freiburg), Volker Buchbender, Eber-hard Fischer and Andreas Solga (Koblenz) for dupli-cates of Plagiochila specimens. May-Ling So (HongKong) kindly confirmed the identification of the Pla-giochila javanica DNA voucher. Financial support ofthe Deutsche Forschungsgemeinschaft (grant HE3584/1) and the Hungarian Research Fund (OTKA,grant U 1861) is gratefully acknowledged.
REFERENCES
Carl H. 1931. Die Arttypen und die systematische Gliederungder Gattung Plagiochila Dum. Annales Bryologici, Supple-mentum 2: 1–170.
Dugas M. 1929. Contribution a l’etude du genre ‘Plagiochila’Dum. Annales des sciences naturelles Botanique, Série 10(11): 1–199.
Felsenstein J. 1985. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39: 783–791.
Gottsche CM. 1863–7. De Mexicanske Levermosser. BescrevneEfter Prof. Fr. Liebmanns Samling. Copenhagen: BiancoLuna.
Gradstein SR, Pócs T, Vána J. 1983. Disjunct Hepaticae intropical America and Africa. Acta Botanica Hungarica 29:127–171.
Groth H, Heinrichs J. 2003. Reinstatement of ChiastocaulonCarl (Plagiochilaceae), based on evidence from nuclear ribo-somal ITS and chloroplast gene rps4 sequences. Plant Biol-ogy 5: 615–622.
Groth H, Lindner M, Wilson R, Hartmann FA,Schmull M, Gradstein SR, Heinrichs J. 2003. Bioge-ography of Plagiochila (Hepaticae): natural species groupsspan several floral kingdoms. Journal of Biogeography 30:965–978.
Groth H, Lindner M, Heinrichs J. 2004. Phylogeny and bio-geography of Plagiochila based on nuclear and chloroplastDNA sequences. Molecular systematics of bryophytes.Monographs in Systematic Botany from the Missouri Botan-ical Garden 98: 365–387.
Hall TA. 1999. BioEdit: a user-friendly biological sequencealignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposia Series 41: 95–98.
Heinrichs J. 2002. A taxonomic revision of Plagiochila sect.Hylacoetes, sect. Adiantoideae and sect. Fuscoluteae in theNeotropics with a preliminary subdivision of NeotropicalPlagiochilaceae into nine lineages. Bryophytorum Biblioth-eca 58: 1–184, Append. 1–5.
Heinrichs J, Gradstein SR. 2000. A revision of Plagiochilasect. Crispatae and sect. Hypnoides (Hepaticae) in the Neo-tropics. I. Plagiochila disticha, P. montagnei and P. raddi-ana. Nova Hedwigia 70: 161–184.
Heinrichs J, Pröschold T, Renker C, Groth H, RycroftDS. 2002a. Plagiochila virginica A.Evans rather than Pla-giochila dubia Lindenb. & Gottsche occurs in Macaronesia;placement in sect. Contiguae Carl is supported by ITSsequences of nuclear ribosomal DNA. Plant Systematics andEvolution 230: 221–230.
Heinrichs J, Sauer M, Grolle R. 2002b. Lectotypificationand synonymy of Plagiochila sect. Vagae Lindenb. (Hepati-cae). Cryptogamie, Bryologie 23: 5–9.
Heinrichs J, Gradstein SR, Groth H, Lindner M. 2003.Plagiochila cucullifolia var. anomala var. nov. from Ecuador,with notes on discordant molecular and morphological vari-ation in Plagiochila. Plant Systematics and Evolution 242:205–216.
Heinrichs J, Lindner M, Pócs T. 2004. Nr DNA internaltranscribed spacer data reveal that RhodoplagiochilaR.M.Schust. (Jungermanniales, Marchantiophyta) is a mem-ber of Plagiochila sect. Arrectae Carl. Organisms, Diversityand Evolution 4: 109–118.
Herzog T. 1932. Beiträge zur Kenntnis der Gattung Plagio-chila. I. Neotropische Arten. Hedwigia 72: 195–242.
Inoue H. 1984. The genus Plagiochila in Southeast Asia.Tokyo: Academic Scientific Book Inc.
Inoue H. 1989. Plagiochila. In: Geissler P, Bischler H, eds.Naiadea to Pycnoscenus, Index Hepaticarum 11. Berlin:Cramer, 106–237.
Jones EW. 1962. African hepatics. XV. Plagiochila in tropicalAfrica. Transactions of the British Bryological Society 4: 254–325.
Ochyra R, Bednarek-Ochyra H, Pócs T, Crosby M. 1992.The moss Adelothecium bogotense in Continental Africa,with a review of its world range. Bryologist 95: 287–295.
Pócs T. 1975. Affinities between the bryoflora of East Africaand Madagascar. In: Miège J, Stork A, eds. Origines desFlores africaines et malgaches. Nature, Spéciation: Comptes-rendus de la VIIIe réunion de l’AETFAT 1. Boissiera 24a:125–128.
Pócs T. 1985. East African Bryophytes, VII. The Hepaticae ofthe Usambara Rain Forest Project Expedition, 1982. ActaBotanica Academiae Scientiarum Hungarica 31: 113–133.
Pócs T. 1999a. Bryophyte speciation and diversity in the EastAfrican mountains. Bryobrothera 5: 237–245.
Pócs T. 1999b. Trachylejeunea grolleana, a new representa-tive of the Neotropical subgenus Hygrolejeuneopsis in Mada-gascar. Haussknechtia Beiheft 9: 283–290.
Posada D, Crandall KA. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14: 817–818.
Renker C, Heinrichs J, Pröschold T, Groth H, Holz I.2002. ITS sequences of nuclear ribosomal DNA support thegeneric placement and the disjunct range of Plagiochila(Adelanthus) carringtonii. Cryptogamie, Bryologie 23: 23–29.
Sass-Gyarmati A, Pócs T. 2002. A second species of Symbiez-idium from Africa. Cryptogamie, Bryologie 23: 31–39.
INTERCONTINENTAL DISTRIBUTION OF PLAGIOCHILA CORRUGATA 481
Schaumann F, Frey W, Hässel de Menéndez G, Pfeiffer T.2003. Geomolecular divergence in the Gondwanan dendroidSymphogyna complex (Pallaviciniaceae, Hepaticophytina,Bryophyta). Flora 198: 404–412.
Schuster RM. 1980. The Hepaticae and Anthocerotae of NorthAmerica 4. New York: Columbia University Press.
Shaw AJ. 2001. Biogeographic patterns and cryptic speciationin bryophytes. Journal of Biogeography 28: 253–261.
Shaw AJ, Allen B. 2000. Phylogenetic relationships,morphological incongruence, and geographic speciation inthe Fontinalaceae (Bryophyta). Molecular Phylogenetics andEvolution 16: 225–237.
Shaw AJ, Werner O, Ros RM. 2003. Intercontinental Medi-terranean disjunct mosses: morphological and molecularpatterns. American Journal of Botany 90: 540–550.
So M-L. 2000. Plagiochila sect. Contiguae (Hepaticae) inAustralasia and the Pacific, with description of Plagiochilasubjavanica sp. nov. Australian Systematic Botany 13: 803–815.
So M-L, Grolle R. 2000. Checklist of Plagiochila (Hepaticae)in Asia. Journal of the Hattori Botanical Laboratory 88: 199–243.
Spruce R. 1884–5. Hepaticae of the Amazon and the Andes ofEcuador and Peru. Transactions and Proceedings of theBotanical Society of Edinburgh 15: i–xi, 1–589, plates I–xxii.
Swofford DL. 2003. PAUP*: phylogenetic analysis using par-simony (*and other methods). Sunderland, MA: SinauerAssociates.
Tamura K, Nei M. 1993. Estimation of the number of nucle-otide substitutions in the control region of mitochondrialDNA in humans and chimpanzees. Molecular Biology andEvolution 10: 512–526.
Vanden Berghen C. 1981. Le genre Plagiochila (Dumort.)Dumort. (Hepaticae) à Madagascar et aux Mascareignes,principalement d’apres les recoltes de M. Onraedt. Bulletindu Jardin Botanique National de Belgique 51: 41–103.