Introduction to a Virtual Special Issue on plant ecological strategy axes in leaf and wood traits
Moving beyond the economics spectrum?
The study of plant ecological strategy axes has emerged as animportant area of research in the past decade. A strategy axisis defined by suites of covarying traits that are studied acrossspecies. One of the axes that has received considerable attentionis the suite of traits related to photosynthesis and leaf life span,recently dubbed the ‘leaf economics spectrum’ (Wright et al.,2004), with a continuum of variation from species with short-lived, high specific leaf area (SLA) leaves of high nutrientconcentrations and photosynthetic rates, to those with theopposite set of trait values at the other end. The basic contrastof these photosynthetic strategies was initially recognized incomparisons of evergreen vs deciduous species (e.g. Mooney& Dunn, 1970). Kikuzawa (1991) and others then introducedthe critical distinction between leaf life span (the length oftime that an individual leaf persists on a tree) and leaf habit(whether all leaves fall off the canopy at any one time). Reichet al. (1997) were the first to recognize clearly the convergencein leaf function across diverse ecosystems, setting the stagefor the global synthesis described by Wright et al. Traitscomprising the woody tissue of plants, while long appreciatedfor their structural and hydraulic importance (Tyree & Ewers,1991; Niklas, 1992), have not been as successfully placedwithin a general predictive framework.
The first set of papers in the Virtual Special Issue of NewPhytologist on plant ecological strategy axes continue to probethe generality of the physiological mechanisms that generatetrade-offs in leaf form and function, and their broader ecologicaland evolutionary implications. He et al. (2006) addressed agap in global knowledge, studying several dozen species of theTibetan plateau. They found that the overall pattern of traitcorrelations was similar at this climatic extreme, further sup-porting the generality of the relationships, although photo-synthetic rates were slightly lower relative to leaf nitrogen.Wright et al. (2006) assessed patterns in leaf dark respiration,and reported shifts in respiration relative to other traits as afunction of irradiance and site temperature. The temperatureeffects, however, appear to be direct effects on respiration, asthere are no detectable differences when respiration is measuredat a common temperature. Leishman et al. (2007) examined
data on native vs invasive species, and found that invasives areshifted towards the ‘fast’ end of the spectrum, along the sharedaxis of trait covariation. In an interesting twist, Lusk & Warton(2007) have discovered that the relationship between leaf traitsand regeneration strategies in forest plants depend on leaf habitand ontogeny. In deciduous plants, the correlations betweenleaf mass per area (the inverse of SLA) and shade toleranceshift between seedlings and saplings, and in different lightenvironments, suggesting a more complex set of growth strategiesin strongly seasonal environments.
In comparative studies, leaf life span is viewed as a speciestrait, and metabolic traits are generally reported for newlymatured leaves. Within plants, leaf life span varies among leavesand reflects the dynamics of self-shading, nitrogen resorptionand leaf carbon balance. Oikawa et al. (2006) examine therole of soil fertility on internal canopy dynamics, and Pornon& Lamaze (2007) consider variation in leaf life span withinthe canopy, and the relative efficiency of nutrient use amongdifferent sets of leaves. These papers suggest an importantavenue for future research to integrate more thoroughly ourunderstanding of within-canopy nitrogen allocation and leaffunction with the broad patterns of interspecific variation andecological strategies.
Studies of leaf function in an evolutionary and comparativeframework provide an important perspective on our knowledgeof ecological strategies. Karst & Lechowicz (2007) examinedleaf traits of temperate, understory ferns, and found that theyfall within the envelopes reported for seed plants at a globalscale, again supporting the generality of the overall relationships.Watanabe et al. (2007) examined leaf nutrient concentrationsin a taxonomic framework, and found that about 25% of thevariation in various nutrients is associated with plant family.The significance of such patterns in relation to the evolutionof leaf function through time remains virtually unexplored.
The structural and functional properties of wood have alsoreceived considerable recent attention from plant biologistsexploring the possibilities of adaptive trade-offs or patterns ofcoordinated evolution among the anatomical and physiologicaltraits making up this complex tissue. Are there broadlygeneralizable strategy axes among woody tissue traits analogousto those of the leaf economics spectrum? Woody growth servesthree primary roles in plants: conducting water and nutrientsfrom root to shoot; providing structural support for leaves;and serving as a storage reservoir. The cellular building blocksall plants must work with in growing wood are the hollowconducting elements (xylem vessels and tracheids), thick-walledfibers and parenchyma cells. If the numbers and dimensionsof these cells can vary independently, a great variety in woodanatomy is possible, with a correspondingly wide range in
stem structural and functional performance. For example,wood composed primarily of large-diameter, thin-walledvessels bears a low carbon construction cost and has high water-conducting capacity, but provides relatively weak structuralsupport and low resistance to drought-induced rupture of thewater column (embolism). Reducing vessel diameter and addingfibers and parenchyma all increase wood density and bothstructural and functional integrity but at the cost of reducedhydraulic conductivity, higher construction costs and lowerwhole-plant growth rate.
The second group of papers in this Virtual Special Issueall consider aspects of the structural and water-conductingproperties of wood and their variation within and across plantcommunities. Working in the wet tropical forest of Bolivia,van Gelder et al. (2006) examined the mechanical property ofwood from 30 species with contrasting juvenile-stage lightrequirements, finding consistent differences between shade-tolerant species (dense and tough wood) and pioneer species(little structural margin of safety from breakage). In an accom-panying commentary, Falster (2006) presents an overview ofthe engineering principles involved in wood biomechanicsand puts variation in wood density in a broader context ofvariation in other traits of importance to plant performancein the tropical forest understory, such as seed size and leafeconomics. Moving to the Mediterranean environment of coastalCalifornia and a different set of environmental demands onwoody plant vasculature, Preston et al. (2006) analyzed patternsof variation in xylem vessel density and lumen area in 51 species,ranging from vines to trees. Their results, including evidencefrom phylogenetic independent contrasts, suggested opposingevolutionary trends in these two key determinants of wooddensity. Within a much narrower phylogenetic range (ninespecies of Rhamnaceae from California), Pratt et al. (2007)found a positive relationship between the evolution of highstem wood density and resistance to embolism under waterstress. Interestingly, there was no apparent trade-off betweenwood density and hydraulic conductance in these species.Anfodillo et al. (2006), in a survey of 30 species, providebroad support for the model of West et al. (1999), predictingchanges in vessel lumen area with tree height that minimizeconduit length effects on hydraulic conductance.
How have wood vascular traits evolved in response tochanges in water availability? Three final papers presented hereaddress this question, taking advantage of known phylogeniesof species growing across distinct precipitation gradients.Among nine Cordia species from the neotropics, Choat et al.(2007) reported greater resistance to embolism in drier sitespecies, but interspecific variation in other wood traits thatwas independent of mean annual precipitation. A similarresult was reported by Bhaskar et al. (2007) among six speciespairs of chaparral shrubs from California and Mexico. InPereskia, a basal group of Cactacea lacking stem succulence,Edwards (2006) found strong support for correlated evolutionamong several stem and leaf traits of importance in plant
water relations but found no evidence for any trait correla-tions with habitat water availability. These results highlight,yet again, the importance of taking phylogeny into accountwhen making inferences from comparative studies aboutadaptive trait evolution.
Collectively, these papers affirm the predictive value of theunderlying relationships among plant functional traits, andthe utility of the leaf economic spectrum as a framework tounderstand variation among sites and among different groupsof species. The substantive progress relating wood anatomicaltraits to plant function and ecological adaptation suggests thatadditional strategy axes soon may be uncovered to complementthis framework.
Peter S. Curtis and David D. Ackerly
Editors
References
Anfodillo T, Carraro V, Carrer M, Fior C, Rossi S. 2006. Convergent tapering of xylem conduits in different woody species. New Phytologist 169: 279–290.
Bhaskar R, Valiente-Banuet A, Ackerly DD. 2007. Evolution of hydraulic traits in closely related species pairs from mediterranean and nonmediterranean environments of North America. New Phytologist 176: 718–726.
Choat B, Sack L, Holbrook NM. 2007. Diversity of hydraulic traits in nine Cordia species growing in tropical forests with contrasting precipitation. New Phytologist 175: 686–698.
Edwards EJ. 2006. Correlated evolution of stem and leaf hydraulic traits in Pereskia (Cactaceae). New Phytologist 172: 479–489.
Falster DS. 2006. Sapling strength and safety: the importance of wood density in tropical forests. New Phytologist 171: 237–239.
van Gelder HA, Poorter L, Sterck FJ. 2006. Wood mechanics, allometry, and life-history variation in a tropical rain forest tree community. New Phytologist 171: 367–378.
He JS, Wang ZH, Wang XP, Schmid B, Zuo WY, Zhou M, Zheng CY, Wang MF, Fang JY. 2006. A test of the generality of leaf trait relationships on the tibetan plateau. New Phytologist 170: 835–848.
Karst AL, Lechowicz MJ. 2007. Are correlations among foliar traits in ferns consistent with those in the seed plants? New Phytologist 173: 306–312.
Kikuzawa K. 1991. A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. The American Naturalist 138: 1250–1263.
Leishman MR, Haslehurst T, Ares A, Baruch Z. 2007. Leaf trait relationships of native and invasive plants: community- and global-scale comparisons. New Phytologist 176: 635–643.
Lusk CH, Warton DI. 2007. Global meta-analysis shows that relationships of leaf mass per area with species shade tolerance depend on leaf habit and ontogeny. New Phytologist 176: 764–774.
Mooney HA, Dunn EL. 1970. Photosynthetic systems of Mediterranean-climate shrubs and trees of California and Chile. The American Naturalist 104: 447–453.
Niklas KJ. 1992. Plant biomechanics: an engineering approach to plant form and function. Chicago, IL, USA: University of Chicago Press.
Oikawa S, Hikosaka K, Hirose T. 2006. Leaf lifespan and lifetime carbon balance of individual leaves in a stand of an annual herb, xanthium canadense. New Phytologist 172: 104–116.
Pornon A, Lamaze T. 2007. Nitrogen resorption and photosynthetic activity over leaf life span in an evergreen shrub, rhododendron ferrugineum, in a subalpine environment. New Phytologist 175: 301–310.
Pratt RB, Jacobsen AL, Ewers FW, Davis SD. 2007. Relationships among xylem transport, biomechanics and storage in stems and roots of nine Rhamnaceae species of the California chaparral. New Phytologist 174: 787–798.
Preston KA, Cornwell WK, DeNoyer JL. 2006. Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms. New Phytologist 170: 807–818.
Reich PB, Walters MB, Ellsworth DS. 1997. From tropics to tundra: global convergence in plant functioning. Proceedings of the National Academy of Sciences, USA 94: 13730–13734.
Tyree MT, Ewers FW. 1991. Tansley Review, 34: the hydraulic architecture of trees and other woody plants. New Phytologist 119: 345–360.
Watanabe T, Broadley M, Jansen S, White P, Takada J, Satake K, Takamatsu T, Tuah S, Osaki M. 2007. Evolutionary control of leaf element composition in plants. New Phytologist 174: 516–523.
West GB, Brown JH, Enquist BJ. 1999. A general model for the structure and allometry of plant vascular systems. Nature 400: 664–667.
Wright IJ, Reich PB, Atkin OK, Lusk CH, Tjoelker MG, Westoby M. 2006. Irradiance, temperature and rainfall influence leaf dark respiration in woody plants: evidence from comparisons across 20 sites. New Phytologist 169: 309–319.
Wright IJ, Westoby M, Reich PB, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin FS, Cornelissen J, Diemer M et al. 2004. The leaf economic spectrum world-wide. Nature 428: 821–827.
Key words: covarying traits, leaf economics spectrum, leaf habit, leaf life span, photosynthesis, plant evolution, specific leaf area (SLA), strategy axis.259610.1111/j. 1469-8137.2008.02596.xJuly 200800901???903???CommentaryCommentary
Commentary
Commentary
Living with salinity
Halophytes are plants that can tolerate saline soils, and cancomplete their life cycle in soils with salinity concentrationsabove 200 mm NaCl (Flowers & Colmer, this issue, pp. 945–963). Some species require salt concentrations of this order foroptimal growth, and grow poorly without it. The mechanismsthat allow this remarkable adaptation are largely unknown, sothe review by Flowers & Colmer is timely.
Halophytes constitute 1% of the world’s flora, and arefound in diverse taxonomic groups, and in a wide range ofenvironments from arid land to coastal marshes. Manyhalophytes grow in waterlogged or flooded soils, and seagrassesare totally immersed in seawater. When salinity and waterloggingoccur in combination, the effects on most plants can be severe(Barrett-Lennard, 2003), yet many halophytes can toleratethis combination. The mechanisms associated with floodingtolerance are the subject of the companion review by Colmer& Flowers, also in this issue (pp. 964–974).
‘It is curious that no monocotyledonous species is able
to benefit from the presence of salinity to the same
extent as dicotyledonous species.’
The species that are most tolerant of salinity, and show amarked growth improvement when the salinity increases, are
all dicotyledonous species. It is curious that no monocotyle-donous species is able to benefit from the presence of salinityto the same extent. This phenomenon has not been explained.Do dicotyledonous and monocotyledonous species use verydifferent mechanisms? In their review, Flowers & Colmerstress the importance of choosing several contrasting ‘model’species for further research into mechanisms of salinity tolerance,to cover the mechanistic diversity that exists across a spectrumof halophytes. However, we suggest that fastest progress canbe made by initially focusing on comparisons between speciesthat are genetically closely related but where one is tolerant ofsalinity and the other is not.
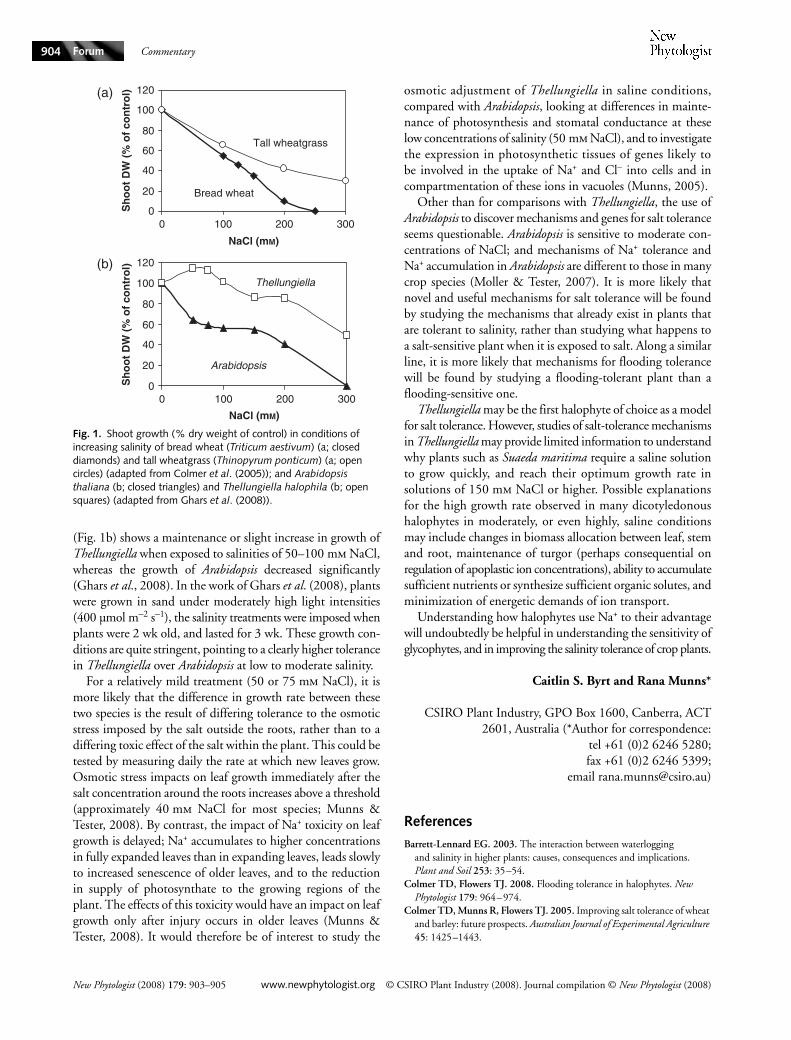
Differences between halophytes and glycophytes are illustratedin Fig. 1, using a pair of closely related monocotyledonous speciesand a pair of closely related dicotyledenous species. The exampleschosen are wheat and its halophytic relative, and Arabidopsisand its halophytic relative. The typical monocotyledonousresponse is shown by the salt-sensitive wheat (Triticum aestivum)and the salt-tolerant tall wheatgrass (Thinopyrum ponticum).Moderate salinity reduces growth of both species, but highersalinities (over 150 mm NaCl) reveal differences (Fig. 1a). Incontrast, a large difference between the salt-tolerant Thellungiellahalophila and salt-sensitive Arabidopis thaliana is evident inlow salinity, around 50 mm NaCl (Fig. 1b).
Thellungiella, although it is not especially salt-tolerant,appears to us to be the halophyte of choice as a model plant,owing to its fast generation time, ease of transformation, publiclyavailable genetic information, and because, compared with mostother halophytes, it is easy to grow in laboratory conditions.The growth of Thellungiella is illustrated as dropping markedlywith increasing salinity in the comparative salinity tolerancediagram presented by Flowers & Colmer, particularly in com-parison with other much more salt-tolerant species. However,a recent comparison between Thellungiella and Arabidopsis
(Fig. 1b) shows a maintenance or slight increase in growth ofThellungiella when exposed to salinities of 50–100 mm NaCl,whereas the growth of Arabidopsis decreased significantly(Ghars et al., 2008). In the work of Ghars et al. (2008), plantswere grown in sand under moderately high light intensities(400 µmol m−2 s−1), the salinity treatments were imposed whenplants were 2 wk old, and lasted for 3 wk. These growth con-ditions are quite stringent, pointing to a clearly higher tolerancein Thellungiella over Arabidopsis at low to moderate salinity.
For a relatively mild treatment (50 or 75 mm NaCl), it ismore likely that the difference in growth rate between thesetwo species is the result of differing tolerance to the osmoticstress imposed by the salt outside the roots, rather than to adiffering toxic effect of the salt within the plant. This could betested by measuring daily the rate at which new leaves grow.Osmotic stress impacts on leaf growth immediately after thesalt concentration around the roots increases above a threshold(approximately 40 mm NaCl for most species; Munns &Tester, 2008). By contrast, the impact of Na+ toxicity on leafgrowth is delayed; Na+ accumulates to higher concentrationsin fully expanded leaves than in expanding leaves, leads slowlyto increased senescence of older leaves, and to the reductionin supply of photosynthate to the growing regions of theplant. The effects of this toxicity would have an impact on leafgrowth only after injury occurs in older leaves (Munns &Tester, 2008). It would therefore be of interest to study the
osmotic adjustment of Thellungiella in saline conditions,compared with Arabidopsis, looking at differences in mainte-nance of photosynthesis and stomatal conductance at theselow concentrations of salinity (50 mm NaCl), and to investigatethe expression in photosynthetic tissues of genes likely tobe involved in the uptake of Na+ and Cl– into cells and incompartmentation of these ions in vacuoles (Munns, 2005).
Other than for comparisons with Thellungiella, the use ofArabidopsis to discover mechanisms and genes for salt toleranceseems questionable. Arabidopsis is sensitive to moderate con-centrations of NaCl; and mechanisms of Na+ tolerance andNa+ accumulation in Arabidopsis are different to those in manycrop species (Moller & Tester, 2007). It is more likely thatnovel and useful mechanisms for salt tolerance will be foundby studying the mechanisms that already exist in plants thatare tolerant to salinity, rather than studying what happens toa salt-sensitive plant when it is exposed to salt. Along a similarline, it is more likely that mechanisms for flooding tolerancewill be found by studying a flooding-tolerant plant than aflooding-sensitive one.
Thellungiella may be the first halophyte of choice as a modelfor salt tolerance. However, studies of salt-tolerance mechanismsin Thellungiella may provide limited information to understandwhy plants such as Suaeda maritima require a saline solutionto grow quickly, and reach their optimum growth rate insolutions of 150 mm NaCl or higher. Possible explanationsfor the high growth rate observed in many dicotyledonoushalophytes in moderately, or even highly, saline conditionsmay include changes in biomass allocation between leaf, stemand root, maintenance of turgor (perhaps consequential onregulation of apoplastic ion concentrations), ability to accumulatesufficient nutrients or synthesize sufficient organic solutes, andminimization of energetic demands of ion transport.
Understanding how halophytes use Na+ to their advantagewill undoubtedly be helpful in understanding the sensitivity ofglycophytes, and in improving the salinity tolerance of crop plants.
Caitlin S. Byrt and Rana Munns*
CSIRO Plant Industry, GPO Box 1600, Canberra, ACT2601, Australia (*Author for correspondence:
Barrett-Lennard EG. 2003. The interaction between waterlogging and salinity in higher plants: causes, consequences and implications. Plant and Soil 253: 35–54.
Colmer TD, Flowers TJ. 2008. Flooding tolerance in halophytes. New Phytologist 179: 964–974.
Colmer TD, Munns R, Flowers TJ. 2005. Improving salt tolerance of wheat and barley: future prospects. Australian Journal of Experimental Agriculture 45: 1425–1443.
Fig. 1. Shoot growth (% dry weight of control) in conditions of increasing salinity of bread wheat (Triticum aestivum) (a; closed diamonds) and tall wheatgrass (Thinopyrum ponticum) (a; open circles) (adapted from Colmer et al. (2005)); and Arabidopsis thaliana (b; closed triangles) and Thellungiella halophila (b; open squares) (adapted from Ghars et al. (2008)).
Flowers TJ, Colmer TD. 2008. Salinity tolerance in halophytes. New Phytologist 179: 945–963.
Ghars MA, Parre E, Debez A, Bordenave M, Richard L, Leport L, Bouchereau A, Savoure A, Abdelly C. 2008. Comparative salt tolerance analysis between Arabidopsis thaliana and Thellungiella halophila, with special emphasis on K+/Na+ selectivity and proline accumulation. Journal of Plant Physiology 165: 588–599.
Moller IS, Tester M. 2007. Salinity tolerance in Arabidopsis: a good model for cereals? Trends in Plant Science 12: 534–540.
Munns R. 2005. Genes and salt tolerance: bringing them together. New Phytologist 167: 645–663.
Munns R, Tester M. 2008. Mechanisms of salinity tolerance. Annual Review of Plant Biology 59: 651–681.
Water vapour loss is often regarded as a necessary evil fororganisms exchanging O2 and CO2 with the atmosphere. Asthe gases are metabolized in solution inside cells, there has tobe a wet cell-surface across which the gases exchange, andwater vapour is lost to the almost invariably unsaturatedatmosphere. In plants the latent heat of evaporation of watercomes in part from the light energy unavoidably dissipated inphotosynthesis. As an unavoidable process, there have beensuggestions of additional functions of transpiration, especiallyin view of the very large range of transpiratory water losses perunit growth among C3 vascular plants and the occurrence oftranspiration in the dark in C3 and C4 plants that do not havenet CO2 fixation. In this issue of New Phytologist, Cramer et al.(pp. 1048–1057) have built on knowledge that transpiratorymass flow of water through soil is important in moving somenutrients to the root surface and present results consistentwith the stimulation of nutrient supply to plant roots by massflow through the soil as a component of the regulation of therate of transpiration.
‘... can it be concluded that the increased water loss is
a result of natural selection in evolution increasing
mass flow of nutrients to the root surface?’
The simplistic view of how embryophytes living on land dealwith the unavoidable loss of water vapour in photosynthesisand the frequently irregular supply of water in precipitation
divides them into two categories: poikilohydric (i.e. with little orno capacity to limit transpiratory water loss) and homoiohydric(i.e. with significant capacity to limit transpiratory water loss).Unless the poikilohydric organisms live in permanently wethabitats they are subject to periodic drying out, and persistencein drier habitats requires desiccation tolerance. Homoiohydricorganisms are generally not desiccation tolerant except inreproductive and dispersal stages, such as dispersed spores(including pollen grains) and seeds and fruits. Homoiohydricplants comprise most terrestrial vegetation today: ContinenceConquers Continents. This neat pigeonholing of plants onland into the categories poikilohydric or homoiohydric, or intothe partly similar categories desiccation-tolerant or desiccation-intolerant, has been criticized by Proctor & Tuba (2002), whorightly point out that strict poikilohydry and strict homoiohydryare the extremes of a continuum, as are complete tolerance ortotal intolerance of desiccation.
Nevertheless, it is convenient to consider sporophytes ofterrestrial vascular plants as predominantly homoiohydricand desiccation intolerant, obtaining photosynthetically activeradiation (PAR) and essentially all of their carbon as CO2from the atmosphere, and water and the great majority ofnutrient elements (other than carbon) from the soil. Theenergy balance of terrestrial plant canopies is described by thePenman–Monteith equation, with energy storage in net primaryproductivity accounting for a minor component of the absorbedshort-wavelength radiation (mainly PAR), and the rest con-tributing the latent heat of evaporation in transpiration andthe loss of sensible heat (Jones, 1992).
Stomata are major determinants of the fate of absorbedshort-wave radiation as energy storage in net photosynthesisand energy dissipation by transpiration (although these twofates are not in direct proportion as a function of stomatalaperture), on the one hand, and increased canopy temperatureand energy dissipation other than through transpiration, onthe other hand. The stomata respond to the atmospheric andinternal leaf environment, the latter crucially including chemicalsignals that have been generated by below-ground parts of theplant in response to the soil environment and permittingthe pre-emptive function of stomata in restricting gas exchangebefore the plant becomes significantly desiccated (Hetherington& Woodward, 2003). The Cowan–Farquhar (Cowan, 1977;Cowan & Farquhar, 1977) approach to modelling stomatalbehaviour, in terms of optimizing carbon gain per unit waterlost in transpiration, has been very successful.
However, Cramer et al. point out that the optimizationmodelling is based on stomatal behaviour during periods onnet CO2 assimilation, so does not take into account nocturnaltranspiration in C3 and C4 plants (Caird et al., 2007). Thesame applies to the use of δ13C as a surrogate for stomatalconductance relative to the biochemical capacity for photo-synthesis. There is also (Cramer et al.) evidence that transpiratorymass flow of water through soils is important in moving somenutrients to the root surface (Barber & Cushman, 1981).
Cramer et al. (2008) point out that the water transpired perunit dry matter accumulated by plants increases when growthis limited by nutrient supply (Raven et al., 2004), which isconsistent with a role for increased water flow through soilcompensating in part for decreased availability of nutrients inthe soil. The fact that such transpiratory responses also occurin hydroponic cultures where, under well-stirred conditions,there would be no increase in nutrient flux to the root surface,does not rule out a role of increasing nutrient fluxes throughthe soil (Cramer et al.) if the signal for increased water flux isrelated to nutrient availability at the root surface. In thisconnection, Cramer et al. point out that nitrate delivered tothe leaf in the xylem is among the determinants of stomatalbehaviour (Wilkinson et al., 2007). Finally, Cramer et al. com-ment on the very wide variation in the measured transpiratorywater loss per unit dry matter accumulation among C3 plants(Wright et al., 2004).
These possibilities led Cramer et al. to carry out experimentsdesigned to examine the effects of varying the dependence ofnutrient delivery to the root surface by mass flow through thesoil on the ratio of transpiration to photosynthetic carbonassimilation. They used the South African winter rainfall grassEnhartia calycina J.E. Sim from nine sites along a rainfallgradient growing in sand. One treatment was ‘interception’because the roots and the fertilizer pellets were in the samevolume of sand, and nutrients could reach the root surface byinterception (root growth) as well as by diffusion and massflow. The other treatment was ‘mass flow’, in which the fertilizerpellets were supplied in sand in a compartment at the centreof the growth vessel that was separated from the sand containingthe roots by a mesh-screened polyvinyl chloride (PVC) pipethat roots could not penetrate but which allowed diffusionand, especially, mass flow of nutrients to the roots.
The ‘mass flow’ plants had statistically significantly lower(mean of 35%) relative growth rates than the ‘interception’plants. The ‘mass flow’ plants also had statistically significantlyhigher stomatal conductance, transpiration rate and intercellularspace CO2 concentration than the ‘interception’ plants, andalso a statistically significantly higher net photosyntheticrate, although the difference between the ‘mass flow’ and the‘interception’ plants was not as great as for the other threeparameters. The higher stomatal conductance and intercellularCO2 concentration resulted, as expected, in a more negativeδ13C value of leaves of ‘mass flow’ than of ‘interception’ plants.There was no difference between treatments in the fraction ofthe plants contributed by roots.
Cramer et al. suggested that the higher leaf area-based rateof net photosynthesis, despite the lower relative growth rate,of the ‘mass flow’ plants than of the ‘interception’ plants couldbe attributed to a greater respiratory carbon loss by the rootsof the ‘mass flow’ plants as a result of the (assumed) lower con-centration of nutrients at the root surface, and hence a greaterenergy cost per unit nutrient taken up by the roots. However,in the absence of measurements of the leaf area per plant dry
matter (leaf area ratio or LAR) it is not possible to be certainthat the higher photosynthetic rate of ‘mass flow’ plants on aleaf area basis means a greater rate of carbon fixation in netphotosynthesis per unit plant dry matter than would beneeded to support the measured lower relative growth, assumingthe same fractional respiratory loss for the two treatments.However, this possibility would require a so much lower LARin the ‘mass flow’ than in the ‘interception’ plants that it wouldhave been noticed when examining the plants. The same pointapplies to the measured transpiration rate on leaf area basis inrelation to the rate of water loss from the whole plant; there isno estimate of water loss per unit dry matter, or per unit drymatter gain, by the plants in the two treatments.
The content of nitrogen (N) and potassium (K) on a leaf drymatter basis was not significantly different between ‘interception’and ‘mass flow’ plants. The leaf dry matter of ‘mass flow’ plantshad significantly more sodium (Na), calcium (Ca), zinc (Zn),manganese (Mn), phosphorus (P) and copper (Cu), andsignificantly less iron (Fe) and magnesium (Mg), than did theleaf dry matter of ‘interception’ plants. The finding that theleaf content on a dry matter basis of eight of the ten elementsexamined was either the same (two) or higher (six) in the ‘massflow’ plants than for the ‘interception’ plants is taken by Crameret al. to indicate that increased mass flow of most nutrientelements compensates, or more than compensates, for theabsence of the interception component of nutrient acquisi-tion in the ‘mass flow’ plants. This conclusion is reasonable,although there are implicit assumptions about the relevance ofthe transpiration rate measured in the LICOR gas-exchangeapparatus on a leaf area basis to the water loss by the growingplant, and that leaf dry matter (for the elemental analysis) hasthe same relation to leaf area (for the gas exchange) in the‘interception’ plants as in the ‘mass flow’ plants.
Accepting that there is indeed increased water loss per unitdry matter gain in the nutrient-limited ‘mass flow’ plants, canit be concluded that the increased water loss is a result ofnatural selection in evolution increasing the mass flow ofnutrients to the root surface? While this is a possibility, it isalso possible that it is an emergent property of the operationof mechanisms that have evolved in relation to other functions.This is, of course, a comment which could be made aboutmany traits that are suggested to have significance in naturalselection, so it is helpful to bring in data from plants whosegrowth is limited by a factor other than nutrient uptake fromsoil. Such a factor is PAR; using δ13C as a surrogate for waterloss per unit dry matter gain (see Raven & Farquhar, 1990),there is a greater potential for water loss per unit dry mattergain during slower growth at low PAR than during fastergrowth at higher PAR in the C3 aroid Dieffenbachia longispathaEngler and Krause (Skillman et al., 2005), although thephenotypic effect of PAR for growth on water-use efficiencyin nature is complex (Bacon, 2004). Here it is difficult to seea role for increased mass flow through soil per unit plantgrowth when there would be expected to be relatively no less
nutrient acquisition by interception or diffusion for the small,low-PAR plants than for the larger, high-PAR plants. Thereare also data showing a lower water-use efficiency and growthrate of C3 plants grown at the lower CO2 levels found at thelast glacial maximum than of similar plants grown in present-day CO2 (Polley et al., 1993); work on a red alga (which, ofcourse, lacks stomata) that relies on diffusive CO2 entry forphotosynthesis (Kübler et al., 1999) confirms that the increaseddiffusive conductance relative to photosynthetic capacity invascular plants grown at low CO2 is a stomatal response. Thesetwo examples show that decreased growth rate as a result oflower PAR or lower CO2, like nutrient limitation, increasetranspiration per unit dry matter gain.
The work reported by Cramer et al. (2008) is certainlyinteresting, and should be followed up by, for example, con-sidering soil rather than sand where elements such as P and Feare expected to be less mobile, and also by investigatingmycorrhizal plants.
John A. Raven
Division of Plant Sciences, University of Dundee at SCRI,Scottish Crop Research Institute, Invergowrie, Dundee DD2
5DA, UK (Author for correspondence: tel.+44 1382 562731; fax +44 113 568275; email
Bacon MS, ed. 2004. Water use efficiency in plant biology. Oxford, UK: Blackwell.Barber SA, Cushman JH. 1981. Nitrogen uptake model for agronomic
crops. In: Iskander JK, ed. Modelling waste water renovation-land treatment. New York, NY, USA: Wiley-Interscience, 382–409.
Caird MA, Richards JH, Donovan LA. 2007. Nighttime stomatal conductance and transpiration in C3 and C4 plants. Plant Physiology 143: 4–10.
Cowan IR. 1977. Stomatal behaviour and environment. Advances in Botanical Research 4: 1176–1227.
Cowan IR, Farquhar GD. 1977. Stomatal function in relation to leaf metabolism and environment. Symposia of the Society for Experimental Biology 31: 471–505.
Cramer MD, Hoffmann V, Verboom GA. 2008. Nutrient availability moderates transpiration in Ehrhartia calycina. New Phytologist 179: 1048–1057.
Hetherington AM, Woodward FI. 2003. The role of stomata in sensing and driving environmental change. Nature 424: 901–908.
Jones HG. 1992. Plants and microclimate. A quantitative approach to environmental plant physiology, 2nd edn. Cambridge, UK: Cambridge University Press.
Kübler JE, Johnston AM, Raven JA. 1999. The effect of reduced and elevated CO2 and O2 on the seaweed Lomentaria articulate. Plant, Cell & Envirironment 22: 1303–1310.
Polley HW, Johnston HB, Marino BP, Mayeux HS. 1993. Increase in water-use efficiency and biomass over glacial to present CO2 concentrations. Nature 361: 61–64.
Proctor MCF, Tuba Z. 2002. Poikilohydry and homoihydry: antithesis or spectrum of possibilities. New Phytologist 156: 327–349.
Raven JA, Farquhar GD. 1990. The influence of N metabolism and organic acid synthesis in the natural abundance of C isotopes in plants. New Phytologist 116: 505–529.
Raven JA, Handley LL, Wollenweber B. 2004. Plant nutrition and water use efficiency. In: Bacon MS, ed. Water use efficiency in plant biology. Oxford, UK: Blackwell, 162–188.
Skillman JB, Garcia M, Virgo A, Winter K. 2005. Growth irradiance effects on photosynthesis and growth in two co-occurring shade-tolerant neotropical perennials of contrasting photosynthetic pathways. American Journal of Botany 98: 1811–1819.
Wilkinson S, Bacon MAZ, Davies WJ. 2007. Nitrate signaling to stomata and growing leaves: interactions with soil drying, ABA, and xylem sap pH in maize. Journal of Experimental Botany 58: 1705–1716.
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z, Bongers F, Cavender-Bares J, Chapin FS, Cornelissen JHC, Diemer M et al. 2004. The world-wide leaf economics spectrum. Nature 428: 821–827.
Secretomic climax in plant–fungal interactions9th European Conference on Fungal Genetics, Edinburgh, April, 2008
The 9th European Conference on Fungal Genetics (ECFG9)was held in the beautiful city of Edinburgh (UK). Over
five-hundred participants assembled in the venerable hallsof the University of Edinburgh to learn about currentprogress in the field of fungal genetics research. An excellentscientific programme accompanied by a catalogue of socialevents made for a memorable meeting. Malt whisky tasting,Scottish dancing and the organizing committee dressed inkilts at the conference dinner are just some examples ofwhat was experienced by the delegates during their stay inEdinburgh.
‘... small secreted proteins could act as putative effectors
manipulating plant detection systems to establish
compatible interactions’
Recent insights in plant–fungal interactions
The ECFG covers a wide range of fungal genetics and molecularresearch. However, one striking observation from this year’smeeting was the considerable number of plant–fungal interactionpresentations and posters given, underlining just how fast thisparticular field is advancing. During the morning plenary sessions,Nick Talbot (University of Exeter, UK) brought us more detailsabout the biology of the model pathogen, Magnaporthe grisea,responsible for rice blast disease. He described throughcytological approaches the mitotic control that takes placeduring the development of the appressorium, the specializedinfection structure necessary for successful plant infection. Theextensive data now available for this plant pathogen makes it anexcellent model, which contributes greatly to our understandingof the plant infection process. Francis Martin (INRA Nancy,France) presented some of the typical features found in the codingspace of the genome of the symbiotic fungus Laccaria bicolor.He reported the presence of hundreds of L. bicolor-specificgenes coding for small secreted proteins, some being highlyregulated in the symbiotic tissues compared with free-livingmycelium. Using immunolocalization it has been demonstratedthat one promising mycorrhiza-induced small secreted protein(MISSP7) accumulated in the Hartig net, the plant–fungalmixed tissue where nutrient exchange takes place between thetwo partners of the mutualistic association (Martin et al., 2008).
The inside men – getting deeper into fungal cells
Living cell biology has reached an amazing level of descriptionfor biological systems. Aside from the obvious scientific reasons,one not in touch with the latest developments in this fast-moving field of science may have felt like they were viewing a‘Hollywood blockbuster’ with special effects when watchingsome of the presentations in this session. At every ECFG, cellbiologists are able to take us deeper into fungal cells, allowingus to experience ‘live’ highly complex biological mechanisms.In this way, several talks from the plenary morning sessions madequite an impression on the ECFG9 audience. For example,the ‘ping-pong’ response of Green Fluorescent Protein (GFP)-labeled Mitogen Activated Protein (MAK2) kinase at the tipsof anastomosis tubes during Neurospora crassa conidial germlingfusion presented by Nick Read (University of Edinburgh, UK) wasone excellent example of how close we can now get to a biological
process. Gero Steinberg (University of Exeter, UK) introducedthe laser-excitation microscopy device – that he stated as beingthe most sensitive – which provides a promising technique fordeciphering the cytoskeleton dynamics in hyphal growth usingthe plant pathogen model Ustilago maydis. For the first time thenumber of dynein units delivered to the tips of developing fungalcells, to remain there or go backwards on microtubules, wasquantified. Such counting was made possible by approximatingthe level of fluorescence of a single GFP unit signal in an ingeniouscounting system set on the nucleopore protein complex. Thesedevelopments, together with new imaging technologies andthe improved quality of fluorescent labeling, will stronglyinfluence the field of fungal biology in the future.
Secretomics: towards the holy grail of plant–fungal interactions
In recent years, the search for secreted polypeptide effectormolecules has emerged as one of the ‘hot’ topics in plant–microbe interactions. Just glancing at the meeting abstracts itcould be seen that this trend has reached a new climax. Afterthe publication of several fungal genomes belonging to speciesthat interact with plants, comparative genomics is now offeringmore clues and leads about how a microbe is able to breach itshost defense and establish a biotrophic interaction or gainaccess to its nutrient source. One of the key areas involved incontrolling plant–fungal interactions is the extracellular interfacebetween the two organisms. The complex interplay among signals,cell-wall-degrading and/or remodelling enzymes, and smallsecreted proteins that are released into this extracellular spaceand eventually delivered into the host cell, determines the ability,or inability, of the host to trigger its defense system against theinvading organism (de Witt, 2007). The comprehension ofgeneral mechanisms controlling such events is yet to be reachedowing to the high diversity of secreted proteins described inthe proteomes of many fungal species interacting with plants.However, recent publications have identified impressive catalogsof small secreted proteins expressed during plant–fungusinteractions (Catanzariti et al., 2006; Kämper et al., 2006;Martin et al., 2008). More candidates presented at the ECFG9in symbiont or pathogen fungal species extended this list (seebelow). The number of specific genes described for a givenspecies is quite striking and may underline the specificity ofthe interaction process between a pathogen and a symbiont,or between distinct pathogens and their plant-hosts after along-lasting interaction history. To date, very few small fungal-secreted proteins have been explored in great detail; however,more studies are expected after in-depth examination offungal genome contents (Mueller et al., 2008a,b). The NewPhytologist Trust will support a symposium entitled ‘Effectorsin plant–microbe interactions’ in September 2009 with theaim of developing a better understanding of effector proteins.
Many of the presentations given in the plant–fungal inter-action parallel session stated the importance of modulating
the plant defense response or recognition of the symbioticpartner in pathogenic as well as in mutualistic interactions.The experimental set ups included examination of wholesecretomes (U. maydis, L. bicolor), immunolocalization, theyeast-based signal sequence trap (YSST), sequencing of tissue-specific cDNA libraries and infection studies with bacterialtype III delivery vectors. The Yeast 2 Hybrid system, whichsuffered bad press as a result of its tedious screening processand high false-positive rates, appears to have experienced arenaissance, with many presentations and posters utilizing thisapproach to track proteins targeted by fungal effectors in planta.
Regine Kahmann (Max Planck Institute, Marburg, Germany)presented the latest advances concerning gene clusters fromU. maydis that encode secreted proteins. Candidate genesexpressed during plant infection influence pathogenicity ofthe fungus, and the selective deletion of specific clusters or genesimpacts upon the distinct defense responses from the plant(Kämper et al., 2006). Kahmann described, in great detail,how these secreted effectors may shape the interaction withthe host after secretion. The research she presented addressedthe localization of secreted effector molecules, their interact-ing partners and how plant defense responses are impacted.Several genes encode proteins presumably translocated intothe nucleus of the host plant cell, and a deletion strain of aninteresting candidate fails to penetrate and elicits plant celldeath, thus suggesting a possible involvement in the suppres-sion of the host defense response. Despite these new insightsinto the biology of this model pathogen, Kahmann expressedcaution because most of the data were obtained using heter-ologous systems rather than from experiments performed inthe host plant, which is necessary to demonstrate unequivo-cally the presumed roles of these U. maydis-secreted proteins.
Several other presentations focused on the latest advancesin our understanding of different plant–fungal interactions(with both pathogens and symbionts); these also describedrepertoires of secreted proteins. Richard O’Connell (MaxPlanck Institute, Köln, Germany) and Barbara Valent (KansasState University, Manhattan, USA) reported sets of smallsecreted proteins in Colleotrichium higginsianum and M. grisea,respectively, which could contain putative effector genes.Similarly, Natalia Requena (University of Karlsruhe, Germany)presented prospective work that aims to identify secreted proteinsfrom the endomycorrhizal fungus, Glomus intraradices, whichcould have roles in the control of the plant defense responseand orientation to symbiosis development. She introducedthe use of a yeast signal sequence trap method (Lee et al.,2006) to identify effectors in presymbiotic and symbiotictissues of Medicago truncatula.
Ecogenomic perspectives with more basidiomycete genomes to come
Understanding the growth, health and physiology of plants innature cannot be achieved without taking into account the
diversity of microbial species that interact continuously intheir changing ecosystems. An excellent example of this is thePopulus community genome project that aims to sequence notonly the genome of the model tree species Populus trichocarpabut also those of several microbes interacting with this tree(Martin et al., 2004, 2008; Tuskan et al., 2006). Followingthe close of the ECFG9, Basidio3, the third basidiomycetesatellite meeting, was held. A statement was made by IgorGrigoriev ( Joint Genome Institute ( JGI), USA) about thebasidiomycete genomes that would be sequenced in the comingyears at the JGI. After this opening contribution, severalpresentations discussed future studies where these genomeswould aid or underpin the research. The fungal genome projectspresented included saprotrophs, such as the brown-rot Postiaplacenta, the dry-rot Serpula lacrymans and Pleurotus ostreatus;the gymnosperm root pathogen Heterobasidion annosum andthe poplar leaf rust Melampsora larici-populina; and theectomycorrhizal symbiont Paxillus involutus. All these speciesplay major roles in diverse niches of terrestrial ecosystems andgreatly impact upon diversity through direct interactions –beneficial or detrimental – or through mobilization andcycling of organic matter. Such progress in the genomics ofbasidiomycetes will provide an unparalleled opportunity toidentify key points in interspecific and fungal–environmentinteractions and increase our understanding of how these specieseffect the functioning of the ecosystem (Martin & Slater, 2007).
The shape of things to come
Recent research in the field of plant–fungal interactions hasdrawn attention to the possibility that small secreted proteinscould act as putative effectors manipulating plant detectionsystems to establish compatible interactions. Over the last fewyears fungal genomics has uncovered hundreds of candidateeffectors of potential interest to the scientific community.These not only help in deciphering the mechanisms thatunderlie plant–fungal interactions but also in addressingcrucial issues such as compatibility and recognition. As shownat the ECFG9, postgenomic studies are now underway inmodel plant–fungus interactions, and there is more to comefrom other models that are benefiting from recent genomicefforts. These studies, together with comparative genomics(possibly with more fungal genomes to come), provide a verypromising outlook for the ECFG10.
S. Duplessis1* and H. Kuhn2
1UMR 1136 INRA/UHP-Nancy 1, Interactions Arbres/Micro-organismes, Centre INRA de Nancy, F-54280,
Champenoux, France; 2KIT, University of Karlsruhe (TH),Institute for Applied Biosciences, Heisenberg-Group
‘Plant–Fungal-Interactions’, Hertzstrasse 16, D-76187,Karlsruhe, Germany (*Author for correspondence:
Catanzariti A-M, Dodds PN, Lawrence GJ, Ayliffe MA, Ellis JG. 2006. Haustorially expressed secreted proteins from flax rust are highly enriched for avirulence elicitors. Plant Cell 18: 243–256.
Kämper J, Kahmann R, Bölker M, Ma L-J, Brefort T, Saville BJ, Banuett F, Kronstad JW, Gold SE, Müller O et al. 2006. Insights from the genome of the biotrophic fungal pathogen Ustilago maydis. Nature 444: 97–101.
Lee SJ, Kelley BS, Damasceno CM, St John B, Kim BS, Kim BD, Rose JK. 2006. A functional screen to characterize the secretomes of eukaryotic pathogens and their hosts in planta. Molecular Plant–Microbe Interactions 19: 1368–77.
Martin F, Aerts A, Ahrén D, Brun A, Danchin EGJ, Duchaussoy F, Gibon J, Kohler A, Lindquist E, Pereda V et al. 2008. The genome of Laccaria bicolor provides insights into mycorrhizal symbiosis. Nature 452: 88–92.
Martin F, Slater H. 2007. New Phytologist – an evolving host for mycorrhizal research. New Phytologist 174: 225–228.
Martin F, Tuskan GA, DiFazio SP, Lammers P, Newcombe G, Podila GK. 2004. Symbiotic sequencing for the Populus mesocosm: DOE tackles the genomes of endomycorrhizal Glomus intraradices and ectomycorrhizal Laccaria bicolor. New Phytologist 161: 330–335.
Mueller O, Kahmann R, Aguilar G, Trejo-Aguilar B, Wu A, de Vries RP. 2008a. The secretome of the maize pathogen Ustilago maydis. Fungal Genetics and Biology, in press, doi:10.1016/j.fgb.2008.03.012.
Müller O, Schreier PH, Uhrig JF. 2008b. Identification and characterization of secreted and pathogenesis-related proteins in Ustilago maydis. Molecular Genetics and Genomics 279: 27–39.
Tuskan G, DiFazio S, Bohlmann J, Grigoriev I, Hellsten U, Jansson S, Putnam N, Ralph S, Rombauts S, Salamov A et al. 2006. The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). Science 313: 1596–1604.
de Witt PJGM. 2007. How plants recognize pathogens and defend themselves. Cellular and Molecular Life Sciences 64: 2726–2732.
• New Phytologist is owned by a non-profit-making charitable trust dedicated to the promotion of plant science, facilitating projectsfrom symposia to open access for our Tansley reviews. Complete information is available at www.newphytologist.org.
• Regular papers, Letters, Research reviews, Rapid reports and both Modelling/Theory and Methods papers are encouraged.We are committed to rapid processing, from online submission through to publication ‘as-ready’ via Early View – our averagesubmission to decision time is just 29 days. Online-only colour is free, and essential print colour costs will be met if necessary.We also provide 25 offprints as well as a PDF for each article.
• For online summaries and ToC alerts, go to the website and click on ‘Journal online’. You can take out a personal subscription tothe journal for a fraction of the institutional price. Rates start at £135 in Europe/$251 in the USA & Canada for the online edition(click on ‘Subscribe’ at the website).
• If you have any questions, do get in touch with Central Office ([email protected]; tel +44 1524 594691) or, for a localcontact in North America, the US Office ([email protected]; tel +1 865 576 5261).