(This is a sample cover image for this issue. The actual cover is not yet available at this time.) This article appeared in a journal published by Elsevier. The attached copy is furnished to the author for internal non-commercial research and education use, including for instruction at the authors institution and sharing with colleagues. Other uses, including reproduction and distribution, or selling or licensing copies, or posting to personal, institutional or third party websites are prohibited. In most cases authors are permitted to post their version of the article (e.g. in Word or Tex form) to their personal website or institutional repository. Authors requiring further information regarding Elsevier’s archiving and manuscript policies are encouraged to visit: http://www.elsevier.com/copyright
Transcript
(This is a sample cover image for this issue. The actual cover is not yet available at this time.)
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
Polyunsaturated fatty acids cause apoptosis in C. albicans and C. dubliniensis biofilms
Vuyisile S. Thibane a, Ruan Ells a, Arno Hugo a, Jacobus Albertyn a, Walter J. Janse van Rensburg b,Pieter W.J. Van Wyk c, Johan L.F. Kock a, Carolina H. Pohl a,⁎a Department of Microbial, Biochemical and Food Biotechnology, University of the Free State, P. O. Box 339, Bloemfontein, 9300, South Africab Department of Haematology and Cell Biology, University of the Free State, P. O. Box 339, Bloemfontein, 9300, South Africac Centre for Microscopy, University of the Free State, P. O. Box 339, Bloemfontein, 9300, South Africa
a b s t r a c ta r t i c l e i n f o
Article history:Received 16 February 2012Received in revised form 24 April 2012Accepted 10 May 2012Available online 17 May 2012
Background: Polyunsaturated fatty acids (PUFAs) have antifungal properties, but the mode by which they in-duce their action is not always clear. The aim of the study was to investigate apoptosis as a mode of action ofantifungal PUFAs (stearidonic acid, eicosapentaenoic acid and docosapentaenoic acid) which are inhibitorytowards biofilm formation of C. albicans and C. dubliniensis.Methods: Candida biofilms were grown in the absence or presence of 1 mM PUFAs (linoleic acid, stearidonicacid, eicosapentaenoic acid, docosapentaenoic acid) for 48 h at 37 °C. The effect of these PUFAs on the mem-brane fatty acid profile and unsaturation index, oxidative stress, mitochondrial transmembrane potential andapoptosis was evaluated.Results: When biofilms of C. albicans and C. dubliniensis were exposed to certain PUFAs there was an increasein unsaturation index of the cellular membranes and accumulation of intracellular reactive oxygen species(ROS). This resulted in apoptosis, evidenced by reduced mitochondrial membrane potential and nuclear con-densation and fragmentation. The most effective PUFA was stearidonic acid.Conclusions: The resultant cell death of both C. albicans and C. dubliniensis is due to apoptosis.General significance: Due to the increase in drug resistance, alternative antifungal drugs are needed. A groupof natural antifungal compounds is PUFAs. However, understanding their mechanisms of action is importantfor further use and development of these compounds as antifungal drugs. This paper provides insight into apossible mode of action of antifungal PUFAs.
In an earlier study [1] itwas found that certainmarine polyunsaturat-ed fatty acids (PUFAs) [i.e. stearidonic acid (C18:4 n-3), eicosapentaenoicacid (C20:5 n-3) and docosapentaenoic acid (C22:5 n-3)] cause inhibi-tion of Candida albicans and Candida dubliniensis biofilm biomass pro-duction as well as metabolic activity. The authors speculated that theseeffects may be due to oxidative stress, which could lead to apoptosis.
Apoptosis is characterized by a set of distinct morphological markersand yeast cells show typical apoptotic markers such as the externali-zation of phosphatidylserine, DNA fragmentation as well as chromatinfragmentation and condensation. Furthermore, the cytochrome c releasefrom the mitochondria [2], reduced mitochondrial membrane potential[3] and formation ofmembrane enclosed cell fragments, called apoptoticbodies, all mark programmed death of yeast cells [4]. Madeo and co-workers [5] first discovered the apoptotic pathway similar to those in
multicellular organisms, in the yeast Saccharomyces cerevisiae. Sincethen several endogenous and exogenous stimuli have been shown tobe responsible for the induction of apoptosis in this yeast. Some ofthese stimuli include mutations or deletions of specific genes [5] andexogenous stimuli such as acetic acid and hydrogen peroxide [6–8].Research has also shown the existence of apoptotic pathways in thehuman pathogen Candida albicans. In this yeast, antimicrobial peptidessuch as melittin [9], psacotheasin [10], pleurocidin [11], papiliocin [12]and arenicin-1 [13] were found to be antifungal through the inductionof apoptosis. These peptides were reported to induce oxidative stressthrough increased production of reactive oxygen species (ROS) andlipid peroxidation of mitochondrial membranes. Accumulation of ROShas long been proposed to be a key event in the apoptotic pathway [14].
The toxicity of fatty acids is dose dependent and relates to chainlength and unsaturation, especially with PUFAs, causing mammaliancell death through apoptosis [15]. In the recent work by Ferreiraand co-workers [16], it was reported that supplementation of mediawith PUFAs could induce the apoptotic pathway in S. cerevisiae.
The aim of the study was therefore to investigate whether the ob-served antifungal activity of marine PUFAs, as reported by Thibaneand co-workers [1], is through the induction of apoptosis in C. albicansand C. dubliniensis biofilms.
Biochimica et Biophysica Acta 1820 (2012) 1463–1468
Candida albicans CBS 562 T and Candida dubliniensisNRRL Y-17841 Twere used in this study and were maintained on yeast malt extract(YM) agar plates (10 g/L glucose, 3 g/L yeast extract, 3 g/L malt extract,5 g/L peptone, 16 g/L agar) at room temperature. The strains were alsostored on agar slants at 4 °C.
2.2. Biofilm formation
Cells of C. albicans and C. dubliniensis were grown separately onYM agar plates and incubated at 30 °C for 24 h. After incubation, aloop-full of the cells was inoculated into 20 mL of yeast nitrogenbase (YNB) glucose medium (10 g/L glucose, 6.7 g/L YNB) and incu-bated at 30 °C for 48 h. Cells were washed twice with sterile phos-phate buffered saline (PBS) by centrifugation for 5 min at 4000 ×gwith a Heraeus®Megafuge® 1.0R Centrifuge and diluted in filter ster-ilized RPMI-1640 medium (Sigma Aldrich, USA) to a standardizedcell concentration of 1×106 cells/mL. Biofilms were allowed to formby first incubating the cells at 37 °C for 1 h, to allow adherence ofcells to the surface [17]. Non-adherent cells were removed by washingtwice with sterile PBS. Different PUFAs [linoleic acid (C18:2 n-6),C18:4 n-3, C20:5 n-3 and C22:5 n-3] with a final concentration of1 mM, as used in Thibane and co-workers [1], were added after 1 hourincubation and mature biofilms were formed at 37 °C for an additional47 h. Ethanol was used as a control.
2.3. Phospholipid fatty acid analyses
Biofilms, formed in Petri dishes and supplemented with PUFAs, werewashed twice with sterile PBS, scraped off using a cell scraper andresuspended in sterile PBS. Total lipids of the biofilms were extractedwith chloroform:methanol (2:1) [18]. The total lipids were separatedinto different fractions using solid phase 0.50 g Si extraction columns(Separations, SA) according to themethod of Bossio and Scow [19]. Brief-ly, columnswere conditionedwith 2 mL chloroform followed by transferof the lipid samples, resuspended in 300 μL chloroform, to the columns.The neutral lipids were eluted with 5 mL chloroform, followed by theglycolipids with 10 mL acetone and finally the phospholipids with5 mL methanol. The eluates were collected and dried under N2 at 32 °C.Fatty acidmethyl esters (FAMEs) of the phospholipid fractionswere pre-pared using methanol-BF3 [20] and quantified using a Varian 430-GCgas chromatograph, with a fused silica capillary column (ChrompackCPSIL 88, 100 m length, 0.25 mm ID, 0.2 μm film thickness). The columntemperature was 40–230 °C (hold 2 min; 4 °C/min; hold 10 min). Fattyacid methyl esters in hexane (1 μL) were injected into the columnusing aVarian CP-8400Autosamplerwith a split ratio of 100:1. The injec-tion port and detector temperatures were maintained at 250 °C. Hydro-gen, at 45 psi, functioned as the carrier gas, while nitrogen wasemployed as themakeup gas. Galaxy Chromatography Data System soft-ware recorded the chromatograms. Fatty acidmethyl ester sampleswereidentified by comparing the relative retention times of FAMEs peaksfrom samples with those of standards. Fatty acids were expressed asthe relative percentage of each individual fatty acid to the total of allfatty acids present in the samples. The unsaturation indexwas calculatedusing the following equation: Unsaturation Index=1×[% monoenoicfatty acids]+2×[% dienoic fatty acids]+3×[% trienoic fatty acids]+4×[% tetraenoic fatty acids]+5×[% pentaenoic fatty acids]. This experi-ment was performed in duplicate on different occasions.
2.4. Measurement of reactive oxygen species (ROS)
Accumulation of intracellular ROS production was measured usingthe fluorescent dye, 2’,7-dichlorofluorescin diacetate (DCFHDA) (Sigma
Aldrich, USA). Briefly, biofilms were prepared as described above inblack 96-well microtiter plates (Corning Incorporated, Costar®, USA)by incubating plates at 37 °C for 24 h. Biofilms were washed twicewith sterile PBS to remove non-adherent cells and 100 μL of filter steril-ized RPMI-1640 medium containing 1 mM of the PUFAs were added.An ethanol controlwas included. A volume of 10 μMDCFHDAwas simul-taneously added with the PUFAs and the plate was incubated at 37 °Cfor an additional 12 h [21]. Fluorescence was measured after incu-bation using a SpectraMax M2 plate reader (Molecular Devices, USA)at an excitation/emission wavelength of 485/535 nm, respectively.
2.5. Measurement of mitochondrial membrane potential
The mitochondrial membrane potential of biofilms prepared inblack 96-well microtiter plates, and supplemented with PUFAs asdescribed above, was measured. After incubation, wells were washedtwice with sterile PBS and the plates assayed according to the JC-1Mitochondrial Membrane Potential Assay Kit (Cayman chemicals,USA) using a SpectraMax M2 plate reader. The lipophilic cationicdye, 5,5′,6,6′-terachloro-1,1′,3,3′-tetraethylbenzimidazolylcarbocya-nine iodide (JC-1), selectively enters the mitochondria. In healthycells, with high mitochondrial transmembrane potential (ΔΨm),JC-1 forms J-aggregates with an intense red fluorescence at excita-tion/emission wavelength of 540/570 nm, respectively. In apoptoticor unhealthy cells, with low ΔΨm, JC-1 remains in the monomericform which fluoresces green at excitation/emission wavelength of485/535 nm, respectively [22]. Mitochondrial membrane potentialwas presented as the ratio of the J-aggregates (healthy cells) andmonomeric forms (apoptotic cells).
2.6. DNA fragmentation (TUNEL) assay
For the analyses of DNA strand breaks, biofilms were prepared inPetri dishes and supplemented with PUFAs as described above. Cellswere washed twice with PBS and fixed in 3.6% formaldehyde. Fixedcells were washed twice with PBS and protoplasted as describedand permeabilized in permeabilization solution (0.1% Triton X-100and 0.1% sodium citrate) for 2 min on ice. DNA strands were labeledwith an In Situ Cell Death Detection Fluorescein Kit (Roche AppliedScience, Germany) at 37 °C for 1 h [23]. Cells were analyzed on theFACSCalibur Flowcytometer using CellQuest Pro software. At least75 000 cells were counted per analyses. Instrument-settings for stan-dard human platelets were used. Hydrogen peroxide (10 mM) wasused as positive control.
Table 1Phospholipid fatty acid profiles of biofilms of Candida albicans following supplementationwith C18:2 n-6, C18:4 n-3, C20:5 n-3 and C22:5 n-3. Values represent mean of duplicateexperiments with the range in brackets.
2.7. Nuclear condensation and fragmentation (DAPI staining)
Biofilms were prepared in chamber slides (Lab-Tek® ChamberSlide™ System, USA) and supplemented with PUFAs as describedabove. Nuclear condensation and fragmentation were analyzed by4′, 6-diamidino-2-phenylindole (DAPI) staining [5]. Biofilms werewashed twice with PBS, permeabilized in permeabilization solution(0.1% Triton X-100 and 0.1% sodium citrate) for 2 min on ice, andincubated with 1 μg/mL of DAPI in the dark at 37 °C for 20 min [12].Cells were qualitatively examined by confocal laser microscopy.
2.8. Statistical analyses
Unless stated otherwise all experiments were conducted in tripli-cate on separate occasions. Averages and standard deviations werecalculated and the student's t-test performed to determine the signif-icance of the data sets. A P value of≤0.01 was considered significant.
3. Results and discussion
3.1. PUFAs influence phospholipid fatty acid profiles and unsaturationindices
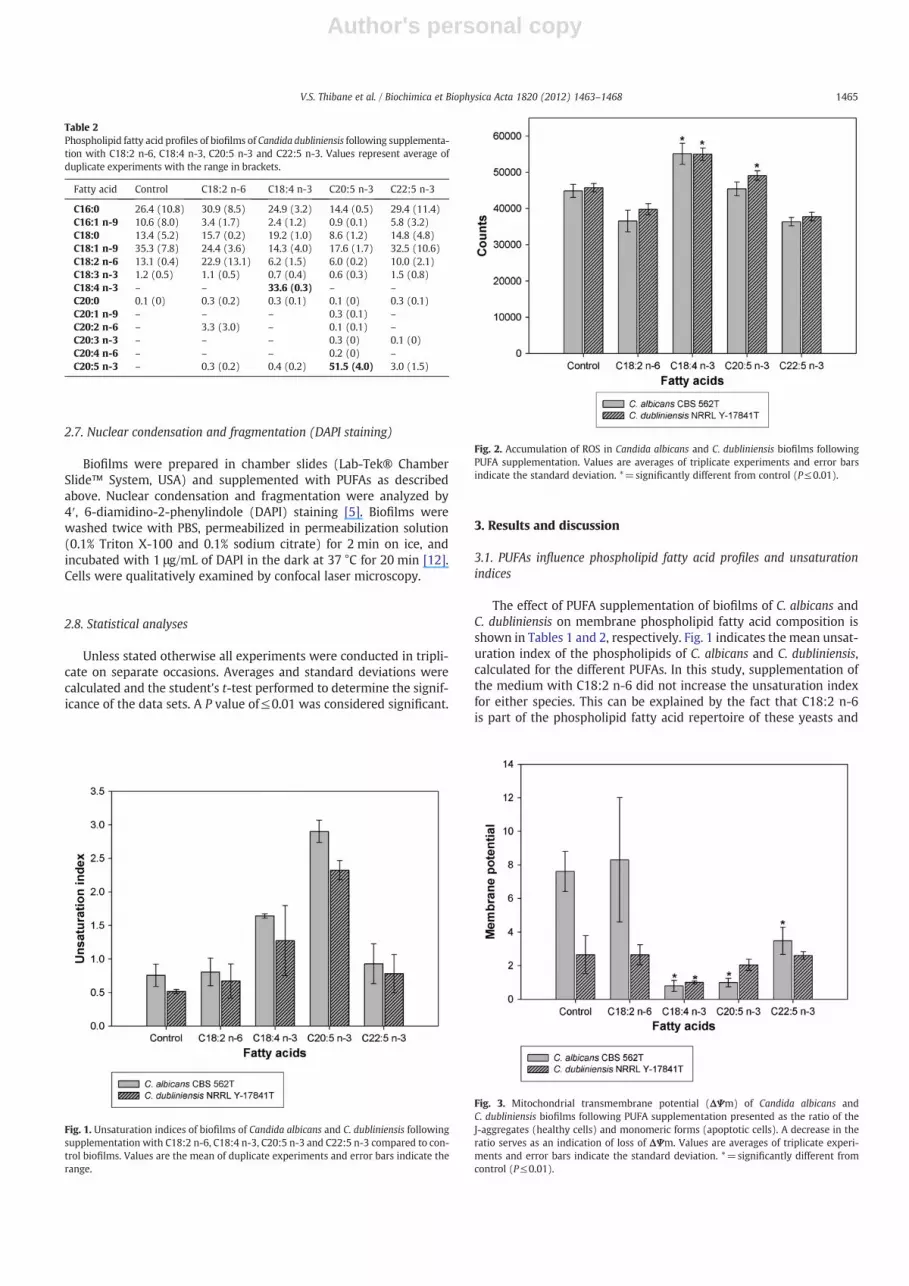
The effect of PUFA supplementation of biofilms of C. albicans andC. dubliniensis on membrane phospholipid fatty acid composition isshown in Tables 1 and 2, respectively. Fig. 1 indicates the mean unsat-uration index of the phospholipids of C. albicans and C. dubliniensis,calculated for the different PUFAs. In this study, supplementation ofthe medium with C18:2 n-6 did not increase the unsaturation indexfor either species. This can be explained by the fact that C18:2 n-6is part of the phospholipid fatty acid repertoire of these yeasts and
Table 2Phospholipid fatty acid profiles of biofilms of Candida dubliniensis following supplementa-tion with C18:2 n-6, C18:4 n-3, C20:5 n-3 and C22:5 n-3. Values represent average ofduplicate experiments with the range in brackets.
Fig. 1. Unsaturation indices of biofilms of Candida albicans and C. dubliniensis followingsupplementation with C18:2 n-6, C18:4 n-3, C20:5 n-3 and C22:5 n-3 compared to con-trol biofilms. Values are the mean of duplicate experiments and error bars indicate therange.
Fig. 2. Accumulation of ROS in Candida albicans and C. dubliniensis biofilms followingPUFA supplementation. Values are averages of triplicate experiments and error barsindicate the standard deviation. *=significantly different from control (P≤0.01).
Fig. 3. Mitochondrial transmembrane potential (ΔΨm) of Candida albicans andC. dubliniensis biofilms following PUFA supplementation presented as the ratio of theJ-aggregates (healthy cells) and monomeric forms (apoptotic cells). A decrease in theratio serves as an indication of loss of ΔΨm. Values are averages of triplicate experi-ments and error bars indicate the standard deviation. *=significantly different fromcontrol (P≤0.01).
1465V.S. Thibane et al. / Biochimica et Biophysica Acta 1820 (2012) 1463–1468
is recognized and readily metabolized through β-oxidation. Interest-ingly, supplementation with C18:4 n-3 and C20:5 n-3, which wereproposed to be inhibitory [1], lead to the incorporation of thesefatty acids into membrane phospholipids of both species, as high-lighted in Tables 1 and 2, and a resultant increase in unsaturationindex (Fig. 1). However, C22:5 n-3 was poorly incorporated into themembrane phospholipids. This may be explained by the increasedhydrophobicity of longer chain fatty acids.
3.2. PUFAs increase reactive oxygen species production
Fig. 2 shows a significant increase (P≤0.01) in ROS production aftersupplementation with C18:4 n-3 in both species as well as with C20:5n-3, in the case of C dubliniensis. This indicates that incorporation of
these two PUFAs into the phospholipids could potentially increaseoxidative stress in biofilms of C. albicans and C. dubliniensis, whichcould lead to an increase in lipid peroxidation and cell damage.
The loss of mitochondrial transmembrane potential (ΔΨm) is one ofseveral key events that occur in the mitochondria during early stages ofapoptosis [24]. There was a significant loss in ΔΨm (P≤0.01) when bio-films of C. albicans were supplemented with C18:4 n-3, C20:5 n-3 andC22:5 n-3 (Fig. 3). However, supplementation of the biofilm with C18:2n-6 did not have a significant effect compared to the control.
Although supplementation of C. dubliniensis biofilms with C18:4n-3 and C20:5 n-3 resulted in a significant increase in ROS production(P≤0.01) (Fig. 2) and supplementation with C18:4 n-3 also resultedin a significant loss in ΔΨm (P≤0.01) (Fig. 3), there were no signifi-cant differences in ΔΨm, compared to the control after supplementa-tion with C18:2 n-6, C20:5 n-3 and C22:5 n-3. This indicates thatespecially C18:4 n-3 has the ability to cause loss of ΔΨm in biofilmsof both Candida species.
3.4. PUFAs cause apoptosis
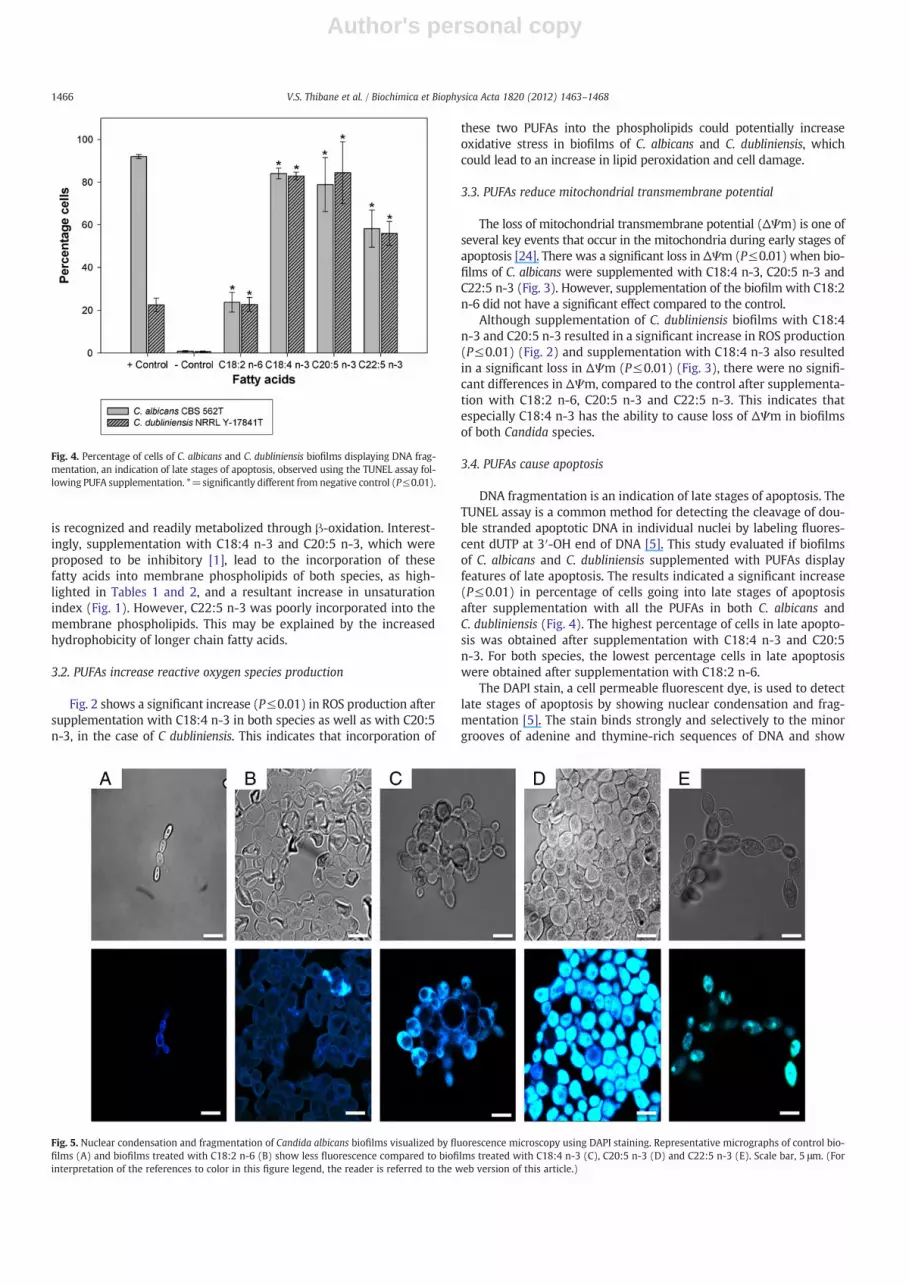
DNA fragmentation is an indication of late stages of apoptosis. TheTUNEL assay is a common method for detecting the cleavage of dou-ble stranded apoptotic DNA in individual nuclei by labeling fluores-cent dUTP at 3′-OH end of DNA [5]. This study evaluated if biofilmsof C. albicans and C. dubliniensis supplemented with PUFAs displayfeatures of late apoptosis. The results indicated a significant increase(P≤0.01) in percentage of cells going into late stages of apoptosisafter supplementation with all the PUFAs in both C. albicans andC. dubliniensis (Fig. 4). The highest percentage of cells in late apopto-sis was obtained after supplementation with C18:4 n-3 and C20:5n-3. For both species, the lowest percentage cells in late apoptosiswere obtained after supplementation with C18:2 n-6.
The DAPI stain, a cell permeable fluorescent dye, is used to detectlate stages of apoptosis by showing nuclear condensation and frag-mentation [5]. The stain binds strongly and selectively to the minorgrooves of adenine and thymine-rich sequences of DNA and show
Fig. 4. Percentage of cells of C. albicans and C. dubliniensis biofilms displaying DNA frag-mentation, an indication of late stages of apoptosis, observed using the TUNEL assay fol-lowing PUFA supplementation. *=significantly different from negative control (P≤0.01).
Fig. 5. Nuclear condensation and fragmentation of Candida albicans biofilms visualized by fluorescence microscopy using DAPI staining. Representative micrographs of control bio-films (A) and biofilms treated with C18:2 n-6 (B) show less fluorescence compared to biofilms treated with C18:4 n-3 (C), C20:5 n-3 (D) and C22:5 n-3 (E). Scale bar, 5 μm. (Forinterpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
1466 V.S. Thibane et al. / Biochimica et Biophysica Acta 1820 (2012) 1463–1468
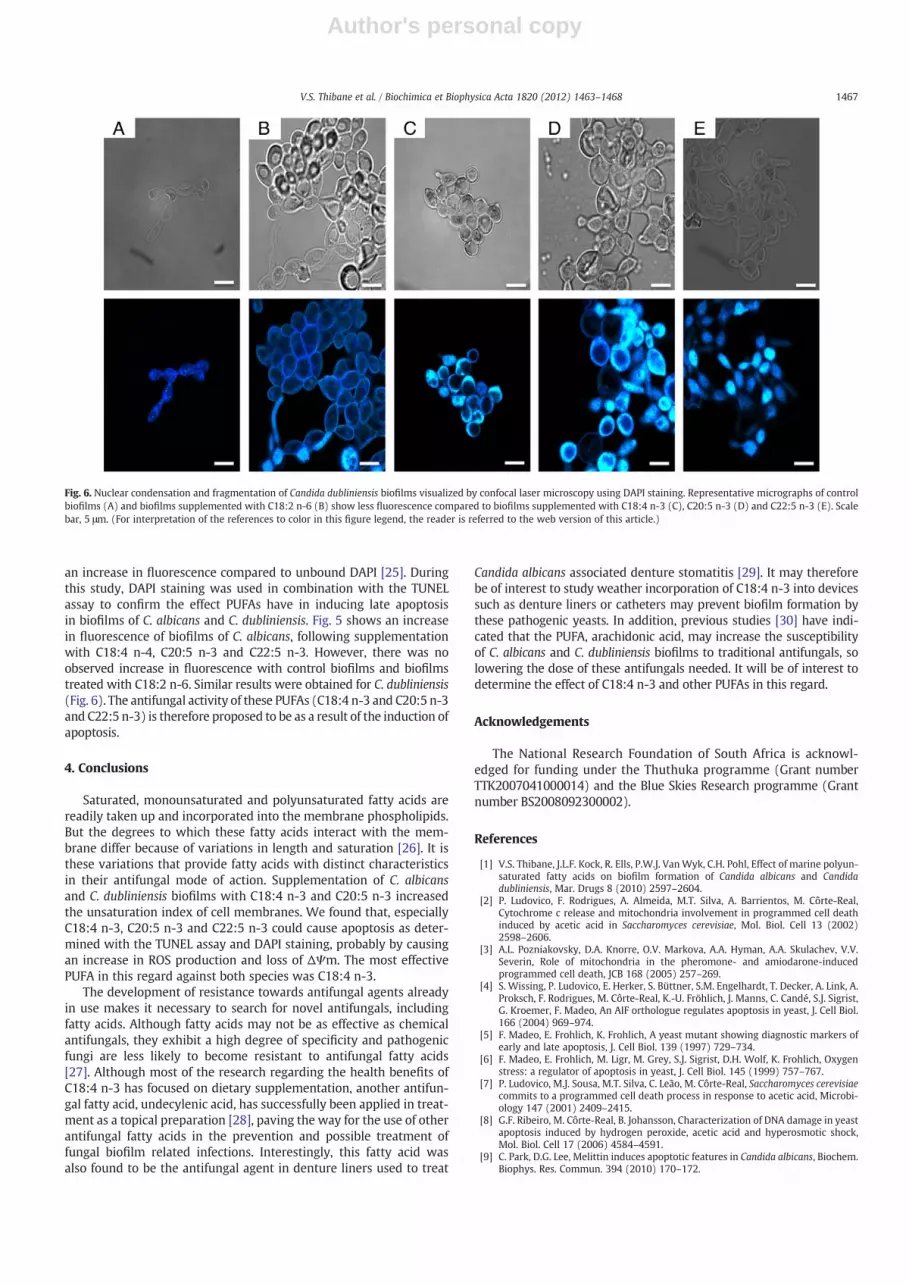
an increase in fluorescence compared to unbound DAPI [25]. Duringthis study, DAPI staining was used in combination with the TUNELassay to confirm the effect PUFAs have in inducing late apoptosisin biofilms of C. albicans and C. dubliniensis. Fig. 5 shows an increasein fluorescence of biofilms of C. albicans, following supplementationwith C18:4 n-4, C20:5 n-3 and C22:5 n-3. However, there was noobserved increase in fluorescence with control biofilms and biofilmstreated with C18:2 n-6. Similar results were obtained for C. dubliniensis(Fig. 6). The antifungal activity of these PUFAs (C18:4 n-3 and C20:5 n-3and C22:5 n-3) is therefore proposed to be as a result of the induction ofapoptosis.
4. Conclusions
Saturated, monounsaturated and polyunsaturated fatty acids arereadily taken up and incorporated into the membrane phospholipids.But the degrees to which these fatty acids interact with the mem-brane differ because of variations in length and saturation [26]. It isthese variations that provide fatty acids with distinct characteristicsin their antifungal mode of action. Supplementation of C. albicansand C. dubliniensis biofilms with C18:4 n-3 and C20:5 n-3 increasedthe unsaturation index of cell membranes. We found that, especiallyC18:4 n-3, C20:5 n-3 and C22:5 n-3 could cause apoptosis as deter-mined with the TUNEL assay and DAPI staining, probably by causingan increase in ROS production and loss of ΔΨm. The most effectivePUFA in this regard against both species was C18:4 n-3.
The development of resistance towards antifungal agents alreadyin use makes it necessary to search for novel antifungals, includingfatty acids. Although fatty acids may not be as effective as chemicalantifungals, they exhibit a high degree of specificity and pathogenicfungi are less likely to become resistant to antifungal fatty acids[27]. Although most of the research regarding the health benefits ofC18:4 n-3 has focused on dietary supplementation, another antifun-gal fatty acid, undecylenic acid, has successfully been applied in treat-ment as a topical preparation [28], paving the way for the use of otherantifungal fatty acids in the prevention and possible treatment offungal biofilm related infections. Interestingly, this fatty acid wasalso found to be the antifungal agent in denture liners used to treat
Candida albicans associated denture stomatitis [29]. It may thereforebe of interest to study weather incorporation of C18:4 n-3 into devicessuch as denture liners or catheters may prevent biofilm formation bythese pathogenic yeasts. In addition, previous studies [30] have indi-cated that the PUFA, arachidonic acid, may increase the susceptibilityof C. albicans and C. dubliniensis biofilms to traditional antifungals, solowering the dose of these antifungals needed. It will be of interest todetermine the effect of C18:4 n-3 and other PUFAs in this regard.
Acknowledgements
The National Research Foundation of South Africa is acknowl-edged for funding under the Thuthuka programme (Grant numberTTK2007041000014) and the Blue Skies Research programme (Grantnumber BS2008092300002).
References
[1] V.S. Thibane, J.L.F. Kock, R. Ells, P.W.J. VanWyk, C.H. Pohl, Effect of marine polyun-saturated fatty acids on biofilm formation of Candida albicans and Candidadubliniensis, Mar. Drugs 8 (2010) 2597–2604.
[2] P. Ludovico, F. Rodrigues, A. Almeida, M.T. Silva, A. Barrientos, M. Côrte-Real,Cytochrome c release and mitochondria involvement in programmed cell deathinduced by acetic acid in Saccharomyces cerevisiae, Mol. Biol. Cell 13 (2002)2598–2606.
[3] A.L. Pozniakovsky, D.A. Knorre, O.V. Markova, A.A. Hyman, A.A. Skulachev, V.V.Severin, Role of mitochondria in the pheromone- and amiodarone-inducedprogrammed cell death, JCB 168 (2005) 257–269.
[4] S. Wissing, P. Ludovico, E. Herker, S. Büttner, S.M. Engelhardt, T. Decker, A. Link, A.Proksch, F. Rodrigues, M. Côrte-Real, K.-U. Fröhlich, J. Manns, C. Candé, S.J. Sigrist,G. Kroemer, F. Madeo, An AIF orthologue regulates apoptosis in yeast, J. Cell Biol.166 (2004) 969–974.
[5] F. Madeo, E. Frohlich, K. Frohlich, A yeast mutant showing diagnostic markers ofearly and late apoptosis, J. Cell Biol. 139 (1997) 729–734.
[6] F. Madeo, E. Frohlich, M. Ligr, M. Grey, S.J. Sigrist, D.H. Wolf, K. Frohlich, Oxygenstress: a regulator of apoptosis in yeast, J. Cell Biol. 145 (1999) 757–767.
[7] P. Ludovico, M.J. Sousa, M.T. Silva, C. Leão, M. Côrte-Real, Saccharomyces cerevisiaecommits to a programmed cell death process in response to acetic acid, Microbi-ology 147 (2001) 2409–2415.
[8] G.F. Ribeiro, M. Côrte-Real, B. Johansson, Characterization of DNA damage in yeastapoptosis induced by hydrogen peroxide, acetic acid and hyperosmotic shock,Mol. Biol. Cell 17 (2006) 4584–4591.
[9] C. Park, D.G. Lee, Melittin induces apoptotic features in Candida albicans, Biochem.Biophys. Res. Commun. 394 (2010) 170–172.
Fig. 6. Nuclear condensation and fragmentation of Candida dubliniensis biofilms visualized by confocal laser microscopy using DAPI staining. Representative micrographs of controlbiofilms (A) and biofilms supplemented with C18:2 n-6 (B) show less fluorescence compared to biofilms supplemented with C18:4 n-3 (C), C20:5 n-3 (D) and C22:5 n-3 (E). Scalebar, 5 μm. (For interpretation of the references to color in this figure legend, the reader is referred to the web version of this article.)
1467V.S. Thibane et al. / Biochimica et Biophysica Acta 1820 (2012) 1463–1468
Author's personal copy
[10] B. Hwang, J. Hwang, J. Lee, D.G. Lee, The antimicrobial peptide, psacotheasininduces reactive oxygen species and triggers apoptosis in Candida albicans, Bio-chem. Biophys. Res. Commun. 405 (2011) 267–271.
[11] J. Cho, D.G. Lee, Oxidative stress by antimicrobial peptide pleurocidin triggersapoptosis in Candida albicans, Biochimie 93 (2011) 1873–1879.
[12] B. Hwang, J. Hwang, J. Lee, J. Kim, S.R. Kim, Y. Kim, D.G. Lee, Induction of yeastapoptosis by an antimicrobial peptide, Papiliocin, Biochem. Biophys. Res. Commun.408 (2011) 89–93.
[13] J. Cho, D.G. Lee, The antimicrobial peptide arenicin-1 promotes generation ofreactive oxygen species and induction of apoptosis, Biochim. Biophys. Acta 1810(2011) 1246–1251.
[14] G.G. Perrone, S. Tan, I.W. Dawes, Reactive oxygen species and yeast apoptosis,Biochim. Biophys. Acta 1783 (2008) 1354–1368.
[15] T.M. Lima, C.C. Kanunfre, C. Pompéia, R. Verlengia, R. Curi, Ranking the toxicity of fattyacids on Jurkat and Raji cells by flow cytometric analyses, Toxicol. In Vitro 16 (2002)741–747.
[16] T.C. Ferreira, L.M. Pepe de Moraes, E.G. Campos, Cell density-dependent linoleicacid toxicity to Saccharomyces cerevisiae, FEMS 11 (2011) 408–417.
[17] Y.M. Samaranayake, P.C. Wu, L.P. Samaranayake, Relationship between the cellsurface hydrophobicity and adherence of Candida krusei and Candida albicans toepithelial and denture acrylic surfaces, APMIS 103 (1995) 707–713.
[18] J. Folch, M. Lees, S.G.H. Sloane, A simple method for the isolation and purificationof total lipids from animal tissues, J. Biol. Chem. 226 (1957) 497–509.
[19] D.A. Bossio, K.M. Scow, Impacts of carbon and flooding on soil microbial commu-nities: phospholipid fatty acid profiles and substrate utilization patterns, Microb.Ecol. 35 (1998) 265–278.
[20] H.T. Slover, E. Lanza, Quantitative analyses of food fatty acids by capillary gaschromatography, J. Am. Oil Chem. Soc. 56 (1979) 933–943.
[21] D. Kobayashi, K. Kondo, N. Uehara, S. Otokozawa, N. Tsiju, A. Yagihashi, N.Watanabe, Endogenous reactive oxygen species is an important mediatorof miconazole antifungal effect, Antimicrob. Agents Chemother. 46 (2002)3113–3117.
[22] W. Cheng, H. Tong, E.W. Miller, C.J. Chang, J. Remington, R.M. Zucker, P.A. Bromberg,J.M. Samet, T.P.J. Hofer, An integrated imaging approach to the study of oxidativestress generation by mitochondrial dysfunction in living cells, Environ. HealthPerspect. 118 (2010) 902–908.
[23] A.J. Phillips, I. Sudbery, M. Ramsdale, Apoptosis induced by environmental stress-es and amphotericin B in Candida albicans, PNAS 100 (2003) 14327–14332.
[24] D.R. Green, J.C. Reed, Mitochondria and apoptosis, Science 281 (1998) 1309–1312.[25] J. Kapuscinski, DAPI: a DNA-specific fluorescent probe, Biotech. Histochem. 70
(1995) 220–233.[26] S. Liu, R. Weibin, L. Jing, X. Hua, W. Jingan, G. Yubao, W. Jingguo, Biological control
of phytopathogenic fungi by fatty acids, Mycopathologia 166 (2008) 93–103.[27] C.H. Pohl, J.L.F. Kock, V.S. Thibane, Antifungal free fatty acids: a review, in: A.
Méndez-Vilas (Ed.), Science against microbial pathogens: current researchand technological advances, vol. 1, Formatex Research Center, Spain, 2011,pp. 61–71.
[28] X.-C. Li, M.R. Jacob, S.I. Khan, M.K. Ashfaq, K.S. Babu, A.K. Agarwal, H.N. ElShody,S.P. Manly, A.M. Clark, Potent in vitro antifungal activity of naturally occurringacetylenic acids, Antimicrob. Agents Chemother. 52 (2008) 2442–2448.
[29] N. McLain, R. Ascanio, C. Baker, R.A. Strohaver, J.W. Dolan, Undecylenic acidinhibits morphogenesis of Candida albicans, Antimicrob. Agents Chemother. 44(2000) 2873–2875.
[30] R. Ells, J.L.F. Kock, P.W.J. van Wyk, P.J. Botes, C.H. Pohl, Arachidonic acid increasesantifungal susceptibility of Candida albicans and Candida dubliniensis, J. Anti-microb. Chemother. 63 (2009) 124–128.
1468 V.S. Thibane et al. / Biochimica et Biophysica Acta 1820 (2012) 1463–1468