RESEARCH New Phytol. (2000), 146, 535–544 Saprotrophic invasion by the soil-borne fungal plant pathogen Rhizoctonia solani and percolation thresholds D. J. BAILEY*, W. OTTEN, C. A. GILLIGAN Department of Plant Sciences, University of Cambridge, Downing Street, Cambridge CB2 3EA, UK Received 9 August 1999 ; accepted 16 February 2000 In this paper we distinguish between invasive and noninvasive (finite) saprotrophic spread of the soil-borne fungal plant pathogen, Rhizoctonia solani amongst discrete sites of nutrient resource. Using simple concepts of percolation theory, we predict the critical threshold distance, associated with a threshold probability, between donor (colonized) and recipient (uncolonized) nutrient sites at which R. solani can spread invasively by mycelial growth through a population of nutrient sites on a lattice. The critical distance for invasive spread is estimated from colonization profiles derived from placement experiments that summarize the probability of colonization with distance between replicated pairs of colonized and uncolonized sites. Colonization profiles were highly nonlinear, decaying sigmoidally with distance. Thresholds for invasive spread were predicted at inter-site distances of 8.1 mm and 11.8 mm for sites of low and high nutrient agar, respectively. In population experiments with inter-site distances below the predicted thresholds, the spread of the fungus was invasive in all replicates. At large distances ("10 mm for low, and "14 mm for high nutrient sites) the spread of the fungus was always finite, with the proportion of finite replicates decreasing sharply close to the percolation threshold. Invasive spread did not depend on the furthest extent of growth of the fungus but on distances predicted by the percolation thresholds. Invasive spread of the fungus is also examined in a more natural and variable, nonsterile system involving the growth and colonization of a lattice of poppy seeds over sand. The system is characterized by a decay in the probability of colonization between older poppy seeds, which effectively ‘ quenches ’ saprotrophic spread. Hence in the population experiments with poppy seeds all growth was ultimately finite. The threshold distance, corresponding to the critical percolation probability for invasive growth changed from 18 mm to 4 mm over 21d leading to a switch from invasive to finite growth. We conclude that percolation theory can be used to link the growth of individual mycelial colonies to the formation of patches that result from the colonization of particulate organic matter. The nonlinearity of the colonization profiles combined with the presence of a percolation threshold means that small changes in the distance between nutrient sites can result in large differences in final patch size. The rapid decay of particulate organic matter in a more natural system can have a profound effect on the dynamics of colonization, restricting saprotrophic invasion of the soil. The consequences of invasion thresholds for colony growth of saprotrophic and parasitic fungi in dynamical systems are briefly discussed. Key words : percolation, heterogeneity, Rhizoctonia solani, invasive spread, pathozone. One important feature in the invasion and per- sistence of many ecologically and economically important fungal parasites and saprotrophs in soil is the ability of the fungus to spread by mycelial growth and expansion of fungal colonies. The lateral spread of parasites depends on the endogenous supply and translocation of nutrients within the fungal colony, the growth-habit of the colony and the distances between susceptible host roots or other organs. If susceptible hosts are too far apart, local *Author for correspondence (tel ›44 1223 333 900 ; fax ›44 1223 333 953 ; e-mail djb21!cus.cam.ac.uk). invasion ceases as the fungus exhausts its nutrient supply before infecting a new host. Similar con- straints apply to the transmission of mycorrhizal fungi between infected and uninfected roots (Smith & Read, 1997) and to the transmission of saprotrophs between discrete fragments of colonized and uncolonized organic matter. In each case it is convenient, at least initially, to visualize spread occurring through a population of discrete sites on a lattice. The sites correspond to discrete nutrient sources, comprising susceptible roots or discrete fragments of organic matter, and the entire lattice represents the soil matrix in which spread occurs. The lattice might
Transcript
RESEARCH New Phytol. (2000), 146, 535–544
Saprotrophic invasion by the soil-borne
fungal plant pathogen Rhizoctonia solani
and percolation thresholds
D. J. BAILEY*, W. OTTEN, C. A. GILLIGAN
Department of Plant Sciences, University of Cambridge, Downing Street,
Cambridge CB2 3EA, UK
Received 9 August 1999 ; accepted 16 February 2000
In this paper we distinguish between invasive and noninvasive (finite) saprotrophic spread of the soil-borne fungal
plant pathogen, Rhizoctonia solani amongst discrete sites of nutrient resource. Using simple concepts of
percolation theory, we predict the critical threshold distance, associated with a threshold probability, between
donor (colonized) and recipient (uncolonized) nutrient sites at which R. solani can spread invasively by mycelial
growth through a population of nutrient sites on a lattice. The critical distance for invasive spread is estimated
from colonization profiles derived from placement experiments that summarize the probability of colonization
with distance between replicated pairs of colonized and uncolonized sites. Colonization profiles were highly
nonlinear, decaying sigmoidally with distance. Thresholds for invasive spread were predicted at inter-site
distances of 8.1 mm and 11.8 mm for sites of low and high nutrient agar, respectively. In population experiments
with inter-site distances below the predicted thresholds, the spread of the fungus was invasive in all replicates. At
large distances ("10 mm for low, and "14 mm for high nutrient sites) the spread of the fungus was always finite,
with the proportion of finite replicates decreasing sharply close to the percolation threshold. Invasive spread did
not depend on the furthest extent of growth of the fungus but on distances predicted by the percolation thresholds.
Invasive spread of the fungus is also examined in a more natural and variable, nonsterile system involving the
growth and colonization of a lattice of poppy seeds over sand. The system is characterized by a decay in the
probability of colonization between older poppy seeds, which effectively ‘quenches’ saprotrophic spread. Hence
in the population experiments with poppy seeds all growth was ultimately finite. The threshold distance,
corresponding to the critical percolation probability for invasive growth changed from 18 mm to 4 mm over 21d
leading to a switch from invasive to finite growth. We conclude that percolation theory can be used to link the
growth of individual mycelial colonies to the formation of patches that result from the colonization of particulate
organic matter. The nonlinearity of the colonization profiles combined with the presence of a percolation threshold
means that small changes in the distance between nutrient sites can result in large differences in final patch size.
The rapid decay of particulate organic matter in a more natural system can have a profound effect on the dynamics
of colonization, restricting saprotrophic invasion of the soil. The consequences of invasion thresholds for colony
growth of saprotrophic and parasitic fungi in dynamical systems are briefly discussed.
Fig. 3. Change in the number of agar sites colonized by Rhizoctonia solani with time for replicate populations
with different distances between sites (given in the top left of each plot) arranged in a triangular lattice for high
nutrient (10% potato dextrose agar (PDA)) and low nutrient (1% PDA) agar. Solid lines indicate replicates
exhibiting invasive spread (patches had reached the edge of the system after 21 d) and dotted lines, replicates
showing finite spread (patches which had not spread to the edge of the system).
System II: percolation with substrate decay (poppy
seed)
Changes in the probability of colonization with distance
and time. To examine the effect of seed (inoculum)
decay on the threshold distance for invasive spread,
rc, probability profiles were constructed using seeds
that had been incubated for three different times (0,
7 and 21 d) on nonsterile sand. Poppy seeds (Papaver
somniferum L.) measuring approx. 1 mm in diameter
were initially sterilized by autoclaving for 1 h at
121 kPa. Large Petri plates (14-cm diameter) were
filled with 200 g of sand (Hepworth Minerals &
Chemicals Ltd, Redhill, UK; Grade 16}30, with
10% gravimetric water content). Donor seeds were
positioned on the sand surface at 0, 3, 6, 9, 12, 15, 18,
21, 25 and 27 mm from uncolonized seeds. Donor
seeds were inoculated with a single hyphal strand
of R. solani removed from the growing edge of a
4-d-old colony grown on water agar either immedi-
RESEARCH Saprotrophic invasion by Rhizoctonia solani 539
ately or after 7 or 21 d incubation of the seeds on
sand in the dark at 23°C. There were 15 replicates
per treatment and the plates were assessed daily for
colonization until no change in the number of seeds
colonized was detected.
Patch formation and seed density. The dynamics of
patch formation were examined for R. solani growing
through a population of poppy seeds. Replicate seed
trays (240¬360 ¬50 mm deep) were filled to a depth
of 4 cm with sand (Hepworth Minerals & Chemicals
Ltd). Sterile poppy seeds were placed on the surface
of the sand in a triangular lattice. Nine seed densities
were prepared with distances between neighbouring
seeds of 12, 14, 16, 18, 20, 22, 24, 26, and 28 mm;
each density was replicated five times. Spread and
colonization of the seeds was initiated from the
central poppy seed, colonized by R. solani. The trays
were sealed in plastic bags to reduce evaporation and
incubated in the dark at 23°C. The bags were opened
daily and the poppy seeds were assessed for
colonization using a binocular microscope (¬40)
for 27 d.
System I: percolation with no decay of substrate
(agar spots)
Predicting the threshold distance for invasion from
profiles describing changes in the probability of
colonization with distance. The probability of
colonization, P, decayed sigmoidally as the distance,
r, between donor and recipient sites increased (Fig.
2). Threshold distances, rc, for bond percolation on
a lattice (corresponding to P ¯ Pc
¯ 0.35) were
obtained by inverse prediction from a logistic model,
P ¯ θ"}(1exp (θ
#(r®θ
$))), where θ
iare parameters.
The model was fitted to the experimental data by
maximum likelihood under the assumption of bi-
nomial errors using Genstat (Numerical Algorithms
Group Ltd, Oxford, UK). Increasing the nutrient
status of the agar resulted in a steeper profile
shifted to the right and an increase in the threshold
distance between sites from rc¯ 8.1³0.49 mm for
low nutrient sites to rc
¯ 11.8³0.93 mm for sites
with a high nutrient status (Fig. 2).
Effect of inter-site distance and nutrient status on finite
and invasive spread in populations. Increasing the
distance between sites and reducing the nutrient
status of sites created finite patches (Figs 3, 4). For
high nutrient sites, all replicates with inter-site
distances % 10 mm displayed invasive growth. At
12 mm, five out of six replicates produced finite
patches and at 14 and 16 mm all replicates were finite.
The threshold distance between sites for invasive
spread was estimated from the population data
(Fig. 3) at approx. 8.0 mm and 12 mm for low and
Invasive spread Finite spreadTime after
innoculation(d)
4
12
20
Fig. 4. Examples of replicate microcosms showing patches
with invasive and finite growth of Rhizoctonia solanibetween sites of low (1% potato dextrose agar) nutrient
agar. Note that gaps occur in invasive patches. Distance
between sites : 6 mm, invasive spread; 8 mm, finite spread.
high nutrient sites, respectively. These estimates
were consistent with the predicted thresholds of 8.1
and 11.8 mm obtained from the colonization profiles
for pairs of nutrient sites (Fig. 2). However, maps
of invasive spread (Fig. 4) show that coverage is
not complete and gaps occur even within invasive
patches.
Effect of the probability of colonization on finite and
invasive spread. The proportion of patches exhibiting
invasive spread was plotted against inter-site dis-
tance (Fig. 5a) and the corresponding probability
(derived from Fig. 2; Fig. 5b). The relationships
were highly nonlinear with a very marked change in
the proportion of invasive patches over short
distances. Whereas invasive spread occurred at
different inter-site distances for high and low
nutrient status (Fig. 5a), there was no difference in
the empirical estimate of the critical probability (Fig.
5b). The latter probability was estimated as
0.365³0.07 by inverse prediction after fitting a
logistic function to the data for the combined profiles
(Fig. 5b).
System II: percolation with decay of substrate
(poppy seed)
Changes in the probability of colonization with distance
and time. The probability of colonization of recipient
poppy seeds declined sigmoidally with distance from
the donor (Fig. 6). Compared with the more artificial
540 RESEARCH D. J. Bailey et al.
1.0
0.8
0.6
0.4
0.2
0.0
0 2 4 6 8 10 12 14 16 18
Inter-site distance (r, mm)
0.0 0.2 0.4 0.6 0.8 1.0
Probability of colonization (P) betweendonor and recipient
1.0
0.8
0.6
0.4
0.2
0.0
(a) (b)
Pro
po
rtio
n o
f p
atch
es s
ho
win
gin
vasi
ve s
pre
ad (
Pi)
Fig. 5. Change in the proportion of patches of Rhizoctonia solani exhibiting invasive spread (Pi) with (a) distance
between sites and (b) probability of colonization between donor and recipient sites (P), for growth amongst a
population of sites of low (1% potato dextrose agar (PDA), open circles) and high (10% PDA, crosses) nutrient
agar. The probability of colonization (P) was estimated from the relationship in Fig. 2, given the inter-site
distance, r. The fitted curves relating the proportion of invasive patches to inter-site distance are given by the
logistic functions Pi¯ 1}(1exp(5.0(r® 11.18) and P
i¯ 1}(1exp(5.0(r®7.5) for high and low nutrient agar,
respectively (Fig. 5a), and by Pi¯ 1}(1exp(®25(P®0.45))) relating the proportion of invasive patches to the
probability of colonization between individual sites (Fig. 5b). Note that, whilst invasive spread occurs at
different inter-site distances for high and low nutrient sites, the probability at which invasive spread occurs is
not affected.
1.0
0.8
0.6
0.4
0.2
0.0
0 5 10 15 20 25 30
Distance (r, mm)
Pro
bab
ility
of
colo
niz
atio
n (
P)
Fig. 6. Change in the probability of colonization, P, by Rhizoctonia solani with distance, r, between pairs of
poppy seeds 0 (circles), 7 (squares) or 21 (triangles) d old. Data are fitted with the logistic functions P ¯1}(1exp(0±25(r®15.83))), P ¯ 1}(1exp(0.71(r®7.66))) and P ¯ 1}(1exp(0.93(r®3.62))) for 0, 7 and 21-
d-old poppy seed, respectively, to provide profiles for the probability of colonization with distance.
agar spots, the poppy seed profile was shifted further
to the right but dropped less steeply. Ageing of
inoculum before exposure of recipients suppressed
colonization, shifting the curve to the left (Fig. 6).
Threshold probabilities, rc, for invasion (P
c¯ 0.35,
for bond percolation on a triangular lattice) corre-
sponded with a maximum distance of 18 mm
between neighbouring seeds for young inoculum
but declined to 8.0 mm for seeds incubated on sand
for 7 d and to 4.0 mm for seed incubated for 21 d
(Fig. 6).
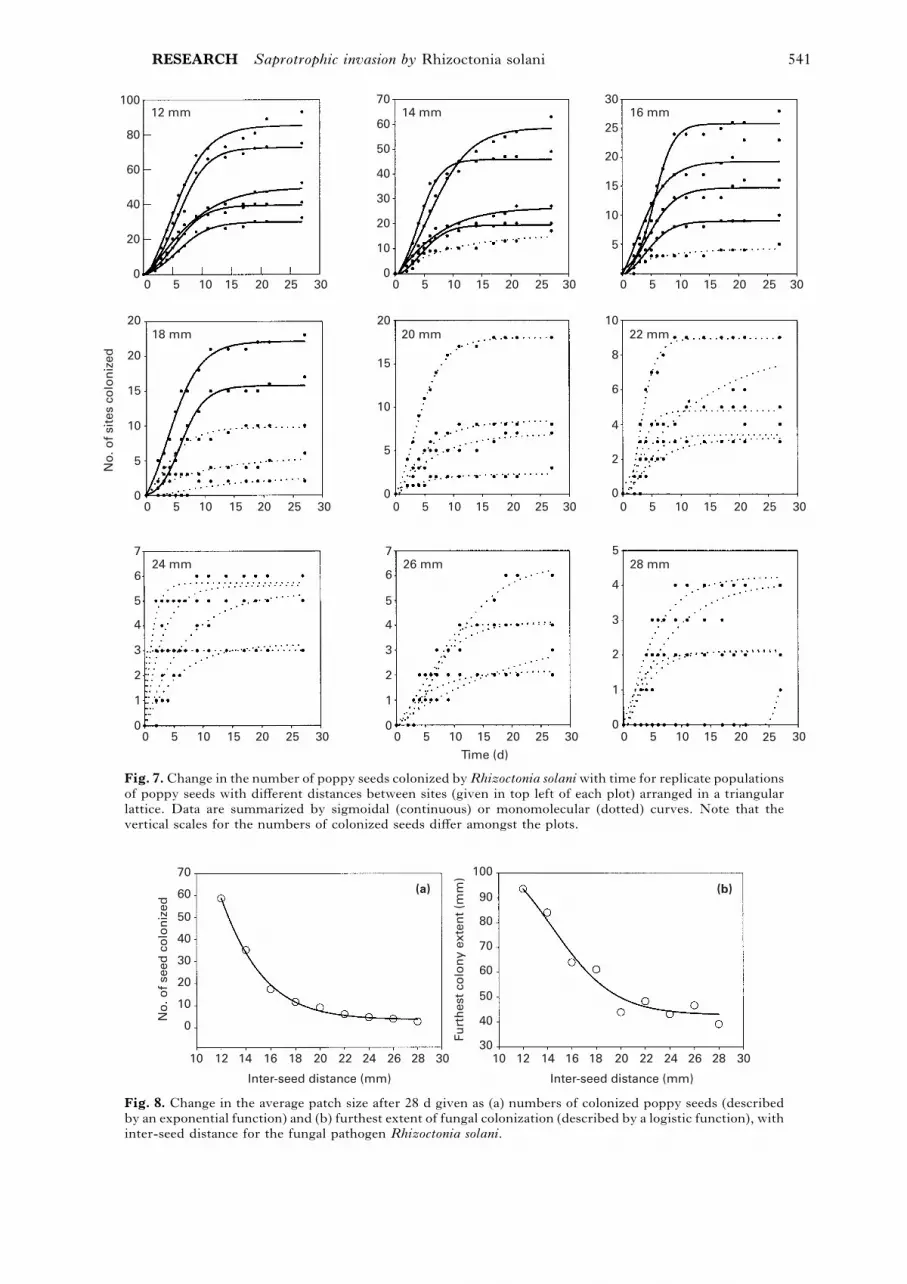
Dynamics of fungal spread. We summarize the
dynamics of fungal spread for individual replicates
by describing changes in the number of colonized
seeds over time for each replicate population (Fig. 7).
Although none of the patches had reached the edge
of the system after 28 d, the rate of colonization had
slowed to almost zero in all replicates, with the
exception of a late and meagre start of one replicate
at the maximum distance.
There was a marked difference in the extent and
rate of spread of the fungus above and below the
RESEARCH Saprotrophic invasion by Rhizoctonia solani 541