Side-stepping secondary symbionts widespread horizontal transfer across and beyond the Aphidoidea
J A RUSSELL
A LATORRE
dagger
B SABATER-MUNtildeOZ
dagger
A MOYA
dagger
and N A MORAN
University of Arizona Department of Ecology and Evolutionary Biology Biological Sciences West Tucson Arizona 85721 USA
dagger
Universitat de Valegravencia mdash Institut Cavanilles Genegravetica Evolutiva Edifici drsquoInstitutos de Paterna Apartado 22085 Valegravencia Spain
Abstract
To elucidate the co-evolutionary relationships between phloem-feeding insects and theirsecondary or facultative bacterial symbionts we explore the distributions of three suchmicrobes mdash provisionally named the R-type (or PASS or S-sym) T-type (or PABS) and U-type mdash across a number of aphid and psyllid hosts through the use of diagnostic molecularscreening techniques and DNA sequencing Although typically maternally transmittedphylogenetic and pairwise divergence analyses reveal that these bacteria have been inde-pendently acquired by a variety of unrelated insect hosts indicating that horizontal transferhas helped to shape their distributions Based on the high genetic similarity between symbiontsin different hosts we argue that transfer events have occurred recently on an evolutionarytimescale In several instances however closely related symbionts associate with relatedhosts suggesting that horizontal transfer between distant relatives may be rarer than trans-mission between close relatives Our findings on the prevalence of these symbionts withinmany aphid taxa along with published observations concerning their effects on host fit-ness imply a significant role of facultative symbiosis in aphid ecology and evolution
Keywords
aphid PASS psyllid R-type T-type U-type
Received 30 August 2002 revision received 10 December 2002 accepted 10 December 2002
Introduction
Many insects and bacteria engage in intimate associationsor symbioses Molecular analyses such as DNA sequencingand comparative sequence analyses which have allowedfor additional approaches such as
in situ
hybridization anddiagnostic polymerase chain reaction (PCR) have playeda key role in the identification of the various microbialparticipants These techniques have also been used todescribe a number of intriguing patterns regarding thediversity of bacterial symbionts within a given host cladethe distributions of closely related symbionts the modes oftransmission which shape these distributions and the agesof associations between hosts and symbionts
Aphids (Hemiptera Aphidoidea) represent an idealgroup of organisms for studies of symbiosis Nearly allmembers of the Aphidoidea harbour obligate endosym-bionts known as
Buchnera aphidicola
(Munson
et al
1991but see Fukatsu
et al
1994) mdash vertically transmitted (ie fromparent to offspring) bacteria housed within specialized
host cells known as bacteriocytes (or mycetocytes)
Buch-nera aphidicola
supplement the phloem-sap diets of theiraphid hosts through the synthesis of essential amino acids(Douglas amp Prosser 1992 Baumann
et al
1995 Sandstroumlmamp Moran 1999) The distributions of these symbionts canbe explained by a pattern of strict vertical transmission(Munson
et al
1991 Rouhbakhsh
et al
1996 Clark
et al
2000 Funk
et al
2000) since the time of a common ancestor84ndash164 million years ago (von Dohlen amp Moran 2000)
In addition many aphids harbour vertically transmittedbacteria that are not required for growth or reproductionThese are often referred to as secondary endosymbiontsMolecular and microscopy studies of several aphid specieshave revealed that these symbionts reside in a variety oflocations inside their hosts are sporadically distributedwithin and across host taxa and comprise a smallerproportion of an embryorsquos symbiotic flora than does theprimary symbiont
B aphidicola
(Buchner 1965 Unterman
et al
1989 Chen
et al
1996 Chen amp Purcell 1997 Fukatsu
et al
2000 Fukatsu 2001 Fukatsu
et al
2001 Sandstroumlm
et al
2001) Secondary symbionts of aphids belong to a numberof distinct lineages within the
γ
- and
α
-Proteobacteria(Unterman
et al
1989 Chen
et al
1996 Chen amp Purcell 1997
Correspondence J A Russell Fax 520ndash621ndash9190 E-mailjarusseluarizonaedu
1062
J A R U S S E L L
E T A L
copy 2003 Blackwell Publishing Ltd
Molecular Ecology
12 1061ndash1075
Fukatsu 2001 Sandstroumlm
et al
2001) and the Mollicutes(Fukatsu
et al
2001) Close relatives of aphid secondarieswith 16S rDNA sequence identities ranging from 98 toover 99 have been found in distantly related arthropodspecies (Chen
et al
1996 Fukatsu
et al
2001 Sandstroumlm
et al
2001) Thus for the few hosts that have been exam-ined the distributions of several secondary symbiontsappear to be partly the result of horizontal transmission(ie movement between hosts other than that from parentto offspring) between host species However some evid-ence suggests that certain associations between aphidsand secondary symbionts are specialized and have co-evolved (Fukatsu 2001)
To date most DNA sequence-based studies of aphid sec-ondary symbionts have focused on hosts within the tribeMacrosiphini Moreover although these studies providedsome evidence for horizontal transfer (Chen amp Purcell1997 Darby
et al
2001 Sandstroumlm
et al
2001) few haveemployed statistical analyses to demonstrate this pointand little effort has been made to assess the extent to whichhorizontal transfer has shaped secondary symbiont dis-tributions Here we use diagnostic molecular screeningtechniques and DNA sequencing along with pairwisedivergence and phylogenetic analyses to explore the dis-tributions of three facultative
γ
-Proteobacterial secondarysymbionts of aphids known as the R-type (or S-sym orPea Aphid Secondary Symbiont mdash PASS) T-type (or PeaAphid
Bemisia
-like Symbiont mdash PABS) and U-type (Chenamp Purcell 1997 Unterman
et al
1989 Darby
et al
2001Sandstroumlm
et al
2001) across a variety of aphid and psyllid(Hemiptera Psylloidea) species Specifically we comparethe phylogenies of hosts and symbionts to determine whe-ther there is any evidence for co-speciation Also we com-pare 16S rDNA divergence values between primary andsecondary symbionts residing within pairs of hosts sharingthe same most recent common ancestor Differences inthese values are used to determine the likelihood that cur-rent secondary infections could possibly be the result ofexclusive vertical transmission since the time of this com-mon host ancestor We also compare the rates of substitu-tion in the 16S rRNA genes of R- T- and U-type symbiontsto those of free-living bacteria and
B aphidicola
and esti-mate the ages of symbiont common ancestors through theuse of a molecular clock Finally we use data on symbiontprevalence within and across host taxa along with obser-vations of symbiont-associated fitness effects to assess thepotential importance of these microbes in aphid biology
Materials and methods
Samples
Members of 75 species spanning 15 subfamilies and tribesacross the Aphidoidea (Remaudiegravere amp Remaudiegravere 1997)
were collected from a variety of locations in the UnitedStates and Eurasia In addition members of four aphidspecies were obtained from laboratory stocks A list of allaphid species involved in the survey their taxonomicaffiliations and information regarding their collection ispresented in Table 1
Representatives of 27 psyllid species from North andSouth America along with Europe (see Thao
et al
2000b forcollection information) were also surveyed as psyllidshave been shown to harbor a number of secondary sym-bionts (Thao
et al
2000a) yet have not been screened forthe R- T- or U-types The samples included were
Acizziauncatoides
(two samples)
Aphalaroida inermis
Bactericeracockerelli
Blastopsylla occidentalis
Boreioglycaspis melaleucae
Cacopsylla brunneipennis
Cacopsylla myrthi
Cacopsylla pere-grina
Cacopsylla pyri
Calophya schini
Ctenarytaina eucalypti
Ctenarytaina longicauda
Ctenarytaina spatulata
Diaphorinacitri
Glycaspis brimblecombei
Heteropsylla cubana
Heterop-sylla texana
Neotriozella hirsuta
Psylla buxi
Trioza urticae
Pachypsylla celtidismamma
Pachypsylla venusta
Panisopelma
sp
Psylla
sp
Russelliana intermedia
Spanioneura fonscolombii
and
Trioza eugeniae
To determine the phylogenetic proximity of related sym-
bionts found within the same host species the 16S rRNAgenes of R- T- or U-type symbionts from several
Acyrthosi-phon pisum
clones maintained in the Moran laboratory atthe University of Arizona were sequenced directly fromPCR products as described below These aphids were notincluded in the symbiont survey as it was known whichsymbionts they harboured prior to the start of the studyTheir 16S rDNA sequences were deposited in GenBankunder the following accession numbers AY136138 (U-typeof
A pisum
clone 2a from New York) AY136139 (R-type of
A pisum
clone 2BB from Wisconsin) AY136140 (R-type of
A pisum
clone 9-2-1 from New York) AY136141 (T-typeof
A pisum
clone 8-2b from New York)
DNA extractions
A modified protocol of Bender
et al
(1983) was used toextract DNA from American and Israeli aphids First asingle aphid was placed in a 15-mL tube which wasdipped into liquid nitrogen The aphid was then crushedwith a plastic pestle Next 100ndash200
micro
L of lsquolysis bufferrsquo[01
m
NaCl 02
m
sucrose 01
m
TrisndashHCl (pH 91) 005
m
ethylenediaminetetraacetic acid (EDTA) and 05 sodiumdodecyl sulphate] was added to each tube and thecontents were further homogenized Samples wereincubated at 65
deg
C for 30 min then 8
m
potassium acetatewas added to give a final concentration of 1
m
Sampleswere kept on ice for a minimum of 30 min thencentrifuged at 16435
g
for 15 min The supernatant wasdecanted into a new 15-mL tube and an equal volume of100 ethanol was added After 5 min at room temperature
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S
1063
copy 2003 Blackwell Publishing Ltd
Molecular Ecology
12 1061ndash1075
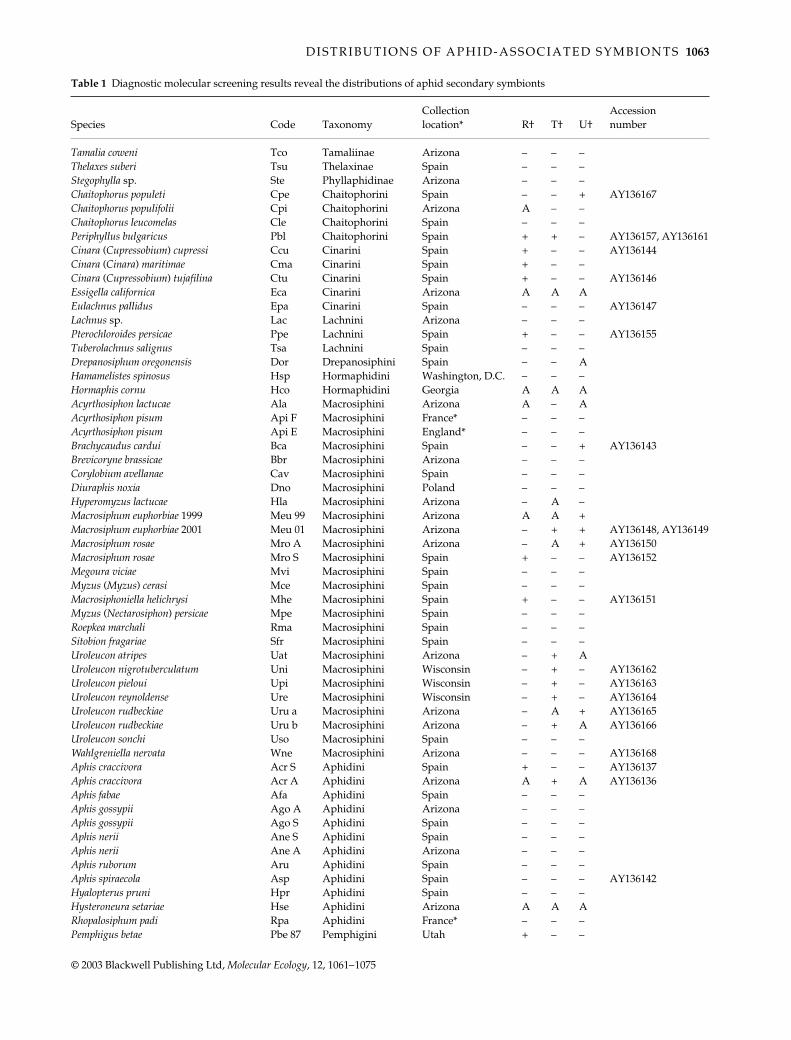
Table 1
Diagnostic molecular screening results reveal the distributions of aphid secondary symbionts
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
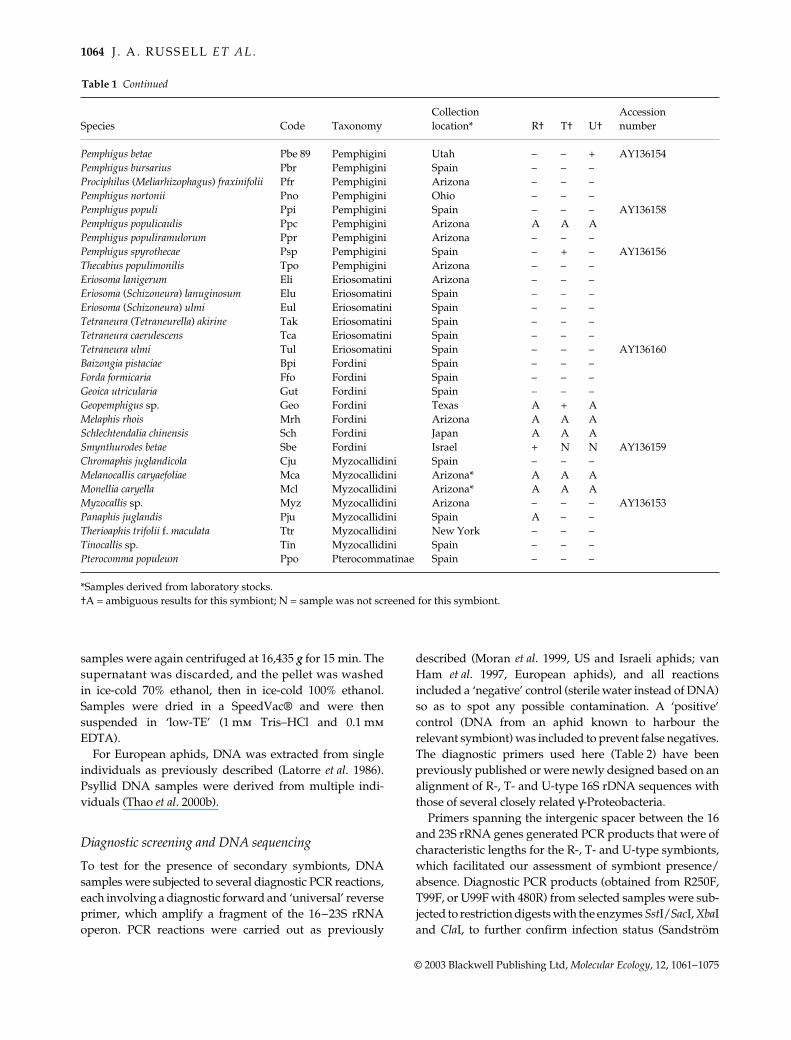
Pemphigus betae Pbe 89 Pemphigini Utah ndash ndash + AY136154Pemphigus bursarius Pbr Pemphigini Spain ndash ndash ndashProciphilus (Meliarhizophagus) fraxinifolii Pfr Pemphigini Arizona ndash ndash ndashPemphigus nortonii Pno Pemphigini Ohio ndash ndash ndashPemphigus populi Ppi Pemphigini Spain ndash ndash ndash AY136158Pemphigus populicaulis Ppc Pemphigini Arizona A A APemphigus populiramulorum Ppr Pemphigini Arizona ndash ndash ndashPemphigus spyrothecae Psp Pemphigini Spain ndash + ndash AY136156Thecabius populimonilis Tpo Pemphigini Arizona ndash ndash ndashEriosoma lanigerum Eli Eriosomatini Arizona ndash ndash ndashEriosoma (Schizoneura) lanuginosum Elu Eriosomatini Spain ndash ndash ndashEriosoma (Schizoneura) ulmi Eul Eriosomatini Spain ndash ndash ndashTetraneura (Tetraneurella) akirine Tak Eriosomatini Spain ndash ndash ndashTetraneura caerulescens Tca Eriosomatini Spain ndash ndash ndashTetraneura ulmi Tul Eriosomatini Spain ndash ndash ndash AY136160Baizongia pistaciae Bpi Fordini Spain ndash ndash ndashForda formicaria Ffo Fordini Spain ndash ndash ndashGeoica utricularia Gut Fordini Spain ndash ndash ndashGeopemphigus sp Geo Fordini Texas A + AMelaphis rhois Mrh Fordini Arizona A A ASchlechtendalia chinensis Sch Fordini Japan A A ASmynthurodes betae Sbe Fordini Israel + N N AY136159Chromaphis juglandicola Cju Myzocallidini Spain ndash ndash ndashMelanocallis caryaefoliae Mca Myzocallidini Arizona A A AMonellia caryella Mcl Myzocallidini Arizona A A AMyzocallis sp Myz Myzocallidini Arizona ndash ndash ndash AY136153Panaphis juglandis Pju Myzocallidini Spain A ndash ndashTherioaphis trifolii f maculata Ttr Myzocallidini New York ndash ndash ndashTinocallis sp Tin Myzocallidini Spain ndash ndash ndashPterocomma populeum Ppo Pterocommatinae Spain ndash ndash ndash
Samples derived from laboratory stocksdaggerA = ambiguous results for this symbiont N = sample was not screened for this symbiont
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
Accession number
Table 1 Continued
samples were again centrifuged at 16435 g for 15 min Thesupernatant was discarded and the pellet was washedin ice-cold 70 ethanol then in ice-cold 100 ethanolSamples were dried in a SpeedVacreg and were thensuspended in lsquolow-TErsquo (1 mm TrisndashHCl and 01 mmEDTA)
For European aphids DNA was extracted from singleindividuals as previously described (Latorre et al 1986)Psyllid DNA samples were derived from multiple indi-viduals (Thao et al 2000b)
Diagnostic screening and DNA sequencing
To test for the presence of secondary symbionts DNAsamples were subjected to several diagnostic PCR reactionseach involving a diagnostic forward and lsquouniversalrsquo reverseprimer which amplify a fragment of the 16ndash23S rRNAoperon PCR reactions were carried out as previously
described (Moran et al 1999 US and Israeli aphids vanHam et al 1997 European aphids) and all reactionsincluded a lsquonegativersquo control (sterile water instead of DNA)so as to spot any possible contamination A lsquopositiversquocontrol (DNA from an aphid known to harbour therelevant symbiont) was included to prevent false negativesThe diagnostic primers used here (Table 2) have beenpreviously published or were newly designed based on analignment of R- T- and U-type 16S rDNA sequences withthose of several closely related γ-Proteobacteria
Primers spanning the intergenic spacer between the 16and 23S rRNA genes generated PCR products that were ofcharacteristic lengths for the R- T- and U-type symbiontswhich facilitated our assessment of symbiont presenceabsence Diagnostic PCR products (obtained from R250FT99F or U99F with 480R) from selected samples were sub-jected to restriction digests with the enzymes SstISacI XbaIand ClaI to further confirm infection status (Sandstroumlm
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1065
et al 2001) All PCR and restriction digest products wererun on agarose gels stained in ethidium bromide and visu-alized with UV light
PCR products spanning portions of the 16S and 23SrRNA genes were obtained for samples that amplifiedin diagnostic screening reactions These products werepurified and sequenced directly (Sandstroumlm et al 2001American aphids) or after cloning (Marchuk et al 1992Martinez-Torres et al 2001 European aphids) All sequen-cing was performed at the University of Arizona GATCSequencing Facility Primers used in the sequencing reac-tions are described in Table 2
Assessment of diagnostic methods
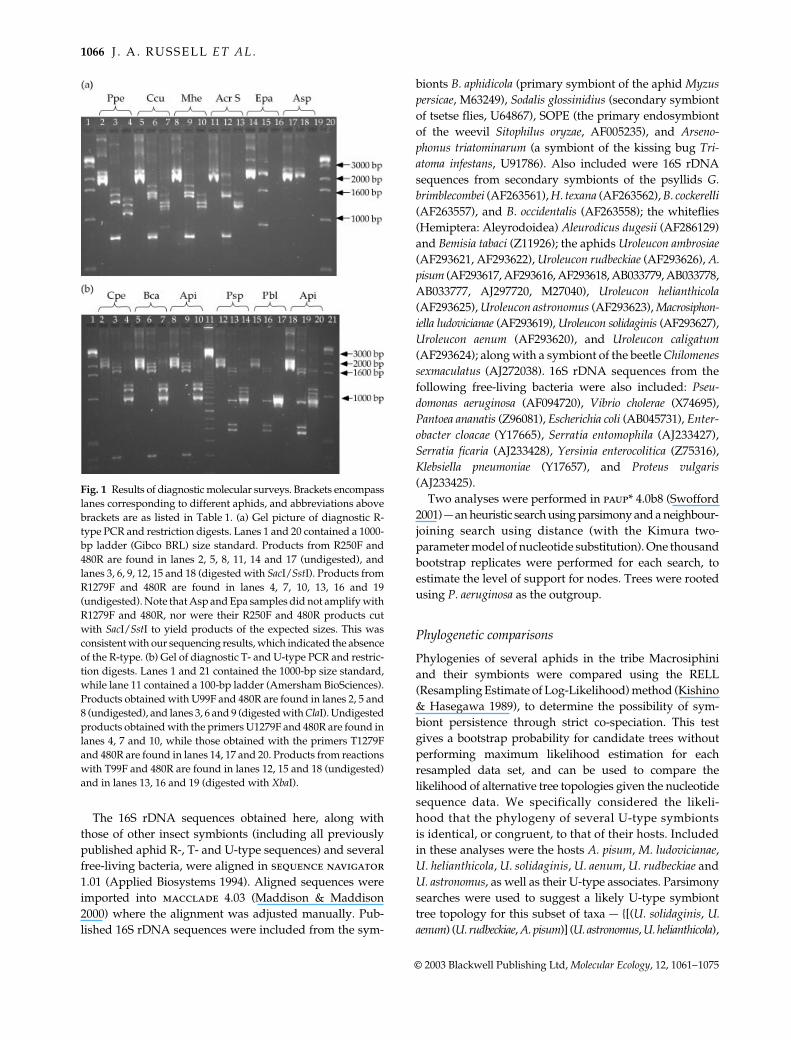
Diagnostic PCR reactions gave amplification in a numberof aphid and psyllid species However not all amplifiedproducts were identical in length to those of positive R- T-and U-type controls from A pisum or to samples in whichthese bacteria were diagnosed through DNA sequencing(Fig 1) In particular the primer R250F exhibited severalcases of nonspecific amplification which resulted inthe discovery of 16S rDNA sequences similar to those ofseveral free-living bacteria (these were considered likelycontaminants) along with symbionts of other insectsSequencing and blastn searches revealed that the com-bination of diagnostic PCR and restriction digest reactionswas sufficient to identify symbionts as no samples testedpositive with both of these techniques yet lacked a 16SrDNA sequence more than 95 similar to previouslydescribed R- T- or U-type symbionts (data not shown)Thus samples were scored as positive (Table 1) only if theytested positive with both of these methods or if associated
16S rDNA sequences were at least 95 similar to apreviously described R- T- or U-type symbiont Similarlywe will refer to all bacteria discovered here as R- T- or U-type symbionts only if they are at least 95 similar to allpreviously described members of these clades as revealedby blastn searches We use this 95 cut-off criterion as allsequences that fell into a monophyletic group with the R-T- or U-types were less than 5 divergent from all othersequences within their respective clades
In contrast samples were scored as negative if they didnot amplify or yielded improperly sized products in tworelevant diagnostic PCR reactions (eg no amplificationwith R250F and 480R or with R1279F and 480R indicated asample that was negative for the R-type) Alternativelysamples were declared negative if the partial 16S rDNAsequence of a diagnostic PCR product was less than 95similar to previously described secondary symbionts Toavoid potential misdiagnosis the few samples not meetingeach of the criteria required to be declared negative or pos-itive were considered ambiguous
16S rDNA phylogeny
Overlapping contigs from sequencing reactions wereassembled using SEQMANII (DNASTAR 1997) Sequenceswere then compared to those in GenBank using blastnsearches Sequences at least 95 similar to all previouslydescribed R- T- or U-type 16S rDNA sequences wereassigned to the relevant symbiont group The results of blastnsearches were also used to choose other symbiotic and free-living bacteria for inclusion in subsequent phylogeneticanalyses Completed sequences were submitted to GenBankand their accession numbers are presented in Table 1
Primer name Primer sequence (5prime rarr 3prime)
Gene and positiondagger UsesDagger
R1279F CGAGAGCAAGCGGACCTCAC 16S 1267ndash1286 DPT1279F CGAGGGAAAGCGGAACTCAG 16S 1267ndash1286 DPU1279F CGAACGTAAGCGAACCTCAT 16S 1267ndash1286 DPR250Fsect GGTAGGTGGGGTAACGGCTC 16S 250ndash269 DP AS ST99Fsectpara AGTGAGCGCAGTTTACTGAG 16S 75ndash87 DP AS SU99Fsect ATCGGGGAGTAGCTTGCTAC 16S 71ndash90 DP AS S480Rsect CACGGTACTGGTTCACTATCGGTC 23S 453ndash476 DP AS1502R GTTACGACTTCACCCCAG 16S 1484ndash1502 S10Fsect AGTTTGATCATGGCTCAGATTG 16S 10ndash31 S AS559F CGTGCCAGCAGCCGCGGTAATAC 16S 514ndash536 S
Names of diagnostic primers begin with an lsquoRrsquo lsquoTrsquo or lsquoUrsquo depending on the symbiont they were designed fordaggerPositions of primers within the 16S and 23S rRNA genes are with respect to the sequences of Escherichia coli (GenBank accession numbers AE000460 and AE000453 respectively)DaggerDP = diagnostic PCR S = sequencing AS = amplification for products to be sequencedsectPreviously described in Sandstroumlm et al (2001)paraPrimer spans a six base pair insertion unique to the T-type
Table 2 Primer sequences their positionsin 16S or 23S rRNA genes and their uses inthis study
The 16S rDNA sequences obtained here along withthose of other insect symbionts (including all previouslypublished aphid R- T- and U-type sequences) and severalfree-living bacteria were aligned in sequence navigator101 (Applied Biosystems 1994) Aligned sequences wereimported into macclade 403 (Maddison amp Maddison2000) where the alignment was adjusted manually Pub-lished 16S rDNA sequences were included from the sym-
bionts B aphidicola (primary symbiont of the aphid Myzuspersicae M63249) Sodalis glossinidius (secondary symbiontof tsetse flies U64867) SOPE (the primary endosymbiontof the weevil Sitophilus oryzae AF005235) and Arseno-phonus triatominarum (a symbiont of the kissing bug Tri-atoma infestans U91786) Also included were 16S rDNAsequences from secondary symbionts of the psyllids Gbrimblecombei (AF263561) H texana (AF263562) B cockerelli(AF263557) and B occidentalis (AF263558) the whiteflies(Hemiptera Aleyrodoidea) Aleurodicus dugesii (AF286129)and Bemisia tabaci (Z11926) the aphids Uroleucon ambrosiae(AF293621 AF293622) Uroleucon rudbeckiae (AF293626) Apisum (AF293617 AF293616 AF293618 AB033779 AB033778AB033777 AJ297720 M27040) Uroleucon helianthicola(AF293625) Uroleucon astronomus (AF293623) Macrosiphon-iella ludovicianae (AF293619) Uroleucon solidaginis (AF293627)Uroleucon aenum (AF293620) and Uroleucon caligatum(AF293624) along with a symbiont of the beetle Chilomenessexmaculatus (AJ272038) 16S rDNA sequences from thefollowing free-living bacteria were also included Pseu-domonas aeruginosa (AF094720) Vibrio cholerae (X74695)Pantoea ananatis (Z96081) Escherichia coli (AB045731) Enter-obacter cloacae (Y17665) Serratia entomophila (AJ233427)Serratia ficaria (AJ233428) Yersinia enterocolitica (Z75316)Klebsiella pneumoniae (Y17657) and Proteus vulgaris(AJ233425)
Two analyses were performed in paup 40b8 (Swofford2001) mdash an heuristic search using parsimony and a neighbour-joining search using distance (with the Kimura two-parameter model of nucleotide substitution) One thousandbootstrap replicates were performed for each search toestimate the level of support for nodes Trees were rootedusing P aeruginosa as the outgroup
Phylogenetic comparisons
Phylogenies of several aphids in the tribe Macrosiphiniand their symbionts were compared using the RELL(Resampling Estimate of Log-Likelihood) method (Kishinoamp Hasegawa 1989) to determine the possibility of sym-biont persistence through strict co-speciation This testgives a bootstrap probability for candidate trees withoutperforming maximum likelihood estimation for eachresampled data set and can be used to compare thelikelihood of alternative tree topologies given the nucleotidesequence data We specifically considered the likeli-hood that the phylogeny of several U-type symbiontsis identical or congruent to that of their hosts Includedin these analyses were the hosts A pisum M ludovicianaeU helianthicola U solidaginis U aenum U rudbeckiae andU astronomus as well as their U-type associates Parsimonysearches were used to suggest a likely U-type symbionttree topology for this subset of taxa mdash [(U solidaginis Uaenum) (U rudbeckiae A pisum)] (U astronomus U helianthicola)
Fig 1 Results of diagnostic molecular surveys Brackets encompasslanes corresponding to different aphids and abbreviations abovebrackets are as listed in Table 1 (a) Gel picture of diagnostic R-type PCR and restriction digests Lanes 1 and 20 contained a 1000-bp ladder (Gibco BRL) size standard Products from R250F and480R are found in lanes 2 5 8 11 14 and 17 (undigested) andlanes 3 6 9 12 15 and 18 (digested with SacISstI) Products fromR1279F and 480R are found in lanes 4 7 10 13 16 and 19(undigested) Note that Asp and Epa samples did not amplify withR1279F and 480R nor were their R250F and 480R products cutwith SacISstI to yield products of the expected sizes This wasconsistent with our sequencing results which indicated the absenceof the R-type (b) Gel of diagnostic T- and U-type PCR and restric-tion digests Lanes 1 and 21 contained the 1000-bp size standardwhile lane 11 contained a 100-bp ladder (Amersham BioSciences)Products obtained with U99F and 480R are found in lanes 2 5 and8 (undigested) and lanes 3 6 and 9 (digested with ClaI) Undigestedproducts obtained with the primers U1279F and 480R are found inlanes 4 7 and 10 while those obtained with the primers T1279Fand 480R are found in lanes 14 17 and 20 Products from reactionswith T99F and 480R are found in lanes 12 15 and 18 (undigested)and in lanes 13 16 and 19 (digested with XbaI)
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
1062
J A R U S S E L L
E T A L
copy 2003 Blackwell Publishing Ltd
Molecular Ecology
12 1061ndash1075
Fukatsu 2001 Sandstroumlm
et al
2001) and the Mollicutes(Fukatsu
et al
2001) Close relatives of aphid secondarieswith 16S rDNA sequence identities ranging from 98 toover 99 have been found in distantly related arthropodspecies (Chen
et al
1996 Fukatsu
et al
2001 Sandstroumlm
et al
2001) Thus for the few hosts that have been exam-ined the distributions of several secondary symbiontsappear to be partly the result of horizontal transmission(ie movement between hosts other than that from parentto offspring) between host species However some evid-ence suggests that certain associations between aphidsand secondary symbionts are specialized and have co-evolved (Fukatsu 2001)
To date most DNA sequence-based studies of aphid sec-ondary symbionts have focused on hosts within the tribeMacrosiphini Moreover although these studies providedsome evidence for horizontal transfer (Chen amp Purcell1997 Darby
et al
2001 Sandstroumlm
et al
2001) few haveemployed statistical analyses to demonstrate this pointand little effort has been made to assess the extent to whichhorizontal transfer has shaped secondary symbiont dis-tributions Here we use diagnostic molecular screeningtechniques and DNA sequencing along with pairwisedivergence and phylogenetic analyses to explore the dis-tributions of three facultative
γ
-Proteobacterial secondarysymbionts of aphids known as the R-type (or S-sym orPea Aphid Secondary Symbiont mdash PASS) T-type (or PeaAphid
Bemisia
-like Symbiont mdash PABS) and U-type (Chenamp Purcell 1997 Unterman
et al
1989 Darby
et al
2001Sandstroumlm
et al
2001) across a variety of aphid and psyllid(Hemiptera Psylloidea) species Specifically we comparethe phylogenies of hosts and symbionts to determine whe-ther there is any evidence for co-speciation Also we com-pare 16S rDNA divergence values between primary andsecondary symbionts residing within pairs of hosts sharingthe same most recent common ancestor Differences inthese values are used to determine the likelihood that cur-rent secondary infections could possibly be the result ofexclusive vertical transmission since the time of this com-mon host ancestor We also compare the rates of substitu-tion in the 16S rRNA genes of R- T- and U-type symbiontsto those of free-living bacteria and
B aphidicola
and esti-mate the ages of symbiont common ancestors through theuse of a molecular clock Finally we use data on symbiontprevalence within and across host taxa along with obser-vations of symbiont-associated fitness effects to assess thepotential importance of these microbes in aphid biology
Materials and methods
Samples
Members of 75 species spanning 15 subfamilies and tribesacross the Aphidoidea (Remaudiegravere amp Remaudiegravere 1997)
were collected from a variety of locations in the UnitedStates and Eurasia In addition members of four aphidspecies were obtained from laboratory stocks A list of allaphid species involved in the survey their taxonomicaffiliations and information regarding their collection ispresented in Table 1
Representatives of 27 psyllid species from North andSouth America along with Europe (see Thao
et al
2000b forcollection information) were also surveyed as psyllidshave been shown to harbor a number of secondary sym-bionts (Thao
et al
2000a) yet have not been screened forthe R- T- or U-types The samples included were
Acizziauncatoides
(two samples)
Aphalaroida inermis
Bactericeracockerelli
Blastopsylla occidentalis
Boreioglycaspis melaleucae
Cacopsylla brunneipennis
Cacopsylla myrthi
Cacopsylla pere-grina
Cacopsylla pyri
Calophya schini
Ctenarytaina eucalypti
Ctenarytaina longicauda
Ctenarytaina spatulata
Diaphorinacitri
Glycaspis brimblecombei
Heteropsylla cubana
Heterop-sylla texana
Neotriozella hirsuta
Psylla buxi
Trioza urticae
Pachypsylla celtidismamma
Pachypsylla venusta
Panisopelma
sp
Psylla
sp
Russelliana intermedia
Spanioneura fonscolombii
and
Trioza eugeniae
To determine the phylogenetic proximity of related sym-
bionts found within the same host species the 16S rRNAgenes of R- T- or U-type symbionts from several
Acyrthosi-phon pisum
clones maintained in the Moran laboratory atthe University of Arizona were sequenced directly fromPCR products as described below These aphids were notincluded in the symbiont survey as it was known whichsymbionts they harboured prior to the start of the studyTheir 16S rDNA sequences were deposited in GenBankunder the following accession numbers AY136138 (U-typeof
A pisum
clone 2a from New York) AY136139 (R-type of
A pisum
clone 2BB from Wisconsin) AY136140 (R-type of
A pisum
clone 9-2-1 from New York) AY136141 (T-typeof
A pisum
clone 8-2b from New York)
DNA extractions
A modified protocol of Bender
et al
(1983) was used toextract DNA from American and Israeli aphids First asingle aphid was placed in a 15-mL tube which wasdipped into liquid nitrogen The aphid was then crushedwith a plastic pestle Next 100ndash200
micro
L of lsquolysis bufferrsquo[01
m
NaCl 02
m
sucrose 01
m
TrisndashHCl (pH 91) 005
m
ethylenediaminetetraacetic acid (EDTA) and 05 sodiumdodecyl sulphate] was added to each tube and thecontents were further homogenized Samples wereincubated at 65
deg
C for 30 min then 8
m
potassium acetatewas added to give a final concentration of 1
m
Sampleswere kept on ice for a minimum of 30 min thencentrifuged at 16435
g
for 15 min The supernatant wasdecanted into a new 15-mL tube and an equal volume of100 ethanol was added After 5 min at room temperature
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S
1063
copy 2003 Blackwell Publishing Ltd
Molecular Ecology
12 1061ndash1075
Table 1
Diagnostic molecular screening results reveal the distributions of aphid secondary symbionts
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
Pemphigus betae Pbe 89 Pemphigini Utah ndash ndash + AY136154Pemphigus bursarius Pbr Pemphigini Spain ndash ndash ndashProciphilus (Meliarhizophagus) fraxinifolii Pfr Pemphigini Arizona ndash ndash ndashPemphigus nortonii Pno Pemphigini Ohio ndash ndash ndashPemphigus populi Ppi Pemphigini Spain ndash ndash ndash AY136158Pemphigus populicaulis Ppc Pemphigini Arizona A A APemphigus populiramulorum Ppr Pemphigini Arizona ndash ndash ndashPemphigus spyrothecae Psp Pemphigini Spain ndash + ndash AY136156Thecabius populimonilis Tpo Pemphigini Arizona ndash ndash ndashEriosoma lanigerum Eli Eriosomatini Arizona ndash ndash ndashEriosoma (Schizoneura) lanuginosum Elu Eriosomatini Spain ndash ndash ndashEriosoma (Schizoneura) ulmi Eul Eriosomatini Spain ndash ndash ndashTetraneura (Tetraneurella) akirine Tak Eriosomatini Spain ndash ndash ndashTetraneura caerulescens Tca Eriosomatini Spain ndash ndash ndashTetraneura ulmi Tul Eriosomatini Spain ndash ndash ndash AY136160Baizongia pistaciae Bpi Fordini Spain ndash ndash ndashForda formicaria Ffo Fordini Spain ndash ndash ndashGeoica utricularia Gut Fordini Spain ndash ndash ndashGeopemphigus sp Geo Fordini Texas A + AMelaphis rhois Mrh Fordini Arizona A A ASchlechtendalia chinensis Sch Fordini Japan A A ASmynthurodes betae Sbe Fordini Israel + N N AY136159Chromaphis juglandicola Cju Myzocallidini Spain ndash ndash ndashMelanocallis caryaefoliae Mca Myzocallidini Arizona A A AMonellia caryella Mcl Myzocallidini Arizona A A AMyzocallis sp Myz Myzocallidini Arizona ndash ndash ndash AY136153Panaphis juglandis Pju Myzocallidini Spain A ndash ndashTherioaphis trifolii f maculata Ttr Myzocallidini New York ndash ndash ndashTinocallis sp Tin Myzocallidini Spain ndash ndash ndashPterocomma populeum Ppo Pterocommatinae Spain ndash ndash ndash
Samples derived from laboratory stocksdaggerA = ambiguous results for this symbiont N = sample was not screened for this symbiont
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
Accession number
Table 1 Continued
samples were again centrifuged at 16435 g for 15 min Thesupernatant was discarded and the pellet was washedin ice-cold 70 ethanol then in ice-cold 100 ethanolSamples were dried in a SpeedVacreg and were thensuspended in lsquolow-TErsquo (1 mm TrisndashHCl and 01 mmEDTA)
For European aphids DNA was extracted from singleindividuals as previously described (Latorre et al 1986)Psyllid DNA samples were derived from multiple indi-viduals (Thao et al 2000b)
Diagnostic screening and DNA sequencing
To test for the presence of secondary symbionts DNAsamples were subjected to several diagnostic PCR reactionseach involving a diagnostic forward and lsquouniversalrsquo reverseprimer which amplify a fragment of the 16ndash23S rRNAoperon PCR reactions were carried out as previously
described (Moran et al 1999 US and Israeli aphids vanHam et al 1997 European aphids) and all reactionsincluded a lsquonegativersquo control (sterile water instead of DNA)so as to spot any possible contamination A lsquopositiversquocontrol (DNA from an aphid known to harbour therelevant symbiont) was included to prevent false negativesThe diagnostic primers used here (Table 2) have beenpreviously published or were newly designed based on analignment of R- T- and U-type 16S rDNA sequences withthose of several closely related γ-Proteobacteria
Primers spanning the intergenic spacer between the 16and 23S rRNA genes generated PCR products that were ofcharacteristic lengths for the R- T- and U-type symbiontswhich facilitated our assessment of symbiont presenceabsence Diagnostic PCR products (obtained from R250FT99F or U99F with 480R) from selected samples were sub-jected to restriction digests with the enzymes SstISacI XbaIand ClaI to further confirm infection status (Sandstroumlm
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1065
et al 2001) All PCR and restriction digest products wererun on agarose gels stained in ethidium bromide and visu-alized with UV light
PCR products spanning portions of the 16S and 23SrRNA genes were obtained for samples that amplifiedin diagnostic screening reactions These products werepurified and sequenced directly (Sandstroumlm et al 2001American aphids) or after cloning (Marchuk et al 1992Martinez-Torres et al 2001 European aphids) All sequen-cing was performed at the University of Arizona GATCSequencing Facility Primers used in the sequencing reac-tions are described in Table 2
Assessment of diagnostic methods
Diagnostic PCR reactions gave amplification in a numberof aphid and psyllid species However not all amplifiedproducts were identical in length to those of positive R- T-and U-type controls from A pisum or to samples in whichthese bacteria were diagnosed through DNA sequencing(Fig 1) In particular the primer R250F exhibited severalcases of nonspecific amplification which resulted inthe discovery of 16S rDNA sequences similar to those ofseveral free-living bacteria (these were considered likelycontaminants) along with symbionts of other insectsSequencing and blastn searches revealed that the com-bination of diagnostic PCR and restriction digest reactionswas sufficient to identify symbionts as no samples testedpositive with both of these techniques yet lacked a 16SrDNA sequence more than 95 similar to previouslydescribed R- T- or U-type symbionts (data not shown)Thus samples were scored as positive (Table 1) only if theytested positive with both of these methods or if associated
16S rDNA sequences were at least 95 similar to apreviously described R- T- or U-type symbiont Similarlywe will refer to all bacteria discovered here as R- T- or U-type symbionts only if they are at least 95 similar to allpreviously described members of these clades as revealedby blastn searches We use this 95 cut-off criterion as allsequences that fell into a monophyletic group with the R-T- or U-types were less than 5 divergent from all othersequences within their respective clades
In contrast samples were scored as negative if they didnot amplify or yielded improperly sized products in tworelevant diagnostic PCR reactions (eg no amplificationwith R250F and 480R or with R1279F and 480R indicated asample that was negative for the R-type) Alternativelysamples were declared negative if the partial 16S rDNAsequence of a diagnostic PCR product was less than 95similar to previously described secondary symbionts Toavoid potential misdiagnosis the few samples not meetingeach of the criteria required to be declared negative or pos-itive were considered ambiguous
16S rDNA phylogeny
Overlapping contigs from sequencing reactions wereassembled using SEQMANII (DNASTAR 1997) Sequenceswere then compared to those in GenBank using blastnsearches Sequences at least 95 similar to all previouslydescribed R- T- or U-type 16S rDNA sequences wereassigned to the relevant symbiont group The results of blastnsearches were also used to choose other symbiotic and free-living bacteria for inclusion in subsequent phylogeneticanalyses Completed sequences were submitted to GenBankand their accession numbers are presented in Table 1
Primer name Primer sequence (5prime rarr 3prime)
Gene and positiondagger UsesDagger
R1279F CGAGAGCAAGCGGACCTCAC 16S 1267ndash1286 DPT1279F CGAGGGAAAGCGGAACTCAG 16S 1267ndash1286 DPU1279F CGAACGTAAGCGAACCTCAT 16S 1267ndash1286 DPR250Fsect GGTAGGTGGGGTAACGGCTC 16S 250ndash269 DP AS ST99Fsectpara AGTGAGCGCAGTTTACTGAG 16S 75ndash87 DP AS SU99Fsect ATCGGGGAGTAGCTTGCTAC 16S 71ndash90 DP AS S480Rsect CACGGTACTGGTTCACTATCGGTC 23S 453ndash476 DP AS1502R GTTACGACTTCACCCCAG 16S 1484ndash1502 S10Fsect AGTTTGATCATGGCTCAGATTG 16S 10ndash31 S AS559F CGTGCCAGCAGCCGCGGTAATAC 16S 514ndash536 S
Names of diagnostic primers begin with an lsquoRrsquo lsquoTrsquo or lsquoUrsquo depending on the symbiont they were designed fordaggerPositions of primers within the 16S and 23S rRNA genes are with respect to the sequences of Escherichia coli (GenBank accession numbers AE000460 and AE000453 respectively)DaggerDP = diagnostic PCR S = sequencing AS = amplification for products to be sequencedsectPreviously described in Sandstroumlm et al (2001)paraPrimer spans a six base pair insertion unique to the T-type
Table 2 Primer sequences their positionsin 16S or 23S rRNA genes and their uses inthis study
The 16S rDNA sequences obtained here along withthose of other insect symbionts (including all previouslypublished aphid R- T- and U-type sequences) and severalfree-living bacteria were aligned in sequence navigator101 (Applied Biosystems 1994) Aligned sequences wereimported into macclade 403 (Maddison amp Maddison2000) where the alignment was adjusted manually Pub-lished 16S rDNA sequences were included from the sym-
bionts B aphidicola (primary symbiont of the aphid Myzuspersicae M63249) Sodalis glossinidius (secondary symbiontof tsetse flies U64867) SOPE (the primary endosymbiontof the weevil Sitophilus oryzae AF005235) and Arseno-phonus triatominarum (a symbiont of the kissing bug Tri-atoma infestans U91786) Also included were 16S rDNAsequences from secondary symbionts of the psyllids Gbrimblecombei (AF263561) H texana (AF263562) B cockerelli(AF263557) and B occidentalis (AF263558) the whiteflies(Hemiptera Aleyrodoidea) Aleurodicus dugesii (AF286129)and Bemisia tabaci (Z11926) the aphids Uroleucon ambrosiae(AF293621 AF293622) Uroleucon rudbeckiae (AF293626) Apisum (AF293617 AF293616 AF293618 AB033779 AB033778AB033777 AJ297720 M27040) Uroleucon helianthicola(AF293625) Uroleucon astronomus (AF293623) Macrosiphon-iella ludovicianae (AF293619) Uroleucon solidaginis (AF293627)Uroleucon aenum (AF293620) and Uroleucon caligatum(AF293624) along with a symbiont of the beetle Chilomenessexmaculatus (AJ272038) 16S rDNA sequences from thefollowing free-living bacteria were also included Pseu-domonas aeruginosa (AF094720) Vibrio cholerae (X74695)Pantoea ananatis (Z96081) Escherichia coli (AB045731) Enter-obacter cloacae (Y17665) Serratia entomophila (AJ233427)Serratia ficaria (AJ233428) Yersinia enterocolitica (Z75316)Klebsiella pneumoniae (Y17657) and Proteus vulgaris(AJ233425)
Two analyses were performed in paup 40b8 (Swofford2001) mdash an heuristic search using parsimony and a neighbour-joining search using distance (with the Kimura two-parameter model of nucleotide substitution) One thousandbootstrap replicates were performed for each search toestimate the level of support for nodes Trees were rootedusing P aeruginosa as the outgroup
Phylogenetic comparisons
Phylogenies of several aphids in the tribe Macrosiphiniand their symbionts were compared using the RELL(Resampling Estimate of Log-Likelihood) method (Kishinoamp Hasegawa 1989) to determine the possibility of sym-biont persistence through strict co-speciation This testgives a bootstrap probability for candidate trees withoutperforming maximum likelihood estimation for eachresampled data set and can be used to compare thelikelihood of alternative tree topologies given the nucleotidesequence data We specifically considered the likeli-hood that the phylogeny of several U-type symbiontsis identical or congruent to that of their hosts Includedin these analyses were the hosts A pisum M ludovicianaeU helianthicola U solidaginis U aenum U rudbeckiae andU astronomus as well as their U-type associates Parsimonysearches were used to suggest a likely U-type symbionttree topology for this subset of taxa mdash [(U solidaginis Uaenum) (U rudbeckiae A pisum)] (U astronomus U helianthicola)
Fig 1 Results of diagnostic molecular surveys Brackets encompasslanes corresponding to different aphids and abbreviations abovebrackets are as listed in Table 1 (a) Gel picture of diagnostic R-type PCR and restriction digests Lanes 1 and 20 contained a 1000-bp ladder (Gibco BRL) size standard Products from R250F and480R are found in lanes 2 5 8 11 14 and 17 (undigested) andlanes 3 6 9 12 15 and 18 (digested with SacISstI) Products fromR1279F and 480R are found in lanes 4 7 10 13 16 and 19(undigested) Note that Asp and Epa samples did not amplify withR1279F and 480R nor were their R250F and 480R products cutwith SacISstI to yield products of the expected sizes This wasconsistent with our sequencing results which indicated the absenceof the R-type (b) Gel of diagnostic T- and U-type PCR and restric-tion digests Lanes 1 and 21 contained the 1000-bp size standardwhile lane 11 contained a 100-bp ladder (Amersham BioSciences)Products obtained with U99F and 480R are found in lanes 2 5 and8 (undigested) and lanes 3 6 and 9 (digested with ClaI) Undigestedproducts obtained with the primers U1279F and 480R are found inlanes 4 7 and 10 while those obtained with the primers T1279Fand 480R are found in lanes 14 17 and 20 Products from reactionswith T99F and 480R are found in lanes 12 15 and 18 (undigested)and in lanes 13 16 and 19 (digested with XbaI)
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S
1063
copy 2003 Blackwell Publishing Ltd
Molecular Ecology
12 1061ndash1075
Table 1
Diagnostic molecular screening results reveal the distributions of aphid secondary symbionts
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
Pemphigus betae Pbe 89 Pemphigini Utah ndash ndash + AY136154Pemphigus bursarius Pbr Pemphigini Spain ndash ndash ndashProciphilus (Meliarhizophagus) fraxinifolii Pfr Pemphigini Arizona ndash ndash ndashPemphigus nortonii Pno Pemphigini Ohio ndash ndash ndashPemphigus populi Ppi Pemphigini Spain ndash ndash ndash AY136158Pemphigus populicaulis Ppc Pemphigini Arizona A A APemphigus populiramulorum Ppr Pemphigini Arizona ndash ndash ndashPemphigus spyrothecae Psp Pemphigini Spain ndash + ndash AY136156Thecabius populimonilis Tpo Pemphigini Arizona ndash ndash ndashEriosoma lanigerum Eli Eriosomatini Arizona ndash ndash ndashEriosoma (Schizoneura) lanuginosum Elu Eriosomatini Spain ndash ndash ndashEriosoma (Schizoneura) ulmi Eul Eriosomatini Spain ndash ndash ndashTetraneura (Tetraneurella) akirine Tak Eriosomatini Spain ndash ndash ndashTetraneura caerulescens Tca Eriosomatini Spain ndash ndash ndashTetraneura ulmi Tul Eriosomatini Spain ndash ndash ndash AY136160Baizongia pistaciae Bpi Fordini Spain ndash ndash ndashForda formicaria Ffo Fordini Spain ndash ndash ndashGeoica utricularia Gut Fordini Spain ndash ndash ndashGeopemphigus sp Geo Fordini Texas A + AMelaphis rhois Mrh Fordini Arizona A A ASchlechtendalia chinensis Sch Fordini Japan A A ASmynthurodes betae Sbe Fordini Israel + N N AY136159Chromaphis juglandicola Cju Myzocallidini Spain ndash ndash ndashMelanocallis caryaefoliae Mca Myzocallidini Arizona A A AMonellia caryella Mcl Myzocallidini Arizona A A AMyzocallis sp Myz Myzocallidini Arizona ndash ndash ndash AY136153Panaphis juglandis Pju Myzocallidini Spain A ndash ndashTherioaphis trifolii f maculata Ttr Myzocallidini New York ndash ndash ndashTinocallis sp Tin Myzocallidini Spain ndash ndash ndashPterocomma populeum Ppo Pterocommatinae Spain ndash ndash ndash
Samples derived from laboratory stocksdaggerA = ambiguous results for this symbiont N = sample was not screened for this symbiont
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
Accession number
Table 1 Continued
samples were again centrifuged at 16435 g for 15 min Thesupernatant was discarded and the pellet was washedin ice-cold 70 ethanol then in ice-cold 100 ethanolSamples were dried in a SpeedVacreg and were thensuspended in lsquolow-TErsquo (1 mm TrisndashHCl and 01 mmEDTA)
For European aphids DNA was extracted from singleindividuals as previously described (Latorre et al 1986)Psyllid DNA samples were derived from multiple indi-viduals (Thao et al 2000b)
Diagnostic screening and DNA sequencing
To test for the presence of secondary symbionts DNAsamples were subjected to several diagnostic PCR reactionseach involving a diagnostic forward and lsquouniversalrsquo reverseprimer which amplify a fragment of the 16ndash23S rRNAoperon PCR reactions were carried out as previously
described (Moran et al 1999 US and Israeli aphids vanHam et al 1997 European aphids) and all reactionsincluded a lsquonegativersquo control (sterile water instead of DNA)so as to spot any possible contamination A lsquopositiversquocontrol (DNA from an aphid known to harbour therelevant symbiont) was included to prevent false negativesThe diagnostic primers used here (Table 2) have beenpreviously published or were newly designed based on analignment of R- T- and U-type 16S rDNA sequences withthose of several closely related γ-Proteobacteria
Primers spanning the intergenic spacer between the 16and 23S rRNA genes generated PCR products that were ofcharacteristic lengths for the R- T- and U-type symbiontswhich facilitated our assessment of symbiont presenceabsence Diagnostic PCR products (obtained from R250FT99F or U99F with 480R) from selected samples were sub-jected to restriction digests with the enzymes SstISacI XbaIand ClaI to further confirm infection status (Sandstroumlm
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1065
et al 2001) All PCR and restriction digest products wererun on agarose gels stained in ethidium bromide and visu-alized with UV light
PCR products spanning portions of the 16S and 23SrRNA genes were obtained for samples that amplifiedin diagnostic screening reactions These products werepurified and sequenced directly (Sandstroumlm et al 2001American aphids) or after cloning (Marchuk et al 1992Martinez-Torres et al 2001 European aphids) All sequen-cing was performed at the University of Arizona GATCSequencing Facility Primers used in the sequencing reac-tions are described in Table 2
Assessment of diagnostic methods
Diagnostic PCR reactions gave amplification in a numberof aphid and psyllid species However not all amplifiedproducts were identical in length to those of positive R- T-and U-type controls from A pisum or to samples in whichthese bacteria were diagnosed through DNA sequencing(Fig 1) In particular the primer R250F exhibited severalcases of nonspecific amplification which resulted inthe discovery of 16S rDNA sequences similar to those ofseveral free-living bacteria (these were considered likelycontaminants) along with symbionts of other insectsSequencing and blastn searches revealed that the com-bination of diagnostic PCR and restriction digest reactionswas sufficient to identify symbionts as no samples testedpositive with both of these techniques yet lacked a 16SrDNA sequence more than 95 similar to previouslydescribed R- T- or U-type symbionts (data not shown)Thus samples were scored as positive (Table 1) only if theytested positive with both of these methods or if associated
16S rDNA sequences were at least 95 similar to apreviously described R- T- or U-type symbiont Similarlywe will refer to all bacteria discovered here as R- T- or U-type symbionts only if they are at least 95 similar to allpreviously described members of these clades as revealedby blastn searches We use this 95 cut-off criterion as allsequences that fell into a monophyletic group with the R-T- or U-types were less than 5 divergent from all othersequences within their respective clades
In contrast samples were scored as negative if they didnot amplify or yielded improperly sized products in tworelevant diagnostic PCR reactions (eg no amplificationwith R250F and 480R or with R1279F and 480R indicated asample that was negative for the R-type) Alternativelysamples were declared negative if the partial 16S rDNAsequence of a diagnostic PCR product was less than 95similar to previously described secondary symbionts Toavoid potential misdiagnosis the few samples not meetingeach of the criteria required to be declared negative or pos-itive were considered ambiguous
16S rDNA phylogeny
Overlapping contigs from sequencing reactions wereassembled using SEQMANII (DNASTAR 1997) Sequenceswere then compared to those in GenBank using blastnsearches Sequences at least 95 similar to all previouslydescribed R- T- or U-type 16S rDNA sequences wereassigned to the relevant symbiont group The results of blastnsearches were also used to choose other symbiotic and free-living bacteria for inclusion in subsequent phylogeneticanalyses Completed sequences were submitted to GenBankand their accession numbers are presented in Table 1
Primer name Primer sequence (5prime rarr 3prime)
Gene and positiondagger UsesDagger
R1279F CGAGAGCAAGCGGACCTCAC 16S 1267ndash1286 DPT1279F CGAGGGAAAGCGGAACTCAG 16S 1267ndash1286 DPU1279F CGAACGTAAGCGAACCTCAT 16S 1267ndash1286 DPR250Fsect GGTAGGTGGGGTAACGGCTC 16S 250ndash269 DP AS ST99Fsectpara AGTGAGCGCAGTTTACTGAG 16S 75ndash87 DP AS SU99Fsect ATCGGGGAGTAGCTTGCTAC 16S 71ndash90 DP AS S480Rsect CACGGTACTGGTTCACTATCGGTC 23S 453ndash476 DP AS1502R GTTACGACTTCACCCCAG 16S 1484ndash1502 S10Fsect AGTTTGATCATGGCTCAGATTG 16S 10ndash31 S AS559F CGTGCCAGCAGCCGCGGTAATAC 16S 514ndash536 S
Names of diagnostic primers begin with an lsquoRrsquo lsquoTrsquo or lsquoUrsquo depending on the symbiont they were designed fordaggerPositions of primers within the 16S and 23S rRNA genes are with respect to the sequences of Escherichia coli (GenBank accession numbers AE000460 and AE000453 respectively)DaggerDP = diagnostic PCR S = sequencing AS = amplification for products to be sequencedsectPreviously described in Sandstroumlm et al (2001)paraPrimer spans a six base pair insertion unique to the T-type
Table 2 Primer sequences their positionsin 16S or 23S rRNA genes and their uses inthis study
The 16S rDNA sequences obtained here along withthose of other insect symbionts (including all previouslypublished aphid R- T- and U-type sequences) and severalfree-living bacteria were aligned in sequence navigator101 (Applied Biosystems 1994) Aligned sequences wereimported into macclade 403 (Maddison amp Maddison2000) where the alignment was adjusted manually Pub-lished 16S rDNA sequences were included from the sym-
bionts B aphidicola (primary symbiont of the aphid Myzuspersicae M63249) Sodalis glossinidius (secondary symbiontof tsetse flies U64867) SOPE (the primary endosymbiontof the weevil Sitophilus oryzae AF005235) and Arseno-phonus triatominarum (a symbiont of the kissing bug Tri-atoma infestans U91786) Also included were 16S rDNAsequences from secondary symbionts of the psyllids Gbrimblecombei (AF263561) H texana (AF263562) B cockerelli(AF263557) and B occidentalis (AF263558) the whiteflies(Hemiptera Aleyrodoidea) Aleurodicus dugesii (AF286129)and Bemisia tabaci (Z11926) the aphids Uroleucon ambrosiae(AF293621 AF293622) Uroleucon rudbeckiae (AF293626) Apisum (AF293617 AF293616 AF293618 AB033779 AB033778AB033777 AJ297720 M27040) Uroleucon helianthicola(AF293625) Uroleucon astronomus (AF293623) Macrosiphon-iella ludovicianae (AF293619) Uroleucon solidaginis (AF293627)Uroleucon aenum (AF293620) and Uroleucon caligatum(AF293624) along with a symbiont of the beetle Chilomenessexmaculatus (AJ272038) 16S rDNA sequences from thefollowing free-living bacteria were also included Pseu-domonas aeruginosa (AF094720) Vibrio cholerae (X74695)Pantoea ananatis (Z96081) Escherichia coli (AB045731) Enter-obacter cloacae (Y17665) Serratia entomophila (AJ233427)Serratia ficaria (AJ233428) Yersinia enterocolitica (Z75316)Klebsiella pneumoniae (Y17657) and Proteus vulgaris(AJ233425)
Two analyses were performed in paup 40b8 (Swofford2001) mdash an heuristic search using parsimony and a neighbour-joining search using distance (with the Kimura two-parameter model of nucleotide substitution) One thousandbootstrap replicates were performed for each search toestimate the level of support for nodes Trees were rootedusing P aeruginosa as the outgroup
Phylogenetic comparisons
Phylogenies of several aphids in the tribe Macrosiphiniand their symbionts were compared using the RELL(Resampling Estimate of Log-Likelihood) method (Kishinoamp Hasegawa 1989) to determine the possibility of sym-biont persistence through strict co-speciation This testgives a bootstrap probability for candidate trees withoutperforming maximum likelihood estimation for eachresampled data set and can be used to compare thelikelihood of alternative tree topologies given the nucleotidesequence data We specifically considered the likeli-hood that the phylogeny of several U-type symbiontsis identical or congruent to that of their hosts Includedin these analyses were the hosts A pisum M ludovicianaeU helianthicola U solidaginis U aenum U rudbeckiae andU astronomus as well as their U-type associates Parsimonysearches were used to suggest a likely U-type symbionttree topology for this subset of taxa mdash [(U solidaginis Uaenum) (U rudbeckiae A pisum)] (U astronomus U helianthicola)
Fig 1 Results of diagnostic molecular surveys Brackets encompasslanes corresponding to different aphids and abbreviations abovebrackets are as listed in Table 1 (a) Gel picture of diagnostic R-type PCR and restriction digests Lanes 1 and 20 contained a 1000-bp ladder (Gibco BRL) size standard Products from R250F and480R are found in lanes 2 5 8 11 14 and 17 (undigested) andlanes 3 6 9 12 15 and 18 (digested with SacISstI) Products fromR1279F and 480R are found in lanes 4 7 10 13 16 and 19(undigested) Note that Asp and Epa samples did not amplify withR1279F and 480R nor were their R250F and 480R products cutwith SacISstI to yield products of the expected sizes This wasconsistent with our sequencing results which indicated the absenceof the R-type (b) Gel of diagnostic T- and U-type PCR and restric-tion digests Lanes 1 and 21 contained the 1000-bp size standardwhile lane 11 contained a 100-bp ladder (Amersham BioSciences)Products obtained with U99F and 480R are found in lanes 2 5 and8 (undigested) and lanes 3 6 and 9 (digested with ClaI) Undigestedproducts obtained with the primers U1279F and 480R are found inlanes 4 7 and 10 while those obtained with the primers T1279Fand 480R are found in lanes 14 17 and 20 Products from reactionswith T99F and 480R are found in lanes 12 15 and 18 (undigested)and in lanes 13 16 and 19 (digested with XbaI)
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
Pemphigus betae Pbe 89 Pemphigini Utah ndash ndash + AY136154Pemphigus bursarius Pbr Pemphigini Spain ndash ndash ndashProciphilus (Meliarhizophagus) fraxinifolii Pfr Pemphigini Arizona ndash ndash ndashPemphigus nortonii Pno Pemphigini Ohio ndash ndash ndashPemphigus populi Ppi Pemphigini Spain ndash ndash ndash AY136158Pemphigus populicaulis Ppc Pemphigini Arizona A A APemphigus populiramulorum Ppr Pemphigini Arizona ndash ndash ndashPemphigus spyrothecae Psp Pemphigini Spain ndash + ndash AY136156Thecabius populimonilis Tpo Pemphigini Arizona ndash ndash ndashEriosoma lanigerum Eli Eriosomatini Arizona ndash ndash ndashEriosoma (Schizoneura) lanuginosum Elu Eriosomatini Spain ndash ndash ndashEriosoma (Schizoneura) ulmi Eul Eriosomatini Spain ndash ndash ndashTetraneura (Tetraneurella) akirine Tak Eriosomatini Spain ndash ndash ndashTetraneura caerulescens Tca Eriosomatini Spain ndash ndash ndashTetraneura ulmi Tul Eriosomatini Spain ndash ndash ndash AY136160Baizongia pistaciae Bpi Fordini Spain ndash ndash ndashForda formicaria Ffo Fordini Spain ndash ndash ndashGeoica utricularia Gut Fordini Spain ndash ndash ndashGeopemphigus sp Geo Fordini Texas A + AMelaphis rhois Mrh Fordini Arizona A A ASchlechtendalia chinensis Sch Fordini Japan A A ASmynthurodes betae Sbe Fordini Israel + N N AY136159Chromaphis juglandicola Cju Myzocallidini Spain ndash ndash ndashMelanocallis caryaefoliae Mca Myzocallidini Arizona A A AMonellia caryella Mcl Myzocallidini Arizona A A AMyzocallis sp Myz Myzocallidini Arizona ndash ndash ndash AY136153Panaphis juglandis Pju Myzocallidini Spain A ndash ndashTherioaphis trifolii f maculata Ttr Myzocallidini New York ndash ndash ndashTinocallis sp Tin Myzocallidini Spain ndash ndash ndashPterocomma populeum Ppo Pterocommatinae Spain ndash ndash ndash
Samples derived from laboratory stocksdaggerA = ambiguous results for this symbiont N = sample was not screened for this symbiont
Species Code TaxonomyCollection location Rdagger Tdagger Udagger
Accession number
Table 1 Continued
samples were again centrifuged at 16435 g for 15 min Thesupernatant was discarded and the pellet was washedin ice-cold 70 ethanol then in ice-cold 100 ethanolSamples were dried in a SpeedVacreg and were thensuspended in lsquolow-TErsquo (1 mm TrisndashHCl and 01 mmEDTA)
For European aphids DNA was extracted from singleindividuals as previously described (Latorre et al 1986)Psyllid DNA samples were derived from multiple indi-viduals (Thao et al 2000b)
Diagnostic screening and DNA sequencing
To test for the presence of secondary symbionts DNAsamples were subjected to several diagnostic PCR reactionseach involving a diagnostic forward and lsquouniversalrsquo reverseprimer which amplify a fragment of the 16ndash23S rRNAoperon PCR reactions were carried out as previously
described (Moran et al 1999 US and Israeli aphids vanHam et al 1997 European aphids) and all reactionsincluded a lsquonegativersquo control (sterile water instead of DNA)so as to spot any possible contamination A lsquopositiversquocontrol (DNA from an aphid known to harbour therelevant symbiont) was included to prevent false negativesThe diagnostic primers used here (Table 2) have beenpreviously published or were newly designed based on analignment of R- T- and U-type 16S rDNA sequences withthose of several closely related γ-Proteobacteria
Primers spanning the intergenic spacer between the 16and 23S rRNA genes generated PCR products that were ofcharacteristic lengths for the R- T- and U-type symbiontswhich facilitated our assessment of symbiont presenceabsence Diagnostic PCR products (obtained from R250FT99F or U99F with 480R) from selected samples were sub-jected to restriction digests with the enzymes SstISacI XbaIand ClaI to further confirm infection status (Sandstroumlm
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1065
et al 2001) All PCR and restriction digest products wererun on agarose gels stained in ethidium bromide and visu-alized with UV light
PCR products spanning portions of the 16S and 23SrRNA genes were obtained for samples that amplifiedin diagnostic screening reactions These products werepurified and sequenced directly (Sandstroumlm et al 2001American aphids) or after cloning (Marchuk et al 1992Martinez-Torres et al 2001 European aphids) All sequen-cing was performed at the University of Arizona GATCSequencing Facility Primers used in the sequencing reac-tions are described in Table 2
Assessment of diagnostic methods
Diagnostic PCR reactions gave amplification in a numberof aphid and psyllid species However not all amplifiedproducts were identical in length to those of positive R- T-and U-type controls from A pisum or to samples in whichthese bacteria were diagnosed through DNA sequencing(Fig 1) In particular the primer R250F exhibited severalcases of nonspecific amplification which resulted inthe discovery of 16S rDNA sequences similar to those ofseveral free-living bacteria (these were considered likelycontaminants) along with symbionts of other insectsSequencing and blastn searches revealed that the com-bination of diagnostic PCR and restriction digest reactionswas sufficient to identify symbionts as no samples testedpositive with both of these techniques yet lacked a 16SrDNA sequence more than 95 similar to previouslydescribed R- T- or U-type symbionts (data not shown)Thus samples were scored as positive (Table 1) only if theytested positive with both of these methods or if associated
16S rDNA sequences were at least 95 similar to apreviously described R- T- or U-type symbiont Similarlywe will refer to all bacteria discovered here as R- T- or U-type symbionts only if they are at least 95 similar to allpreviously described members of these clades as revealedby blastn searches We use this 95 cut-off criterion as allsequences that fell into a monophyletic group with the R-T- or U-types were less than 5 divergent from all othersequences within their respective clades
In contrast samples were scored as negative if they didnot amplify or yielded improperly sized products in tworelevant diagnostic PCR reactions (eg no amplificationwith R250F and 480R or with R1279F and 480R indicated asample that was negative for the R-type) Alternativelysamples were declared negative if the partial 16S rDNAsequence of a diagnostic PCR product was less than 95similar to previously described secondary symbionts Toavoid potential misdiagnosis the few samples not meetingeach of the criteria required to be declared negative or pos-itive were considered ambiguous
16S rDNA phylogeny
Overlapping contigs from sequencing reactions wereassembled using SEQMANII (DNASTAR 1997) Sequenceswere then compared to those in GenBank using blastnsearches Sequences at least 95 similar to all previouslydescribed R- T- or U-type 16S rDNA sequences wereassigned to the relevant symbiont group The results of blastnsearches were also used to choose other symbiotic and free-living bacteria for inclusion in subsequent phylogeneticanalyses Completed sequences were submitted to GenBankand their accession numbers are presented in Table 1
Primer name Primer sequence (5prime rarr 3prime)
Gene and positiondagger UsesDagger
R1279F CGAGAGCAAGCGGACCTCAC 16S 1267ndash1286 DPT1279F CGAGGGAAAGCGGAACTCAG 16S 1267ndash1286 DPU1279F CGAACGTAAGCGAACCTCAT 16S 1267ndash1286 DPR250Fsect GGTAGGTGGGGTAACGGCTC 16S 250ndash269 DP AS ST99Fsectpara AGTGAGCGCAGTTTACTGAG 16S 75ndash87 DP AS SU99Fsect ATCGGGGAGTAGCTTGCTAC 16S 71ndash90 DP AS S480Rsect CACGGTACTGGTTCACTATCGGTC 23S 453ndash476 DP AS1502R GTTACGACTTCACCCCAG 16S 1484ndash1502 S10Fsect AGTTTGATCATGGCTCAGATTG 16S 10ndash31 S AS559F CGTGCCAGCAGCCGCGGTAATAC 16S 514ndash536 S
Names of diagnostic primers begin with an lsquoRrsquo lsquoTrsquo or lsquoUrsquo depending on the symbiont they were designed fordaggerPositions of primers within the 16S and 23S rRNA genes are with respect to the sequences of Escherichia coli (GenBank accession numbers AE000460 and AE000453 respectively)DaggerDP = diagnostic PCR S = sequencing AS = amplification for products to be sequencedsectPreviously described in Sandstroumlm et al (2001)paraPrimer spans a six base pair insertion unique to the T-type
Table 2 Primer sequences their positionsin 16S or 23S rRNA genes and their uses inthis study
The 16S rDNA sequences obtained here along withthose of other insect symbionts (including all previouslypublished aphid R- T- and U-type sequences) and severalfree-living bacteria were aligned in sequence navigator101 (Applied Biosystems 1994) Aligned sequences wereimported into macclade 403 (Maddison amp Maddison2000) where the alignment was adjusted manually Pub-lished 16S rDNA sequences were included from the sym-
bionts B aphidicola (primary symbiont of the aphid Myzuspersicae M63249) Sodalis glossinidius (secondary symbiontof tsetse flies U64867) SOPE (the primary endosymbiontof the weevil Sitophilus oryzae AF005235) and Arseno-phonus triatominarum (a symbiont of the kissing bug Tri-atoma infestans U91786) Also included were 16S rDNAsequences from secondary symbionts of the psyllids Gbrimblecombei (AF263561) H texana (AF263562) B cockerelli(AF263557) and B occidentalis (AF263558) the whiteflies(Hemiptera Aleyrodoidea) Aleurodicus dugesii (AF286129)and Bemisia tabaci (Z11926) the aphids Uroleucon ambrosiae(AF293621 AF293622) Uroleucon rudbeckiae (AF293626) Apisum (AF293617 AF293616 AF293618 AB033779 AB033778AB033777 AJ297720 M27040) Uroleucon helianthicola(AF293625) Uroleucon astronomus (AF293623) Macrosiphon-iella ludovicianae (AF293619) Uroleucon solidaginis (AF293627)Uroleucon aenum (AF293620) and Uroleucon caligatum(AF293624) along with a symbiont of the beetle Chilomenessexmaculatus (AJ272038) 16S rDNA sequences from thefollowing free-living bacteria were also included Pseu-domonas aeruginosa (AF094720) Vibrio cholerae (X74695)Pantoea ananatis (Z96081) Escherichia coli (AB045731) Enter-obacter cloacae (Y17665) Serratia entomophila (AJ233427)Serratia ficaria (AJ233428) Yersinia enterocolitica (Z75316)Klebsiella pneumoniae (Y17657) and Proteus vulgaris(AJ233425)
Two analyses were performed in paup 40b8 (Swofford2001) mdash an heuristic search using parsimony and a neighbour-joining search using distance (with the Kimura two-parameter model of nucleotide substitution) One thousandbootstrap replicates were performed for each search toestimate the level of support for nodes Trees were rootedusing P aeruginosa as the outgroup
Phylogenetic comparisons
Phylogenies of several aphids in the tribe Macrosiphiniand their symbionts were compared using the RELL(Resampling Estimate of Log-Likelihood) method (Kishinoamp Hasegawa 1989) to determine the possibility of sym-biont persistence through strict co-speciation This testgives a bootstrap probability for candidate trees withoutperforming maximum likelihood estimation for eachresampled data set and can be used to compare thelikelihood of alternative tree topologies given the nucleotidesequence data We specifically considered the likeli-hood that the phylogeny of several U-type symbiontsis identical or congruent to that of their hosts Includedin these analyses were the hosts A pisum M ludovicianaeU helianthicola U solidaginis U aenum U rudbeckiae andU astronomus as well as their U-type associates Parsimonysearches were used to suggest a likely U-type symbionttree topology for this subset of taxa mdash [(U solidaginis Uaenum) (U rudbeckiae A pisum)] (U astronomus U helianthicola)
Fig 1 Results of diagnostic molecular surveys Brackets encompasslanes corresponding to different aphids and abbreviations abovebrackets are as listed in Table 1 (a) Gel picture of diagnostic R-type PCR and restriction digests Lanes 1 and 20 contained a 1000-bp ladder (Gibco BRL) size standard Products from R250F and480R are found in lanes 2 5 8 11 14 and 17 (undigested) andlanes 3 6 9 12 15 and 18 (digested with SacISstI) Products fromR1279F and 480R are found in lanes 4 7 10 13 16 and 19(undigested) Note that Asp and Epa samples did not amplify withR1279F and 480R nor were their R250F and 480R products cutwith SacISstI to yield products of the expected sizes This wasconsistent with our sequencing results which indicated the absenceof the R-type (b) Gel of diagnostic T- and U-type PCR and restric-tion digests Lanes 1 and 21 contained the 1000-bp size standardwhile lane 11 contained a 100-bp ladder (Amersham BioSciences)Products obtained with U99F and 480R are found in lanes 2 5 and8 (undigested) and lanes 3 6 and 9 (digested with ClaI) Undigestedproducts obtained with the primers U1279F and 480R are found inlanes 4 7 and 10 while those obtained with the primers T1279Fand 480R are found in lanes 14 17 and 20 Products from reactionswith T99F and 480R are found in lanes 12 15 and 18 (undigested)and in lanes 13 16 and 19 (digested with XbaI)
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1065
et al 2001) All PCR and restriction digest products wererun on agarose gels stained in ethidium bromide and visu-alized with UV light
PCR products spanning portions of the 16S and 23SrRNA genes were obtained for samples that amplifiedin diagnostic screening reactions These products werepurified and sequenced directly (Sandstroumlm et al 2001American aphids) or after cloning (Marchuk et al 1992Martinez-Torres et al 2001 European aphids) All sequen-cing was performed at the University of Arizona GATCSequencing Facility Primers used in the sequencing reac-tions are described in Table 2
Assessment of diagnostic methods
Diagnostic PCR reactions gave amplification in a numberof aphid and psyllid species However not all amplifiedproducts were identical in length to those of positive R- T-and U-type controls from A pisum or to samples in whichthese bacteria were diagnosed through DNA sequencing(Fig 1) In particular the primer R250F exhibited severalcases of nonspecific amplification which resulted inthe discovery of 16S rDNA sequences similar to those ofseveral free-living bacteria (these were considered likelycontaminants) along with symbionts of other insectsSequencing and blastn searches revealed that the com-bination of diagnostic PCR and restriction digest reactionswas sufficient to identify symbionts as no samples testedpositive with both of these techniques yet lacked a 16SrDNA sequence more than 95 similar to previouslydescribed R- T- or U-type symbionts (data not shown)Thus samples were scored as positive (Table 1) only if theytested positive with both of these methods or if associated
16S rDNA sequences were at least 95 similar to apreviously described R- T- or U-type symbiont Similarlywe will refer to all bacteria discovered here as R- T- or U-type symbionts only if they are at least 95 similar to allpreviously described members of these clades as revealedby blastn searches We use this 95 cut-off criterion as allsequences that fell into a monophyletic group with the R-T- or U-types were less than 5 divergent from all othersequences within their respective clades
In contrast samples were scored as negative if they didnot amplify or yielded improperly sized products in tworelevant diagnostic PCR reactions (eg no amplificationwith R250F and 480R or with R1279F and 480R indicated asample that was negative for the R-type) Alternativelysamples were declared negative if the partial 16S rDNAsequence of a diagnostic PCR product was less than 95similar to previously described secondary symbionts Toavoid potential misdiagnosis the few samples not meetingeach of the criteria required to be declared negative or pos-itive were considered ambiguous
16S rDNA phylogeny
Overlapping contigs from sequencing reactions wereassembled using SEQMANII (DNASTAR 1997) Sequenceswere then compared to those in GenBank using blastnsearches Sequences at least 95 similar to all previouslydescribed R- T- or U-type 16S rDNA sequences wereassigned to the relevant symbiont group The results of blastnsearches were also used to choose other symbiotic and free-living bacteria for inclusion in subsequent phylogeneticanalyses Completed sequences were submitted to GenBankand their accession numbers are presented in Table 1
Primer name Primer sequence (5prime rarr 3prime)
Gene and positiondagger UsesDagger
R1279F CGAGAGCAAGCGGACCTCAC 16S 1267ndash1286 DPT1279F CGAGGGAAAGCGGAACTCAG 16S 1267ndash1286 DPU1279F CGAACGTAAGCGAACCTCAT 16S 1267ndash1286 DPR250Fsect GGTAGGTGGGGTAACGGCTC 16S 250ndash269 DP AS ST99Fsectpara AGTGAGCGCAGTTTACTGAG 16S 75ndash87 DP AS SU99Fsect ATCGGGGAGTAGCTTGCTAC 16S 71ndash90 DP AS S480Rsect CACGGTACTGGTTCACTATCGGTC 23S 453ndash476 DP AS1502R GTTACGACTTCACCCCAG 16S 1484ndash1502 S10Fsect AGTTTGATCATGGCTCAGATTG 16S 10ndash31 S AS559F CGTGCCAGCAGCCGCGGTAATAC 16S 514ndash536 S
Names of diagnostic primers begin with an lsquoRrsquo lsquoTrsquo or lsquoUrsquo depending on the symbiont they were designed fordaggerPositions of primers within the 16S and 23S rRNA genes are with respect to the sequences of Escherichia coli (GenBank accession numbers AE000460 and AE000453 respectively)DaggerDP = diagnostic PCR S = sequencing AS = amplification for products to be sequencedsectPreviously described in Sandstroumlm et al (2001)paraPrimer spans a six base pair insertion unique to the T-type
Table 2 Primer sequences their positionsin 16S or 23S rRNA genes and their uses inthis study
The 16S rDNA sequences obtained here along withthose of other insect symbionts (including all previouslypublished aphid R- T- and U-type sequences) and severalfree-living bacteria were aligned in sequence navigator101 (Applied Biosystems 1994) Aligned sequences wereimported into macclade 403 (Maddison amp Maddison2000) where the alignment was adjusted manually Pub-lished 16S rDNA sequences were included from the sym-
bionts B aphidicola (primary symbiont of the aphid Myzuspersicae M63249) Sodalis glossinidius (secondary symbiontof tsetse flies U64867) SOPE (the primary endosymbiontof the weevil Sitophilus oryzae AF005235) and Arseno-phonus triatominarum (a symbiont of the kissing bug Tri-atoma infestans U91786) Also included were 16S rDNAsequences from secondary symbionts of the psyllids Gbrimblecombei (AF263561) H texana (AF263562) B cockerelli(AF263557) and B occidentalis (AF263558) the whiteflies(Hemiptera Aleyrodoidea) Aleurodicus dugesii (AF286129)and Bemisia tabaci (Z11926) the aphids Uroleucon ambrosiae(AF293621 AF293622) Uroleucon rudbeckiae (AF293626) Apisum (AF293617 AF293616 AF293618 AB033779 AB033778AB033777 AJ297720 M27040) Uroleucon helianthicola(AF293625) Uroleucon astronomus (AF293623) Macrosiphon-iella ludovicianae (AF293619) Uroleucon solidaginis (AF293627)Uroleucon aenum (AF293620) and Uroleucon caligatum(AF293624) along with a symbiont of the beetle Chilomenessexmaculatus (AJ272038) 16S rDNA sequences from thefollowing free-living bacteria were also included Pseu-domonas aeruginosa (AF094720) Vibrio cholerae (X74695)Pantoea ananatis (Z96081) Escherichia coli (AB045731) Enter-obacter cloacae (Y17665) Serratia entomophila (AJ233427)Serratia ficaria (AJ233428) Yersinia enterocolitica (Z75316)Klebsiella pneumoniae (Y17657) and Proteus vulgaris(AJ233425)
Two analyses were performed in paup 40b8 (Swofford2001) mdash an heuristic search using parsimony and a neighbour-joining search using distance (with the Kimura two-parameter model of nucleotide substitution) One thousandbootstrap replicates were performed for each search toestimate the level of support for nodes Trees were rootedusing P aeruginosa as the outgroup
Phylogenetic comparisons
Phylogenies of several aphids in the tribe Macrosiphiniand their symbionts were compared using the RELL(Resampling Estimate of Log-Likelihood) method (Kishinoamp Hasegawa 1989) to determine the possibility of sym-biont persistence through strict co-speciation This testgives a bootstrap probability for candidate trees withoutperforming maximum likelihood estimation for eachresampled data set and can be used to compare thelikelihood of alternative tree topologies given the nucleotidesequence data We specifically considered the likeli-hood that the phylogeny of several U-type symbiontsis identical or congruent to that of their hosts Includedin these analyses were the hosts A pisum M ludovicianaeU helianthicola U solidaginis U aenum U rudbeckiae andU astronomus as well as their U-type associates Parsimonysearches were used to suggest a likely U-type symbionttree topology for this subset of taxa mdash [(U solidaginis Uaenum) (U rudbeckiae A pisum)] (U astronomus U helianthicola)
Fig 1 Results of diagnostic molecular surveys Brackets encompasslanes corresponding to different aphids and abbreviations abovebrackets are as listed in Table 1 (a) Gel picture of diagnostic R-type PCR and restriction digests Lanes 1 and 20 contained a 1000-bp ladder (Gibco BRL) size standard Products from R250F and480R are found in lanes 2 5 8 11 14 and 17 (undigested) andlanes 3 6 9 12 15 and 18 (digested with SacISstI) Products fromR1279F and 480R are found in lanes 4 7 10 13 16 and 19(undigested) Note that Asp and Epa samples did not amplify withR1279F and 480R nor were their R250F and 480R products cutwith SacISstI to yield products of the expected sizes This wasconsistent with our sequencing results which indicated the absenceof the R-type (b) Gel of diagnostic T- and U-type PCR and restric-tion digests Lanes 1 and 21 contained the 1000-bp size standardwhile lane 11 contained a 100-bp ladder (Amersham BioSciences)Products obtained with U99F and 480R are found in lanes 2 5 and8 (undigested) and lanes 3 6 and 9 (digested with ClaI) Undigestedproducts obtained with the primers U1279F and 480R are found inlanes 4 7 and 10 while those obtained with the primers T1279Fand 480R are found in lanes 14 17 and 20 Products from reactionswith T99F and 480R are found in lanes 12 15 and 18 (undigested)and in lanes 13 16 and 19 (digested with XbaI)
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
The 16S rDNA sequences obtained here along withthose of other insect symbionts (including all previouslypublished aphid R- T- and U-type sequences) and severalfree-living bacteria were aligned in sequence navigator101 (Applied Biosystems 1994) Aligned sequences wereimported into macclade 403 (Maddison amp Maddison2000) where the alignment was adjusted manually Pub-lished 16S rDNA sequences were included from the sym-
bionts B aphidicola (primary symbiont of the aphid Myzuspersicae M63249) Sodalis glossinidius (secondary symbiontof tsetse flies U64867) SOPE (the primary endosymbiontof the weevil Sitophilus oryzae AF005235) and Arseno-phonus triatominarum (a symbiont of the kissing bug Tri-atoma infestans U91786) Also included were 16S rDNAsequences from secondary symbionts of the psyllids Gbrimblecombei (AF263561) H texana (AF263562) B cockerelli(AF263557) and B occidentalis (AF263558) the whiteflies(Hemiptera Aleyrodoidea) Aleurodicus dugesii (AF286129)and Bemisia tabaci (Z11926) the aphids Uroleucon ambrosiae(AF293621 AF293622) Uroleucon rudbeckiae (AF293626) Apisum (AF293617 AF293616 AF293618 AB033779 AB033778AB033777 AJ297720 M27040) Uroleucon helianthicola(AF293625) Uroleucon astronomus (AF293623) Macrosiphon-iella ludovicianae (AF293619) Uroleucon solidaginis (AF293627)Uroleucon aenum (AF293620) and Uroleucon caligatum(AF293624) along with a symbiont of the beetle Chilomenessexmaculatus (AJ272038) 16S rDNA sequences from thefollowing free-living bacteria were also included Pseu-domonas aeruginosa (AF094720) Vibrio cholerae (X74695)Pantoea ananatis (Z96081) Escherichia coli (AB045731) Enter-obacter cloacae (Y17665) Serratia entomophila (AJ233427)Serratia ficaria (AJ233428) Yersinia enterocolitica (Z75316)Klebsiella pneumoniae (Y17657) and Proteus vulgaris(AJ233425)
Two analyses were performed in paup 40b8 (Swofford2001) mdash an heuristic search using parsimony and a neighbour-joining search using distance (with the Kimura two-parameter model of nucleotide substitution) One thousandbootstrap replicates were performed for each search toestimate the level of support for nodes Trees were rootedusing P aeruginosa as the outgroup
Phylogenetic comparisons
Phylogenies of several aphids in the tribe Macrosiphiniand their symbionts were compared using the RELL(Resampling Estimate of Log-Likelihood) method (Kishinoamp Hasegawa 1989) to determine the possibility of sym-biont persistence through strict co-speciation This testgives a bootstrap probability for candidate trees withoutperforming maximum likelihood estimation for eachresampled data set and can be used to compare thelikelihood of alternative tree topologies given the nucleotidesequence data We specifically considered the likeli-hood that the phylogeny of several U-type symbiontsis identical or congruent to that of their hosts Includedin these analyses were the hosts A pisum M ludovicianaeU helianthicola U solidaginis U aenum U rudbeckiae andU astronomus as well as their U-type associates Parsimonysearches were used to suggest a likely U-type symbionttree topology for this subset of taxa mdash [(U solidaginis Uaenum) (U rudbeckiae A pisum)] (U astronomus U helianthicola)
Fig 1 Results of diagnostic molecular surveys Brackets encompasslanes corresponding to different aphids and abbreviations abovebrackets are as listed in Table 1 (a) Gel picture of diagnostic R-type PCR and restriction digests Lanes 1 and 20 contained a 1000-bp ladder (Gibco BRL) size standard Products from R250F and480R are found in lanes 2 5 8 11 14 and 17 (undigested) andlanes 3 6 9 12 15 and 18 (digested with SacISstI) Products fromR1279F and 480R are found in lanes 4 7 10 13 16 and 19(undigested) Note that Asp and Epa samples did not amplify withR1279F and 480R nor were their R250F and 480R products cutwith SacISstI to yield products of the expected sizes This wasconsistent with our sequencing results which indicated the absenceof the R-type (b) Gel of diagnostic T- and U-type PCR and restric-tion digests Lanes 1 and 21 contained the 1000-bp size standardwhile lane 11 contained a 100-bp ladder (Amersham BioSciences)Products obtained with U99F and 480R are found in lanes 2 5 and8 (undigested) and lanes 3 6 and 9 (digested with ClaI) Undigestedproducts obtained with the primers U1279F and 480R are found inlanes 4 7 and 10 while those obtained with the primers T1279Fand 480R are found in lanes 14 17 and 20 Products from reactionswith T99F and 480R are found in lanes 12 15 and 18 (undigested)and in lanes 13 16 and 19 (digested with XbaI)
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1067
M ludovicianae All nodes in this tree were supported bybootstrap values of 80 or higher We then compared theapproximate bootstrap probability of this topology tothose of three possible host trees based on mitochon-drial and nuclear gene sequences (from Figs 1c in Moranet al 1999) mdash [(U solidaginis U aenum (U astronomusU rudbeckiae)) U helianthicola] M ludovicianae A pisumAll RELL analyses were executed in molphy 23 (Adachi ampHasegawa 1996)
Relative rates and pairwise divergences
To determine the rates at which symbiont 16S rRNAgenes are evolving relative rates tests were performedThe 16S rDNA sequences included all exceeded 1300 nucleo-tides in length and were derived from the R-typesymbionts of A pisum (six individuals) and U caligatumthe T-type symbionts of Uroleucon nigrotuberculatum Uambrosiae (two individuals) U rudbeckiae (two individuals)Uroleucon pieloui Uroleucon reynoldense Aphis craccivoraA pisum (two individuals) Periphyllus bulgaricus Csexmaculatus and B tabaci the U-type symbionts of Usolidaginis Brachycaudus cardui Macrosiphum euphorbiae (twoindividuals) Macrosiphum rosae M ludovicianae U aenumPemphigus betae A pisum U astronomus and Chaitophoruspopuleti B aphidicola and the free-living bacteria E coli Sentomophila S ficaria and V cholerae This trimmed-downalignment file (otherwise identical to that used for the 16SrDNA phylogeny) was opened in paupb8 where missingand ambiguous sites were removed Vibrio cholerae wasdefined as the outgroup and a guide tree was constructed(using parsimony and 100 bootstrap replicates) forweighting in relative rates tests (Robinson et al 1998) Thebootstrap 60 majority-rule consensus tree used in relativerates tests had the following topology (V cholerae ((((Apisum California R A pisum Japan R A pisum Japan R Apisum Japan R A pisum Wisconsin R A pisum ArizonaR) U caligatum R) (S ficaria S entomophila)) E coli((((U solidaginis U (B cardui U C populeti U) U aenum UP betae U) (M euphorbiae U M euphorbiae U) M rosae U Apisum U U astronomus U) M ludovicianae U) (((Unigrotuberculatum T U ambrosiae Arizona T U reynoldenseT U pieloui T U ambrosiae Minnesota T C pyri T) (Urudbeckiae T U rudbeckiae T A pisum New York T A pisumWisconsin T) P bulgaricus T C sexmaculatus T) Acraccivora T B tabaci T)) Buchnera)) The ungapped andunambiguous 1301 base pair alignment was openedin rrtree (Robinson-Rechavi amp Huchon 2000 httppbiluniv-lyon1frsoftwarerrtreehtml) 16S rRNA ratesof substitution were compared between R-type T-typeU-type B aphidicola E coli and Serratia species withV cholerae defined as the outgroup Divergences werecomputed using the Kimura two-parameter model ofnucleotide substitution and relative rates tests were
performed as described by Robinson et al (1998) Thealignment file created for relative rates tests was alsoused to compute pairwise divergences between the 16SrDNA sequences of free-living and symbiotic bacteriaBoth the absolute number of differences and Kimuratwo-parameter corrected distances were computed usingpaup 40b8
Primary and secondary symbiont pairwise divergence comparisons
Finally pairwise divergence comparison analyses wereperformed in an attempt to help determine whether R-T- or U-type symbionts could have persisted withinhost lineages through strict vertical transmission since thetime of a common host ancestor (ie whether they haveundergone strict co-speciation) If such a scenario hasoccurred genetic divergences between secondaries infectinga pair of host species from two distinct taxa shouldresemble those between B aphidicola from the same taxaSpecifically because B aphidicola undergoes strict verticaltransmission (Munson et al 1991 Rouhbakhsh et al 1996Clark et al 2000 Funk et al 2000) and because bacterial 16SrRNA substitution rates range from 1 to 4 per hundredmillion years (Moran et al 1993 Ochman amp Wilson 1987Ochman et al 1999) differences in divergences (betweenB aphidicola and secondaries in equi-distant hosts) shouldnot exceed four-fold under strict vertical transmission ofsecondary symbionts Violations of these conditions indicatehorizontal transfer however cases in which conditionsare not violated do not imply co-speciation as similardivergences could simply be chance This approach doesnot require knowledge of phylogenetic relatedness andthus provides an alternative approach to examining thepossibility of co-speciation
We constructed an alignment including several R-T- and U-type 16S rDNA sequences from symbionts asso-ciated with hosts from the tribes Chaitophorini Macro-siphini Pemphigini and Aphidini and from the generaAcyrthosiphon and Uroleucon Also included were 16S rDNAsequences of B aphidicola symbionts from species in thesame tribes and genera A pisum (M27039) Diuraphis noxia(M63251) M persicae (M63249) and Uroleucon sonchi (M63250)of the Macrosiphini Chaitophorus viminalis (M63252) of theChaitophorini P betae (M63254) and Pemphigus populi(AJ296750) of the Pemphigini and Rhopalosiphum maidis(M63247) Rhopalosiphum padi (M63248) and Schizaphisgraminum (M63246) of the Aphidini The alignment wasperformed as described above Pairwise distances werethen calculated over 1296 nucleotides (no missing orambiguous sites) using the Kimura two-parameter modelof nucleotide substitution and compared between Baphidicola and secondaries infecting one member each of thesame two host taxa For example the 16S rDNA divergence
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
between B aphidicola of P populi (Pemphigini) and Mpersicae (Macrosiphini) was compared to that betweenthe U-type symbionts of P betae (Pemphigini) and M ludo-vicianae (Macrosiphini) When sequences were availablefor symbionts of more than one host species within a giventaxon the smallest B aphidicola divergence value wascompared to the largest secondary symbiont divergencevalue This minimized the differences in 16S rDNA diver-gences making any conclusions of horizontal transferquite conservative
Results
Symbiont distributions
The results of our diagnostic molecular screening sur-vey are summarized in Table 1 Each of the R- T- and U-type symbionts was found in one or more previouslyunscreened members of the Macrosiphini The R-type wasfound in members of six new aphid tribes mdash the AphidiniChaitophorini Cinarini Fordini Lachnini and Pem-phigini The T-type was found in one psyllid Cacopsyllapyri (GenBank accession no AY136145) and in four newaphid tribes mdash the Aphidini Chaitophorini Fordini andPemphigini Finally the U-type was found in two new aphidtribes mdash the Chaitophorini and Pemphigini In total withinthe Aphidoidea the R-type was found in 1079 surveyedspecies the T-type in 1178 and the U-type in 676Within the Psylloidea 027 surveyed species were foundto associate with the R- or U-types while only 127 psyllidspecies was positive for the T-type Though more aphidand psyllid hosts tested negative than positive for eachsymbiont it should be noted that negative results here donot imply that the symbionts of interest are absent from aspecies as only one to a few individuals were surveyed
These results indicate that R- T- and U-type symbiontsare certainly not confined to members of the Macrosiphinior even to the superfamily Aphidoidea in the case of theT-type Indeed blastn searches using T-type 16S rDNAsequences revealed that such molecules share a highdegree of similarity (over 98) with that of a putativemale-killing bacterium described from the ladybird beetleChilomenes sexmaculatus (MEN Majerus personal commu-nication GenBank accession no AJ272038) Also T-type16S rDNA sequences have previously been shown to shareover 98 identity with that from a secondary symbiont ofthe whitefly Bemisia tabaci (Sandstroumlm et al 2001)
Diagnostic screening revealed two cases of potentiallystable associations between a symbiont type and a hostclade First all three surveyed members of the genusCinara were found to harbour the R-type (Table 1) Alsothree previously unscreened Nearctic members of the sub-genus Uroleucon mdash U pieloui U reynoldense and U nigrotu-berculatum mdash were found to associate with the T-type
(Table 1) adding to previous findings of the T-type in therelated North American species Uroleucon ambrosiae Uro-leucon astronomus and Uroleucon rudbeckiae (Sandstroumlm et al2001)
Phylogenetic analysis
The 16S rDNA phylogeny of aphid- and psyllid-associatedbacteria described here plus their symbiotic and free-living relatives is presented in Fig 2 Several bacterialsequences obtained from aphids (and one from a psyllid)fell into one of three strongly supported clades termedR [parsimony (BS) = 72] T (parsimony BS = 100) and U(parsimony BS = 100) Within these clades relationshipswere often unresolved because of high genetic similarityAs previously demonstrated (Sandstroumlm et al 2001) the T-and U-types were sister clades (parsimony BS = 92) andmembers of the R-clade clustered with species of the genusSerratia (parsimony BS = 94)
Additionally 16S rDNA sequences of aphid-associatedbacteria obtained here fell into three other clades con-taining previously described insect endosymbionts(Fig 2) First bacteria from the distantly related aphidsAphis spiraecola a Myzocallis sp and Wahlgreniella nervataformed a strongly supported clade (parsimony BS = 100)which included Arsenophonus triatominarum (Hypsa amp Dale1997) a symbiont of the kissing bug Triatoma infestans(Hemiptera Reduviidae) along with secondary symbiontsof several psyllids (Thao et al 2000a) and a whitefly(Spaulding amp von Dohlen 2001) In our analysis this cladewas the sister taxon to the free-living bacterium Proteus vul-garis We will refer to this symbiotic group as Ars mdash short forArsenophonus the genus name of the T infestans symbiontand of the male-killing symbiont of Nasonia vitripennis(Gherna et al 1991) to which our bacteria were also related(data not shown) Next a bacterium associated withEulachnus pallidus fell within a clade which included sym-bionts of tsetse flies weevils and some psyllids (parsimonyBS = 77) We will refer to this clade as So-So short forSitophilus oryzae Primary Endosymbiont (Heddi et al 1999)and Sodalis glossinidius (Dale amp Maudlin 1999) mdash names fortwo symbionts within this group Finally bacteria fromPemphigus populi and Tetraneura ulmi formed a mono-phyletic clade (parsimony BS = 100) nested within the cladeincluding the T- and U-types (parsimony BS = 92) We will referto these bacteria as the V-type While we use the term symbiontwhen referring to newly described aphid associates of theArs So-So and V clades we point out that further researchon their transmission and localization is required for defin-itive proof and elucidation of their symbiotic lifestyles
Symbiont 16S rDNA phylogenies conflicted with phylo-genies of their hosts For example the T-type symbiontsof several members of the genus Uroleucon were moreclosely related to a T-type symbiont from the psyllid C pyri
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1069
than to those from other Uroleucon species (Fig 2) Alsothe T-like symbionts of a whitefly and a ladybird beetle didnot occupy a basal position amongst the T-type symbiontsas would be expected under co-speciation A direct compar-ison between the phylogenies of closely related hosts andtheir U-type symbionts revealed several inconsistencies(Fig 3) mdash at least four rearrangements of the (unrooted)symbiont tree are required to make this tree match that of thehosts Finally the RELL test relative support value for thesymbiont topology was a striking 100 (data not shown) mdashindicating that this tree topology was far more likely thanany of the host-tree topologies given our U-type 16S rDNAdataset
Despite these patterns of incongruency several sym-bionts from conspecific aphids collected from differentgeographical locations or at different times were highly related
These included the R- and U-type symbionts from Acyrthosi-phon pisum and the U-type from Macrosiphum euphorbiaeWhereas the A pisum R-type and M euphorbiae U-type wereeach monophyletic (parsimony BS = 79 and 100 respectively)the A pisum U-type sequences fell into a clade which alsoincluded the U-type of U rudbeckiae (parsimony BS = 86)
Relative rates and pairwise divergences
R-type 16S rRNA genes evolved at a rate consistent withthose of free-living bacteria such as Escherichia coli andSerratia but more slowly than those of Buchnera aphidicolathe T-type and the U-type (Table 3) T-type 16S rDNAsequences were found to evolve at a rate not significantlydifferent from U-type or B aphidicola 16S rDNA sequencesbut significantly faster than 16S rDNA of free-living lineages
Fig 2 Maximum parsimony phylogeny ofaphid-associated bacteria and their relativesOne thousand bootstrap replicates wereperformed for both distance and parsimonysearches Bootstrap values from the parsimonysearch are located above the relevantbranches (or arrows) while those from thedistance analysis in agreement with theparsimony tree topology are located belowbranches (or arrows) Names of free-livingbacteria are in underlined italics names ofaphid symbionts (named after their hostsexcept for Buchnera aphidicola) are in plainitalics names of symbionts of other insects(all but Sodalis glossinidius and Arsenophonustriatominarum are named after their hosts)are presented in bold italics Bacteria whichare considered secondary symbionts (iethey reside in a host with an obligateprimary symbiont) are denoted by an lsquoSSrsquofollowing their listed names Clades includingsuch symbionts are called R T U V Arsand So-So and are encompassed by bars onthe right Several aphid species names arepresent in multiple positions on the tree Todistinguish these collection locations(NY = New York WI = Wisconsin AZ =Arizona CA = California MN = Minnesota)or dates (prime01 = 2001 prime99 = 1999) are placedat the ends of host names
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
Finally U-type 16S rDNA sequences evolved more quicklythan did those of free-living bacteria yet more slowly thanB aphidicola 16S rDNA
Pairwise divergence analyses of symbiont and free-livingbacteria revealed that secondary symbionts of a given typeshow high levels of sequence similarity (table availablefrom corresponding author) Within the R- T- and U-cladesall compared sequences were less than 05 16 and 22divergent respectively Strikingly T-type symbionts in thepsyllid C pyri and several Nearctic aphids of the subgenusUroleucon were identical over 1301 compared nucleotidesThe U-clade was the most genetically diverse with anaverage of 12 16S rDNA divergence between symbiontsin different host species compared to 04 for the T-type
Pairwise divergence comparisons
Comparisons between the 16S rDNA distances of B aphidicolaand secondary symbionts infecting pairs of hosts sharingthe same most recent common ancestor are presented inTable 4 In all but one case the divergences between Baphidicola sequences were at least twice as large as thosebetween secondary symbionts of equidistant hosts Only412 comparisons were consistent with the possibilityof strict secondary symbiont co-speciation under theassumption that 16S rDNA substitution rates should notdiffer by more than four-fold the U-type infections inPemphigus betae and Macrosiphoniella ludovicianae (Pemphiginivs Macrosiphini) Chaitophorus populeti and M ludovicianae(Chaitophorini vs Macrosiphini) and A pisum and Uastronomus (Acyrthosiphon vs Uroleucon) and the R-type
infection in A pisum and U caligatum (Acyrthosiphon vsUroleucon) Preliminary analyses suggested that many R-type16S rDNA sequence divergences were inconsistent withco-speciation For example divergences between sym-bionts infecting hosts of different tribes were as low as044 when compared over approximately 1000 basepairs However because these sequences were incompletethey were excluded from formal divergence analyses
Discussion
Symbiont diversity
Previous studies that did not include DNA sequenceinformation suggested that a number of aphid speciesharbour vertically transmitted facultative bacterial asso-ciates that reside in various host tissues (eg Buchner1965 Fukatsu amp Ishikawa 1998) Observations on theirdis-tributions revealed that they had been independentlyacquired by and possibly lost from aphid lineagesHowever because the bacteria were not identified noconclusions could be made regarding symbiont diversitythe phylogenetic breadth of the hosts infected by relatedmicrobes or the likelihood of occasional co-speciationIncluding the six groups of bacteria described here aphidshave been reported to associate with secondary symbiontsspanning 10 distinct bacterial lineages (Unterman et al1989 Chen et al 1996 Jeyaprakash amp Hoy 2000 Darbyet al 2001 Fukatsu 2001 Fukatsu et al 2001 Sandstroumlmet al 2001) Given our serendipitous discovery of three
Table 3 Relative rates tests on the 16S rRNA genes of secondarysymbionts
B aphidicola Buchnera aphidicolaKx3 = 16S rDNA distance between taxon x and the outgroup taxon 3 (ie Vibrio cholerae)daggerDifference is significant at α = 005
Fig 3 Incongruency between host and symbiont trees revealshorizontal transfer of U-type symbionts of the Macrosiphini Thehost phylogeny (left) is redrawn from Fig 1(c) of Moran et al 1999The unrooted U-type symbiont phylogeny (right) was obtainedthrough parsimony searches including only the U-type 16S rDNAsequences of the taxa shown here Dashed arrows connect hoststo their respective symbionts (named after hosts) and suggestseveral inconsistencies This host topology was one of just threepossible alternatives because of a trichotomy involving (Uae)(Usl) and (Uru Uas) the other topologies were also incongruentwith that of the symbiont tree (not shown) Name abbreviationsnot presented in Table 1 are as follows Mlu = Macrosiphoniellaludovicianae Uhe = Uroleucon helianthicola Usl = Uroleucon solida-ginis Uae = Uroleucon aenum Uas = Uroleucon astronomus
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1071
new aphid associates it is likely that a substantial numberof the microbial guests residing in aphids have yet to bedescribed
Research on other members of the Sternorrhyncha hasrevealed similar patterns of symbiont diversity For exam-ple the psyllid Diaphorina citri was found to associate withfive distinct bacteria which were occasionally all found toreside within the same host individuals (Subandiyah et al2000) In addition at least four distinct bacterial symbiontshave been found in whiteflies (Clark et al 1992 Costa et al1995 Jeyaprakash amp Hoy 2000 Spaulding amp von Dohlen2001) and at least five infect scale insects (Fukatsu amp Nikoh2000 Thao et al 2002) Thus these related plant sap feedersare common habitats for a number of independently derivedsymbionts
Symbiont transmission
Within laboratory colonies aphid secondary symbiontsundergo highly efficient vertical transmission with noobserved instances of horizontal transfer (Chen amp Purcell1997 Sandstroumlm et al 2001) These observations suggestthat vertical transfer plays a large role in the persistence ofsymbionts within populations Despite this importanceour examination of a wide range of host species providesno definitive evidence for co-speciation indicating thathorizontal transfer is in large part responsible for R-T- and U-type distributions across species Given ourobservations of low 16S rDNA divergences betweensymbionts in different hosts it appears that a number ofhorizontal transfer events occurred recently with T-type
symbionts having undergone an especially recent radiationonto their hosts Thus interspecies movement is probablyan ongoing process for these bacteria
Evidence similar to that presented here has been used toargue that a number of facultative symbionts includingWolbachia pipientis Sodalis glossinidius and several psyllidassociates have been independently acquired by differenthost lineages (OrsquoNeill et al 1992 Werren et al 1995 Aksoyet al 1997 Thao et al 2000a) Despite this evidence themeans by which facultative symbionts move betweenarthropod species remain obscure Some researchers haveproposed a role of transmission through host plants(Darby et al 2001) Given the similarity of symbionts inaphids and ladybird beetles (Werren et al 1994 Chen et al1996 Majerus et al 1999 Fukatsu et al 2001) it is temptingto propose the involvement of predatorndashprey interactionsSeveral investigations have found similar W pipientis inhosts and their parasites suggesting possible interspecifictransmission through hostndashparasite interactions (Vavreet al 1999 Cordaux et al 2001 Noda et al 2001) Accord-ingly experimental studies have documented the transferof W pipientis between a host and its parasitoid andbetween conspecific parasitoids infecting the same host(Heath et al 1999 Huigens et al 2000) However there isstill no evidence of a means by which nonparasitoids canacquire symbionts We note that even rare transfer eventsmay be sufficient to explain the observed patterns
In contrast to patterns suggesting interspecific hori-zontal transfer of secondary symbionts several of ourobservations are suggestive of symbionts persisting withina single host species These cases do not necessarily imply
Host species abbreviations not described in Table 1 or Fig 3 are as follows Sgr = Schizaphis graminum Cvi = Chaitophorus viminalis Rma = Rhopalosiphum maidis Uso = Uroleucon sonchi Uam = Uroleucon ambrosiae from Arizona Uca = Uroleucon caligatumdaggerThe secondary symbiont types (ie R T or U) for which divergence values are presented are listed next to their host abbreviation codesDaggerThe smallest distance between B aphidicola infecting members of these taxa is presented heresectThe largest distance between secondary symbionts infecting members of these taxa is presented here
Table 4 16S rDNA divergences of Buchneraaphidicola vs those of secondary symbiontsfor pairs of aphid hosts sharing same mostrecent common ancestor (see text for details)
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
strict vertical transmission they could rather reflect hori-zontal transfer between only conspecifics Overall the phylo-genetic and genetic similarities of symbionts in a singlespecies would not be expected if rampant interspecifichorizontal transfer was common We therefore take theseobservations to indicate the existence of impediments tosuch lateral movement Such barriers could include limitedopportunities for interspecific transfer limited host rangesinfluenced by survival in and transmission to offspring anew host or frequent extinction of newly infected lineagesas a result of maladaptive phenotypic effects caused bysymbionts Thus despite the appareat vecency of somehorizontal transfer events apparently being recent the per-sistence of symbionts within host species suggests thatintraspecific transfer is more common than interspecificmovement enhancing opportunities for one-to-one co-evolution between aphids and secondary symbionts
Symbiont distributions
Based on our observations of R- T- and U-type symbiontdistributions across multiple aphid tribes and beyond wecan rule out the possibility that these clades are specializedon hosts within the tribe Macrosiphini or that T-typesymbionts are confined to the Sternorrhyncha Howeverthe rarity of these bacteria within the Psylloidea (relative tothe Aphidoidea) suggests that the R- T- and U-typesymbionts may be a more significant component of aphidbiology
Several other facultative symbionts of arthropods havebeen found to infect even more phylogenetically diverseranges of host species For example closely related W pip-ientis symbionts have been found in species spanning theArthropoda (Jeyaprakash amp Hoy 2000 Werren amp Windsor2000) and in nematodes (Sironi et al 1995) Closely relatedsymbionts (over 98 16S rDNA identity) of the genusSpiroplasma have been found in hosts of the insect ordersColeoptera (Majerus et al 1999) Lepidoptera (Jiggins et al2000) Hemiptera (Fukatsu et al 2001) as well as in ticks(Weisburg et al 1989) Also Rickettsia with over 99 16SrDNA similarity infect beetles (Werren et al 1994) aphids(Chen et al 1996) and ticks (Philip et al 1983) These obser-vations indicate that transfer among distant host taxa is acommon feature of facultative symbioses between bacteriaand arthropods and suggest that some symbiont lineagesretain a generalized ability to live in distant host speciesAlternatively gene acquisition by symbionts may permitthe colonization of novel hosts such acquisition couldbe mediated by bacteriophage that are known to associ-ate with secondary symbionts of aphids (van der Wilket al 1999 Sandstroumlm et al 2001) In contrast to theseobservations high genetic similarity of symbionts withinAcyrthosiphon pisum and within Macrosiphum euphorbiaesuggests that some symbionts are adapted to a lifestyle
within their current hosts lowering their potential forinterspecific horizontal transfer To address questionsregarding the potential host ranges of secondary sym-bionts we propose an experimental approach involvingmicroinjection of symbionts into novel host species
History of associations
The overall lack of phylogenetic congruency and thedisparity in the divergences of secondary symbiontsand obligate symbionts as shown here suggest thatassociations between specific hosts and their currentsecondary symbionts are quite young While high geneticsimilarity between R-type symbionts and free-livingSerratia (ie 23 divergence between the Aphis craccivoraR-type and Serratia entomophila over 1135 nucleotides)suggests a recent origin of symbiosis the story for the T-and U-types is more complicated Given that T-type 16SrRNA substitution rates are similar to those of Buchneraaphidicola (Table 3) applying the rate calibrated for Baphidicola (ie 2ndash4 substitution per hundred million yearsMoran et al 1993) suggests that those described in thisstudy shared a common ancestor no more than 20ndash40million years ago (based on the maximum divergence of16 between the T-types of Bemisia tabaci and Chilomenessexmaculatus) However the monophyly of the T- U- andV-types (Fig 2) suggests that the age of symbiosis in thisclade of microbes is ancient mdash average sequence diver-gence between T- and U-types was around 8 revealinga minimum age of 100 million years Also the elevatedsubstitution rates of T- and U-type bacteria vs those offree-living bacteria are consistent with a long historyof symbiosis To determine more conclusively whetherthe ancestor of these bacteria was a symbiont extensivescreening of nonsymbiotic environments for bacteria inthis clade would be necessary It would also be usefulto determine whether the similar lifestyles of T- U- andpossibly V-type symbionts rely on homologous pathwaysand whether the genes for these pathways were present intheir common ancestor
Importance of facultative symbiosis
Only a few attempts have been made to estimate pheno-typic effects associated with infection by secondary sym-bionts and each of these has focused on symbionts of thepea aphid A pisum Two bacteria the R-type and sym-bionts of the genus Spiroplasma have been shown todepress host fitness slightly under certain environmentalconditions (Chen et al 2000 Fukatsu et al 2001) Yet theR-type symbiont confers benefits at high temperatures(Chen et al 2000 Montllor et al 2002) possibly expandingthe range of niches available to infected A pisumFurthermore a recent study suggests that single isolates of
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1073
R- and T-type symbionts confer resistance to parasitismby the hymenopteran parasitoid Aphidius ervi (Oliver et alin press)
Several studies have revealed a high frequency of sec-ondary symbionts within aphid populations For examplethe T-type was found in 4040 Uroleucon ambrosiae collectedfrom various regions throughout the USA (Sandstroumlm et al2001) while another γ-Proteobacterial symbiont known asYSMS was found to infect 100 of surveyed individualsbelonging to two species within the aphid genus Yamato-callis (Fukatsu 2001) In addition a survey of CaliforniaA pisum populations described the R-type in 5057 indi-viduals (Chen amp Purcell 1997) while the U-type symbiontwas found in 219858 individuals surveyed across Japan(Tsuchida et al 2002) Given the observations on their pre-valence within and incidence across species along withtheir significant effects on host fitness in a variety of eco-logical and environmental contexts we conclude thatfacultative symbionts have probably played a significantrole in the ecology and evolution of aphids
Acknowledgements
We thank J M Michelena for insect collecting and identificationand Paul Baumann for providing psyllid DNA samples Also wethank Olga Cuesta Kim Hammond Ben Wang and Helen Dunbarfor technical assistance This research was funded by National Sci-ence Foundation Grant DEB-9978518 to N Moran grant GV01-177from Oficina de Ciegravencia y Tecnologia del Govern Valenciagrave to ALatorre and grant BFM2000-1313 from Ministerio de Ciencia yTeconologiacutea (MCYT) to A Moya
References
Adachi J Hasegawa M (1996) MOLPHY Programs for MolecularPhylogenetics Based on Maximum Likelihood Version 23 TheInstitute of Statistical Mathematics Tokyo
Aksoy S Chen X Hypsa V (1997) Phylogeny and potential trans-mission routes of midgut-associated endosymbionts of tsetse(Diptera Glossinidae) Insect Molecular Biology 6 183ndash190
Baumann P Baumann L Lai CY et al (1995) Genetics physiologyand evolutionary relationships of the genus Buchnera mdash intra-cellular symbionts of aphids Annual Review of Microbiology 4955ndash94
Bender W Spierer P Hogness DS (1983) Chromosomal walkingand jumping to isolate the Ace and rosy loci and the bithoraxcomplex in Drosophila melanogaster Journal of Molecular Biology168 17ndash33
Buchner P (1965) Endosymbionts of Animals with Plant MicroorganismsInterscience New York
Chen DQ Purcell AH (1997) Occurrence and transmission offacultative endosymbionts in aphids Current Microbiology 34220ndash225
Chen DQ Campbell BC Purcell AH (1996) A new Rickettsia froma herbivorous insect the pea aphid Acyrthosiphon pisum(Harris) Current Microbiology 33 123ndash128
Chen DQ Montllor CB Purcell AH (2000) Fitness effects of twofacultative endosymbiotic bacteria on the pea aphid Acyrthosi-
phon pisum and the blue alfalfa aphid A kondoi EntomologiaExperimentalis et Applicata 95 315ndash323
Clark MA Baumann L Munson MA et al (1992) The Eubacterialendosymbionts of whiteflies (Homoptera Aleyrodoidea) con-stitute a lineage distinct from the endosymbionts of aphids andmealybugs Current Microbiology 25 119ndash123
Clark MA Moran NA Baumann P Wernegreen JJ (2000) Co-speciation between bacterial endosymbionts (Buchnera) and arecent radiation of aphids (Uroleucon) and pitfalls of testing forphylogenetic congruence Evolution 54 517ndash525
Cordaux R Michel-Salzat A Bouchon D (2001) Wolbachia infectionin crustaceans novel hosts and potential routes for horizontaltransmission Journal of Evolutionary Biology 14 237ndash243
Costa HS Westcot DM Ullman DE et al (1995) Morphologicalvariation in Bemisia endosymbionts Protoplasma 189 194ndash202
Dale C Maudlin I (1999) Sodalis gen nov and Sodalis glossinidiussp nov a microaerophilic secondary endosymbiont of the tsetse flyGlossina morsitans morsitans International Journal of SystematicBacteriology 49 267ndash275
Darby AC Birkle LM Turner SL Douglas AE (2001) An aphid-borne bacterium allied to the secondary symbionts of whiteflyFEMS Microbiology Ecology 36 43ndash50
Douglas AE Prosser WA (1992) Synthesis of essential amino-acidtryptophan in the pea aphid (Acyrthosiphon pisum) symbiosisJournal of Insect Physiology 38 565ndash568
Fukatsu T (2001) Secondary intracellular symbiotic bacteria in aphidsof the genus Yamatocallis (Homoptera aphididae Drepanosiphinae)Applied and Environmental Microbiology 67 5315ndash5320
Fukatsu T Ishikawa H (1998) Differential immunohistochemicalvisualization of the primary and secondary intracellularsymbiotic bacteria of aphids Applied Entomology and Zoology 33321ndash326
Fukatsu T Nikoh N (2000) Endosymbiotic microbiota of thebamboo pseudococcid Antonina crawii (Insecta Homoptera)Applied and Environmental Microbiology 66 643ndash650
Fukatsu T Aoki S Kurosu U Ishikawa H (1994) Phylogeny ofCerataphidini aphids revealed by their symbiotic micro-organisms and basic structure of their galls mdash implications forhost-symbiont coevolution and evolution of sterile soldier castesZoological Science 11 613ndash623
Fukatsu T Nikoh N Kawai R Koga R (2000) The secondary endo-symbiotic bacterium of the pea aphid Acyrthosiphon pisum(Insecta Homoptera) Applied and Environmental Microbiology66 2748ndash2758
Fukatsu T Tsutomu T Nikoh N Koga R (2001) Spiroplasmasymbiont of the pea aphid Acyrthosiphon pisum (Insecta Hom-optera) Applied and Environmental Microbiology 67 1284ndash1291
Funk DJ Helbling L Wernegreen JJ Moran NA (2000) Intra-specific phylogenetic congruence among multiple symbiontgenomes Proceedings of the Royal Society of London Series B-Biological Sciences 267 2517ndash2521
Gherna RL Werren JH Weisburg W (1991) Arsenophonus nasoniaegen nov sp nov the causative agent of the son-killer trait in theparasitic wasp Nasonia vitripennis International Journal of System-atic Bacteriology 41 563ndash565
Heath BD Butcher RDJ Whitfield WGF Hubbard SF (1999)Horizontal transfer of Wolbachia between phylogenetically dis-tant insect species by a naturally occurring mechanism CurrentBiology 9 313ndash316
Heddi A Grenier AM Khatchadourian C Charles H Nardon P(1999) Four intracellular genomes direct weevil biologyNuclear mitochondrial principal endosymbiont and Wolbachia
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
Proceedings of the National Academy of Sciences of the United Statesof America 96 6814ndash6819
Huigens ME Luck RF Klaassen RHG et al (2000) Infectiousparthenogenesis Nature 405 178ndash179
Hypsa V Dale C (1997) In vitro culture and phylogenetic analysisof lsquoCandidatus Arsenophonus triatominarumrsquo an intracellularbacterium from the triatomine bug Triatoma infestans Inter-national Journal of Systematic Bacteriology 47 1140ndash1144
Jeyaprakash A Hoy MA (2000) Long PCR improves WolbachiaDNA amplification wsp sequences found in 76 of sixty-threearthropod species Insect Molecular Biology 9 393ndash405
Jiggins FM Hurst GDD Jiggins CD von der Schulenburg JHGMajerus MEN (2000) The butterfly Danaus chrysippus is infectedby a male-killing Spiroplasma bacterium Parasitology 120 439ndash446
Kishino H Hasegawa M (1989) Evaluation of the maximum like-lihood estimate of the evolutionary tree topologies from DNAsequence data and the branching order in Hominoidea Journalof Molecular Evolution 29 170ndash179
Latorre A Moya A Ayala FJ (1986) Mitochondrial DNA poly-morphism of Drosophila subobscura Proceedings of the NationalAcademy of Sciences of the United States of America 83 8649ndash8653
Maddison DR Maddison WP (2000) Macclade Analysis of Phylogenyand Character Evolution Sinauer Associates Sunderland MA
Majerus TMO von der Schulenburg JHG Majerus MEN HurstGDD (1999) Molecular identification of a male-killing agent inthe ladybird Harmonia axyridis (Pallas) (Coleoptera Coccinellidae)Insect Molecular Biology 8 551ndash555
Marchuk D Drumm M Saulino A Collins FS (1992) Constructionof T-vectors a rapid general system for direct cloning ofunmodified PCR products Nucleic Acids Research 19 1154
Martinez-Torres D Buades C Latorre A Moya A (2001) Molecularsystematics of aphids and their primary endosymbionts Mole-cular Phylogenetics and Evolution 20 437ndash449
Montllor CB Maxmen A Purcell AH (2002) Facultative bacterialendosymbionts benefit pea aphids Acyrthosiphon pisum underheat stress Ecological Entomology 27 189ndash195
Moran NA Kaplan ME Gelsey MJ Murphy TG Scholes EA (1999)Phylogenetics and evolution of the aphid genus Uroleucon basedon mitochondrial and nuclear DNA sequences Systematic Ento-mology 24 85ndash93
Moran NA Munson MA Baumann P Ishikawa H (1993) A mole-cular clock in endosymbiont bacteria is calibrated using theinsect host Proceedings of the Royal Society of London Series B-Biological Sciences 253 167ndash171
Munson MA Baumann P Clark MA et al (1991) Aphid-eubacterialendosymbiosis evidence for its establishment in an ancestor offour aphid families Journal of Bacteriology 173 6321ndash6324
Noda H Miyoshi T Zhang Q et al (2001) Wolbachia infectionshared among planthoppers (Homoptera Delphacidae) andtheir endoparasite (Strepsiptera Elenchidae) a probable case ofinterspecies transmission Molecular Ecology 10 2101ndash2106
Ochman H Wilson AC (1987) Evolution in bacteria mdash evidencefor a universal substitution rate in cellular genomes Journalof Molecular Evolution 26 74ndash86
Ochman H Elwyn S Moran NA (1999) Calibrating bacterial evo-lution Proceedings of the National Academy of Sciences of the UnitedStates of America 96 12638ndash12643
Oliver KM Russell JA Moran NA Hunter MS (2003) Facultativebacterial symbionts in aphids confer resistance to parasiticwasps Proceedings of the National Academy of Sciences of the UnitedStates of America 100 1803ndash1807
OrsquoNeill SL Giordano R Colbert AME Karr TL Robertson HM(1992) 16S rRNA phylogenetic analysis of the bacterial endo-symbionts associated with cytoplasmic incompatibility ininsects Proceedings of the National Academy of Sciences of theUnited States of America 89 2699ndash2702
Philip RN Casper EA Anacker RL et al (1983) lsquoRickettsia belliirsquo spnov a tick borne rickettsia widely distributed in the UnitedStates that is distinct from the spotted fever and typhusbiogroups International Journal of Systematic Bacteriology 3394ndash106
Remaudiegravere G Remaudiegravere M (1997) Catalogue des Aphididae duMonde Homoptera Aphidoidea INRA Paris
Robinson M Gouy M Gautier C Mouchiroud D (1998) Sensitivityof the relative-rate test to taxonomic sampling Molecular Biologyand Evolution 15 1091ndash1098
Robinson-Rechavi M Huchon D (2000) RRTree Relative-RateTests between groups of sequences on a phylogenetic tree Bio-informatics 16 296ndash297
Rouhbakhsh D Lai CY von Dohlen CD et al (1996) The tryp-tophan biosynthetic pathway of aphid endosymbionts (Buchn-era) genetics and evolution of plasmid-associated anthranilatesynthase (trpEG) within the Aphididae Journal of Molecular Evo-lution 42 414ndash421
Sandstroumlm J Moran N (1999) How nutritionally imbalanced isphloem sap for aphids Entomologia Experimentalis et Applicata91 203ndash210
Sandstroumlm J Russell JA White JP Moran NA (2001) Independentorigins and horizontal transfer of bacterial symbionts of aphidsMolecular Ecology 10 217ndash228
Sironi M Bandi C Sachhi L DiSacco B Damiani G Genchi C(1995) Molecular evidence for a close relative of the arthropodendosymbiont Wolbachia in a filarial worm Molecular and Bio-chemical Parasitology 74 223ndash227
Spaulding AW von Dohlen CD (2001) Psyllid endosymbiontsexhibit patterns of cospeciation with hosts and destabilizingsubstitutions in ribosomal RNA Insect Molecular Biology 10 57ndash67
Subandiyah S Nikoh N Tsuyumu S Somowiyarjo S Fukatsu T(2000) Complex endosymbiotic microbiota of the citrus psyllidDiaphorina citri (Homoptera Psylloidea) Zoological Science 17983ndash989
Swofford DL (2001) PAUP Phylogenetic Analysis Using Parsimony(And Other Methods) Sinauer Associates Sunderland MA
Thao ML Clark MA Baumann L et al (2000a) Secondary endo-symbionts of psyllids have been acquired multiple timesCurrent Microbiology 41 300ndash304
Thao ML Moran NA Abbot P et al (2000b) Cospeciation ofpsyllids and their primary prokaryotic endosymbionts Appliedand Environmental Microbiology 66 2898ndash2905
Thao ML Gullan PJ Baumann P (2002) Secondary (γ-Proteobacteria)endosymbionts infect the primary (β-Proteobacteria) endo-symbionts of mealybugs multiple times and coevolve withtheir hosts Applied and Environmental Microbiology 68 3190ndash3197
Tsuchida T Koga R Shibao H Matsumoto T Fukatsu T (2002)Diversity and geographic distribution of secondary endosymbioticbacteria in natural populations of the pea aphid Acyrthosiphonpisum Molecular Ecology 11 2123ndash2135
Unterman BM Baumann P McLean DL (1989) Pea aphid symbiontrelationships established by analysis of 16S ribosomal-RNAsJournal of Bacteriology 171 2970ndash2974
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts
D I S T R I B U T I O N S O F A P H I D - A S S O C I A T E D S Y M B I O N T S 1075
van der Wilk F Dullemans AM Verbeek M van den Heuvel JF(1999) Isolation and characterization of APSE-1 a bacteriophageinfecting the secondary endosymbiont of Acyrthosiphon pisumVirology 262 104ndash113
van Ham RC Moya A Latorre A (1997) Putative evolutionaryorigin of plasmids carrying the genes involved in leucinebiosynthesis in Buchnera aphidicola (endosymbiont of aphids)Journal of Bacteriology 179 4768ndash4777
Vavre F Fleury F Lepetit D Fouillet P Bouletreau M (1999)Phylogenetic evidence for horizontal transmission of Wolbachiain hostndashparasitoid associations Molecular Biology and Evolution16 1711ndash1723
von Dohlen CD Moran NA (2000) Molecular data support a rapidradiation of aphids in the Cretaceous and multiple origins ofhost alternation Biological Journal of the Linnean Society 71 689ndash717
Weisburg WG Tully JG Rose DL et al (1989) A phylogenetic ana-lysis of the mycoplasmas basis for their classification Journal ofBacteriology 171 6455ndash6467
Werren JH Windsor DM (2000) Wolbachia infection frequencies ininsects evidence of a global equilibrium Proceedings of the RoyalSociety of London Series B-Biological Sciences 267 1277ndash1285
Werren JH Hurst GDD Zhang W Breeuwer JAJ Stouthamer RMajerus MEN (1994) Rickettsial relative associated with malekilling in the ladybird beetle (Adalia bipunctata) Journal of Bac-teriology 176 388ndash394
Werren JH Zhang W Guo LR (1995) Evolution and phylogenyof Wolbachia reproductive parasites of arthropods Proceedingsof the Royal Society of London Series B-Biological Sciences 26155ndash71
Jacob Russell is a PhD candidate in the department of Ecology andEvolution at the University of Arizona in Tucson His researchfocuses on the interactions between aphids and their facultativebacterial symbionts Nancy Moran is a Regentsrsquo Professor in thesame department and studies the biology and evolution ofbacterial symbionts and their insect hosts Professors AmparoLatorre and Andregraves Moya and PhD candidate Beatriz Sabater-Muntildeoz belong to the department of Evolutionary Genetics at theUniversity of Valenciagrave and study the molecular basis of symbiosisin insects as well as the co-evolutionary interactions betweenaphids and their symbionts