THE MIDDLE ORDOVICIAN PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN AND ITS BIOSTRATIGRAPHIC CORRELATION by JO ¨ RG MALETZ* and DAVID L. BRUTON *Department of Geology, University at Buffalo, SUNY, Buffalo, New York, 14260-3050, USA; e-mail: [email protected]Geological Museum, University of Oslo, Boks 1172, Blindern, N-0318 Oslo, Norway; e-mail: [email protected]Typescript received 20 March 2007; accepted in revised form 18 January 2008 Abstract: The Proventocitum procerulum radiolarian assem- blage from the Valhallfonna Formation of Spitsbergen is described for the first time in some detail from material inves- tigated in transmitted light. A number of species are recognized in the well-preserved material, showing unexpected diversity. The age of the fauna is restricted to the early Darriwilian (Da1-2), based on comparison with precisely dated radiolarian assemblages from the Cow Head and Table Head Groups of western Newfoundland. Most species, however, are left in open nomenclature until additional material is available for SEM investigation. Inanigutta magnifica n. sp. is described as new. Key words: Ordovician, Radiolaria, biostratigraphy, Spits- bergen, Proventocitum procerulum. PROVENTOCITUM PROCERULUM is a characteristic radi- olarian species of uncertain relationship that was first described in some detail as Popofskyellum? procerulum Nazarov in Nazarov and Popov (1980) from the Ordovi- cian of Kazakhstan. Illustrations of the type material include a number of line drawings and reflected light photographs, but no SEM photos have been provided. Fortey and Holdsworth (1971) already recognized the uniqueness of this species in their material from Spitsber- gen and identified specimens as Radiolaria familia nova spp., conical form, without providing a detailed descrip- tion and illustration. The only further description of this species is from the Arenig of the Ballantrae Complex in Scotland (Aitchison 1998). The material comes from chert layers and is poorly preserved, but was illustrated from SEM photos. The characteristic shape of the skeleton of Proventoci- tum procerulum made it an easily recognizable form in the Ordovician and it has been used as an important index for biostratigraphy, especially as the associated fau- nal elements were poorly known and mostly consisted of indeterminate, nondescript spherical radiolarians (Aitchi- son 1998). The biostratigraphic distribution of the Proventocitum procerulum – Haplentactinia armillata radiolarian assem- blage (Nazarov 1988), here Proventocitum procerulum assemblage in short, was considered to cover the entire Arenig and the Lower Llanvirn (Nazarov 1988; Noble and Aitchison 2000), the equivalent to the Floian to Lower Darriwilian. Danelian and Popov (2003) found the Lower Arenig Inanihella bakansensis – Inanihella(?) akzhala assemblage below the Proventocitum procerulum assem- blage in Kazakhstan. The term assemblage does not imply that all faunal ele- ments may be present in an individual faunal assemblage and even index species may be lacking. Data from the Cow Head and Table Head Groups of western Newfoundland show that faunal compositions are quite variable and some faunas are represented by very monot- onous or even monospecific assemblages. Biozones based on first appearance (FAD) are impossible to provide for Ordovician radiolarians, as most faunas are individual finds not recorded in any biostratigraphic context. Bio- stratigraphic ranges have not been established for any Ordovician radiolarian species. Faunas of the Proventocitum procerulum assemblage were discovered in a few localities widely scattered over the globe. Nazarov and Popov (1980) described a rich and moderately diverse faunal assemblage from the Kur- chilik Formation of Kazakhstan. They documented the fauna in some detail and illustrated the specimens mostly in light photographs. Nazarov and Popov (1980: p. 7) listed nine radiolarian species, three of which in open nomenclature, and the sponge spicule Anakrusa miriacan- tha (Remarks on the species and a revised list can be found in Appendix 1). [Palaeontology, Vol. 51, Part 5, 2008, pp. 1181–1200] ª The Palaeontological Association doi: 10.1111/j.1475-4983.2008.00803.x 1181
Transcript
THE MIDDLE ORDOVICIAN PROVENTOCITUMPROCERULUM RADIOLARIAN ASSEMBLAGE OFSPITSBERGEN AND ITS BIOSTRATIGRAPHICCORRELATION
by JORG MALETZ* and DAVID L. BRUTON!*Department of Geology, University at Buffalo, SUNY, Buffalo, New York, 14260-3050, USA; e-mail: [email protected]!Geological Museum, University of Oslo, Boks 1172, Blindern, N-0318 Oslo, Norway; e-mail: [email protected]
Typescript received 20 March 2007; accepted in revised form 18 January 2008
Abstract: The Proventocitum procerulum radiolarian assem-
blage from the Valhallfonna Formation of Spitsbergen is
described for the first time in some detail from material inves-
tigated in transmitted light. A number of species are recognized
in the well-preserved material, showing unexpected diversity.
The age of the fauna is restricted to the early Darriwilian
(Da1-2), based on comparison with precisely dated radiolarian
assemblages from the Cow Head and Table Head Groups of
western Newfoundland. Most species, however, are left in open
nomenclature until additional material is available for SEM
investigation. Inanigutta magnifica n. sp. is described as new.
PROVENTOCITUM PROCERULUM is a characteristic radi-olarian species of uncertain relationship that was firstdescribed in some detail as Popofskyellum? procerulumNazarov in Nazarov and Popov (1980) from the Ordovi-cian of Kazakhstan. Illustrations of the type materialinclude a number of line drawings and reflected lightphotographs, but no SEM photos have been provided.Fortey and Holdsworth (1971) already recognized theuniqueness of this species in their material from Spitsber-gen and identified specimens as Radiolaria familia novaspp., conical form, without providing a detailed descrip-tion and illustration. The only further description of thisspecies is from the Arenig of the Ballantrae Complex inScotland (Aitchison 1998). The material comes from chertlayers and is poorly preserved, but was illustrated fromSEM photos.
The characteristic shape of the skeleton of Proventoci-tum procerulum made it an easily recognizable form inthe Ordovician and it has been used as an importantindex for biostratigraphy, especially as the associated fau-nal elements were poorly known and mostly consisted ofindeterminate, nondescript spherical radiolarians (Aitchi-son 1998).
The biostratigraphic distribution of the Proventocitumprocerulum – Haplentactinia armillata radiolarian assem-blage (Nazarov 1988), here Proventocitum procerulumassemblage in short, was considered to cover the entireArenig and the Lower Llanvirn (Nazarov 1988; Noble and
Aitchison 2000), the equivalent to the Floian to LowerDarriwilian. Danelian and Popov (2003) found the LowerArenig Inanihella bakansensis – Inanihella(?) akzhalaassemblage below the Proventocitum procerulum assem-blage in Kazakhstan.
The term assemblage does not imply that all faunal ele-ments may be present in an individual faunal assemblageand even index species may be lacking. Data from theCow Head and Table Head Groups of westernNewfoundland show that faunal compositions are quitevariable and some faunas are represented by very monot-onous or even monospecific assemblages. Biozones basedon first appearance (FAD) are impossible to provide forOrdovician radiolarians, as most faunas are individualfinds not recorded in any biostratigraphic context. Bio-stratigraphic ranges have not been established for anyOrdovician radiolarian species.
Faunas of the Proventocitum procerulum assemblagewere discovered in a few localities widely scattered overthe globe. Nazarov and Popov (1980) described a richand moderately diverse faunal assemblage from the Kur-chilik Formation of Kazakhstan. They documented thefauna in some detail and illustrated the specimens mostlyin light photographs. Nazarov and Popov (1980: p. 7)listed nine radiolarian species, three of which in opennomenclature, and the sponge spicule Anakrusa miriacan-tha (Remarks on the species and a revised list can befound in Appendix 1).
[Palaeontology, Vol. 51, Part 5, 2008, pp. 1181–1200]
ª The Palaeontological Association doi: 10.1111/j.1475-4983.2008.00803.x 1181
Aitchison (1998) documented a similar assemblagefrom the Ballantrae Complex of Scotland. This faunaincludes several species of Proventocitum together withHaplentactinia(?) armillata and a number of further,poorly preserved spherical spumellarians. It was inter-preted to be of mid-Arenig age based on the closeproximity to levels with the Upper Bendigonian toChewtonian graptolite fauna 5 of Stone and Smellie(1988). Maletz and Bruton (2005) referred the Proventoci-tum procerulum assemblage to the Lower to MiddleDarriwilian, based on the observation that the assemblagediffers considerably from Chewtonian to Castlemainian(Upper Floian to Lower Dapingian) assemblages ofwestern Newfoundland and Spitsbergen. A comparison ofradiolarian assemblages from the Lower to Middle Ordo-vician is here attempted to get a better idea on the exactbiostratigraphic distribution of the Proventocitum proce-rulum assemblage and to understand its composition.
THE SPITSBERGEN FAUNA
R. A. Fortey collected the first samples in 1967 and laterFortey and Holdsworth (1971) described the Proventoci-tum procerulum assemblage from a level 185 m above thebase of the Valhallfonna Formation in the ProfilbekkenMember of a beach section south of the Buldebreen armof the Valhallfonna glacier, northern Ny Friesland, Spits-bergen (Fortey and Bruton 1973) (Text-fig. 1). This levelcompares to the Yapeenian to Lower Darriwilian (Maletzand Bruton 2005) or the Upper Arenig to Lower Llanvirnas first suggested by Fortey and Holdsworth (1971). Theauthors reported radiolarians from three levels within aninterval of five meters, but did not document these faunasseparately.
Holdsworth (1977) and Maletz and Bruton (2007)described a number of further radiolarian faunal assem-blages from various levels in the Vallhallfonna Formation.Holdsworth’s (1977) material is largely comparable withthe fauna from sample 2 of Maletz and Bruton (2007)bearing numerous well-preserved radiolarians. The bio-stratigraphy of the Vallhallfonna Formation is establishedfrom the excellent graptolite record (Cooper and Fortey1982) and the radiolarian records are easily correlatedinto the succession (Maletz and Bruton 2007). The heredescribed and illustrated radiolarian material originatesfrom the slides prepared by Holdsworth and used in theoriginal description and illustration by Fortey and Holds-worth (1971). Two sets of slides with a Proventocitumprocerulum assemblage are preserved in the Holdsworthcollection at the Geological Museum (PMO, Oslo, Nor-way):
1) P: slide Nrs. 01-98. The slides include the specimensillustrated by Fortey and Holdsworth (1971). The material
is strongly deteriorated and most of the specimens areindeterminable.
2) SP-P: Nrs. 100-112. The material comes from thesame level. This material is much better preserved and isthe main base for the descriptions and illustrations in thispaper.
Photographic documentation and identification of allspecimens is deposited at the Geological Museum (PMO,Oslo) together with the collection.
The Proventocitum procerulum faunal assemblage is onlyknown from this material in Spitsbergen and additionalspecimens are not available at the moment. Thus, Holds-worth’s microscopic slides remain the only source tounderstand the faunal composition, diversity structureand biostratigraphy of the Proventocitum procerulum radi-olarian fauna of Spitsbergen. However, a detailed compar-ison with radiolarians investigated with the SEM remainsdifficult, as some structural details are not visible in trans-mitted light.
A comparison of the Spitsbergen material is made withMiddle Ordovician radiolarian faunas from the CowHead and Table Head Groups of western Newfoundland(Maletz 2007a,b; and unpublished data) to understandfaunal composition and evolutionary changes, as well ascomparability of SEM and transmitted light representa-tions (Text-fig. 2). Unfortunately, the poor preservationof the Spitsbergen specimens prevents in most instances
TEXT -F IG . 1 . Map of Spitsbergen showing the location of theradiolarian locality (modified from Maletz and Bruton 2007).
1182 PALAEONTOLOGY, VOLUME 51
the detection of internal structures like the initial spiculeor a microsphere.
The faunal composition. It is extremely difficult to giveprecise percentages of the faunal elements, as more than22% of all specimens in the samples are indeterminable,possibly resulting in a considerable bias (Table 1). Theindex species of the P. procerulum assemblage (P. proceru-lum, H.(?) armillata) occur each at only about 3%. Thus,it might be difficult to detect these faunal elements inpoorly preserved and less rich faunas.
The most common species in the assemblage is Sphae-roentactinia hexaspinosa with 13% of all specimens. Thegenus Sphaeroentactinia was discovered first in the UpperFloian (mid Arenig) of Spitsbergen (Maletz and Bruton2007), where it occurs in a number of species. The speci-mens here identified as S. hexaspinosa are very similar tothe slightly older Spitsbergen material and represent theyoungest known specimens of the genus. Species of thegenus Sphaeroentactinia may eventually form good indexspecies for this time interval.
Haplotaeniatum sp. (12%) Inanigutta sp. B (8%) andthe Inanigutta penrosei group (10%) also represent com-mon faunal elements in the P. procerulum faunal assem-blage. However, their taxonomic status is still unclear andmore research is necessary before their usefulness can beestablished.
Antygopora specimens amount to about 8%, thus thegenus is rare in comparison with the older assemblagesfrom Spitsbergen (Maletz and Bruton 2007). None of thespecimens from this sample bears evidence of the pres-ence of outer spines.
Most other faunal elements are rare and form less than1% of the fauna, thus are insignificant for identifying theassemblage. The P. procerulum fauna is dominated bySphaeroentactinia hexaspinosa associated with mediumsized inaniguttids with a variable number of spines, inclu-ing the first forms with few, distinct, long outer spines(Inanigutta penrosei group). The genus Antygopora isstrongly diminished in frequency and diversity. It is notrepresented any more in the Darriwilian assemblages ofthe Table Head Group (Maletz 2007b). Interestingly, spec-imens of the typical Upper Floian genus Nyfrieslandia(Maletz and Bruton 2007) have not been detected in thesample. They, however, may be difficult to differentiatefrom Inanigutta or even Haplotaeniatum in investigationwith transmitted light.
A comparison with the P. procerulum faunal assemblagefrom the Ballantrae Complex (Aitchison 1998) is difficult,as the spherical forms in this assemblage were not differ-entiated due to the poor preservation. The author did notgive detailed information of diversity and frequencies ofindividual species, therefore. The P. procerulum assem-blage from Kazakhsthan (Nazarov and Popov 1980) obvi-
ously lacks the genus Sphaeroentactinia, which is socommon in the Spitsbergen sample and, at the moment,this difference remains unexplained.
THE AGE OF THE PROVENTOCITUMPROCERULUM ASSEMBLAGE
The exact age of the Proventocitum procerulum radiolarianassemblage is difficult to substantiate, as there are nodirectly associated macrofossils available to support anage determination in any of the records. The age determi-nation therefore has to be made indirectly from evidenceof older and younger faunas in successions yieldingmicro- and macrofossils and a comparison with otherradiolarian faunas. The Lower to Middle Ordovicianradiolarian faunas are still poorly known and a biostrati-graphic differentiation does not exist. Nazarov (1988) firstprovided a biostratigraphic zonation for Ordovicianradiolarian faunas (Text-fig. 3). He differentiated theHaplentactinia armillata – Proventocitum procerulum,Haplentactina juncta – Inanigutta unica and Haplentacti-nia baltica – Inanibigutta aculeata zones for the Ordovi-cian, but the Tremadocian interval was left unzoned asfaunas were not available. His Haplentactinia armillata –Proventocitum procerulum interval covered the wholeArenig (Floian to Dapingian). Noble and Aitchison(1995) did not give a biozonation for the Ordovician timeinterval, while Noble and Aitchison (2000) zoned theMiddle to Upper Ordovician subsequently, but did notname index forms. Danelian and Popov (2003) restrictedthe Proventocitum procerulum assemblage to the UpperArenig (Dapingian) and Lower Darriwilian.
The Proventocitum procerulum assemblage fromKazakhstan is from sample 19, collected about 4 kmsoutheast of Sarykum railway station, about 70 km westof the town of Balkhash (Danelian and Popov 2003).Nazarov and Popov (1980) originally considered theradiolarian fauna to be of Upper Arenig age, based onassociated microfaunas, but recent revisions refer theassemblage to the Lower Abereiddian or Lower Llanvirn(Appolonov et al. 1990; Danelian and Popov 2003).Aitchison (1998) described a small and poorly preservedradiolarian fauna from the Ballantrae Complex of Scot-land. The fauna was considered to be of mid Arenig age(Upper Floian), as graptolites of Bendigonian to Chewto-nian age were found in close proximity to the radiolarians(Stone and Rushton 1983; Rushton et al. 1986; Stone andSmellie 1988). The Spitsbergen assemblage with Proven-tocitum procerulum can be considered as of Lower to midDarriwilian age, based on the succession and the presenceof trilobites in the Profilbekken member (Fortey 1974,1975a,b, 1980) that are comparable with those from theTable Head Group of western Newfoundland.
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1183
A
D
E
F
G H
L
B C
I
J K
100 µm100 µm
100 µm
100 µm
100 µm
109.5a
60 µm
50 µm
80 µm
TEXT -F IG . 2 . Comparison of radiolarian specimens in SEM photography and transmitted light photography. Inanigutta sp.,Inanigutta penrosei group, Table Head Group, western Newfoundland. A, GSC 10840, JM 10.167, innermost sphere (microsphere?)with robust outer spines. B, GSC 10841, JM 10.128, inner sphere with partly preserved first outer sphere, showing outer spines andbeams. C, GSC 10842, JM 10.41, two partly preserved spheres with indication of inner microsphere (arrows). Inanigutta sp., Inaniguttapenrosei group, Spitsbergen. D, F, PMO 212.225 ⁄ 18, inner sphere (microsphere?) with strong outer spines and indications of firstouter sphere. Antygopora sp. indet. E, PMO 212.223 ⁄ 20, small sphere without spines. Antygopora irregularia Maletz. G, H, J, PMO212.221 ⁄ 01, sphere with short outer spines and typical antygoporid bandages (J), outer spines continue inside outer sphere (arrows inG), Spitsbergen specimen. I, GSC 130.384, SEM photo, Beothuka terranova assemblage, St. Paul’s Inlet, North section, westernNewfoundland. Sphaeroentactinia hexaspinosa Maletz and Bruton. K, PMO 212.229 ⁄ 04, sample 1 (Maletz and Bruton 2007),Spitsbergen. L, PMO 212.227 ⁄ 5.
1184 PALAEONTOLOGY, VOLUME 51
New knowledge on early to Middle Ordovician radio-larian faunas can help to restrict the biostratigraphicrange of the Proventocitum procerulum assemblage (Fig.3). Rich radiolarian faunas are now found in the Arenig
of western Newfoundland, where the radiolarians occurassociated with graptolites. The graptolite biostratigraphyof the region is very precise (Williams and Stevens 1986,1988) and can be used in detail for international corre-lation. The Cow Head Group of western Newfoundlandmay provide an important key to establish a precise anduseful Ordovician radiolarian biostratigraphy. It provideda number of distinct Upper Cambrian to Middle Ordo-vician radiolarian faunas (Won and Iams 2002; Wonet al. 2005; Maletz and Bruton 2007; Maletz 2007a, b),showing the fast biostratigraphic changes of radiolarianfaunas through time. The youngest of the faunas is fromthe Arienigraptus gracilis Biozone (Castlemainian 4). Thefauna has not yet been described in detail, but a numberof specimens were illustrated by Maletz and Bruton(2007) and Maletz (2007b). The fauna does not includeany specimens of Proventocitum procerulum or Haplen-tactinia(?) armillata. It bears numerous robust Inaniguttaspecimens with long spines that may be included in theInanigutta penrosei group. Other common groupsinclude labyrinthine forms and antygoporids. The CowHead Group ranges into the lower part of the Holmo-graptus lentus Biozone (Maletz, unpublished), but radio-larians have not been found in the uppermost part ofthe unit.
The Middle Ordovician Table Head Group of westernNewfoundland also provided a number of radiolarianspecies (Bergstrom 1979; Renz 1990; Zhang and Pratt2000). The assemblages are of low diversity and bear
TEXT -F IG . 3 . Radiolarian biostratigraphy of the Lower to Middle Ordovician and correlation with graptolite zones.
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1185
common robust Inanigutta specimens of the Inaniguttapenrosei group (Text-fig. 2A–C) associated with less com-mon labyrinthine forms. Proventocitum procerulumand Haplentactinia(?) armillata are not found in the suc-cession, constraining the upper limit of the biostrati-graphic range of this assemblage more precisely. Thesponge Konyrium, however, is present (Bergstrom 1979)as it is in the Kazakhstan assemblage (Nazarov and Popov1980). The Table Head Group covers a biostratigraphicinterval from the upper Holmograptus lentus to the Ptero-graptus elegans Biozone (Albani et al. 2001) or the Histiod-ella tableheadensis and Histiodella kristinae conodontbiozones and the interval with Histiodella bellburnensis (Sto-uge 1984; Albani et al. 2001).
Taxonomy
Repositories. The illustrated material from Spitsbergen is housedin the type collection of the Geological Museum, University ofOslo, Norway (hereafter abbreviated PMO). Material from theCow Head and Table Head Groups are in the collection of theGeological Survey of Canada, Ottawa, Canada (GSC).
Type species. Palaeospiculum burkensis Won in Won and Below1999 from the Middle Cambrian of western Australia.
?Palaeospiculum sp.
Text-fig. 4A–C, G
1971 Possible point-centred, spicular entactiniid spp.Fortey and Holdsworth, p. 38, pl. 10, figs. 6, 9.
Material. 4 complete specimens forming a point-centered spiculewith a variable number of apophyses.
Description. A number of six-rayed, point-centred spicules(hexactine spicules) with few, irregularly placed secondary rays,not produced in whorls, have been discovered in the collection.The primary spines of the spicule are about 100–150 lm longand possess a diameter at the base of 8–10 lm. They slowlytaper towards the tip. Secondary spines or apophyses are com-mon in the central part of the spicule forming a loose centralbody with a diameter of 100–130 lm. The secondary rays or ap-ophyses bend distinctly outwards and can be up to 100 lmlong.
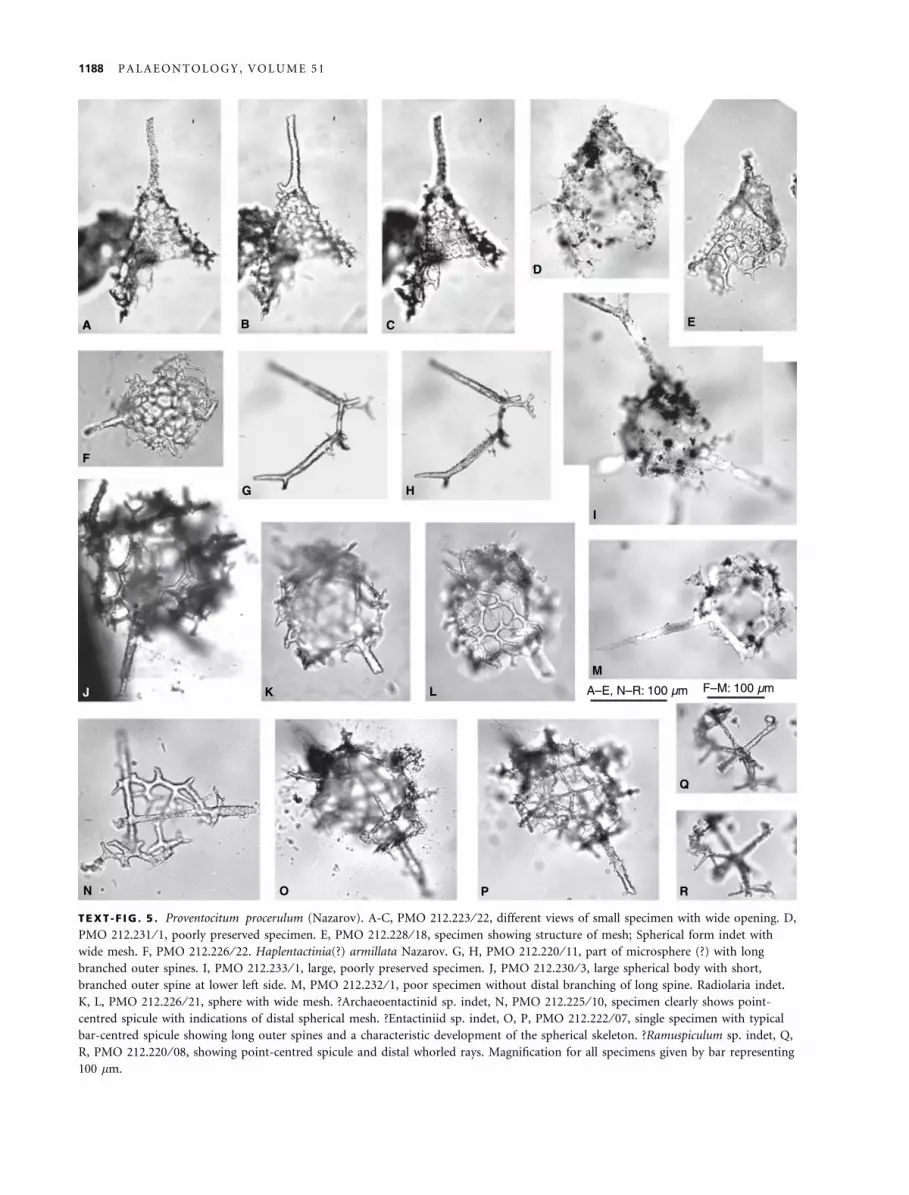
Remarks. The specimens appear to be most close to sim-ple specimens of Palaeospiculum (see Won and Below1999). The form resembles Palaeospiculum curvum speci-mens from the early Tremadocian (Won et al. 2005), buthas a less densely constructed skeleton with few apophy-ses, in which the primary spicule is easily detectable. The‘possible point-centred, spicular entactiniid spp.’ of Wonand Below (1999) may belong to this form, even thoughthe authors describe the specimens as not quite orthogo-nal in symmetry. The specimens represent the youngestknown members of the Paleospiculumidae. A single speci-men has been referred provisionally to Ramuspiculum(Text-fig. 5Q,R), but is too poorly preserved for a specificidentification.
Order ENTACTINARIA Kozur and Mostler, 1982
Family PROTOENTACTINIIDAE Kozur et al., 1996
Genus Protoentactinia Kozur et al., 1996
Type species. Protoentactinia gracilispinosa Kozur et al. 1996 fromthe Tremadocian of North America.
Protoentactinia(?) spp.
Text-figs 4M–P, ?5N–P
1971 Bar centred spicular entactiniid spp. Fortey andHoldsworth, p. 38, pl. 10, figs. 2, 5.
Material. 3 poor specimens.
Remarks. Fortey and Holdsworth (1971) illustrated anddescribed two specimens showing a small spicule withdistal branching and mentioned that these forms are
TEXT -F IG . 4 . ?Palaeospiculum sp., A,B, PMO 212.223 ⁄ 06, spicule with apophyses. C, PMO 212.227 ⁄ 16. G, PMO 212.227 ⁄ 12, smallspecimen. Sphaeroentactinia hexaspinosa Maletz and Bruton, D, H, PMO 212.224 ⁄ 06, small specimen showing sphere development. E,F, PMO 212.227 ⁄ 03, specimen showing three-dimensional sphere development. I, PMO 212.220 ⁄ 18. Spicular form indet, J, PMO212.222 ⁄ 11. K, PMO 212.220 ⁄ 19. L, PMO 212.221 ⁄ 16. Protoentactinia(?) sp., M, PMO 212.223 ⁄ 14, point-centred initial spicule withbranchings. N, O, PMO 212.230 ⁄ 1, large fragment. P, PMO 212.226 ⁄ 11, largest fragment, presence and position of spicule uncertain.Magnification for all specimens given by bar representing 100 lm.
1186 PALAEONTOLOGY, VOLUME 51
A B C
D
100 µmA
H
G F E
I
M
L K J
P O N
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1187
A B C
D
F–M: 100 µmA–E, N–R: 100 µm
A
HG
F
E
I
M
LKJ
P
Q
RON
TEXT -F IG . 5 . Proventocitum procerulum (Nazarov). A-C, PMO 212.223 ⁄ 22, different views of small specimen with wide opening. D,PMO 212.231 ⁄ 1, poorly preserved specimen. E, PMO 212.228 ⁄ 18, specimen showing structure of mesh; Spherical form indet withwide mesh. F, PMO 212.226 ⁄ 22. Haplentactinia(?) armillata Nazarov. G, H, PMO 212.220 ⁄ 11, part of microsphere (?) with longbranched outer spines. I, PMO 212.233 ⁄ 1, large, poorly preserved specimen. J, PMO 212.230 ⁄ 3, large spherical body with short,branched outer spine at lower left side. M, PMO 212.232 ⁄ 1, poor specimen without distal branching of long spine. Radiolaria indet.K, L, PMO 212.226 ⁄ 21, sphere with wide mesh. ?Archaeoentactinid sp. indet, N, PMO 212.225 ⁄ 10, specimen clearly shows point-centred spicule with indications of distal spherical mesh. ?Entactiniid sp. indet, O, P, PMO 212.222 ⁄ 07, single specimen with typicalbar-centred spicule showing long outer spines and a characteristic development of the spherical skeleton. ?Ramuspiculum sp. indet, Q,R, PMO 212.220 ⁄ 08, showing point-centred spicule and distal whorled rays. Magnification for all specimens given by bar representing100 lm.
1188 PALAEONTOLOGY, VOLUME 51
relatively common. However, in the preserved samplethere are no comparable specimens present. A few largerfragments with dichotomous and trichotomous branch-ings are found in the samples and are included here inProtoentactinia(?) sp. A single specimen also resemblingFortey and Holdsworth’s (1971) material has a point-centered spicule and is therefore excluded from Protoen-tactinia. The specimen may belong to the Palaeospiculu-midae, most probably to Ramuspiculum (Text-fig. 5Q,R).There may be more than one species included in thisform, but differentiation is difficult due to the poorpreservation.
Family SPHAEROENTACTINIIDAE Maletz and Bruton, 2007
Genus Sphaeroentactinia Maletz and Bruton, 2007
Type species. Sphaeroentactinia hexaspinosa Maletz and Bruton,2007 from the Middle Arenig (Upper Floian to Lower Dapin-gian) of Spitsbergen.
Diagnosis (Maletz and Bruton 2007). Spherical radiolari-ans with a bar-centred spicule and a sphere formed fromapophyses; spicule eccentrically positioned; spines may ormay not be incorporated in the three-dimensional skele-ton.
Sphaeroentactinia hexaspinosa Maletz and Bruton, 2007
Text-figs 2K, L; 4D–F, H, I
1971 Entactiniid genus novum Fortey and Holdsworth,p. 37, pl. 10, figs. 7, 8.
1971 aff. Entactinia Foreman spp. Fortey and Holdsworth,p. 37, pl. 10, Text-fig. 1.
2007 Sphaeroentactinia hexaspinosa n. sp. Maletz andBruton, p. 264, figs. 6A, 12A-I.
Material. 38 specimens, mostly complete.
Remarks. Maletz and Bruton (2007) described and illus-trated the species in some detail, based on materialmounted on SEM stubs. The here presented material inves-tigated in transmitted light agrees in all observable detailswith the Spitsbergen specimens. Fortey and Holdsworth(1971) described the species as a point-centred entactinid:‘‘six spicule rays, not quite orthogonal in symmetry, meetat a point’’ (Fortey and Holdsworth 1971; p. 37). None ofthe specimens investigated, however, clearly shows a point-centred spicule. The six spines are slightly asymmetrical,suggesting a bar-centred spicule with a short median bar, asis distinctly visible in slightly older specimens from theUpper Floian of Spitsbergen (Text-fig. 2K).
Family HAPLENTACTINIIDAE Nazarov, in Nazarov and Popov
1980
Subfamily HAPLENTACTINIINAE Foreman, 1963
Genus Haplentactinia Foreman, 1963
Type species. Haplentactinia rhinophyusa Foreman, 1963from the Upper Devonian of northern Ohio.
Remarks. The diagnosis of the genus Haplentactinia isfairly vague. The included species originate mainly fromthe Devonian and, thus, the inclusion of an early to Mid-dle Ordovician form is questionable. Unfortunately thedetailed development of this form is unclear as juvenilesand isolated spicules are not available.
Haplentactinia(?) armillata Nazarov, in Nazarov and Popov
1980
Text-fig. 5G–J, ?K–M
?1971 Aff. Stigmosphaera Rust sp. Fortey andHoldsworth, p. 38, pl. 10, Text-fig. 3.
1971 aff. Entactinia Foreman spp. Fortey andHoldsworth, p. 38, pl. 10, Text-fig. 4.
1980 Haplentaetinia armillata Nazarov, sp. nov. Nazarov,p. 55, pl. 6, figs. 1,2; pl. 16, figs. 9,10; Text-Fig. 31.
1988 Haplentactinia armillata (Nazarov); Nazarov, pl. 9,Text-Fig. 12.
1998 Haplentactinia armillata (Nazarov); Aitchison, p. 79,Text-Fig. 2U.
Material. 8 specimens, some broken.
Description. The species may possess a small microsphere of ca.55–60 lm in diameter (Text-fig. 5G, H), but as the only specimenshowing this feature is incomplete, the detailed structure isunclear. The specimen, however, shows a part of a branched struc-ture at the base of two distally branched spines, here interpreted asthe outer spines of Haplentactinia(?) armillata. A branched outerspine is also present in a specimen with a coarse three-dimensionalmesh, measuring ca. 150 lm in diameter (Text-fig. 5I). The outerspines are generally between 100 and 150 lm long, with a stoutbase, slowly tapering towards their branching point. The second-ary spines of these branches are up to 100 lm long, but in mostspecimens they are distinctly shorter, generally 50–60 lm long.They have a maximum diameter of 10 lm. The coarse mesh hasan opening width of 20–25 lm with the bars attaining a diameterof 5–6 lm. The mesh appears to get finer and denser distally, butall specimens are either small or incompletely preserved.
Remarks. Nazarov and Popov (1980) described and illus-trated the species from light photographs and SEM.However, there might be some confusion about the con-cept of this species. Nazarov and Popov (1980), in their
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1189
description (ibid. p. 55) did not mention the SEM illus-trations (ibid. pl. 6, figs. 1, 2), even though they wereclearly labelled in the figure explanation. While the trans-mitted light photo and the drawing of a single specimenshows the characteristically branched outer spines, theSEM illustration does not show this feature. The speci-men might belong to a different labyrinthine species.Unfortunately this specimen is the only example ofHaplentactinia(?) armillata illustrated by Nazarov andOrmiston (1986, pl. 2, figs. 5,6).
Aff. Stigmosphaera Rust described by Fortey and Holds-worth (1971) appears to be a juvenile of Haplentactinia(?)armillata. The authors already recognized similarities toHaplentactinia.
Distribution. The species was found only in Kazakhstan (Naza-rov and Popov 1980) and Britain (Aitchison 1998). The Britishmaterial consists of one poorly preserved specimen from the Bal-lantrae Complex of Scotland, showing as the only distinguishingcharacter the branched outer spines. Haplentactinia(?) armillataoccurs invariably together with Proventocitum procerulum in alllocalities where is has been discovered.
RADIOLARIA ORDER ET FAMILYINDET
Family PROVENTOCITIIDAE Aitchison, 1998
Genus Proventocitum Nazarov and Ormiston, 1984 (emend.
Aitchison 1998)
Type species. Popofskyellum? procerulum Nazarov in Nazarov andPopov 1980 from the Ordovician of Kazakhstan.
Proventocitum procerulum (Nazarov, 1980 in Nazarov and
Popov 1980)
Text-fig. 5A–F
1971 Radiolaria familia nova sp. Fortey and Holdsworth,p. 39, pl. 11, figs. 1, 3, 4, 7, 8; text-Text-Fig. 1b.
1980 Popofskyellum? procerulum Nazarov, sp. nov. Nazarovand Popov, p. 68, pl. 15, figs. 4-9; text-Text-Fig. 42.
1993 Proventocitum procerulum (Nazarov); Nazarov andOrmiston, p. 43, pl. 4, figs. 8, 11; text-Text-Fig. 9b.
1998 Proventocitum procerulum (Nazarov); Aitchison,p. 78, figs. 2A-C, E, G, I, M, N; 3A-F, H-J.
Material. 10 specimens.
Remarks. The species is mainly known from poorly pre-served, possibly recrystallized specimens, not showingmuch detail of the skeleton (see Aitchison 1998). Nazarovand Popov (1980) illustrated good specimens in reflected
light and from drawings, but SEM photos of well-pre-served specimens are not available for this species. Naza-rov and Popov’s (1980) specimens show the thin layer ofmesh formed from bars and the distinct and long outerspines. The relationship between the spines and the elon-gated shell is not visible in the specimens and the drawingin Nazarov and Popov (1980) may be misleading. Thus,the relationship and even inclusion in one of the largergroupings of Palaeozoic radiolarians is impossible at themoment.
Aitchison (1998) included two further forms in thegenus Proventocitum. These, however, are poorly pre-served and known only from the Ballantrae Complex ofScottland. His material appears to be consistently smallerthan the Nazarov and Popov (1980) specimens. The spec-imens, as well as all other material from the BallantraeComplex appear to show strongly coarsened skeletons inwhich most details are destroyed.
The Spitsbergen specimens are well preserved, but frag-mentary. As they are all mounted for transmitted lightinvestigation, surface details cannot be investigated underthe SEM. The material shows that the skeleton is formedfrom thin bars producing irregular meshes.
Spicular form indet.
Text-fig. 4J–L
cf. 1971 Radiolaria familia nova spp., spicular form Forteyand Holdsworth, pl. 11, figs. 2,5; ?text-Text-Fig.1A.
2007 Spicular form indet. Maletz and Bruton, p. 286,Text-Fig. 29A-O.
Material. 6 specimens.
Remarks. This form is fairly rare in the sample. The speci-mens do not add any information to the material describedby Maletz and Bruton (2007), but there is little doubt thatthe specimens from this level belong to the same taxon.
Order SPUMELLARIA Ehrenberg, 1875
Family ANTYGOPORIDAE Maletz and Bruton, 2007
Genus Antygopora Maletz and Bruton, 2005
Type species. Antygopora ordovicica Maletz and Bruton, 2005 fromthe Upper Floian of Spitsbergen.
Diagnosis (emended from Maletz and Bruton2005). Spherical spumellarians with one or more shells,formed by a thin layer pierced by round pores with
1190 PALAEONTOLOGY, VOLUME 51
elevated rims and ornamented on the surface with distinctconnecting ridges forming a honeycomb pattern in some,but not all species; the internal microsphere may producea variable number of external spines.
Remarks. The genus has been recognized in western New-foundland and Spitsbergen (Maletz and Bruton 2005,2007), but may be more widely distributed. Early speciesare spineless, but biostratigraphically younger species beara variable number of outer spines, originating from amicrosphere.
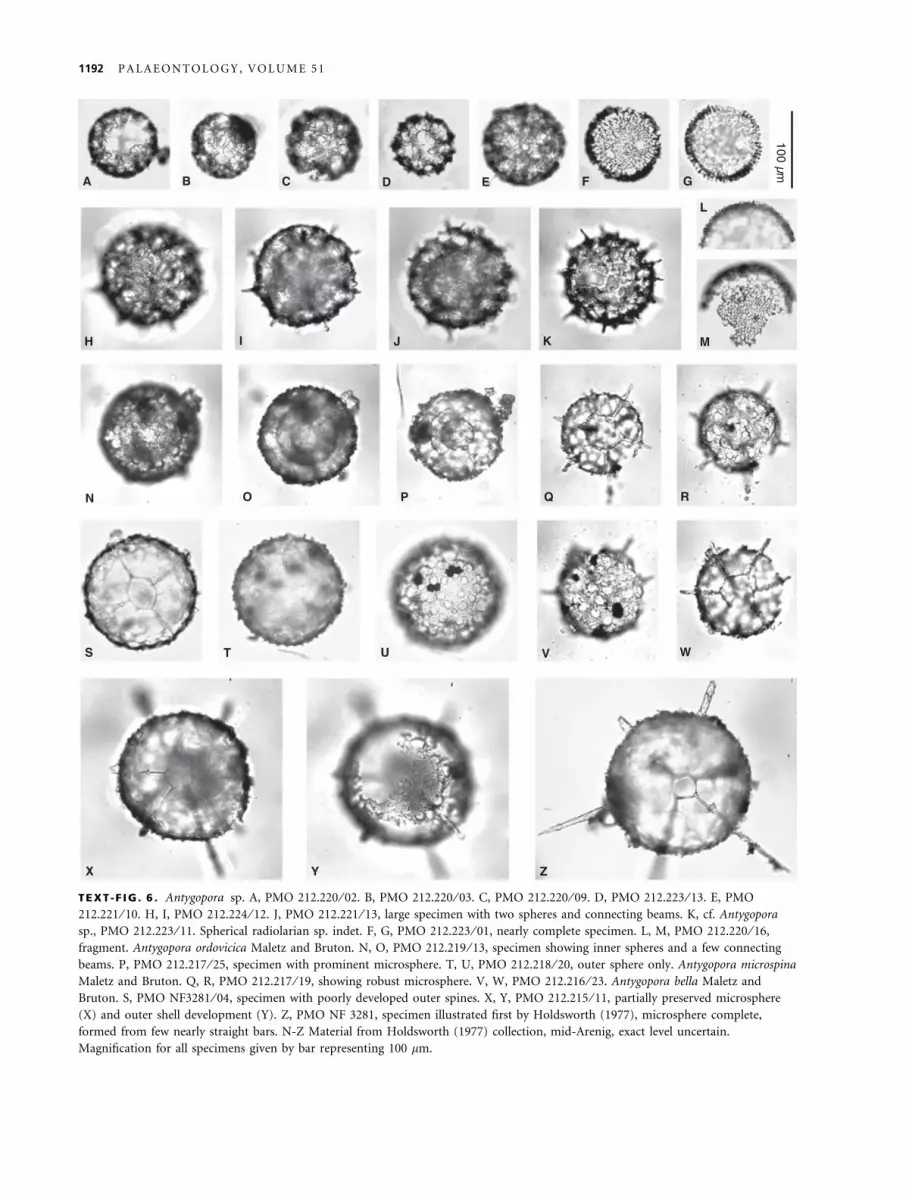
The recognition of Antygopora is based on the roundedpores of their shells (Text-fig. 6U: Antygopora ordovicica).The typical honeycomb pattern of the surface is easilyrecognizable under the SEM (Maletz and Bruton 2005,2007), but impossible to see in transmitted light (Text-fig. 6N, U, Y). A number of older Antygopora specimensfrom the Upper Floian of Spitsbergen are illustrated herefor the first time in transmitted light photographs (Text-fig. 6N–Z). The specimens originate from the Holdsworthcollection. They show the inner spheres and the typicalmicrosphere (Text-fig. 6Q, W, Z) that is difficult to detectunder the SEM and is not preserved in many specimens.
Antygopora sp.
Text-fig. 6A–E, H–K
Material. 24 specimens.
Description. The material includes two groups of specimens thesmaller one of which only shows a single sphere (Text-fig. 6A–D), whereas the second one bears an inner and an outer sphere(Text-fig. 6E, H–J). The material is here interpreted as an onto-genetic series in which the smaller specimens represent the innersphere only. The inner sphere has a diameter of 80–90 lm. Nospines or elaborations are visible. The sphere appears to beformed from ca. 8–10 lm wide bandages surrounding irregularlydistributed, rounded pores. The typical raised rims of Antygo-pora are, unfortunately, not recognizable in the material, inves-tigated under a light microscope. The outer sphere is about110–120 lm in diameter and is similarly developed than theinner sphere. Beams can be recognized to connect the twospheres, and can also be seen on the outer surface of the sphere(Text-fig. 6I, J). They appear to be distributed at regular dis-tances, but it is not possible from the material to recognize adistinct pattern.
Remarks. It is not possible to identify this material withA. ordovicica (Text-fig. 6N–P, T, U) or any other Antygo-pora species. Due to the limitations of the material, it ishere left in open nomenclature. As Antygopora species arecommon and diverse in the Castlemainian (Maletz2007b), the specimens here may represent one of theyoungest species of the genus. A number of specimens
were found in the Holdsworth collection, but had notbeen described before.
Antygopora irregularia Maletz, 2007a
Text-fig. 2G–J
2007a Antygopora irregularia n. sp. Maletz, p. 81,figs. 6A-H, J, K.
Material. 2 specimens mounted on glass slides, additional mate-rial from sample 2 (Maletz and Bruton 2007; Maletz 2007a) con-sidered to belong to the same species.
Remarks. Maletz (2007a) described the species in detailand illustrated the material from SEM photographs, basedon material from the Cow Head Group of western New-foundland. The material here discussed is difficult tocompare as it is investigated with transmitted light only.The specimens agree in their dimensions and develop-ment with the type material from the Cow Head Groupof western Newfoundland (Text-fig. 2I). The preservationmakes it difficult to observe the slightly raised rims onthe bandages (see Text-fig. 2J). However, the typical elon-gated bandages and the rounded pores are obvious in thematerial.
Family INANIGUTTIDAE Nazarov and Ormiston, 1984
Remarks. The family Inaniguttidae includes spericalspumellarians with a variable number of concentricspheres, provided with six or more outer spines. The exactdevelopment of the Inaniguttidae is unknown (De Weveret al. 2001). MacDonald (2006) described the microspherein two species of Plusatispila to have a diameter of33–40 lm. This is the first true record of a microsphere inthe Inaniguttidae, relating the family to the Spumellaria.
MacDonald (2006) and Jones and Noble (2006) dis-cussed the possible synonymy of the Inaniguttidae andthe Haplotaeniatumidae, citing the curved beams of spe-cies of Plusatispila that could be an indiation of intergra-dation of concentric shells (Inaniguttidae) and spiralshells (Haplotaeniatumidae). This is supported by thepresence of a distinct microsphere in both groups, andthe detailed structure of this microsphere might revealtheir true relationships in the future.
Noble (1994: text-fig. 10) gave an overview of the gen-era included in the Inaniguttidae. The differentiation ismainly based on the number of spheres and outer spines,characters that may not be reliable, as it can be seen thatspecies show considerable variation – in part based ontheir ontogeny. MacDonald (2006: p. 33) even suggestedthat the genera Plussatispila, Kalimnasphaera, Inanihella
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1191
B C D
100 µm
A
H
G F E
I M
L
K J
P Q R O N
U V W
X Y Z
T S
TEXT -F IG . 6 . Antygopora sp. A, PMO 212.220 ⁄ 02. B, PMO 212.220 ⁄ 03. C, PMO 212.220 ⁄ 09. D, PMO 212.223 ⁄ 13. E, PMO212.221 ⁄ 10. H, I, PMO 212.224 ⁄ 12. J, PMO 212.221 ⁄ 13, large specimen with two spheres and connecting beams. K, cf. Antygoporasp., PMO 212.223 ⁄ 11. Spherical radiolarian sp. indet. F, G, PMO 212.223 ⁄ 01, nearly complete specimen. L, M, PMO 212.220 ⁄ 16,fragment. Antygopora ordovicica Maletz and Bruton. N, O, PMO 212.219 ⁄ 13, specimen showing inner spheres and a few connectingbeams. P, PMO 212.217 ⁄ 25, specimen with prominent microsphere. T, U, PMO 212.218 ⁄ 20, outer sphere only. Antygopora microspinaMaletz and Bruton. Q, R, PMO 212.217 ⁄ 19, showing robust microsphere. V, W, PMO 212.216 ⁄ 23. Antygopora bella Maletz andBruton. S, PMO NF3281 ⁄ 04, specimen with poorly developed outer spines. X, Y, PMO 212.215 ⁄ 11, partially preserved microsphere(X) and outer shell development (Y). Z, PMO NF 3281, specimen illustrated first by Holdsworth (1977), microsphere complete,formed from few nearly straight bars. N-Z Material from Holdsworth (1977) collection, mid-Arenig, exact level uncertain.Magnification for all specimens given by bar representing 100 lm.
1192 PALAEONTOLOGY, VOLUME 51
(penrosei) and some species of Inanibigutta may belong toa single genus, indicating that the taxonomy of the lateOrdovician to Silurian members of the family Inanigutti-dae is in considerable disarray. However, frequently newgenera are described, neglecting the understanding ofstructural relationships (see MacDonald 2006; Jones andNoble 2006).
Genus Inanigutta Nazarov and Ormiston, 1984
Type species. Entactinia unica Nazarov, 1975 from the UpperOrdovician (Sandbian) of Kazakhstan.
Remarks. Herein, all spherical spumellarians with amicrosphere and a variable number of spheres and outerspines formed from thin rounded bars are provisionallyincluded in the genus Inanigutta, awaiting a revision ofthe inaniguttid genera.
Inanigutta magnifica n. sp.
Text-fig. 7N, R–U
Material. 3 specimens. PMO 212.221 ⁄ 12 (Figs. 7N, R, U) is theholotype. The remaining specimens from the sample are para-types.
Diagnosis. Inanigutta species with poorly developedmicrosphere and two concentric shells formed fromwidely spaced bars; shells connected by beams and outerspines; some irregular curved bars may be present.
Description. The concentric skeleton has a diameter of ca. 180–200 lm in mature specimens with outer spines protruding ca.30–40 lm above the surface of the shell. The microsphere wasdetected through a few spines protruding into the inner pre-served sphere, but details of its development are not available.The inner sphere has a diameter of about 100–120 lm, whilethe second sphere is 180–200 lm in diameter. There are about20 needle-like outer spines, irregularly placed around thespheres. In incomplete specimens the outer spines often showapophyses (Text-fig. 7N, U) that develop in more mature speci-mens into the outer sphere. The spheres are formed from thin,round bars, less than 5 lm in diameter, forming a mesh withmeshes measuring 20–40 lm in width. The bars of these meshesare often slightly curved (Text-fig. 7R). The development of theouter sphere often appears slightly spiral.
Remarks. Inanigutta magnifica n. sp. is easily differenti-ated from Haplotaeniatum sp. through its large size,wide mesh and the presence of two distinct spheres.Haplotaeniatum sp. could easily be mistaken as a juve-nile Inanigutta magnifica, however in poorly preservedspecimens.
Inanigutta penrosei (Ruedemann and Wilson, 1936) group
Text-fig. 8A–R
1936 Haliomma penrosei n. sp. Ruedemann and Wilson,p. 1573, pl. 5, figs. 29, 32; pl. 6, fig. 10.
1980 Helioentactinia penrosei (Ruedemann and Wilson);Nazarov and Popov, p. 50, pl. 4, Text-Fig. 6, pl. 13,Text-Fig. s. 1,2, text-Text-Fig. 27.
1993 Inanihella penrosei (Ruedemann and Wilson);Wang, p. 102, pl. 1, figs. 1-6; pl. 2, figs. 1-9; pl. 3,figs. 1-8; pl. 4, figs. 1-3.
Diagnosis. Robust Inanigutta species with two to threespheres, adorned with ca. 6–8 long and robust outerspines.
Material. 31 specimens in various stages of their development.
Description. Robust spherical radiolarians with at least threespheres (Text-fig. 8P, Q) and a large microsphere (not preservedin most of the specimens) are included in this species. The mostjuvenile specimens consist of a small inner sphere (micro-sphere?) of about 80 lm in diameter (Text-fig. 8A, B). Thesphere shows a wide mesh with the meshes being ca. 15 lmwide and the bars about 2–3 lm wide. Approximately six outerspines may originate from the surface of this microsphere, but acontinuation of the spines inside the microsphere was notobserved. The spines are about 4 lm wide and reach a diameterof 5–6 lm outside the second sphere, before tapering slowlytowards their tips. The spines can be more than 100 lm long.The inner sphere is preserved rarely in larger specimens, possiblyan indication of later resorption as the radiolarian grows. Thesecond sphere has a diameter of 150–170 lm, while the thirdsphere measures 220–240 lm in diameter. The spheres are pro-duced from thin, round bars forming an irregular to hexagonalmeshwork. The density of the mesh varies considerably fromwide (Text-fig. 8G) to dense (Text-fig. 8M), possibly showingontogenetic changes. Initially the developing outer spheres areindicated by apophyses around the outer spines (Text-fig. 8N,R). Beams are present in many specimens supporting the shellstructure.
Remarks. Inanigutta penrosei was first described byRuedemann and Wilson (1936) from thin sections ofNormanskill chert from eastern New York State. Thispoorly preserved material comes from the Upper Ordovi-cian (Sandbian), based on the graptolite faunas of theNormanskill Shales (see Ruedemann 1947). Wang (1993)described it as Inanihella penrosei from the Climacograptusbicornis Biozone of Gansu Province, China, with numer-ous rod-like major spines. However, the illustrations showonly ca. 6–8 outer spines and the specimens compareclosely to the here described material.
The material here described is highly variable andmay belong to more than one species. Specimens withirregular development of the outermost sphere (Text-
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1193
B C D
100
µm
A
H G F
E
I
M L K
J
P Q R O
N
U T S
1194 PALAEONTOLOGY, VOLUME 51
fig. 8J–L) are very similar to Kalimnasphaera. Otherspecimens develop extremely regular spheres with ahigh number of fine beams (Text-fig. 8P, Q), similar toPlussatispila.
Most specimens do not show a preserved microsphere.MacDonald (2006) mentioned that in many inaniguttidsthe microsphere and the first outer sphere are commonlynot preserved. A few juvenile specimens in this collectionshow a well-preserved small inner sphere, possibly a micro-sphere (Text-fig. 5A, B), indicating that the inner skeletonof the inaniguttids may have been resorbed during growthand the material recycled into the outer spheres.
Noble and Aitchison (2000) used the long-spined inan-iguttid zone 1 as a characteristic biostratigraphic intervalcovering the Llanvirn ⁄Llandeilo time interval (=Darriwil-ian 2–4) and correlated it with Nazarov’s (1988) Haplen-tactinia juncta – Inanigutta unica Zone. From this intervalthe first specimens of long-spined inaniguttids wererecorded. However, long-spined inaniguttids appear muchearlier in the late Castlemainian (Maletz and Bruton2007: Fig. 5f). The true identify of these forms, thus,remains uncertain until better data for their specific iden-tification are available.
Inanigutta sp. A
Text-fig. 7M, Q, ?J
Material. 3 specimens.
Diagnosis. Medium sized Inanigutta species with lightframe formed from thin bars; skeleton consists of twospheres and a possible microsphere; spines short and nee-dle-like.
Description. A microsphere is not preserved in any of the speci-mens. The innermost sphere has a diameter of ca. 80 lm. Thesecond sphere measures 140–150 lm in diameter. Both spheresare connected by outer spines that may originate on the surfaceof the inner sphere. The outer spines protrude about 30–40 lmabove the surface of the outer sphere and do not bear apophy-ses. Some of the spines connect the outer and inner spheres, butothers end blind between the two spheres (Text-fig. 7Q). Thebars and spines have a diameter of ca. 3–4 lm. The bars form
the meshes of the spheres with a mesh-width of 20–30 lm.Meshes are irregular and a distinct regular pattern was notobserved.
Remarks. The species differs from Inanigutta magnifica n.sp. through a more delicate meshwork and smaller spherediameter.
Inanigutta sp. B
Text-fig. 9A–L
1971 Sphaeroids – apparently non-spiculate spumellariansFortey and Holdsworth, p. 39, pl. 11, Text-Fig. 6.
Material. 51 specimens.
Diagnosis. Medium sized Inanigutta species with two tothree spherical shells, connected by beams and a variablenumber of slender outer spines.
Description. The skeleton shows at least two spheres connectedwith numerous beams. The inner shell has a diameter of about70–110 lm. The outer sphere varies between 120 and 160 lm,but in some specimens indications of a third sphere are presentas apophyses on the outer spines (Text-fig. 9G, H). The num-ber of outer spines appears to be variable, but at least 8–10spines can be counted in well-preserved specimens. They seemto originate from the surface of the inner sphere and piercethe outer sphere, while a variable number of beams also con-nect between the inner and outer spheres. The development ofboth spheres show identical features, a dense mesh of roundbars, forming irregular pores, measuring 10–15 lm in diameter,slightly less dense in the outer sphere (15–20 lm diameter).The outer spines are ca. 30–50 lm long, measured from thesurface of the second (outer) sphere, and have a diameter ofless than 5 lm.
Remarks. The form shows a high variability in its devel-opment and there may be more than one speciesincluded. Inner spheres are often poorly preserved. Forteyand Holdsworth (1971) described this form as a large,robust form with low spine count and lack of an innershell. The illustration, however, is too poor for a specificidentification, but the dimensions and mesh details visible
TEXT -F IG . 7 . Haplotaeniatum sp. A, F, PMO 212.227 ⁄ 08, juvenile showing indistinct microsphere. B, PMO 212.222 ⁄ 13, juvenilewith spiral arrangement. C, PMO 212.223 ⁄ 08, larger specimen with spiral development, showing density of mesh. D, I, PMO212.220 ⁄ 05, small specimen showing irregular development of mesh. E, PMO 212.224 ⁄ 14, spherical specimen. G, H, PMO 212.223 ⁄ 18,spherical specimen. K, L, PMO 212.227 ⁄ 06, large specimen with typical spiral development of sphere. O, P, PMO 212.224 ⁄ 02,specimen with apparent spherical development showing two spheres. Inanigutta sp. A. J, PMO 212.224 ⁄ 13, larger specimen, irregularshape. M, Q, PMO 212.223 ⁄ 17, specimen with thin and wide mesh, showing distinct microsphere. Inanigutta magnifica n. sp. N, R, U,PMO 212.221 ⁄ 12, holotype, typical specimen with wide mesh formed from thick bars, two spheres are preserved, internal microsphereindistinct. S, T, PMO 212.222 ⁄ 14, specimen with coarse mesh, outer sphere (sphere 2) only preserved. Magnification for all specimensgiven by bar representing 100 lm.
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1195
B C D
100 µm
A
H G F
E
I
M
L K J
P Q R O
N
TEXT -F IG . 8 . Inanigutta penrosei group. A, B, PMO 212.225 ⁄ 18, ?microsphere with robust outer spines and apophyses. C, D, PMO212.223 ⁄ 09, sphere with outer spines and beams. E, PMO 212.226 ⁄ 06, sphere showing mesh, many beams are visible along with outerspines bearing apophyses. F, O, PMO 212.220 ⁄ 15. G, PMO 212.221 ⁄ 03. H, I, PMO 212.222 ⁄ 12. J-L, PMO 212.227 ⁄ 14. M, N, R, PMO212.226 ⁄ 04. P, Q, PMO 212.227 ⁄ 18, specimen showing indications of three spheres connected by beams and a few outer spines.Magnification for all specimens given by bar representing 100 lm.
1196 PALAEONTOLOGY, VOLUME 51
in their illustrations indicate that the material should beassigned to this form.
Family HAPLOTAENIATUMIDAE Won et al., 2002
Remarks. Won et al. (2002) established the family Hapo-taeniatumidae based on the presence of a ‘‘spongy’’ three-dimensional spherical shell. The species have one or morespongy shells, usually arranged in a spiral fashion. Theexact development of the spongy mesh from thin bars isnot known in most species. The internal developmentand the origin of the external spines are uncertain.Spongy or three-dimensionally developed shells are com-mon in Cambrian and Lower Ordovician radiolarian
faunas, but most may not be related to the Haplotaenia-tumidae. A spongy spherical body was developed in thePalaeospiculumidae and the Protoentactiniidae, based onbranching of the primary spicule or apophyses.
The proloculus of Won et al. (2002) and Jones andNoble (2006) may represent the microsphere, visible inother forms attributed to the Haplotaeniatumidae (Mac-Donald 2006; Maletz and Bruton 2007). Its developmenthas never been described in detail.
Genus Haplotaeniatum Nazarov and Ormiston, 1993
Type species. Haplotaeniatum labyrintheum Nazarov andOrmiston, 1993.
B C
D
100 µm
A
H
G F E
I L K J
TEXT -F IG . 9 . Inanigutta sp. B. A, B, PMO 212.221 ⁄ 14, specimen showing mesh development of outer sphere and irregular thirdsphere, inner sphere not preserved. C, PMO 212.220 ⁄ 06, sphere with outer spines and beams. D, H, PMO 212.225 ⁄ 13, specimens withstrong development of spines and beams. E, PMO 212.229 ⁄ 01, poorly preserved specimen with inner and outer sphere. F, G, PMO212.220 ⁄ 10, well preserved specimen with two distinct spheres, showing apparent origination of outer spines on inner sphere. I, L,PMO 212.223 ⁄ 04, small sphere with incomplete development of second sphere, both are closer together than usual. J, K, PMO212.222 ⁄ 10, specimen with numerous outer spines, tentatively referred to this species. Magnification for all specimens given by barrepresenting 100 lm.
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1197
Haplotaeniatum sp.
Text-fig. 7A–I, K, L, O, P
1971 Sphaeroids – apparently non-spiculate spumellariansFortey and Holdsworth, p. 39, pl. 11, figs. 9, ?10.
Material. 37 specimens.
Diagnosis. Labyrinthine spumellarians with a looselyspiral arrangement of the shell and a variable number ofslender outer spines.
Description. The diameter of the sphere is about 110–130 lmin mature specimens. The shape is often slightly irregular dueto the variable development of beams and bars. The speciesappears to bear an inconspicuous inner sphere of ca. 80 lmin diameter, possibly a microsphere, but specimens withoutany external bars were not found to confirm this indication.The smallest specimens (Text-fig. 7A, F) show the indistinctmicrosphere with a moderate number of outer spines andirregular bars indicating the three-dimensional, spiraliformmesh of the larger spherical body. The spiraliform develop-ment is clearly visible in a number of specimens (Text-fig. 7L), but is not invariably developed or observable in trans-mitted light. Some specimens appear to show a more concen-tric development with a number of indistinct spheres (Text-fig. 7H, I). The mesh forms from thin (2–3 lm wide), round,somewhat curved bars connecting the outer spines. Shorterbeams also develop from the inner microsphere and do notdevelop into outer spines, but end at the junction with thecurved bars. Bars often start from the outer spines and beamsand grow in various directions to connect to each other. Thelength of the outer spines, measured from the outer rim ofthe sphere is rarely more than 30 lm. The spines are thin andneedle-like.
Remarks. The species is common in the sample, but isoften difficult to differentiate from Inanigutta specimens.Inanigutta may be differentiated by a more regular devel-opment and mostly distinct shell, rarely showing moreirregular three-dimensional mesh that mimics a spiralarrangement. The two specimens illustrated by Fortey andHoldsworth (1971) may represent different growth stagesof this species.
Forms indet.
A number of rare specimens have been discovered thatcurrently cannot be identified, but are here illustrated forcompleteness. A number of poorly preserved specimensmay belong to Antygopora, but are not worth to beillustrated here. A single better-preserved specimen(Text-fig. 2E) shows a dense net of bandages with smallround to irregularly shaped pores. It is quite characteristic
and distinct, but a description is not warranted untilbetter material is available. A rare form with a thickspherical shell (Text-fig. 6F, G, L, M) has a diameter of100–130 lm. The surface appears to be highly irregularwith deep pits and some irregular pores. Unfortunately,the species is not known from SEM material yet. Thus, aclear understanding of the shell structure is impossible.Extremely rare are possible archaeoentactinids (Text-fig. 5N) and entactiniids (Figs. 5O, P).
Acknowledgements. The radiolarian samples were collected byR. A. Fortey (Fortey and Holdsworth 1971 material) and R. A.Fortey & DLB (Holdsworth 1977 material) and has since beencurated by F. J. Lindemann, collections manager, GeologicalMuseum, University of Oslo. C. E. Mitchell (University atBuffalo, SUNY, Buffalo, N.Y.) provided additional material fromthe Table Head Group of western Newfoundland. Peter Bush(South Campus Instrumentation Center, University at Buffalo,SUNY) provided access to SEM work.
REFERENCES
AITCHISON, J. C. 1998. A lower Ordovician (Arenig) radio-larian fauna from the Ballantrae Complex, Scotland. ScottishJournal of Geology, 34, 73–81.
ALBANI , R., BAGNOLI , G., MALETZ, J. and STOUGE,S. 2001. Integrated chitinozoan, conodont and graptolite bio-stratigraphy from the Upper Cape Cormorant Formation(Middle Ordovician), western Newfoundland. Canadian Jour-nal of Earth Sciences, 38, 387–409.
APPOLONOV, M. K., ZHEMCHUZHNIKOV, V. G. andDUBININA, S. V. 1990. Ordovician of the northwesternPribalkhash area. Izvestiya Akad. Nauk Kazakhskoy SSR, Ser.Geol., 4, 3–16.
BERGSTROM, S. M. 1979. First report of the enigmaticOrdovician microfossil Konyrium in North America. Journal ofPaleontology, 53, 320–327.
CALKINS, G. N. 1909. Protozoology. Lea and Febiger, NewYork, pp. 1–349.
COOPER, R. A. and FORTEY, R. A. 1982. TheOrdovician graptolites of Spitsbergen. Bulletin of the BritishMuseum, Natural History, London (Geology), 36, 157–302, pls1–6.
DANELIAN, T. and POPOV, L. 2003. Ordovician radiolarianbiodiversity: insights based on new and revised data fromKazakhstan. Bulletin de la Societe geologique de Franc, 174,325–335.
DE WEVER, P., DUMITRICA, P., CAULET, J. P., NIG-RINI , C. and CARIDROIT, M. 2001. Radiolarians in theSedimentary Record. Taylor and Francis, 533 pp.
DUMITRICA, P., CARIDROIT, M. and DE WEVER, P.2000. Archaeospicularia, ordre nouveau de radiolaires: unenouvelle etape pour la classification des radiolaires du paleo-zoique inferieur. Comptes rendues, Academie des Sciences, Paris,Sciences de la Terre et des Planete, 330, 563–569.
1198 PALAEONTOLOGY, VOLUME 51
EHRENBERG, C. G. 1838. Uber die Bildung der Kreidefelsenund des Kreidemergels durch unsichtbare Organismen.Konigliche Akademie der Wissenschaften zu Berlin, Abhandlun-gen, 1838, 59–147.
—— 1875. Fortsetzung der mikrogeologischen Studien alsGesamt-Uebersicht der mikroskopischen Palaontologie glei-chartig analysierter Gebirgsarten der Erde, mit speciellerRucksicht auf den Polycystinen-Mergel von Barbados. Ab-handlungen der Koniglichen Akademie der Wissenschaften zuBerlin, Jahrgang, 1875, 1–226.
FOREMAN, H. P. 1963. Upper Devonian Radiolaria from theHuronmember of the Ohio Shale.Micropaleontology, 9, 267–304.
FORTEY, R. A. 1974. The Ordovician trilobites of SpitsbergenI. Olenidae. Norsk Polarinstitutt Skrifter, 160, 1–129.
—— 1975a. Early Ordovician trilobite communities. Fossils andStrata, 4, 339–360.
——1975b. The Ordovician trilobites of Spitsbergen II. Asaphi-dae, Raphiophoridae and Telephinidae of the ValhallfonnaFormation. Norsk Polarinstitutt Skrifter, 162, 1–207.
——1980. The Ordovician trilobites of Spitsbergen III. Remain-ing trilobites of the Valhallfonna Formation. Norsk Polarinsti-tutt Skrifter, 171, 1–163.
—— and BRUTON, D. L. 1973. Cambrian-Ordovician rocksadjacent to Hinlopenstretet, north Ny Friesland, Spitsbergen.Bulletin of the Geological Society of America, 84, 2227–2242.
—— and HOLDSWORTH, B. K. 1971. The oldest knownwell-preserved Radiolaria. Bollettino della Societa PaleontologiaItaliana, 10, 35–41.
HOLDSWORTH, B. K. 1977. Paleozoic Radiolaria: strati-graphic distribution in Atlantic borderlands. 167–184. InSWAIN, F. M. (ed.). Stratigraphic micropaleontology of Atlan-tic basin and borderlands. Developments in Paleontology andStratigraphy 6. Elsevier, New York.
JONES , M. K. and NOBLE, P. J. 2006. Sheinwoodian (upper-most Lower Silurian) Radialaria from the Cape PhillipsFormation, Nunavut, Canada. Micropaleontology, 52 (4), 289–315.
KOZUR, H. W. and MOSTLER, H. 1982. Entactinaria su-bordo nov., a new radiolarian suborder. Geologisch-Palaonto-logische Mitteilungen Innsbruck, 11 (12), 399–414.
—— —— and REPETSKI , J. E. 1996. Well-preserved Trem-adocian Radiolaria from the Windfall Formation of the Ante-lope range, Eureka County, Nevada, USA. Geologisch-Palaontologische Mitteilungen Innsbruck, 121, 245–271.
MACDONALD, E. W. 2006. Haplotaeniatumidae and Inani-guttidae (Radiolaria) from the lower Silurian of the Cape Phil-lips Formation, Cornwallis Island, Nunavut, Canada. Journalof Paleontology, 80 (1), 19–37.
MALETZ, J. 2007a. The early Ordovician Beothuka terranova(Radiolaria) faunal assemblage in western Newfoundland.Palaontologische Zeitschrift, 81, 71–82.
——2007b. Overlooked and ignored: radiolarians and their bio-stratigraphical potential in the Ordovician. Acta Palaeontolog-ica Sinica, 46 (Suppl.), 291–297.
—— and BRUTON, D. L. 2005. The Beothuka terranova(Radiolaria) assemblage and its importance for the under-standing of early Ordovician radiolarian evolution. GeologicalMagazine, 142 (6), 711–721.
————2007. Early Ordovician (Chewtonian to Castlemainian)radiolarians of Spitsbergen. Journal of Systematic Palaeontology,5 (2), 245–288.
MULLER, J. 1858. Uber die Thalassicolen, Polycystinen undAcanthometren des Mittelmeeres. Abhandlungen der Preussis-chen Akademie der Wissenschaften Jahrgang, 1858, 1–62.
NAZAROV, B. B. 1975. Radiolaria of the lower – middlePaleozoic of Kazakstan. Trudy Akademiia NAUK SSSR, Geo-logic Institute, 275, 203 pp. (in Russian).
——1988. Paleozoic Radiolaria Practical manual of microfauna ofthe USSR. Volume 2: Leningrad, Nedra, 232 pp.
—— and ORMISTON, A. R. 1984. Tentative system of Paleo-zoic Radiolaria. 64–87. In PETRUSHEVSKAYA, M. G. andSTEPANJANTS, S. D. (eds). Morphology, ecology and evolu-tion of Radiolaria (Eurorad IV Symposium Volume, October1984). Zoological Institute, Leningrad, Nauka (in Russian withEnglish Abstract).
—— ——1986. Trends in the development of Paleozoic Radio-laria. Marine Micropaleontology, 11, 3–32.
—— —— 1993. New biostratigraphically important PaleozoicRadiolaria of Eurasia and North America. 22–60. In BLUE-FORD, J. R. and MURCHEY, B. (eds). Radiolaria of Giantand Subgiant Fields in Asia. Micropaleontology, Special Publi-cation, No. 6.
—— and POPOV, L. E. 1980. Stratigrafiya i fauna Kremnis-to – Karbonatnykh tolshch ordovika kazakhstana: radiolariii bezza mkovye brakhiopody (Stratigraphy and fauna of thesiliceous-carbonate sequence of the Ordovician of Kazakh-stan; radiolarians and inarticulate brachiopods. Trudy Geo-logicheskiy Institut Akademiy a Nauk SSR, 331, 190 pp. (inRussian).
NOBLE, P. J. 1994. Silurian radiolarian zonation for the Cabal-los Novaculite, Marathon Uplift, west Texas. Bulletin of Ameri-can Paleontology, 106, 1–55.
—— and AITCHISON, J. C. 1995. Status of Ordovician andSilurian radiolarian studies in North America. 19–30. InBLOOM, C. D. et al. (eds). Paleontological Society ShortCourses in Paleontology 9.
————2000. Early Paleozoic radiolarian biozonation. Geology,28, 367–370.
RENZ, G. W. 1990. Ordovician Radiolaria from Nevada andNewfoundland – a comparison at the family level. MarineMicropaleontology, 15, 393–402.
RIEDEL, W. R. 1967. Some new families of Radiolaria.Proceedings of the Geological Society of London, 1640, 148–149.
RUEDEMANN, R. 1947. Graptolites of North America. Geo-logical Society of America Memoir, 19, 1–652.
—— and WILSON, T. Y. 1936. Eastern New York OrdovicianCherts. Bulletin of the Geological Society of America, 47, 1535–1585.
RUSHTON, A. W. A., STONE, P., SMELLIE , J. L. andTUNNICLIFF , S. P. 1986. An early Arenig age for the Pinb-ain sequence of the Ballantrae Complex. Scottish Journal ofGeology, 22, 41–54.
STONE, P. and RUSHTON, A. W. A. 1983. Graptolite faunasform the Ballantrae Ophiolite Complex and their structuralimlications. Scottish Journal of Geology, 19, 297–310.
MALETZ AND BRUTON: PROVENTOCITUM PROCERULUM RADIOLARIAN ASSEMBLAGE OF SPITSBERGEN 1199
—— and SMELLIE , J. L. 1988. The Ballantrae area: descriptionof the solid geology of parts of 1:25.000 sheets NX08, 18 and 19.British Geological Survey London, United Kingdom.
STOUGE, S. S. 1984. Conodonts from the Middle OrdovicianTable Head Formation, western Newfoundland. Fossils andStrata, 16.
WANG, Y. J. 1993. Middle Ordovician radiolarians from thePingliang Formation of Gansu Province, China. Micropaleon-tology Special Publication, 6, 98–114, pls. 1–10.
WILLIAMS, S. H. and STEVENS, R. K. 1986. Summaryaccount of the Lower Ordovician (Arenig) graptolite biostra-tigraphy of the Cow Head Group, western Newfoundland.Bulletin of the Geological Society of Denmark, 35, 259–270.
—— —— 1988. Early Ordovician (Arenig) graptolites of theCow Head Group, western Newfoundland, Canada. Palaeonto-graphica Canadiana, 5, 1–167.
WON, M.-Z. and BELOW, R. 1999. Cambrian Radiolariafrom the Georgina Basin, Queensland, Australia. Micropaleon-tology, 45, 325–363.
——BLODGETT, R. B. and NESTOR, V. 2002. Llandoverian(early Silurian) radiolarians from the Road River Formationof east-central Alaska and the new family Haplotaeniatumidae.Journal of Paleontology, 76, 941–964.
—— and IAMS, W. J. 2002. Late Cambrian radiolarian faunasand biostratigraphy of the Cow Head Group, western New-foundland. Journal of Paleontology, 76, 1–33.
———— and REED, K. 2005. Earliest Ordovician (early to mid-dle Tremadocian) radiolarian faunas of the Cow Head Group,western Newfoundland. Journal of Paleontology, 79, 433–459.
ZHANG, X.-G. and PRATT, B. R. 2000. A varied MiddleOrdovician sponge spicule assemblage from western New-foundland. Journal of Paleontology, 74, 386–393.
APPENDIX 1
Kazakhstan sample 19 (Nazarov and Popov 1980; Danelian andPopov 2003)
Locality: North Balkhash Region, locality ca. 4 km southeastof Sarykum railway station, about 70 km west of the town ofBalkash. Sample comes from the bedded argillaceous limestonesat the lower part of the Kurchilik Formation.
Fauna (original faunal list in Nazarov and Popov 1980, p. 7).Revision and remarks by Jorg Maletz (February 2007).Astroentactinia? miscella Nazarov in Nazarov and Popov 1980.
Sphere diameter ca. 200 lm, mesh with round pores indicatea possible relationship to Antygopora; many outer spines, atleast ca. 20; nothing more is known from the only illustratedspecimen. Similarities also exist to Inanigutta sp. B.
Entactinia hirsuta Nazarov in Nazarov and Popov 1980;. Poorlyillustrated by two photos in reflected light and drawings.Diameter of sphere ca. 230–280 lm; many relatively shortouter spines and possibly beams; mesh appears to be formedfrom bars, development unclear; three-dimensional mesh sur-rounding sphere; Nazarov (in Nazarov and Popov 1980, p. 33,Text-Fig. 13a) indicated an internal spicule; relationshipuncertain.
Entactinia sp. unclear identification, poor transmitted lightphoto, appears to have 6–8 outer spines and numerous beams.
Entactinosphaera? hirta Nazarov in Nazarov and Popov 1980;.fairly large form, inner spicule shown by Nazarov and Popov(1980), Text-Fig. 18b as a drawing, not verified from material.May just be an interpretation from the position of outerspines; if there are two inner spheres instead, it might be simi-lar to some of the Spitsbergen specimens (Inanigutta sp. B), inwhich the outer shell is not preserved or constructed.
Entactinosphaera sp., belongs to the Inanigutta penrosei group withlong and stout outer spines, one small inner sphere is visible.
Haplentactinia(?) armillata Nazarov in Nazarov and Popov 1980.Illustrated by SEM and reflected light, internal structure andrelationship uncertain, characteristic are branched outer spines.
Inanihella bakanasensis (Nazarov 1975), originally Helioentacti-nia? bakanasensis. Type species of Inanihella; two shells withlittle distance between and numerous beams, possible lownumber of outer spines, needs description. Danelian and Po-pov (2003) re-described the species and illustrated it withSEM photos. Similarities exist to Nyfrieslandia, but identifica-tion is unclear.
Helioentactinia sp. unclear, unidentifiable at the moment.Proventocitum procerulum (Nazarov in Nazarov and Popov
1980), well known and illustrated by Nazarov and Popov (1980).Anakrusia miriacantha Nazarov – sponge spicule.