Since the introduction of genetically modified (GM) crops in the mid-1990s, there has been a rapid adoption of the tech-nology and the global acreage of GM crops reached a record 125 million hectares (309 million acres) in 25 countries in 2008.1 The country with the largest area of GM crops (half of the world’s total acreage) is the United States of America (USA), followed by Argentina, Brazil, Canada, India and China. Currently, the major GM field crops are soybean, corn, cotton and canola. Minor GM crops include papaya, sugar-beet, squash, potato and alfalfa. The most widely adopted GM traits are herbicide tolerance and insect resistance. These traits provide growers with benefits of increased yield, reduced insecticide use and simplified management of weed control with fewer and more flexible herbicide applications. Herbicide (glyphosate) tolerance is the most widely used trait and has the largest acreage in soybean, corn, cotton and sugarbeet, fol-lowed by traits for insect resistance in corn and cotton. Other
*Correspondence to: Qiudeng Que; Email: [email protected]: 06/25/10; Revised: 08/25/10; Accepted: 08/27/10Previously published online:www.landesbioscience.com/journals/gmcrops/article/13439DOI: 10.4161/gmcr.1.4.13439
In recent years, there has been a rapid increase in the planting of transgenic crops with stacked traits. Most of these products have been formed by conventional breeding, i.e., the crossing of transgenic plant (event) containing individual transgenes with other event(s) containing single or double transgenic traits. Many biotech companies are developing stacked trait products with increasing numbers of insect and herbicide tolerance genes for controlling a broad range of insect pests and weeds. There has also been an increase in development of technologies for molecular stacking of multiple traits in a single transgene locus. In this review we look at the status of stacked trait products, crop trait stacking technologies and the technical challenges we are facing. We also review recent progress in developing technology for assembling large transgene arrays in vitro (molecular stacks), their delivery to crop plants and issues they pose for transgene expression.
Trait stacking in transgenic cropsChallenges and opportunities
Qiudeng Que,* Mary-Dell M. Chilton, Cheryl M. de Fontes, Chengkun He, Michael Nuccio, Tong Zhu, Yuexuan Wu, Jeng S. Chen and Liang Shi
Syngenta Biotechnology, Inc.; Research Triangle Park, NC USA
Key words: trait stacking, insect resistance, herbicide tolerance, molecular stack, large DNA transformation, mini-chromosome, targeted integration, transgene expression
transgenic traits include virus resistance, male sterility and oil quality. In 2010, 86, 93 and 93% of corn, cotton and soy-bean, respectively, planted in the US contained at least one biotech trait (www.ers.usda.gov/Data/BiotechCrops). Until the introduction of glufosinate tolerance varieties recently, all GM soybean varieties planted in the US contained only one single trait, i.e., glyphosate tolerance. In corn and cotton, there is still significant acreage with a single herbicide tolerance or insect resistance trait. However, the percentage of products with stacked traits (herbicide tolerance and insect resistance) has increased rapidly, reaching 47% in corn and 58% in cotton in 2010 (www.ers.usda.gov/Data/BiotechCrops).
Current Stacked Trait Products and Trends
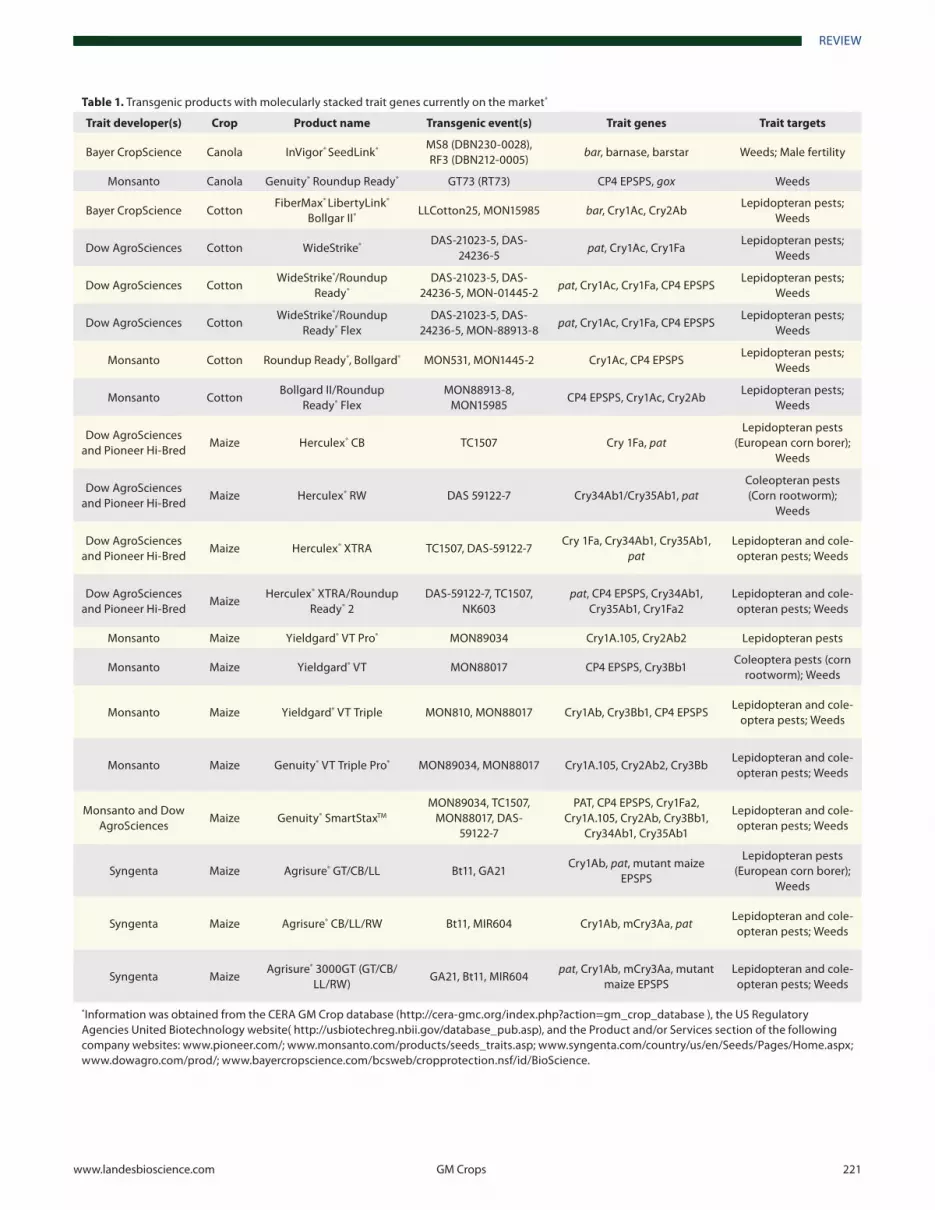
The insect resistant trait based on Bacillus thuringiensis (Bt) toxin is becoming an indispensible tool in modern high effi-ciency agriculture. In order for this trait to be used sustainably, it is important to prevent the emergence and buildup of insects resistant to Bt toxins. To help manage insect resistance, the first Bt crops introduced in the USA were required to have host ref-uge areas without the trait (www.epa.gov/oppbppd1/biopesti-cides/pips/regofbtcrops.htm). In the USA, the Environmental Protection Agency (EPA) required that at least 20% of a grower’s corn acres be planted with non-Bt corn as a refuge area. In cot-ton-growing regions, growers can only plant up to 50% of their corn acreage with Bt corn borer resistant hybrids since maize is also an alternate host for cotton bollworm (www.epa.gov/oppb-ppd1/biopesticides/pips/bt_brad2/4-irm.pdf). If the refuge area could be decreased from the standard 20 to 5%, a signifi-cant production benefit would be realized from this additional 15% Bt crop. Therefore, it makes sense to develop alternative approaches to preventing target pests from developing resistance. Recently, the industry has rapidly moved in the direction of pro-viding two or more modes of action for the control of major insect pests, thus reducing the chance for resistance to develop and decreasing the percentage needed for insect refuges. Table 1 lists some products with stacked traits currently on the mar-ket. Additional information of these transgenic events and other transgenic events in non-stacked products not listed in the table can be found in the CERA database.2 In the US, information on genetically engineered crop plants intended for food or feed that
www.landesbioscience.com GM Crops 221
REVIEW REVIEW
Table 1. Transgenic products with molecularly stacked trait genes currently on the market*
*Information was obtained from the CERA GM Crop database (http://cera-gmc.org/index.php?action=gm_crop_database ), the US Regulatory Agencies United Biotechnology website( http://usbiotechreg.nbii.gov/database_pub.asp), and the Product and/or Services section of the following company websites: www.pioneer.com/; www.monsanto.com/products/seeds_traits.asp; www.syngenta.com/country/us/en/Seeds/Pages/Home.aspx; www.dowagro.com/prod/; www.bayercropscience.com/bcsweb/cropprotection.nsf/id/BioScience.
222 GM Crops Volume 1 Issue 4
(featuring the single mode of action Herculex® RW) offers a RIB option for corn rootworm resistance management but still requires growers to plant a separate 20% corn borer refuge (farmindustrynews.com/seed/0201-acremax-technology/).
The emergence of glyphosate tolerant weeds in many regions of the world will likewise require the use of additional and/or alternative herbicide tolerance traits for complete control of weeds in the future, especially in dicot crops such as canola, soybean and cotton that have fewer available effective herbicide options. It has been reported that several companies are currently devel-oping alternative non-glyphosate weed control technologies.3 For example, in collaboration with the University of Nebraska, Monsanto is developing dicamba-tolerant soybean for control of glyphosate-resistant weeds using the dicamba mono-oxygenase (DMO) gene. DuPont is developing a dual herbicide resistance to both acetolactate synthase (ALS) inhibitors and glyphosate for use in corn and soybean.3 Dow AgroSciences is developing a soybean trait for resistance to 2,4-dichlorophenoxyacetic acid (2, 4-D) and pyridyloxyacetate herbicides based on the AAD-12 (aryloxyalkanoate dioxygenease) gene.3 Bayer CropScience is developing soybean traits for resistance to glufosinate (Liberty herbicide) and HPPD (4-hydroxyphenolpyruvate dioxygenase) inhibitors such as isoxaflutole.3 Syngenta has also announced the development of HPPD tolerant soybeans (native trait fol-lowed by GM) (www2.syngenta.com/en/investor_relations/pdf/Syngenta_SRI_presentation.pdf). Soybean varieties toler-ant to the imidazolinone class of herbicides developed by BASF and EMBRAPA (Empresa Brasileira de Pesquisa Agropecuária) received regulatory approval for commercial cultivation in Brazil recently (www.basf.com/group/pressrelease/P-10-148).
The gene stacking situation in corn is becoming complex. In the US, at least 8 genes will be needed in corn to provide for combined weed control and at least two modes of action for controlling four major pests: Ostrinia nubilalis (European corn borer, ECB), Helicoverpa zea (corn earworm, CEW), Diatraea grandiosella (Southwestern corn borer, SWCB) and Diabrotica virgifera (western corn rootworm, WCR) in the mid-west corn growing region. In addition to the above, several other insect pests, Agrotis ipsilon (black cutworm), Loxagrotis albicosta (west-ern bean cutworm), Spodoptera frugiperda (Fall armyworm, FAW) and Diatraea saccharalis (sugarcane borer) also cause serious leaf, stock or ear damages. The complete control of these corn pests may require use of an additional insecticidal gene such as VIP3A. In addition to the herbicide tolerance and insect resistance traits, there are several agronomic and quality traits that the agricultural biotechnology industry is currently developing, including yield enhancement, drought tolerance, nitrogen utilization efficiency, disease resistance, fertility control (male sterility), grain quality (amino acid composition, protein content and oil composition) and grain processing (phytase for animal feed and amylase for corn ethanol). Thus the number of trait genes that need to be transformed into corn could easily add up to 15 or more, assum-ing that most traits can be achieved with only one or two genes. In addition, multiple genes will probably be needed for robust per-formance of complex agronomic traits such as yield enhancement and drought tolerance. In the past, transgenic events were created
have completed all recommended or required reviews for plant-ing, food or feed use can be found in the National Biological Information Infrastructure (NBII) database (usbiotechreg.nbii.gov/database_pub.asp). The NCGA database also has the lat-est up-to-date listing of the available maize biotech traits and their Japan and EU approval status (www.ncga.com/know-you-grow). In the past few years, the US EPA approved reduction of the refuge areas for cotton and corn products with dual Bt genes with different modes of action. For example, Syngenta’s Agrisure® VipteraTM 3111GT (www.agrisureviptera3111.com/) with two lepidopteran active proteins (Cry1Ab and Vip3A) and one coleopteran active protein (mCry3A) qualifies for a reduced corn borer refuge in cotton growing regions from 50 to 20%. Monsanto’s GenuityTM YieldGard® VT ProTM (with Cry1A.105 and Cry2Ab) and Triple Pro® (with Cry1A.105, Cry2Ab and Cry3Bb) hybrids also qualify for a reduced corn borer refuge in cotton growing regions from 50 to 20% because they have two corn borer traits (www.genuity.com/Traits/Corn/Genuity-VT-Triple-PRO.aspx). Monsanto’s GenuityTM Bollgard II® cotton (with Cry1Ac and Cry2Ab) and Dow AgroSciences’ WideStrike® Insect Protected cotton (with Cry1Fa and Cry1Ac) also use the “natural refuge” option relying on surrounding fields with non-cotton hosts in certain areas to replace the once required non-Bt cotton refuge (www.epa.gov/oppfead1/cb/csb_page/updates/2007/bollgard-cotton.htm). Also, the US EPA has approved a natural refuge option for Syngenta’s VipCotTM cotton which produces both Cry1Ab and Vip3A proteins (news.agropages.com/News/NewsDetail---2482.htm). In 2009, Monsanto and Dow AgroSciences received EPA approval for the SmartStaxTM hybrid corn product with eight different genes for herbicide tolerance and insect-protection (www.epa.gov/oppb-ppd1/biopesticides/pips/smartstax-factsheet.pdf). SmartStaxTM is created by breeding crosses bringing together several previ-ously approved transgenic events including Dow AgroSciences’ Herculex® I (Cry1Fa) and Herculex® RW (Cry34Ab and Cry35Ab) technologies, Monsanto’s GenuityTM YieldGard® VT Rootworm (Cry3Bb)/RR2 and GenuityTM YieldGard® VT PRO® (Cry1A.105 and Cry2Ab) technologies, and the two weed control technologies, Roundup Ready 2® (RR2) based on the 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene from Agrobacterium strain CP4 and LibertyLink® based on the phosphinothricin phosphotransferase (PAT) gene from Streptomyces. The use of dual modes of control for each pest in SmartStaxTM allowed for the refuge requirement to be reduced from 20 to 5% in the US Corn Belt (www.epa.gov/pesticides/biopesticides/pips/smartstax-factsheet.pdf). Syngenta Seeds, Inc., has submitted to the US EPA an application for the reg-istration of its Agrisure® VipteraTM 3,220 trait stack, also fea-turing two modes of action against all major lepidopteran corn pests and requesting a reduced refuge of 5% for the US Corn Belt (agrisureviptera.com/NewsDetail.aspx?NewsID = 118). Another development in the marketplace for managing emergence of resistance is the Refuge in a Bag (RIB) concept, which simplifies the refuge management by incorporating non-GM seeds or seeds without the target insect traits in the Bt seed bag directly. Pioneer Hi-Bred’s Optimum® AcreMax 1TM
www.landesbioscience.com GM Crops 223
transgenic events. Usually, combining several loci carrying sim-ple traits together results in products with good trait efficacy. In some countries, no additional regulatory approval is required for commercializing breeding stacks. Even when a new approval is needed, the approval of breeding stacks of existing traits is usu-ally faster than a new trait event since all traits in the breed-ing stack are previously well characterized. Breeding stacks are also flexible: the trait offering can be tailored to specific markets where the insect pest spectrum might be different, thus requir-ing different trait stacks. However, combination of several trait loci is very time consuming and expensive because of the num-ber of back-crosses that are needed to convert hundreds of elite lines for commercial launch in each crop. When breeding for trait stacks containing two or three loci, it is convenient to use a linked herbicide tolerance trait to identify desired progeny containing the intended trait combinations. However, since the number of herbicide tolerance traits is limited, introgression of multiple loci must be achieved in part by use of more expensive molecular (either DNA or protein) analysis tools. The problem becomes more complicated when multiple third party traits are combined together, because the genetic background could be very different among the donor lines. Seed production of varieties with mul-tiple trait loci is also hard to track from the perspective of quality management and product stewardship. Potential erroneous label-ing or handling of parent lines may result in cross-contaminated seed lots. Currently, from a practical point of view, at least two loci are needed for regulatory or technical reasons. For example, if the regulatory agency requires refuge planting for corn hybrids with insect resistance traits, the refuge plants will probably still require herbicide tolerance for weed management. Thus, it would be convenient to separate traits into an herbicide tolerance trait stack on one inbred and an insect resistance trait stack on a sepa-rate inbred. Inbreds containing each of these separate stacks can be generated individually.
Improving Tools to Deliver Large Multi-Gene Stack Vectors
Generation of molecular stack of multiple traits presents several challenges, including cassette design, vector assembly, transfor-mation and gene expression analysis. Many trait genes in the cur-rent approved products are under the control of a few frequently used promoters such as Cauliflower Mosaic Virus (CaMV) 35S, maize ubiquitin-1 and rice actin-1. For successful molecular stacking it will be necessary to increase the number of available regulatory elements for trait development. Despite a reasonable number of available promoters, there are very few studies describ-ing how they work when combined at a single locus.9 It is also important to understand how other regulatory elements such as enhancers and insulators interact when placed close to each other, and to analyze the impact of the overall chromatin environment on their activity. Routine recombinant DNA technology depends on restriction enzymes for DNA manipulation. Currently, the main workhorse for most laboratories is recombinant DNA technology based on restriction endonucleases and DNA liga-tion, as well as recombinase-mediated assembly. However, as the
by transforming constructs carrying one or two trait genes indi-vidually into plants. Events with the required level of efficacy in the field were generated relatively easily in some cases. Products with single or double traits provide flexibility in trait combina-tions for each region. However, the effort required to bring a large number of trait loci into multiple cultivars even in a single crop quickly becomes unmanageable if each transgenic locus only car-ries one or two traits. From a scientific and breeding point of view, it is very desirable to deliver several traits simultaneously in a sin-gle locus. Delivery of multiple trait genes in one or more pieces of recombinant DNA simultaneously or consecutively into a single transgenic locus is called molecular stacking of traits. Trait molec-ular stacks are tightly linked and exhibit an extremely low rate of segregation, essentially behaving as a single gene, thus making trait introgression and line conversion much simpler.
Current Trait Combination Tools: Breeding Stacks and Molecular Stacks
Molecular stacking of two to three gene expression cassettes is straightforward. Each of these expression cassettes can contain a gene encoding a natural or mutant protein or an engineered chi-meric protein or protein fusion with multiple functions such as dual Bt that can control a broad spectrum of insects. Many of the first generation trait events were generated in this fashion to con-tain stacks of both herbicide tolerance and insect resistance traits, such as Syngenta’s maize Agrisure® event Bt11 which contains a PAT gene expression cassette conferring glufosinate tolerance and a Cry1Ab gene expression cassette conferring European corn borer resistance.2 Currently, the following maize events with dou-ble trait stacks are still on the market (see Table 1): Syngenta’s Bt11 with PAT and Cry1Ab, Dow AgroSciences’ TC1507 with PAT and Cry1Fa, Monsanto’s MON 89034 with Cry1A.105 and Cry2Ab, and Dow AgroSciences’ DAS 59122-7 Herculex® RW event with PAT, Cry34Ab and Cry35Ab. However, as the number of traits increases, the assembly of each stack becomes more cum-bersome. More importantly, the expression of each gene and effi-cacy of each trait may become less predictable.4,5 Co-expression can be achieved by using a linker peptide to make a translational fusion of two different trait genes products.6,7 In tobacco plastids, multiple open reading frames can be placed under the control of a single promoter and be transcribed as an operon, produc-ing multiple separate proteins. Tobacco plastids have also been engineered with vectors carrying multiple genes.8 However, the plastids of major field crops like maize, cotton, canola and soy-bean are not routinely transformed and no commercial plastid trait product is on the market yet. It remains to be seen whether there will be a breakthrough in transformation technology that will allow routine plastid transformation for trait development in major crops. Therefore, plastid transformation technology will not be discussed further here.
All of the current multi-trait stack products on the market are derived by combining previously existing transgenic events by traditional breeding methods; thus they are called breeding stacks. Breeding stacks have the advantage of flexibility. One can utilize previously tested, approved and commercialized
224 GM Crops Volume 1 Issue 4
absolute requirement for obtaining tomato transformants with the BIBAC system.20 Similar to BIBAC, Liu et al.25 reported con-struction of TAC (transformation-ready artificial chromosome) vectors based on P1 phage origin of replication. T-DNA regions of TAC vectors with 40 to 80 kb genomic DNA fragments have been transferred back into Arabidopsis with high efficiency and the transgenes were shown to be inherited faithfully by the prog-eny. Both BAC and TAC vectors contain very low copy bacte-rial origins of replication (f1, P1 and Ri) which allow the insert to be more stable in both E. coli and Agrobacterium. However, BAC and TAC vectors may not be essential for large T-DNA con-struction and delivery as long as the inserts can be stably main-tained in the E. coli and Agrobacterium cells. It has been reported that conventional binary vectors such as one based on the RK2 origin of replication can also be used for construction of large T-DNA vectors.26 Launching of T-DNA from the Agrobacterium chromosome has also been tested for delivery of transgene.27 This design, which minimizes copy number in Agrobacterium, results in a high percentage of single copy plant transformants with intact T-DNA, although the overall transformation frequency is lower compared to binary vectors.27 Delivering one long T-DNA from the Agrobacterium chromosome might be a good alternative to the use of several smaller binary vectors. However, the additional step of inserting T-DNA into the Agrobacterium chromosome in addition to the reduced transformation efficiency make the method more cumbersome in practice.
The molecular stacking approach has been studied with an increasing number of genes assembled into a single plant trans-formation construct.9 Downstream breeding effort will be greatly simplified if multiple traits with different modes of action can be carried in a single transformation vector, for example, an ECB and CRW stack with Cry1Ab, Cry1Fa, VIP3A, mCry3Aa, Cry34Ab and Cry35Ab along with EPSPS conferring glypho-sate tolerance. However, this kind of multi-cassette vector would require coordinated expression of all 7 trait genes in either con-stitutive and tissue-specific fashions. In molecular stacking, trait genes controlled by promoters with different tissue speci-ficities must be placed in close proximity, raising the risk that there might be interaction between different promoter regulatory sequences. This is especially of concern when long-range-active viral enhancer elements such as those from the CaMV 35S pro-moter are used, because they may affect expression of several genes in the array. Encouragingly, the few examples in the litera-ture suggest that molecular stacking of trait genes may not be so formidable. Cao et al.28 have transformed rice plants by T-DNA delivery and expressed several trait genes including herbicide tol-erance and yield enhancement from a ~20 kb 5 gene array under the control of different promoters. Fujisawa et al.29 have described coordinate expression of seven trait genes to enhance carotenoid synthesis in Brassica napus. In this study trait genes were intro-duced on a single ~17 kb T-DNA. Recently it was shown that when a large 32 kb T-DNA vector with 10 genes was introduced into an elite maize line, most trait genes could be expressed well based on ELISA data and lepidopteran and coleopteran insect bioassay results.30 In this study, it was shown that even though transformation frequency of the large 32 kb T-DNA vector by
number of components and size of DNA constructs increase, it becomes more difficult to find unique restriction sites for tailor-ing DNA manipulation. Even though assembly of 8–10 gene expression cassettes onto one T-DNA can be done with careful vector design, detailed planning and use of rare cutting endo-nucleases, application of new tools would be very helpful. Novel restriction endonucleases capable of recognizing and cleaving long sequences are extremely useful for assembly of complex DNA structures. Recently, it has been demonstrated that novel mega-endonucleases and zinc finger nucleases can be custom-designed to allow digestion of specific sequences.10,11 Dafny-Yelin and Tzfira have reviewed the different approaches for assembling multigene vectors, including traditional cloning methods, rare cutting endonucleases, artificial restriction enzymes and recom-binases.12 Encouragingly, recent advances in DNA synthesis technologies may greatly facilitate vector assembly by provid-ing genes with unwanted restriction sites removed and avoiding sub-cloning steps. Advances in DNA synthesis may make it cost-effective to synthesize an entire gene vector and replace routine vector construction and assembly.13 Enzymatic assembly of DNA molecules up to several hundred kilobase (kb) pairs has been demonstrated.14 This kind of large DNA assembly method will be needed for efficient construction of transformation vectors containing large native genomic regions, metabolic pathways and whole redesigned organelle genomes and mini-chromosomes. For further reading on the different aspects of synthetic biology, the reader is encouraged to explore other articles in the same focus issue as for the above ref. 13.
Large multi-gene constructs have been transformed into plants using both Agrobacterium-mediated and physical deliv-ery methods. The upper size limit (if any) for biolistic deliv-ery is unknown. Biolistic bombardment of plant cells has been used to deliver large DNA molecules such as bacterial artificial chromosomes (BACs), yeast artificial chromosomes (YACs) and minichromosomes.15-18 Micro-projectile bombardment-mediated transformation often results in insertion of truncated and/or concatenated plasmid DNA. However, biolistic delivery of a linear construct with no vector backbone has been shown to produce a high percentage of events with intact single copy insertions.19 Use of lower DNA concentrations along with care-ful handling of donor DNA have led to successful biolistic trans-formation and regeneration of maize plants containing a circular minichromosome.17
For Agrobacterium-mediated gene delivery, special binary vectors with low copy bacterial origins of replication have been designed for introducing large T-DNA inserts, especially for genomic DNA containing repetitive sequences. The Binary Bacterial Artificial Chromosome (BIBAC) vector system has been used successfully for transformation of intact high molec-ular weight DNA into plant chromosomes in several species including Arabidopsis, tobacco, tomato and rice.20-23 The BIBAC vector has been reported to deliver at least 150 kb of intact T-DNA to the tobacco nuclear genome. The efficiency of that process was found to be enhanced by the presence of additional Agrobacterium tumefaciens virulence genes.24 In fact, a helper plasmid containing extra copies of virG was found to be an
www.landesbioscience.com GM Crops 225
homing endonucleases or custom-designed zinc-finger nucleases. Site-specific recombinases such as Cre, Flp and λ integrase have been demonstrated to mediate recombination and integration of transgene sequences in important crops like tobacco, rice, soy-bean and maize.36-39 Homing endonucleases are found in nature and are involved in intron homing, a gene conversion process initiated by nuclease-mediated homing site cleavage. More than a dozen well-characterized homing endonucleases have been isolated from different organisms, including bacteria, algae and fungi. Their specificity can also be changed by mutagenesis of the amino acid sequences recognizing the DNA target sequence.10,40 There have been several reports on the use of homing endonu-cleases I-SceI and I-CeuI for directing targeted insertion in dif-ferent plants including tobacco, rice and maize via homologous recombination41-43 or non-homologous end-joining.44 One poten-tial drawback of using the site-specific recombinases and hom-ing endonucleases for site-specific insertion is the need to first introduce a target recognition sequence into the genome before targeted insertion can be carried out efficiently.
In contrast, zinc-finger nucleases (ZFNs) can be designed to cut any desired region of the natural or GM genome of the target plant. ZFNs are engineered chimeric nucleases created by fusing the zinc-finger DNA binding domain with the nucle-ase domain of the type IIs restriction endonuclease FokI.45 The DNA sequence specificity can be altered by changing the zinc-finger protein sequences. ZFNs recognizing specific transgenic and endogenous sequences have also been used to make ds-DNA breaks to direct gene targeting in several plant species, including Arabidopsis, tobacco and maize, and by using different transfor-mation techniques including electroporation, Agrobacterium and silicon carbide whiskers.46-50 Since the binding specificity of the ZFN can be altered to recognize the sequence of interest directly, this technology has the potential to create insertion into any desired location in the genome. For trait development, a remain-ing question is how to determine the best chromosomal location for inserting potential genes of interest. We must also investigate whether stacked trait genes will be expressed as expected to give the desired trait efficacy. One straightforward approach would be to insert new genes into previously characterized loci such as sites of transgene insertion in existing commercial products. Such a strategy has been proposed as an example for converting a commercial variety for transgene stacking.51 Since the tools of
Agrobacterium was much lower, many events containing intact T-DNA could be recovered.
Because the delivery of large T-DNA molecules has resulted in decreased transformation efficiency, it may be more desir-able to generate several smaller molecular stacks from a subset of similarly expressed traits and deliver them in combinations, rather than putting all traits together. Smaller stacks of a subset of traits provide flexibility when combining different traits for product packages to fit the needs of different regions and cus-tomers and in response to the changing regulatory and business environment. These separate T-DNAs can also be delivered at the same time during transformation using either Agrobacterium or biolistic delivery to generate transgenic loci with co-integrated DNA. Such co-integration of transformed DNA molecules has been reported in many co-transformation studies. Komari et al.31 generated co-integrated T-DNA events and also segregating T-DNA events with two different T-DNAs harbored in one or two Agrobacterium strains. McCormac et al.32 also showed that by using a binary plasmid with three selectable T-DNA regions, the frequency of co-integration could be as high as a third within a T
0 subpopulation of co-transformants. Chen et al.33 also co-
delivered 14 different plasmid DNAs into rice embryogenic tissues using biolistic bombardment, and found that many trans-genic plants contained co-integrated DNA molecules. Zhu et al.34 described similar work in maize but individual trait genes were co-bombarded to create a library of transgenic events. Their screen identified several independent events containing func-tional copies of all five trait genes. The reader is also encouraged to read a recent review by Naqvi et al.35 on different options for multigene transfer, especially concerning their application for metabolic pathway engineering.
New Technologies for Delivering Molecular Stacks
In the last decade there has been substantial progress in the development of technologies allowing creation of transgenic crop plants with multiple gene stacks, especially in large DNA deliv-ery, site-specific integration and mini-chromosome technologies. Table 2 lists some of the technology providers that are working on the trait stacking technologies. Site-specific integration can be mediated by site-specific recombinases or double stranded (ds)-DNA breaks created by rare-cutting endonucleases such as
Table 2. Developers of novel trait gene stacking technologies
Developer Technology AgBiotech partnership and licensees* Web site
*From the announcements found in individual developer company’s website.
226 GM Crops Volume 1 Issue 4
are usually expressed constitutively. Even though it has been shown possible to use the same promoter(s) to control several genes in pathway engineering studies,29,34,55 it is not known how stable the expression is over generations and whether gene silenc-ing will occur. Since repeated sequences in a transgene array may increase the chance of homology-dependent gene silencing, it would be desirable to avoid using the same promoter repeatedly in a transformation vector. However, the availability of suitable promoters is limited for practical reasons.9 For example, if it is desirable that certain Bt genes not be expressed in corn pollen, one hurdle is that there is a limited selection of strong promot-ers that have this attribute. Also, it is not well understood what mechanisms contribute to the expression variation between dif-ferent transgenic plants that contain the same gene cassettes, even though it is generally assumed that both the position effect from the insertion site’s neighboring chromosome environment and the transgene’s epigenetic modification such as methylation can be factors. For example, it has been reported that the intra-transformant variability of reporter gene expression could be as high as the inter-transformant variation in comparisons between single copy transformants.56 It has also been shown that site-specific integration of a reporter gene into the same locus can produce alleles that express at a predictable level or alleles that are differentially silenced.57 This effect is depending on the tar-get site of the insertion, but correlates with the amount of DNA methylation of the introduced DNA, presumably by the host cell during the transformation process.57 Thus, there is still much to be learned about the epigenetic control of gene expression in transgene stacking. In her earlier gene stacking review Halpin4 pointed out the challenges associated with multi-gene expression and manipulation in plants. Five years later, many of these same challenges on transgene expression remain despite considerable progress in assembling and delivering transgenes.5 Even for trans-formation vectors containing only one or two traits, it is well-known that many transformants must be generated to identify a few lead events for commercial development. The probability of getting a commercially acceptable event will decrease as the number of trait gene cassettes increases, especially if the stacked traits require very different regulatory sequences and have dif-ferent expression patterns, for example a molecular stack with a lepidopteran insect resistance trait and a rootworm control trait. For trait efficacy, it is probably easier to generate a commercially viable event with a stack containing only a subset of trait genes such as a broad lepidopteran insect control stack of Cry1Ab, Cry1Fa and Vip3Aa that requires similar expression pattern, ver-sus a large combination stack with multiple herbicide tolerance genes and several kinds of insect resistance traits under the con-trol of promoters with different tissue specificities.
In plants, enhancers present in promoters are capable of changing the activity and specificity of nearby promoters.58,59 For example, the CaMV 35S enhancer can affect expression of genes located over 78 kb away.60 This kind of long distance effect of regulatory elements on gene expression can potentially result in problematic expression of neighboring genes in a molecular stack event because strong promoters are frequently used to drive constitutive trait gene expression to ensure insect resistance and
site-specific insertion are now becoming a reality, these questions can be answered in the near future. In addition, rapid develop-ment in crop genome sequencing and gene expression profiling will also be very useful in helping us to understand and predict suitable chromosomal locations for a particular group of trait genes.
Recently, it has been shown that engineered mini-chromosomes can be formed either by deletion of non-essential portions of the supernumerary maize B-chromosomes52 or by in vitro assem-bly from isolated centromere-like sequences.17,18 Carson et al.17 reported the in vitro assembly of artificial mini-chromosomes from cloned centromere-like sequences and transgene cassettes, and delivered them into maize cells to form autonomous func-tional chromosomal units de novo. It was estimated that native maize chromosomes contain from ~300 kb to >2,800 kb of CentC and CRM repeat sequences in different centromeres.53 The smallest centromere of the artificial mini-chromosomes used by Carson et al.17 is less than 20 kb, which is at least 15 times smaller than that of the smallest native maize chromosome centromere of ~300 kb. Genetic mapping shows that the minimal size for cen-tromere function of B-chromosomes is around 150 kb,54 which is still more than seven-fold larger than the centromere of the mini-chromosome reported by Carson et al.17 In spite of the small size of engineered artificial mini-chromosomes, they were shown to be stably transmitted through mitosis and meiosis and were inher-ited through several generations.17 These authors also found that the reporter gene carried on their artificial mini-chromosomes was expressed normally.17 As gene vectors, mini-chromosomes offer some advantages over integration of the transgene into the host chromosomes. Transgenes on a mini-chromosome will not interrupt important endogenous genes or cause undesirable pro-tein fusions. Likewise they avoid the problem of transgene inser-tion into an undesirable region of a normal chromosome resulting in negative phenotypes, such as low transgene expression or yield drag. Because all trait genes on the mini-chromosome are linked and behave genetically as a single locus, breeding to introgress all the traits into new breeding lines should be facile, in contrast to the case with integrated transgenes, which require many backcrosses. However, it still remains to be seen how the transgenes will interact when they are carried on a minichromosome, since it is not known whether the mini-chromosome has the usual chromatin struc-ture or whether the genes will be subjected to similar epigenetic regulations as those integrated into normal chromosomes. It will be interesting to see how the mini-chromosomes are transmitted mitotically and meiotically over time under field conditions and whether trait genes will be expressed properly when the plants are subjected to various environmental stresses.
Expression of Transgenes in Molecular Stacks
One of the major uncertainties of molecular stacking concerns the expression of stacked genes over generations, and whether trait efficacy can be maintained in diverse genetic backgrounds under field conditions. This kind of multi-cassette vector would require coordinated expression of all genes in a tissue-specific fashion while the selectable marker and herbicide tolerance genes
www.landesbioscience.com GM Crops 227
transgenic Arabidopsis plants.58 The fact that only some MARs exhibit true insulating activity may explain their mixed effects on gene expression in plants.
Challenges and Uncertainties
As mentioned above, there have been many technical develop-ments for creating and delivering multiple gene stacks in the last decade. However, the adoption of these technologies will probably depend on several factors, including technical perfor-mance, development cost, grower needs, regulatory environ-ment and fit with the individual biotech company’s breeding program. One of the considerations is the trait needs of grow-ers in different regions, since pest and weed distributions are unique for each geographical location. Another consideration is the flexibility of breeding with another company’s germ-plasm and traits, since many GM traits are also cross-licensed to third party seed companies. In this case, a molecular stack event with all of the company’s traits might not be desirable since some traits might not be compatible with another compa-ny’s existing traits or are not covered by the license. For insect resistance trait programs, it is important to consider resistance management requirements while designing the trait or molecu-lar stacks. Also, for hybrid seed crops like maize, trait stack design should consider whether some traits should be on the male or the female side to make the intended hybrid seeds for future breeding efforts.
A further consideration of whether or not to adopt a large DNA stack vs. breeding stack is the sheer cost and timeline of registering a new transgenic event. In some countries such as the US and Canada, breeding stacks from previously registered events do not require new safety assessment studies. Therefore, it is much cheaper and faster to develop and commercialize a stack product via combination of existing events. However, the US EPA does require separate review of the safety of the trait stacks if a specific hazard can be identified, for example when the parental events carry traits encoding biotoxins (“plant incor-porated protectants”) such as insecticides, since combinations of biotoxins may result in synergetic or adjuvant effects. In Japan and EU countries, stacked events are considered as new GMOs and they need regulatory approval, including risk assessment of their safety, similar to single event. However, even in EU coun-tries the safety assessments of the parental events covering all uses of the stacked events will provide a good basis for the evaluation of stacked trait events, though additional information proving the validity of the studies carried out on the GM parental lines are needed to complete the risk assessment of the GM stacked event.67 Since the cost of performing regulatory studies and reg-istering a transgenic event often exceeds multimillions of dollars for an individual country and may take several years for regula-tory agencies to approve newly generated events, there is little incentive for biotech companies to spend additional millions on regulatory studies for a different transgenic event of a previ-ously approved trait if the new event does not provide clear addi-tional benefits. However, there are situations where a molecular stack might be preferred in order to avoid the cost of regulatory
herbicide tolerance trait efficacy throughout a large part of the crop’s life cycle. Another factor that may affect stacked transgene expression is the nature of the insertion site itself, i.e., the neigh-boring genomic context including activity of the cis-regulatory elements and chromatin environment. Variation ascribed to the locus is generally referred to as position effect. One strategy for reducing variability of transgene expression variation and posi-tion effect is to flank the transgene with chromatin boundary ele-ments such as insulators or matrix attachment regions. Insulators are DNA sequence elements that protect genes from the inappro-priate influences of nearby promoter elements (enhancer blocker) or the spreading of heterochromatin effects into the transgene from the surrounding regions (chromatin barrier). Various ele-ments such as insulators and MARs have been studied in several laboratories to evaluate their effect on transgene expression but results have been mixed. Nagaya et al.61 found that a sea urchin Ars (arylsulfatase) insulator can suppress variation of transgene expression in tobacco BY2 cells. Matrix-attachment regions (MARs), also known as scaffold attachment regions (SARs), are sequence elements with nuclear matrix (scaffold) binding proper-ties and are postulated to be the boundary elements organizing nuclear DNA into discrete chromatin structural domains. One study found that in Arabidopsis a chimeric transgene flanked by tomato HSC80 MARs behaved no differently than a con-trol without MARs.62 These authors also showed that the use of a different MARs, the ARS1 MAR from yeast, significantly decreased expression. However, in two different tobacco stud-ies, yeast and tobacco MARs were shown to increase transgene expression in tobacco cells.63,64 Since these studies utilized biolis-tic delivery, the dramatic improvement observed in the result-ing multi-copy lines may be a protection against gene silencing, which may also explain why no such protection was seen at a very high transgene copy number.63 In maize, three MAR elements, including two from maize (Adh1 5' MAR and Mha1 5' MAR) and one from yeast (ARS1 MAR), had very different effects on transgene expression that bore no relation to their affinity for the nuclear matrix in vitro.65 In this study, it was shown that the Adh1 5' MAR and yeast ARS1 MAR reduced transgene silenc-ing but had no effect on the variability of expression. In stable transgenic maize plants, GUS expression was lower in all tissues of the plants with Adh1 MARs flanking the GUS gene cassette compared to control plants without Adh1 MARs flanking the GUS gene cassette. However, GUS expression was found to be localized to the lateral root initiation sites in transgenic plants with Adh1 MARs flanking the GUS gene cassette.65 To elimi-nate the effect of transgene position and copy number on MAR activity, Cre-lox mediated recombination was used to generate allelic loci.66 In this study, it was found that GUS gene expres-sion in plant populations could be shielded from RNA silencing by the presence of the chicken lysozyme A element, but expres-sion stabilization was only observed in two of three independent tobacco transformants.66 Hily et al.58 showed that out of the three plant MARs tested, only the petunia TBS MAR element, but not the maize ADH1 or tobacco Rb7 MARs, was found to block interactions between the 35S enhancer and AGIP promoter with-out affecting the function of either 35S or AGIP promoters in
228 GM Crops Volume 1 Issue 4
as minimum promoters with well characterized cis-regulatory elements, enhancers and insulators, can be used in combina-tion to precisely control the spatial and temporal expression of each transgene or different transgenes in concert at the desired strength for trait stacks. In addition, the knowledge is rapidly accumulating about epigenetic features in a genome, such as histone modification, DNA methylation and microRNAs.68,69 Genetic features such as homologous recombination hotspots discovered by high density genotyping technologies, as well as information about functional regulation of host gene expression at the chromosomal level, could be used to select and develop the optimal landing sites for site-specific integration. Furthermore, the learning about the chromosomal structure and organiza-tion and their expression control from the host genomes could be applied to improve the design and function of mini-chromo-somes or artificial chromosomes. Finally, integrated analysis of all the omics data at transcript, protein, metabolite levels enables one to reverse-engineer the host systems by identifying key players of the traits. Such knowledge from systems biology studies can be used not only for forward genetic engineering, but also for miti-gation of negative impact from interaction between the trans-genes and host genome. These and probably other unforeseen technological advances will probably have an important impact in this rapidly moving field.
Acknowledgements
We would like to thank Ryan Kurtz, Larry Zeph and Roger Kemble for their critical reviews and helpful suggestions to the manuscript.
approvals of several independent events, for example in a new crop where a number of traits are needed or in a crop for which breeding is extremely difficult, for example in vegetatively propa-gated crops such as potato, cassava and sugarcane.
Transgene stacking presents many interesting challenges, both scientific and tactical, that will require input from molecular and genome biologists, breeders, growers and regulatory agencies in order to choose the most cost-effective solution. It will be very interesting to see whether one or several means of trait stack-ing will ultimately predominate in the market. Will the dark horse of the field, the mini-chromosome, which is providing so much interesting science today, turn out to be a sound and use-ful technology that supplants the current methods of inserting transgenes into the normal chromosomes of the plant? Will the frustrating and yet tantalizing results with insulator technology develop into a valuable tool for stabilizing gene expression?
In this post-structural genomic era, more crop genomes are being fully sequenced and characterized; and more technologies with higher throughput, improved accuracy and broader cover-age are being developed. Various methods have been developed to predict various regulatory features, transcription factor bind-ing sites, tandem repeats, CpG/CpNpG islands and combina-tory transcription factor binding sites, to name a few, by taking advantage of the vast amount of genomic sequence information, methylation or chromatin immunoprecipitation (ChIP) profiles and transcriptome profiling data from diverse species. These predicted features can be used to develop artificial regulators to drive various expression patterns. The artificial regulators, such
References1. James C. A global overview of biotech (GM) Crops:
Adoption, impact and future prospects. GM Crops 2010; 1:8-12.
2. CERA. GM Crop Database. Center for Environmental Risk Assessment (CERA), ILSI Research Foundation, Washington D.C. http://cera-gmc.org/index.php?action=gm_crop_database.
3. Robinson E. Fewer new chemicals chasing resistant weeds? Southwest Farm Press 2009; http://www.south-westfarmpress.com/mag/farming_fewer_new_chemi-cals/index.html.
4. Halpin C. Gene stacking in transgenic plants—the challenge for 21st century plant biotechnology. Plant Biotechnol J 2005; 3:141-55.
5. Dietz-Pfeilstetter A. Stability of transgene expression as a challenge for genetic engineering. Plant Science 2010; 179:164-7.
7. Fan Z, Yuan L. Products of multifunctional chimaeric enzymes in plants: a promising approach for degrading plant cell wall from within. Plant Biotechnol J 2010; 8:308-15.
8. Bock R. Plastid biotechnology: prospects for herbi-cide and insect resistance, metabolic engineering and molecular farming. Curr Opinion Biotechnol 2007; 18:100-6.
9. Peremarti A, Twyman RW, Gómez-Galera S, Naqvi S, Farré G, Sabalza M, et al. Promoter diversity in multi-gene transformation. Plant Mol Biol 2010; 73:363-78.
10. Fajardo-Sanchez E, Stricher F, Pâques F, Mark Isalan M, Serrano L. Computer design of obligate heterodi-mer meganucleases allows efficient cutting of custom DNA sequences. Nucleic Acids Res 2008; 36:2163-73.
11. Zeevi V, Tovkach A, Tzfira T. Increasing cloning pos-sibilities using artificial zinc finger nucleases. Proc Natl Acad Sci USA 2008; 105:12785-90.
12. Dafny-Yelin M, Tzfura T. Delivery of multiple trans-genes to plant cells. Plant Physiol 2007; 145:1118-28.
14. Gibson DG, Young L, Chuang RY, Venter JC, Hutchison CA, III, Smith HO. Enzymatic assembly of DNA molecules up to several hundred kilobases. Nat Methods 2009; 6:343-5.
15. Mullen J, Adam G, Blowers A, Earle E. Biolistic trans-fer of large DNA fragments to tobacco cells using YACs retrofitted for plant transformation. Mol Breeding 1998; 4:449-57.
16. Song R, Segal G, Messing J. Expression of the sorghum 10-member kafirin gene cluster in maize endosperm. Nuc Acid Res 2004; 32:189.
17. Carlson SR, Rudgers GW, Zieler H, Mach JM, Luo S, Grunden E, et al. Meiotic transmission of an in vitro-assembled autonomous maize minichromosome. PLoS Genet 2007; 3:1965-74.
19. Fu X, Duc LT, Fontana S, Bong BB, Tinjuangjun P, Sudhakar D, et al. Linear transgene constructs lacking vector backbone sequences generate low-copy-number transgenic plants with simple integration patterns. Transgenic Res 2000; 9:11-9.
20. Hamilton CM, Frary A, Lewis C, Tanksley SD. Stable transfer of intact high molecular weight DNA into plant chromosomes. Proc Natl Acad Sci USA 1996; 93:9975-9.
21. Frary A, Hamilton CM. Efficiency and stability of high molecular weight DNA transformation: an analysis in tomato. Transgenic Res 2001; 10:121-32.
22. He RF, Wang Y, Shi Z, Ren X, Zhu L, Weng Q, et al. Construction of a genomic library of wild rice and Agrobacterium-mediated transformation of large insert DNA linked to BPH resistance locus. Gene 2003; 321:113-21.
23. Wang W, Wu Y, Li Y, Xie J, Zhang Z, Deng Z, et al. A large insert Thellungiella halophila BIBAC library for genomics and identification of stress tolerance genes. Plant Mol Biol 2010; 72:91-9.
24. Hamilton CM. A binary-BAC system for plant trans-formation with high-molecular-weight DNA. Gene 1997; 200:107-16.
25. Liu YG, Shirano Y, Fukuki H, Yanai Y, Tasaka M, Tabata S, et al. Complementation of plant mutants with large genomic DNA fragments by a transforma-tion-competent artificial chromosome vector acceler-ates positional cloning. Proc Natl Acad Sci USA 1999; 96:6535-40.
26. Tao Q, Zhang HB. Cloning and stable maintenance of DNA fragments over 300 kb in Escherichia coli with conventional plasmid-based vectors. Nucleic Acid Res 1998; 26:4901-9.
27. Oltmanns H, Frame B, Lee LY, Johnson S, Li B, Wang K, et al. Generation of backbone-free, low transgene copy plants by launching T-DNA from the Agrobacterium chromosome. Plant Physiol 2010; 152:1158-66.
28. Cao MX, Huang JQ, Wei ZM, Yao QH, Wan CZ, Lu JA. Engineering higher yield and herbicide resistance in rice by Agrobacterium-mediated multiple gene transformation. Crop Science 2004; 44:2206-13.
29. Fujisawa M, Takita E, Harada H, Sakurai N, Suzuki H, Ohyama K, et al. Pathway engineering of Brassica napus seeds using multiple key enzyme genes involved in ketocarotenoid formation. J Exp Botany 2009; 60:1319-32.
www.landesbioscience.com GM Crops 229
58. Hily JM, Singer SD, Yang Y, Liu Z. A transformation booster sequence (TBS) from Petunia hybrida functions as an enhancer-blocking insulator in Arabidopsis thali-ana. Plant Cell Rep 2009; 28:1095-104.
59. Gudynaite-Savitch L, Johnson DA, Miki BLA. Strategies to mitigate transgene-promoter interactions. Plant Biotechnol J 2009; 7:472-85.
60. Ren S, Johnston JS, Shippen DE, McKnight TD. Telomerase Activator1 induces telomerase activity and potentiates responses to auxin in Arabidopsis. Plant Cell 2004; 16:2910-22.
61. Nagaya S, Yoshida K, Kato K, Akasaka K, Shinmyo A. An insulator element from the sea urchin Hemicentrotus pulcherrimus suppresses variation in transgene expres-sion in cultured tobacco cells. Mol Genet Genomics 2001; 265:405-13.
62. Holmes-Davis R, Comai L. The matrix attachment regions (MARs) associated with the Heat Shock Cognate 80 gene (HSC80) of tomato represent spe-cific regulatory elements. Mol Genet Genomics 2002; 266:891-8.
63. Allen GC, Hall G Jr, Michalowski S, Newman W, Spiker S, Weissinger AK, et al. High-level transgene expression in plant cells: Effects of a strong scaffold attachment region from tobacco. Plant Cell 1996; 8:899-913.
64. Halweg C, Thompson WF, Spiker S. The Rb7 matrix attachment region increases the likelihood and magni-tude of transgene expression in tobacco cells: A flow cytometric study. Plant Cell 2005; 17:418-29.
65. Brouwer C, Bruce W, Maddock S, Avramova Z, Bowen B. Suppression of transgene silencing by matrix attach-ment regions in maize: A dual role for the maize 5' ADH1 matrix attachment region. Plant Cell 2002; 14:2251-64.
66. Mlynárová L, Hricová A, Loonen A, Nap JP. The presence of a chromatin boundary appears to shield a transgene in tobacco from RNA silencing. Plant Cell 2003; 15:2203-17.
67. De Schrijver A, Devosb Y, Van den Bulckea M, Cadotc P, De Loosed M, Reheulb D, et al. Risk assessment of GM stacked events obtained from crosses between GM events. Trends Food Sci Tech 2007; 18:101-9.
68. Li X, Wang X, He K, Ma Y, Su N, He H, et al. High-resolution mapping of epigenetic modifications of the rice genome uncovers interplay between DNA methyla-tion, histone methylation and gene expression. Plant Cell 2008; 20:259-76.
69. Lister R, O’Malley RC, Tonti-Filippini J, Gregory BD, Berry CC, Millar AH, et al. Highly integrated single-base resolution maps of the epigenome in Arabidopsis. Cell 2008; 133:523-36.
45. Urnov FD, Miller JC, Lee YL, Beausejour CM, Rock JM, Augustus S, et al. Highly efficient endogenous human gene correction using designed zinc-finger nucleases. Nature 2005; 435:646-51.
46. Cai CQ, Doyon Y, Ainley WM, Miller JC, DeKelver RC, Moehle EA, et al. Targeted transgene integration in plant cells using designed zinc finger nucleases. Plant Molecular Biology 2009; 69:699-709.
47. De Pater S, Neuteboom LW, Pinas JE, Hooykaas PJJ, van der Zaal BJ. ZFN-induced mutagenesis and gene-targeting in Arabidopsis through Agrobacterium-mediated floral dip transformation. Plant Biotechnol J 2009; 7:821-35.
48. Wright DA, Townsend JA, Winfrey RJ Jr, Irwin PA, Rajagopal J, Lonosky PM, et al. High-frequency homologous recombination in plants mediated by zinc-finger nucleases. Plant J 2005; 44:693-705.
49. Shukla VK, Doyon Y, Miller JC, DeKelver RC, Moehle EA, Worden SE, et al. Precise genome modification in the crop species Zea mays using zinc-finger nucleases. Nature 2009; 459:437-41.
50. Townsend JA, Wright DA, Winfrey RJ, Fu F, Maeder ML, Joung JK, et al. High-frequency modification of plant genes using engineered zinc-finger nucleases. Nature 2009; 459:442-5.
51. Ow DW. GM maize from site-specific recombination technology, what next? Curr Opin Biotechnol 2007; 18:115-20.
52. Yu W, Han F, Gao Z, Vega JM, Birchler JA. Construction and behavior of engineered minichro-mosomes in maize. Proc Natl Acad Sci USA 2007; 104:8924-9.
53. Jin W, Melo JR, Nagaki K, Talbert PB, Henikoff S, Dawe RK, et al. Maize centromeres: organization and functional adaptation in the genetic background of oat. Plant Cell 2004; 16:571-81.
54. Birchler JA, Han F. Maize centromeres: structure, func-tion, epigenetics. Annu Rev Genet 2009; 43:287-303.
55. Naqvi S, Zhu C, Farre G, Ramessar K, Bassie L, Breitenbach J, et al. Transgenic multivitamin corn through biofortification of endosperm with three vita-mins representing three distinct metabolic pathways. Proc Natl Acad Sci USA 2009; 106:7762-7.
56. De Buck S, Windels P, De Loose M, Depicker A. Single-copy T-DNAs integrated at different positions in the Arabidopsis genome display uniform and com-parable b-glucuronidase accumulation levels. Cell Mol Life Sci 2004; 61:2632-45.
57. Day CD, Lee E, Kobayashi JK, Holappa LD, Albert H, Ow DW. Transgene integration into the same chromosome location can produce alleles that express at a predictable level or alleles that are differentially silenced. Genes and Dev 2000; 14:2869-80.
30. Ward J, Xu J, Lamb J, Gilbertson L. The use of Agrobacterium tumefaciens to deliver ten genes on a sin-gle vector to corn. The 9th International Plant Molecular Biology (IPMB) Congress 2009; Poster #1364.
31. Komari T, Hiei Y, Saito Y, Murai N, Kumashiro T. Vectors carrying two separate T-DNAs for co-trans-formation of higher plants mediated by Agrobacterium tumefaciens and segregation of transformants free from selection markers. Plant J 1996; 10:165-74.
32. McCormac AC, Fowler MR, Chen DF, Elliott MC. Efficient co-transformation of Nicotiana tabacum by two independent T-DNAs, the effect of T-DNA size and implications for genetic separation. Transgenic Res 2001; 10:143-55.
33. Chen L, Marmey P, Taylor NJ, Brizard JP, Espinoza C, D’Cruz P, et al. Expression and inheritance of mul-tiple transgenes in rice plants. Nat Biotechnol 1998; 16:1060-4.
34. Zhu C, Naqvi S, Breitenbach J, Sandmann S, Christou P, Capell T. Combinatorial genetic transformation generates a library of metabolic phenotypes for the carotenoid pathway in maize. Proc Natl Acad Sci USA 2008; 105:18232-7.
35. Naqvi S, Farre G, Sanahuja G, Capell T, Zhu C, Christou P. When more is better: multigene engineer-ing in plants. Trends in Plant Science 2009; 15:48-56.
36. Albert H, Dale EC, Ow DW. Site-specific integration of DNA into wild-type and mutant lox sites placed in the plant genome. Plant J 1995; 7:649-59.
37. Srivastava V, Ariza-Nieto M, Wilson AJ. Cre-mediated site-specific gene integration for consistent transgene expression in rice. Plant Biotechnol J 1999; 2:169-79.
38. Suttie JL, Chilton MD, Que Q. Lambda integrase mediated recombination in plants. US Patent 2008; 7:351-877.
39. Li Z, Xing A, Moon BP, McCardell RP, Mills K, Falco SC. Site-specific integration of transgenes in soybean via recombinase mediated DNA cassette exchange. Plant Physiol 2009; 151:1087-95.
40. Prieto J, Epinat JC, Redondo P, Ramos E, Padró D, Cédrone F, et al. Generation and analysis of mesophilic variants of the thermostable archaeal I-DmoI homing endonuclease. J Biol Chem 2008; 283:4364-74.
41. Puchta H, Dujon B, Hohn B. Two different but related mechanisms are used in plants for the repair of genomic double-strand breaks by homologous recombination. Proc Natl Acad Sci USA 1996; 93:5055-60.
42. Que Q. Targeted integration and stacking of DNA through homologous recombination. US Patent Application 2006/0253918.
43. D’Halluin K, Vanderstraeten C, Stals E, Cornelissen M, Ruiter R. Homologous recombination: a basis for targeted genome optimization in crop species such as maize. Plant Biotech J 2007; 6:93-102.
44. Chilton MDM, Que Q. Targeted Integration of T-DNA into the tobacco genome at double-stranded breaks: New insights on the mechanism of T-DNA integration. Plant Physiol 2003; 133:956-65.