Baseline Bloom of Cochlodinium polykrikoides (Dinophyceae: Gymnodiniales) in Bahía de La Paz, Gulf of California Ismael Gárate-Lizárraga ⇑ Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN), Departamento de Plancton y Ecología Marina, Avenida Instituto Politécnico Nacional s/n, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, Mexico article info Keywords: Proliferation Blooms Dinoflagellates Cochlodinium Diatoms New records abstract As part of a continuing toxic microalgae monitoring program, phytoplankton samples were collected on 16–17 August 2012 at several sampling sites in the southern part of the Bahía de La Paz. A bloom of the dinoflagellate Cochlodinium polykrikoides was detected. Abundance of C. polykrikoides ranged from 73 to 276 Â 10 3 cells L 1 on the first day to 980–1425 Â 10 3 cells L 1 on the second day. Study of live speci- mens showed great variation in cell size and form, mainly as single cells or chains of two cells. Live cells were 30–47 lm long and 20–35 lm wide (n = 30). Seawater temperature during the bloom was 29–30 °C. Low densities of Cochlodinium convolutum, Cochlodinium helicoides, and Cochlodinium shuettii were also found in the samples of the bloom event. These high densities of C. polykrikoides did not lead to fish die-offs in the bay. Ó 2012 Elsevier Ltd. All rights reserved. Microalgae blooms have increased in the past 20 years in the Gulf of California (Cortés-Altamirano and Licea-Durán, 2004; Gárate-Lizárraga et al., 2006). These blooms are frequent and peri- odic throughout the year in Bahía de La Paz in the southwestern part of the Gulf of California (Gárate-Lizárraga, 2011). Microalgae blooms could be innocuous or cause negative impacts to marine fauna through poisoning, mechanical damage, or other media (Gárate-Lizárraga, 2011). Bloom-forming species belong to differ- ent phytoplankton groups: dinoflagellates, diatoms, silicoflagel- lates, raphidophytes, planktonic cyanobacteria, and the ciliate Myrionecta rubra (Gárate-Lizárraga et al., 2001, 2006, 2009a,b; Band-Schmidt et al., 2005; López-Cortés et al., 2008; Gárate- Lizárraga and Muciño-Márquez, 2012). Most of blooms in Bahía de La Paz are caused by dinoflagellates (Gárate-Lizárraga et al., 2001, 2006, 2009a,b; Gárate-Lizárraga, 2011, 2012). Unarmored dinoflagellates, such as Noctiluca scintillans, Gymnodinium catena- tum, Gyrodinium instriatum, Katodinium glaucum, and Cochlodinium polykrikoides , are the main blooming species in the Bahía de La Paz (Gárate-Lizárraga et al., 2009a). First blooms of C. polykrikoides were observed during the late summer of 2000 and early autumn 2001. On both occasions, fish die-offs in fish ponds were observed (Gárate-Lizárraga et al., 2004). After that, sporadic proliferation of this species has occurred with no consequences for the marine bio- ta (Gárate-Lizárraga and Muñetón-Gómez, 2008; Gárate-Lizárraga et al., 2009a). This report describes the most recent bloom of C. polykrikoides in Bahía de La Paz. The morphological variation of C. polykrikoides and the microalgae community present during this bloom is also described. Phytoplankton bottle samples were collected during prolifera- tion of microalgae observed during a routine survey performed at five sampling stations in the southern part of Bahía de La Paz (Fig. 1). An algae bloom was detected on 16 and 17 August 2012. Bahía de La Paz is the largest bay on the peninsular side of the Gulf of California. It has constant exchange of water with the Gulf of California via a northern and a southern broad channel (Gómez- Valdés et al., 2003). The southern channel is shallow and associated with a shallow basin about 10 m deep. There is a shallow lagoon, the Ensenada of La Paz, connected to the bay by a narrow inlet (1.2 km wide and 4 km long) with an average depth of 7 m. Sam- pling stations are located in the shallow basin of the southernmost region of the Bahía de La Paz. Phytoplankton samples were fixed with Lugol’s solution. Identification and cell counts were made in 5 ml settling chambers in an inverted Carl Zeiss phase-contrast microscope. Surface and vertical tows from 15 m were made with a 20 lm mesh phytoplankton net. A portion of each tow was immediately fixed with acid Lugol’s solution and later preserved in 4% formalin. These samples were used to identify some uncom- mon species found in the bottle samples. Sea surface temperature (SST) was measured with a bucket thermometer (Kahl Scientific Instruments, El Cajon, CA). A compound microscope was used to measure cells and a digital camera (Konus Optical & Sport Systems, Verona, Italy) were used to record the phytoplankton images. A total of 75 microalgae taxa from the phytoplankton samples were identified during the red tide: 37 were diatoms, 33 dinoflagel- lates, 1 cyanobacterium, 1 ebriid, 1 silicoflagellate, 1 raphydophyte, 0025-326X/$ - see front matter Ó 2012 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.marpolbul.2012.11.031 ⇑ Tel.: +52 612 123 0350x82434; fax: +52 612 122 5322. E-mail addresses: [email protected], [email protected]Marine Pollution Bulletin 67 (2013) 217–222 Contents lists available at SciVerse ScienceDirect Marine Pollution Bulletin journal homepage: www.elsevier.com/locate/marpolbul
Transcript
Marine Pollution Bulletin 67 (2013) 217–222
Contents lists available at SciVerse ScienceDirect
Bloom of Cochlodinium polykrikoides (Dinophyceae: Gymnodiniales) in Bahía de LaPaz, Gulf of California
Ismael Gárate-Lizárraga ⇑Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas (CICIMAR-IPN), Departamento de Plancton y Ecología Marina, Avenida Instituto Politécnico Nacionals/n, Col. Playa Palo de Santa Rita, La Paz, B.C.S. 23096, Mexico
a r t i c l e i n f o a b s t r a c t
Keywords:ProliferationBloomsDinoflagellatesCochlodiniumDiatomsNew records
0025-326X/$ - see front matter � 2012 Elsevier Ltd. Ahttp://dx.doi.org/10.1016/j.marpolbul.2012.11.031
As part of a continuing toxic microalgae monitoring program, phytoplankton samples were collected on16–17 August 2012 at several sampling sites in the southern part of the Bahía de La Paz. A bloom of thedinoflagellate Cochlodinium polykrikoides was detected. Abundance of C. polykrikoides ranged from 73 to276 � 103 cells L�1 on the first day to 980–1425 � 103 cells L�1 on the second day. Study of live speci-mens showed great variation in cell size and form, mainly as single cells or chains of two cells. Live cellswere 30–47 lm long and 20–35 lm wide (n = 30). Seawater temperature during the bloom was 29–30 �C.Low densities of Cochlodinium convolutum, Cochlodinium helicoides, and Cochlodinium shuettii were alsofound in the samples of the bloom event. These high densities of C. polykrikoides did not lead to fishdie-offs in the bay.
� 2012 Elsevier Ltd. All rights reserved.
Microalgae blooms have increased in the past 20 years in theGulf of California (Cortés-Altamirano and Licea-Durán, 2004;Gárate-Lizárraga et al., 2006). These blooms are frequent and peri-odic throughout the year in Bahía de La Paz in the southwesternpart of the Gulf of California (Gárate-Lizárraga, 2011). Microalgaeblooms could be innocuous or cause negative impacts to marinefauna through poisoning, mechanical damage, or other media(Gárate-Lizárraga, 2011). Bloom-forming species belong to differ-ent phytoplankton groups: dinoflagellates, diatoms, silicoflagel-lates, raphidophytes, planktonic cyanobacteria, and the ciliateMyrionecta rubra (Gárate-Lizárraga et al., 2001, 2006, 2009a,b;Band-Schmidt et al., 2005; López-Cortés et al., 2008; Gárate-Lizárraga and Muciño-Márquez, 2012). Most of blooms in Bahíade La Paz are caused by dinoflagellates (Gárate-Lizárraga et al.,2001, 2006, 2009a,b; Gárate-Lizárraga, 2011, 2012). Unarmoreddinoflagellates, such as Noctiluca scintillans, Gymnodinium catena-tum, Gyrodinium instriatum, Katodinium glaucum, and Cochlodiniumpolykrikoides, are the main blooming species in the Bahía de La Paz(Gárate-Lizárraga et al., 2009a). First blooms of C. polykrikoideswere observed during the late summer of 2000 and early autumn2001. On both occasions, fish die-offs in fish ponds were observed(Gárate-Lizárraga et al., 2004). After that, sporadic proliferation ofthis species has occurred with no consequences for the marine bio-ta (Gárate-Lizárraga and Muñetón-Gómez, 2008; Gárate-Lizárragaet al., 2009a). This report describes the most recent bloom ofC. polykrikoides in Bahía de La Paz. The morphological variation of
ll rights reserved.
5322.o.com.mx
C. polykrikoides and the microalgae community present during thisbloom is also described.
Phytoplankton bottle samples were collected during prolifera-tion of microalgae observed during a routine survey performed atfive sampling stations in the southern part of Bahía de La Paz(Fig. 1). An algae bloom was detected on 16 and 17 August 2012.Bahía de La Paz is the largest bay on the peninsular side of the Gulfof California. It has constant exchange of water with the Gulf ofCalifornia via a northern and a southern broad channel (Gómez-Valdés et al., 2003). The southern channel is shallow and associatedwith a shallow basin about 10 m deep. There is a shallow lagoon,the Ensenada of La Paz, connected to the bay by a narrow inlet(1.2 km wide and 4 km long) with an average depth of 7 m. Sam-pling stations are located in the shallow basin of the southernmostregion of the Bahía de La Paz. Phytoplankton samples were fixedwith Lugol’s solution. Identification and cell counts were made in5 ml settling chambers in an inverted Carl Zeiss phase-contrastmicroscope. Surface and vertical tows from 15 m were made witha 20 lm mesh phytoplankton net. A portion of each tow wasimmediately fixed with acid Lugol’s solution and later preservedin 4% formalin. These samples were used to identify some uncom-mon species found in the bottle samples. Sea surface temperature(SST) was measured with a bucket thermometer (Kahl ScientificInstruments, El Cajon, CA). A compound microscope was used tomeasure cells and a digital camera (Konus Optical & Sport Systems,Verona, Italy) were used to record the phytoplankton images.
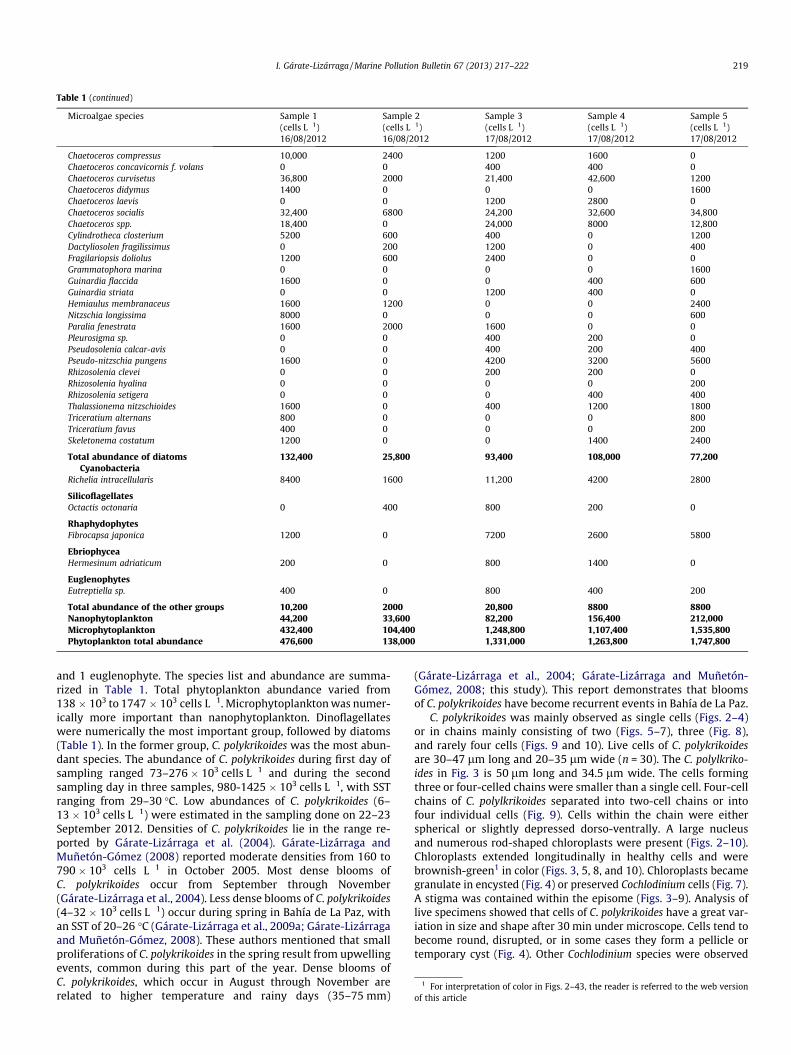
A total of 75 microalgae taxa from the phytoplankton sampleswere identified during the red tide: 37 were diatoms, 33 dinoflagel-lates, 1 cyanobacterium, 1 ebriid, 1 silicoflagellate, 1 raphydophyte,
Total abundance of the other groups 10,200 2000 20,800 8800 8800Nanophytoplankton 44,200 33,600 82,200 156,400 212,000Microphytoplankton 432,400 104,400 1,248,800 1,107,400 1,535,800Phytoplankton total abundance 476,600 138,000 1,331,000 1,263,800 1,747,800
1 For interpretation of color in Figs. 2–43, the reader is referred to the web versionof this article
I. Gárate-Lizárraga / Marine Pollution Bulletin 67 (2013) 217–222 219
and 1 euglenophyte. The species list and abundance are summa-rized in Table 1. Total phytoplankton abundance varied from138 � 103 to 1747 � 103 cells L�1. Microphytoplankton was numer-ically more important than nanophytoplankton. Dinoflagellateswere numerically the most important group, followed by diatoms(Table 1). In the former group, C. polykrikoides was the most abun-dant species. The abundance of C. polykrikoides during first day ofsampling ranged 73–276 � 103 cells L�1 and during the secondsampling day in three samples, 980-1425 � 103 cells L�1, with SSTranging from 29–30 �C. Low abundances of C. polykrikoides (6–13 � 103 cells L�1) were estimated in the sampling done on 22–23September 2012. Densities of C. polykrikoides lie in the range re-ported by Gárate-Lizárraga et al. (2004). Gárate-Lizárraga andMuñetón-Gómez (2008) reported moderate densities from 160 to790 � 103 cells L�1 in October 2005. Most dense blooms ofC. polykrikoides occur from September through November(Gárate-Lizárraga et al., 2004). Less dense blooms of C. polykrikoides(4–32 � 103 cells L�1) occur during spring in Bahía de La Paz, withan SST of 20–26 �C (Gárate-Lizárraga et al., 2009a; Gárate-Lizárragaand Muñetón-Gómez, 2008). These authors mentioned that smallproliferations of C. polykrikoides in the spring result from upwellingevents, common during this part of the year. Dense blooms ofC. polykrikoides, which occur in August through November arerelated to higher temperature and rainy days (35–75 mm)
(Gárate-Lizárraga et al., 2004; Gárate-Lizárraga and Muñetón-Gómez, 2008; this study). This report demonstrates that bloomsof C. polykrikoides have become recurrent events in Bahía de La Paz.
C. polykrikoides was mainly observed as single cells (Figs. 2–4)or in chains mainly consisting of two (Figs. 5–7), three (Fig. 8),and rarely four cells (Figs. 9 and 10). Live cells of C. polykrikoidesare 30–47 lm long and 20–35 lm wide (n = 30). The C. polylkriko-ides in Fig. 3 is 50 lm long and 34.5 lm wide. The cells formingthree or four-celled chains were smaller than a single cell. Four-cellchains of C. polylkrikoides separated into two-cell chains or intofour individual cells (Fig. 9). Cells within the chain were eitherspherical or slightly depressed dorso-ventrally. A large nucleusand numerous rod-shaped chloroplasts were present (Figs. 2–10).Chloroplasts extended longitudinally in healthy cells and werebrownish-green1 in color (Figs. 3, 5, 8, and 10). Chloroplasts becamegranulate in encysted (Fig. 4) or preserved Cochlodinium cells (Fig. 7).A stigma was contained within the episome (Figs. 3–9). Analysis oflive specimens showed that cells of C. polykrikoides have a great var-iation in size and shape after 30 min under microscope. Cells tend tobecome round, disrupted, or in some cases they form a pellicle ortemporary cyst (Fig. 4). Other Cochlodinium species were observed
2
5
13 14 1511 12
N
6
N
7
3 4
8
109
Figs. 2–15. Light microscopy images of Cochlodinium species from Bahía de La Paz, August 2012. Figs. 2–4 show three different single specimens of Cochlodiniumpolykrikoides; N = nucleus. Fig. 4 show a pellicle or temporary cyst of Cochlodinium polykrikoides, red arrow indicates the hyaline membrane; Figs. 5–7 show different two-celled chains of Cochlodinium polykrikoides with morphological variations. Fig. 7 shows a Lugol-fixed two-celled chain of Cochlodinium polykrikoides. Fig. 8 is a three-celledchain of Cochlodinium polykrikoides. Fig. 9 shows a Cochlodinium polykrikoides chain ready to separate. Fig. 10 is a four-celled chain of Cochlodinium polykrikoides. Fig. 11 showsa specimen of Cochlodinium convolutum in ventral view. Figs. 12 and 13 show live and Lugol-fixed temporary cysts of Cochlodinium helicoides, respectively. Fig. 14 shows a livespecimen of Cochlodinium shuetti. Fig. 15 shows a live temporary cyst of Cochlodinium shuettii. White arrows indicate the bright orange stigma in the lateral area of the cell.Black arrows indicate the flagella of the cells. Yellow arrows indicate the rod-shaped chloroplasts of the cells.
220 I. Gárate-Lizárraga / Marine Pollution Bulletin 67 (2013) 217–222
in the samples: Cochlodinium convolutum (Fig. 11), Cochlodiniumhelicoides (Figs. 12 and 13), and Cochlodinium shuettii (Figs. 14 and15). These species were recently reported in this bay in net phyto-plankton samples (Gárate-Lizárraga et al., 2011a). Figs. 16–43 showselected microalgae identified during the Cochlodinium polykrokoidesproliferation. In this survey, four species of microalgae producers ofokadaic acid were observed: Dinophysis acuminata (not illustrated),Dinophysis caudata (Fig. 16), Dinophysis tripos (Fig. 17), and Dinophy-sis fortii (Fig. 18). Dinophysis shuettii (Fig. 19) is a common dinoflagel-late in Bahía de La Paz. Akashiwo sanguinea (Fig. 20) and Kareniamikimotoi (Fig. 21) are also blooming dinoflagellates. Gymnodiniumwilczekii (Fig. 22) and Warnowia sp. (Fig. 23) are recorded for the first
time along the Pacific coasts of Mexico. Gambierdiscus cf. toxicus(Fig. 24) is known to produce the following toxins: ciguatoxin, gam-bieric acid, and maitotoxin (Yasumoto et al., 1993) Coolia monotis(Fig. 25) is a tychoplanktonic species that produces cooliatoxin(Holmes et al., 1995). Prorocentrum balticum (Fig. 26) is a tycho-planktonic species rarely reported in Bahía de La Paz. The dinoflagel-late Palaeophalacroma unicinctum (Fig. 27) is first recorded in theGulf of California. Scrippsiella spinifera (Fig. 28) is a dinoflagellatespecies rarely reported in Bahía de La Paz (Okolodkov andGárate-Lizárraga, 2006). Chaetoceros concavicornis f. volans (Fig. 29)and C. socialis (Fig. 30) are two potentially noxious diatoms. Otherpelagic diatoms were identified: Dactyliosolen fragilissimus (Fig. 31),
16 17
24
26
25
18
27
32
29
31
23
30
38 43
21
28
22
33 373643
39 40 41 42
19 20
35
Figs. 16–43. Show selected microalgae identified during the Cochlodinium polykrokoides proliferation. Dinophysis caudata (Fig. 16), Dinophysis tripos (Fig. 17), Dinophysis fortii(Fig. 18), Dinophysis schuettii (Fig. 19), Akashiwo sanguinea (Fig. 20), Karenia mikimotoi (Fig. 21), Gymnodinium wilczekii (Fig. 22), Warnowia sp. (Fig. 23), Gambierdiscus cf.toxicus (Fig. 24), Coolia monotis (Fig. 25), Prorocentum balticum (Fig. 26), Palaeophalacroma unicinctum (Fig. 27), Scrippsiella spinifera (Fig. 28), Chaetoceros concavicornis f. volans(Fig. 29), Chaetoceros socialis (Fig. 30), Dactyliosolen fragilissimus (Fig. 31), Rhizosolenia setigera (Fig. 32), Guinardia striata (Fig. 33), Rhizosolenia clevei; white arrow indicates thecyanobacterium Richelia intracellularis (Fig. 34), Hemiaulus membranaceus (Fig. 35), Grammatophora marina (Fig. 36), Paralia fenestrata (Fig. 37), Trachyneis aspera girdle view(Fig. 38), Auliscus elegans var. californica (Fig. 39), Hermesinum adriaticum with numerous Synechococcus-like endosymbiotic cyanobacteria, indicated by a white arrow(Fig. 40), Eutreptia sp.; white arrowheads indicate the flagellum (Fig. 41) and black arrow indicates the stigma (Fig. 42), and Fibrocapsa japonica (Fig. 43).
I. Gárate-Lizárraga / Marine Pollution Bulletin 67 (2013) 217–222 221
Rhizosolenia setigera (Fig. 32), Guinardia striata (Fig. 33), R. clevei withRichelia intracellularis, one intracellular endosymbiont (Fig. 34), andHemiaulus membranaceus (Fig. 35). Tychoplanktonic diatomsidentified were Grammatophora marina (Fig. 36), Paralia fenestrata(Fig. 37), Trachyneis aspera (Fig. 38), and Auliscus elegans var. califor-nica (Fig. 39). The ebriid Hermesinum adriaticum (Fig. 40) withnumerous Synechococcus-like endosymbiotic cyanobacteria andEutreptia sp. (Figs. 41 and 42) are two uncommon microalgae inBahía de La Paz that cause blooms in the Gulf of California.Fibrocapsa japonica (Fig. 43) is a fish killer raphidophyte (Pezzolesiet al., 2010).
C. polykrikoides is globally widespread and a red tide species re-lated to extensive fish kills and great economic loss in Japan, Korea,The Philippines, Central America, Cuba, the west coast of NorthAmerica, and the Persian Gulf (Yuki and Yoshimatsu, 1989; Fukuyoet al., 1990; Kim, 1998; Vargas-Montero et al., 2006; Gómez et al.,2007; Azanza et al., 2008; Mulholland et al., 2009; Matsuoka et al.,2010). Kim et al. (1999) reported that reactive oxygen species gen-erated by C. polykrikoides are causative factors responsible for theoxidative damage of gill tissue leading to fish kills. Although C.polykrikoides is widely distributed along the Pacific coasts ofMexico (López, 2000; Gárate-Lizárraga et al., 2000, 2004, 2009c;
222 I. Gárate-Lizárraga / Marine Pollution Bulletin 67 (2013) 217–222
Morales-Blake and Hernández-Becerril, 2001; Okolodkov andGárate-Lizárraga, 2006; Cortés-Altamirano et al., 2006; Gómez-Villarreal et al., 2008), few cases of fish mortalities have been re-ported. Although C. polykrikoides has co-occurred with the toxicdinoflagellate Gymnodinium catenatum in Bahía de Acapulco forseveral years, there were no fish mortalities (Gárate-Lizárragaet al., 2009c; Gárate-Lizárraga et al., 2011b). The only two casesof fish kills by C. polykrikoides occurred in Bahía Banderas, Nayarit(López, 2000; Gómez-Villarreal et al., 2008) and in Bahía de La Pazin summer–autumn 2000 and 2001 (Gárate-Lizárraga et al., 2000,2004). In Bahía de La Paz, fish kills from C. polykrikoides bloomshave occurred in fish and shrimp ponds (Gárate-Lizárraga et al.,2004). During the bloom event of October 2005, reported byGárate-Lizárraga and Muñetón-Gómez (2008) and in this event,no fish kills occurred. Since 2000, a monthly monitoring programof bloom forming and toxin-producing microalgae species in Bahíade La Paz and other areas along the coast of the State of BajaCalifornia Sur is an on-going activity. This program has alloweddetection of more than 50 blooms and more than 34 responsiblespecies.
Acknowledgment
The study was funded by the Instituto Politécnico Nacional(Projects: SIP-20121153 and SIP-20121152). The author is a COFAAand EDI fellow.
References
Azanza, R.V., David, L.T., Borja, R.T., Baula, I.U., Fukuyo, Y., 2008. An extensiveCochlodinium bloom along the western coast of Palawan, Philippines. HarmfulAlgae 7, 324–330.
Band-Schmidt, C., Martínez-López, A., Gárate-Lizárraga, I., 2005. First record ofChattonella marina in Bahía de la Paz, Gulf of California. Harmful Algae News 28,6–7.
Cortés-Altamirano, R., Licea-Durán, S., 2004. Decoloración de proliferaciones demicroalgas como parámetro bioindicador en la Bahía de Mazatlán. MéxicoRevista de Biología Tropical 52 (Suppl. 1), 27–34.
Cortés-Altamirano, R., Lavín, M.F., Sierra-Beltrán, A.P., Cortés-Lara, M.del E., 2006.Hipótesis sobre el transporte de microalgas invasoras del oriente al Golfo deCalifornia por las corrientes marinas. Ciencias del Mar, Universidad Autónomade Sinaloa 18, 19–26.
Fukuyo, Y., Takano, H., Chihara, M., Matsuoka, K., 1990. Red Tide Organisms inJapan. An Illustrated Taxonomic Guide, Uchida Rokakuho, Tokyo, p. 407.
Gárate-Lizárraga, I., 2011. Harmful algae blooms along the coast of the BajaCalifornia Peninsula. Harmful Algae News 44, 6–7.
Gárate-Lizárraga, I., 2012. Proliferation of Amphidinium carterae (Gymnodiniales:Gymnodiniaceae) in Bahía de La Paz, Gulf of California. CICIMAR Oceánides 27,1–13.
Gárate-Lizárraga, I., Muñetón-Gómez, M.S., 2008. Los riesgos de las mareas rojas enel Pacifico Mexicano. Conversus 3, 20–23.
Gárate-Lizárraga, I., Muciño-Márquez, R.E., 2012. Blooms of Trichodesmiumerythraeum and T. thiebautii (Cyanobacteria, Oscillatoriales) in the Bahía de LaPaz, Gulf of California. CICIMAR, Oceánides 27, 61–64.
Gárate-Lizárraga, I., Bustillos-Guzmán, J.J., Morquecho, L.M., Lechuga-Deveze, C.H.,2000. First outbreak of Cochlodinium polykrikoides in the Gulf of California.Harmful Algae News 21, 7.
Gárate-Lizárraga, I., Hernández-Orozco, M.L., Band-Schmidt, C.J., Serrano-Casillas,G., 2001. Red tides along the coasts of the Baja California Sur, México (1984–1999). Oceánides 16, 127–134.
Gárate-Lizárraga, I., López-Cortés, D.J., Bustillos-Guzmán, J.J., Hernández-Sandoval,F.E., 2004. Blooms of Cochlodinium polykrikoides (Gymnodiniaceae) in the Gulf ofCalifornia. Mexico Revista de Biología Tropical 52 (Suppl. 1), 51–58.
Gárate-Lizárraga, I., Muñetón-Gómez, M.S., Maldonado-López, V., 2006.Florecimiento del dinoflagelado Gonyaulax polygramma frente a la Isla EspírituSanto, Golfo de California (Octubre 2004). Revista de Investigaciones Marinas17, 31–39.
Gárate-Lizárraga, I., Band-Schmidt, C.J., Aguirre-Bahena, F., Grayeb-del Álamo, T.,2009a. A multi-species microalgae bloom in Bahía de La Paz, Gulf of California,Mexico (June 2008). CICIMAR Oceánides 24, 1–15.
Gárate-Lizárraga, I., Band-Schmidt, C.J., López-Cortés, D.J., Muñetón-Gómez, M.S.,2009b. Bloom of Scrippsiella trochoidea (Gonyaulacaceae) in a shrimp pond fromthe southwestern Gulf of California, Mexico. Mar. Pollut. Bull. 58, 145–149.
Gárate-Lizárraga, I., Díaz-Ortiz, J.A., Pérez-Cruz, B., Alarcón-Tacuba, M., Torres-Jaramillo, A., Alarcón-Romero, M.A., López-Silva, S., 2009c. Cochlodiniumpolykrikoides and Gymnodinium catenatum in Bahía de Acapulco, Mexico(2005–2008). Harmful Algae News 40, 8–9.
Gárate- Lizárraga, I., García-Domínguez, F., Pérez-Cruz, B., Díaz-Ortíz, J.A., 2011a.First record of Cochlodinium convolutum and C. helicoides (Gymnodiniales:Dinophyceae) in the Gulf of California. Revista de Biología Marina yOceanografía 46, 495–498.
Gárate-Lizárraga, I., Díaz-Ortiz, J.A., Pérez-Cruz, B., Alarcón-Romero, M.A., Chávez-Almazán, M.L., López-Silva, S., 2011b. A multi-species dinoflagellate bloom andshellfish toxicity in Costa Grande, Guerrero, Mexico (December, 2010).CICIMAR, Oceánides 26, 67–71.
Gómez, L., Licea, Y., Díaz, U., Álvarez, I., 2007. Primer reporte de Cochlodiniumpolykrikoides Margalef en aguas cubanas. Revista de Investigaciones Marinas 28,87–89.
Gómez-Valdés, J., Delgado, J.A., Dwora, J.A., 2003. Overtides, compound tides, andtidal-residual current in Ensenada de la Paz lagoon, Baja California Sur, Mexico.Geofísica Internacional 42, 623–634.
Gómez-Villarreal, M.C., Martínez-Gaxiola, M.D., Peña-Manjarrez, J.L., 2008. Algalblooms at Banderas Bay, Mexico (2000–2001), from SeaWiFS-sensor-data.Revista de Biología Tropical 56, 1653–1664.
Holmes, M.J., Lewis, R.J., Jones, A., Wong Hoy, A.W., 1995. Cooliatoxin, the first toxinfrom Coolia monotis (Dinophyceae). Nat. Toxins 3, 355–362.
Kim, H.G., 1998. Cochlodinium polykrikoides blooms in Korean coastal waters andtheir mitigation. In: Reguera, B., Blanco, J., Fernández, Ma, L., Wyatt, T. (Eds.).Harmful Algae, Xunta de Galicia and Intergovernmental OceanographicCommission of UNESCO, Spain, pp. 227–228.
Kim, C.S., Lee, S.G., Lee, C.K., Kim, H.G., Jung, J., 1999. Reactive oxygen species ascausative agents in the ichthyotoxicity of the red tide dinoflagellateCochlodinium polykrikoides. J. Plankton Res. 21, 2105–2115.
López, L., 2000. Marea roja en Bahía de Banderas. Gaceta Universitaria 1, 6.López-Cortés, D.J., Gárate-Lizárraga, I., Bustillos-Guzmán, J.J., Hernández-Sandoval,
F., 2008. Blooms of Myrionecta rubra in Bahía de La Paz, Gulf of California,Mexico in early summer 2005. CICIMAR Oceánides 23, 1–10.
Matsuoka, K., Takano, Y., Kamrani, E., Rezai, H., Puthiyedathu, T.S., Al Gheilani, H.M.,2010. Study on Cochlodinium polykrikoides Margalef (Gymnodiniales,Dinophyceae) occurring in the Oman Sea and the Persian Gulf from August of2008 to August 2009. Current Development in Oceanography 1, 153–171.
Morales-Blake, A., Hernández-Becerril, D.U., 2001. Unusual HABs in Manzanillo Bay,Colima, México. Harmful Algae News 21, 6.
Mulholland, M.R., Morse, R.E., Boneillo, G.E., Bernhardt, P.W., Filippino, K.C., Procise,L.A., Garcia-Blanco, J.L., Marshall, H.G., Egerton, T.A., Hunley, W.S., Moore, K.A.,Berry, D.L., Gobler, C.J., 2009. Understanding the causes and impacts of thedinoflagellate. Cochlodinium polykrikoides blooms in Chesapeake Bay. EstuariesCoasts. http://dx.doi.org/10.1007/s12237-009-9169-5.
Okolodkov, Y.B., Gárate-Lizárraga, I., 2006. An annotated checklist of dinoflagellatesfrom the Mexican Pacific. Acta Botánica Mexicana 74, 1–154.
Pezzolesi, L., Cucchiari, E., Guerrini, F., Pasteris, A., Galletti, P., Tagliavini, E., Totti, C.,Pistocchi, R., 2010. Toxicity evaluation of Fibrocapsa japonica from the NorthernAdriatic Sea through a chemical and toxicological approach. Harmful Algae 9,504–514.
Vargas-Montero, M., Freer-Bustamante, E., Jiménez-Montealegre, R., Guzmán, J.C.,2006. Occurrence and predominance of the fish killer Cochlodinium polykrikoideson the Pacific coast of Costa Rica. Afr. J. Mar. Sci. 28, 215–217.
Yasumoto, T., Satake, M., Fukui, M., Nagai, H., Murata, M., Legrand, A.M., 1993. ATurning Point in Ciguatera Study. In: Smayda, T.J., Shimizu, Y. (Eds.), ToxicPhytoplankton in the Sea. Elsevier, Amsterdam, pp. 455–461.
Yuki, K., Yoshimatsu, S., 1989. Two fish-killing species of Cochlodinium fromHarima-Nada, Seto Inland Sea, Japan. In: Okaichi, T., Anderson, D., Nemoto, T.(Eds.), Red Tides: Biology, Environmental Science, and Toxicology, Elsevier, NewYork, pp. 451–454.