50
Cell Effects on Mechanical Properties of Environment Morgan Boes
| Date post: | 22-Dec-2015 |
| Category: |
Documents |
| Upload: | lisa-bryant |
| View: | 218 times |
| Download: | 0 times |

Cell Effects on Mechanical Properties of Environment
Morgan Boes

Sources
• Beningo, K. A., Dembo, M., Kaverina, I., Small, J. V., & Wang, Y.-l. (2001). Nascent Focal Adhesions are Responsible for the Generation of Strong Propulsive Forces in Migrating Fibroblasts. The Journal of Cell Biology , 153 (4), 881-7.
• Dobereiner, H.-G., Dubin-Thaler, B. J., Giannone, G., & Sheetz, M. P. (2005). Force sensing and generation in cell phases: analysis of complex functions. Journal of Applied Physiology , 98, 1542-6.
• Wang, J. H., & Li, B. (2010). Mechanics rules cell biology. Sports Medicine, Arthroscopy, Rehabilitation, Therapy, & Technology , 2 (16).

Mechanical Forces
• a push or pull exerted by the cell• a push or pull encountered by the cell

“Mechanics rules cell biology”
Cells in the musculoskeletal system are subjected to various mechanical forces in vivo. Years of research have shownthat these mechanical forces, including tension and compression, greatly influence various cellular functions such asgene expression, cell proliferation and differentiation, and secretion of matrix proteins. Cells also usemechanotransduction mechanisms to convert mechanical signals into a cascade of cellular and molecular events.
An overview of cell mechanobiology to highlight the notion that mechanics, mainly in the formof mechanical forces, dictates cell behaviors in terms of both cellular mechanobiological responses andmechanotransduction.
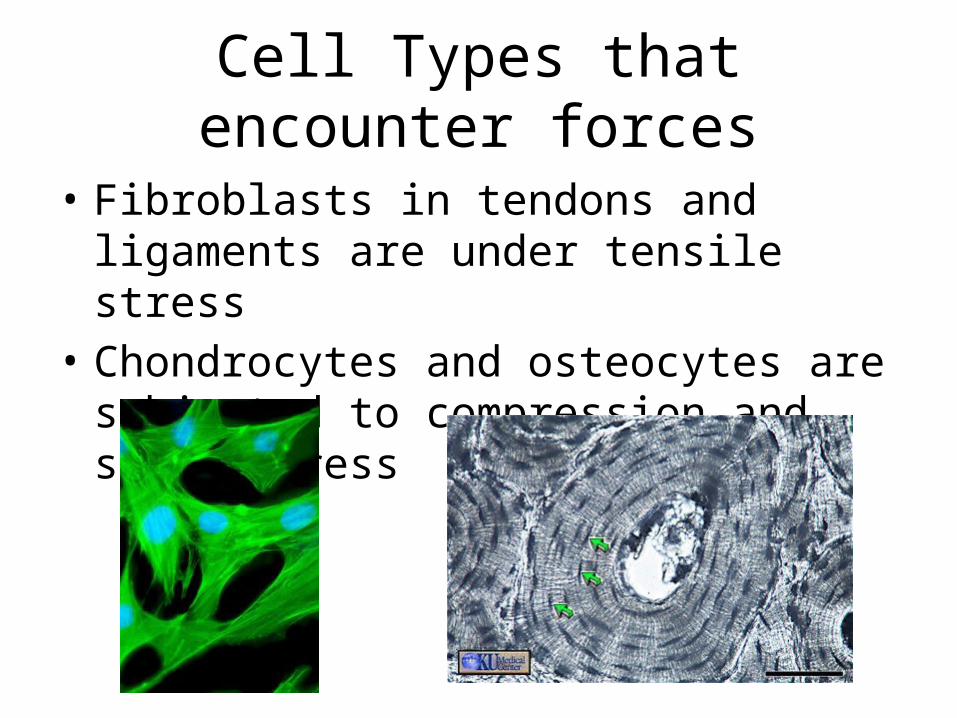
Cell Types that encounter forces
• Fibroblasts in tendons and ligaments are under tensile stress
• Chondrocytes and osteocytes are subjected to compression and shear stress

Internal Mechanical Forces
• Forces generated by the cells themselves– Considered intracellular tension– In non-muscle cells this is created by cross-linking of
actomyosin.• These forces are then transmitted to the ECM via
focal adhesions– These forces are called cell traction forces (CTFs)– CTFs direct ECM assembly, control cell shape, permit
cell movement, and maintain cellular tensional homeostasis.
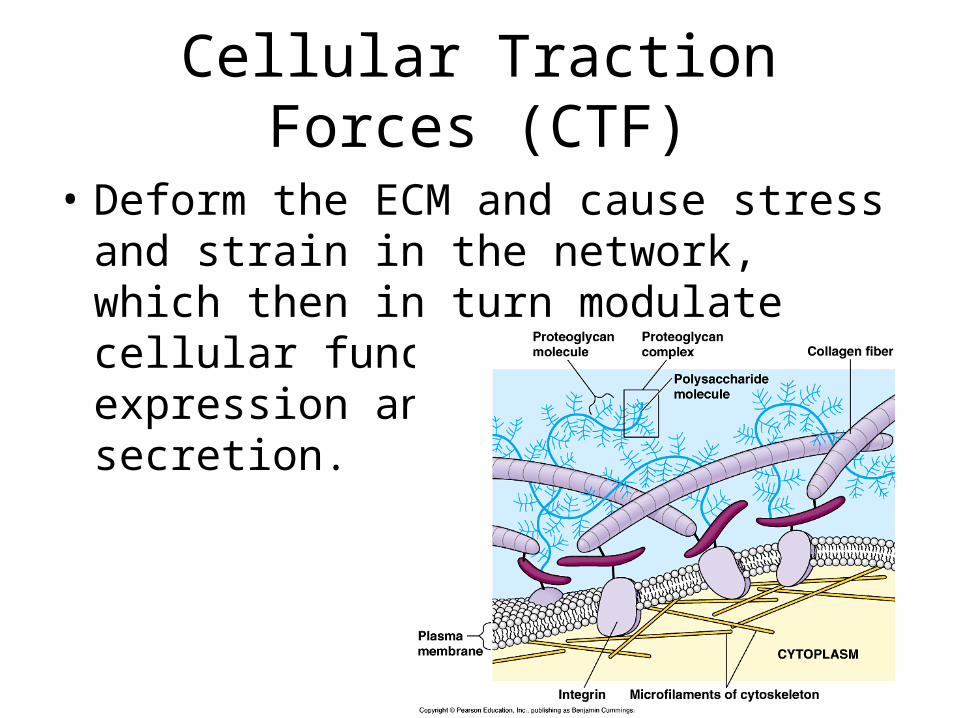
Cellular Traction Forces (CTF)
• Deform the ECM and cause stress and strain in the network, which then in turn modulate cellular functions such as gene expression and protein secretion.

• Cells can also use their internal contractile forces to regulate their own proliferation and differentiation.
• Internal mechanical forces generated by cells themselves regulate cell biology in terms of metabolic state, cell proliferation and differentiation, etc. Especially when these CTFs are transmitted to the ECM, where they regulated many vital cellular functions such as migration and ECM assembly
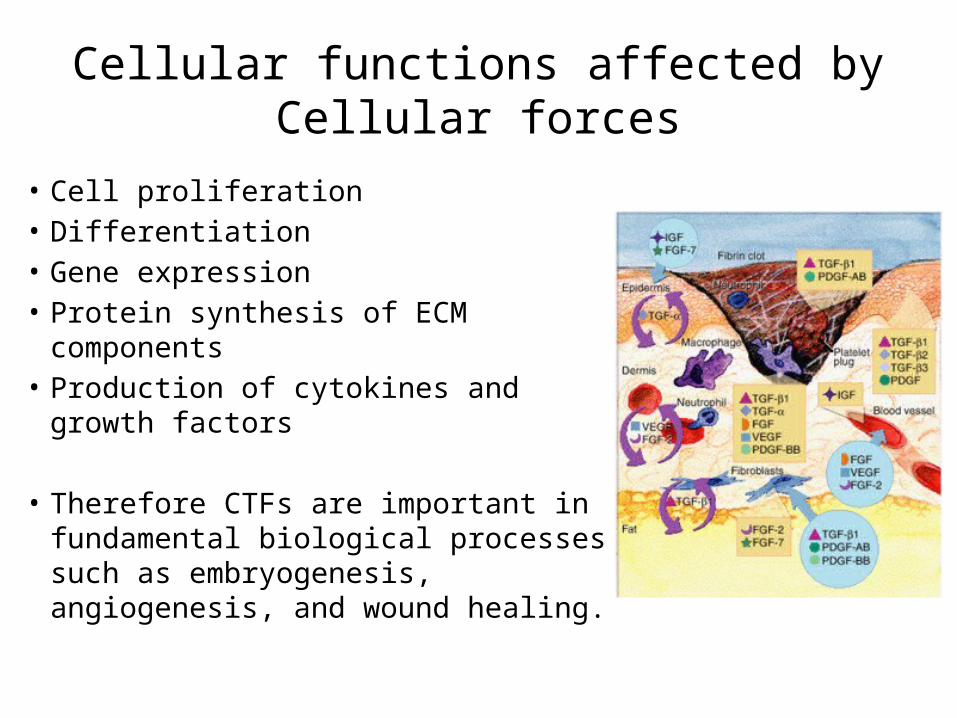
Cellular functions affected by Cellular forces
• Cell proliferation• Differentiation• Gene expression• Protein synthesis of ECM components• Production of cytokines and growth
factors
• Therefore CTFs are important in fundamental biological processes such as embryogenesis, angiogenesis, and wound healing.

Force sensing and generation in cell phases: analyses of complex functions
• Cellular morphology is determined by– Motility– Force sensing– Force generation

Two major explanations for motility
• Either cellular motility depends in a continuous fashion on cell composition
• Or it exhibits phases wherein only a few protein modules are activated locally for a given time.

• Observations of the behavior of cells can be dissected into functional pathways involving key proteins or protein groups that contribute specific function to the overall behavior
• Cells in suspension have a basal level of motility that enables the to probe their immediate environment.

Phase 1- “early spreading”
• Cell goes from a rough sphere to a thick disk on a 2D surface with about the same cross-sectional area
• During this phase there are three necessary steps• This phase is activated by a cell-wide process that
induces disassembly of filaments generally and spreading locally.

Phase 1 Step 1
• Local sensing of the matrix coating of the surface which activates– General breakdown of cortical actin filaments and
cortical structure– Local assembly of actin filaments at matrix-coated
surfaces• In this step there is a threshold to the activation
that is a function of both fibronectin density and time.– Higher concentrations decrease the lag time before
spreading instead of increasing the rate of spreading
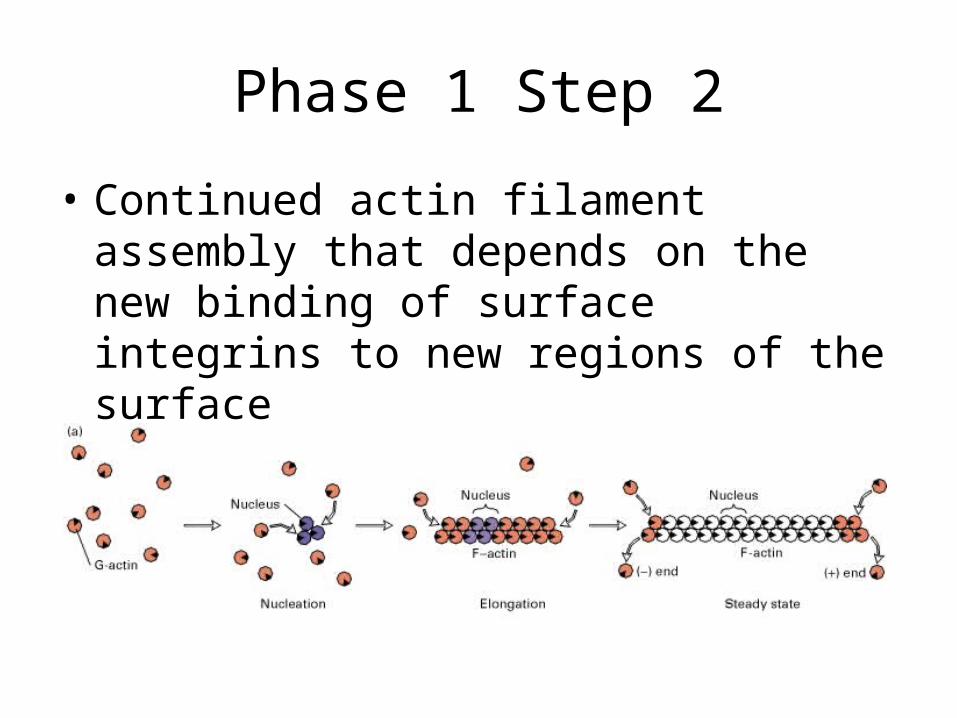
Phase 1 Step 2
• Continued actin filament assembly that depends on the new binding of surface integrins to new regions of the surface

Phase 1 Step 3
• Slow rearward transport of the newly assembled actin filaments.
• This phase will automatically stop when either the cell reaches a critical area or receives another dominant signal

Phase 2: Contractile Phase of Spreading
• Distinguished by a dramatic increase in the rate of rearward actin movement.
• Required for continued spreading• Rigid substrates are also required for
continued spreading• But how does the cell know if the substrate is
rigid?

Rigidity Sensing
• At these early stages of spreading, there are no stress fibers, thus it is difficult to understand how the cytoskeleton is organized to support force generation from one side to the other.
• The tension that the cell creates is clearly due to myosin, and the major question is how the myosin is organized to enable it to generate force to move actin inward and ultimately to generate force on the surround matrix

Ridgidity Sensing
• Rigidity sensing is a major aspect of the contractile phase, and fibroblasts need a rigid substrate to spread fully.
• Neurons prefer a softer substrate.• There are two theories on how the cell senses
the rigidity of the substrate nearby.
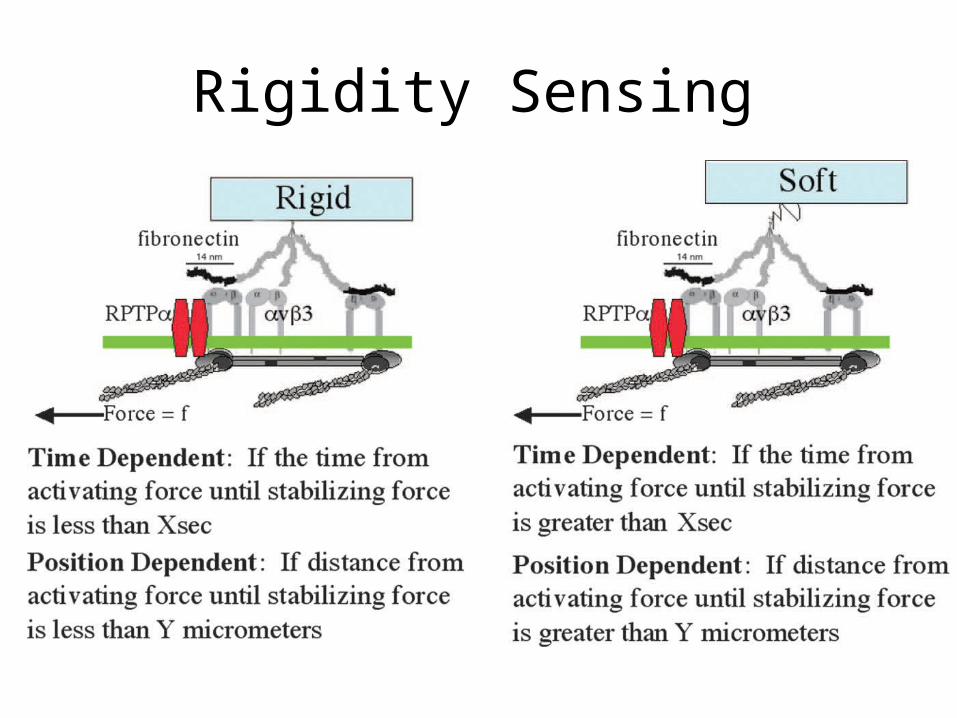
Rigidity Sensing

Phase 3
• On sufficiently stiff substrates, the cell continues spreading, approaching its maximal area of contact. After which the cell moves forward in a particular direction after polarization is triggered by either internal signals or external chemical gradients

• These phases are relatively general and applies to fibroblasts, endothelial cells, and presumably in a similar form to even neuronal growth cones
• Motility phases involve a characteristic subset of functions that are organized in specific spatial and temporal order. They involve distinct sets of protein modules
Phase Transitions
• How does the cell control the transitions between the fast spreading and retraction phases?

Initiating Cell spreading
• Characterized by an increase in the actin polymerization velocity at the leading edge of the lamellipodium, pushing the membrane forward. Increased polymerization is triggered by favorable contact with the ECM.
• Time from contact to spreading decreases with fibronectin density
Transition to periodic contractile phase
• Linked to the activity of myosin light chain kinase (MLCK), a protein control in turn the activity of myosin motors. But there is no direct evidence yet for the involvement of myosin.

Global Cell Phases
• The evolutionary pressure to survive preserves function but not necessarily the associated protein modules.
• So by understanding cell mechanisms and their phases, we can compare across cell types even if the proteins involved vary.
• To better define these phases, define basic functional protein modules

Basic functional modules example
• Regulatory proteins – controlled by a signaling network coordinating spatially distant and/or logically separate functional events in a cell– Must not interact directly with the structural
proteins

Nascent Focal Adhesions Are Responsible for the Generationof Strong Propulsive Forces in Migrating Fibroblasts

Focal Adhesions
• We know that focal adhesions tightly adhere to the extracellular matrix
• But what is their role in force transduction?

The role of focal adhesions in force transduction?
• To figure this out the researchers– Mapped traction stress generated by fibroblasts
expressing GFP-zyxin.• They found– The overall distribution of focal adhesions only
partially resembles the distribution of traction stresses

Leading edge of cells
• Faint small adhesions transmit strong forces• Large, bright, mature focal adhesions exert
weaker forces• This relationship is unique to the leading edge
of motile cells.

• Also traction forces decrease soon after the appearance of focal adhesions
• As focal adhesions mature, changes in structure, protein content, or phsophorylation may cause the focal adhesion to change its function from the transmission of strong propulsive forces, to a passive anchorage device for maintaining a spread cell morphology.

Focal Adhesions
• Involved in anchoring cells to the substrate• What about contractile forces that might be
transmitted through these structures to propel directional movements

• Hundreds of focal adhesions must be coordinated in order to maintain both the direction of migration and the morphology of the cell in an efficient manner. HOW?
• Solution – to generate maps of both dynamic focal adhesions under a migrating fibroblast, and traction forces that a cell exerts on the substrate

Traction stress Measurement
• Tracked by embedding beads into a flexible substrate that the cells are grown on.
• Then using large-scale matrix computations, convert bead movements of substrate deformation to maps of traction stress.

Traction stress calculations
• Plated cells on collagen I –coated flexible polyacrylamide substrates.
• The confinement of traction stress was within the cell boundary, global balance of forces and torques was required
• Used a Monte Carlo simulation to determine force balance.
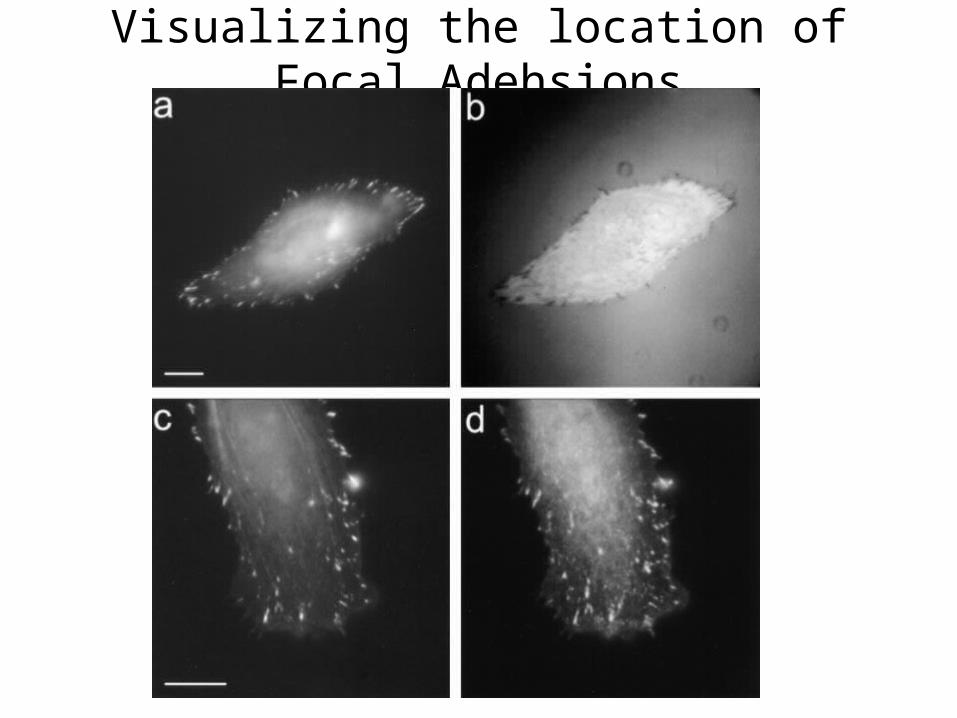
Visualizing the location of Focal Adehsions
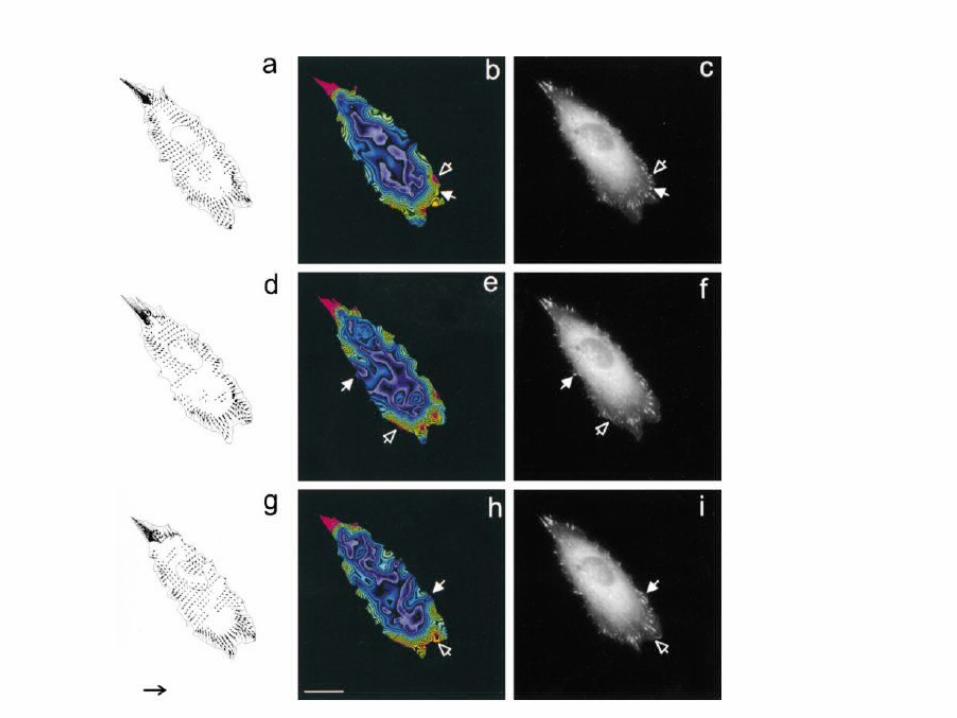

Results
• Small, nascet focal adhesions at the leading edge exert transient forces to move the cell forward.
• Mature focal adhesions serve primarily as anchors to the substrate.
• Allows fibroblasts to migrate efficiently and responsively without complex coordination of the mechanical output among the adhesion foci
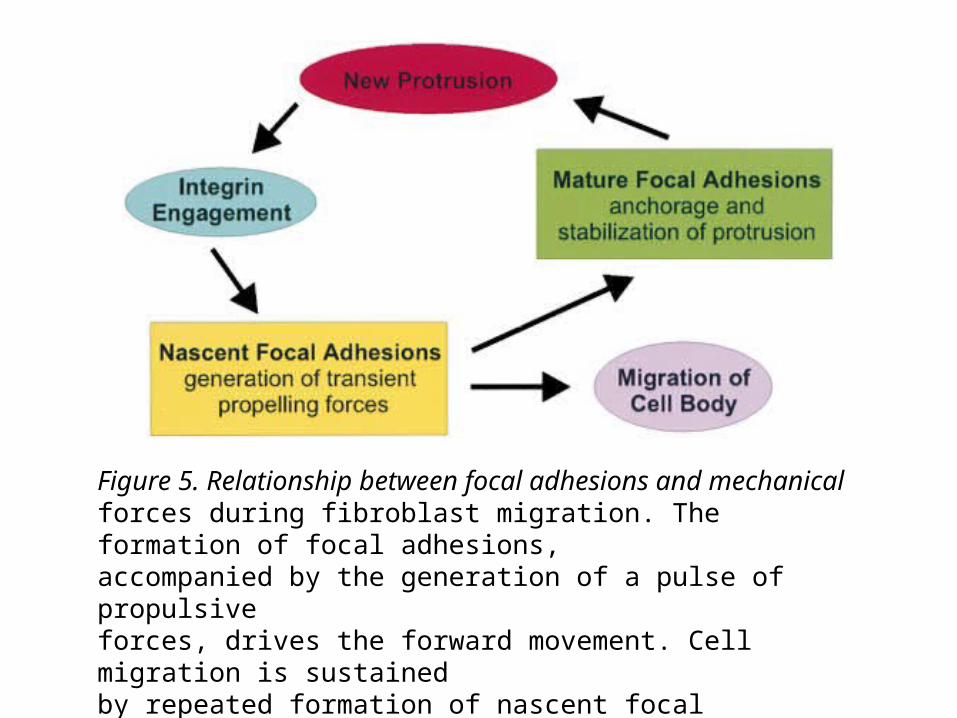
Figure 5. Relationship between focal adhesions and mechanicalforces during fibroblast migration. The formation of focal adhesions,accompanied by the generation of a pulse of propulsiveforces, drives the forward movement. Cell migration is sustainedby repeated formation of nascent focal adhesions, and thus repeatedpulses of propulsive forces. Mature focal adhesions playonly a passive role in anchoring cells to the substrate.

Advantages to this approach 1
• A division of labor between propulsive adhesions and anchorage at the leading edge which allows the cell to migrate while maintaining its spread morphology.

Advantages to this approach 2
• Since cell migration is driven by transient pulses of propulsive forces in the leading lamella, minimal coordination is required among mechanical interactions at a multitude of focal adhesions

Advantages to this approach 3
• This mechanism facilitates rapid reorientation in response to environmental cues, simply by shifting assembly of nascent focal adhesions to a new protrusive region.

Questions?

Sources
• Beningo, K. A., Dembo, M., Kaverina, I., Small, J. V., & Wang, Y.-l. (2001). Nascent Focal Adhesions are Responsible for the Generation of Strong Propulsive Forces in Migrating Fibroblasts. The Journal of Cell Biology , 153 (4), 881-7.
• Dobereiner, H.-G., Dubin-Thaler, B. J., Giannone, G., & Sheetz, M. P. (2005). Force sensing and generation in cell phases: analysis of complex functions. Journal of Applied Physiology , 98, 1542-6.
• Wang, J. H., & Li, B. (2010). Mechanics rules cell biology. Sports Medicine, Arthroscopy, Rehabilitation, Therapy, & Technology , 2 (16).

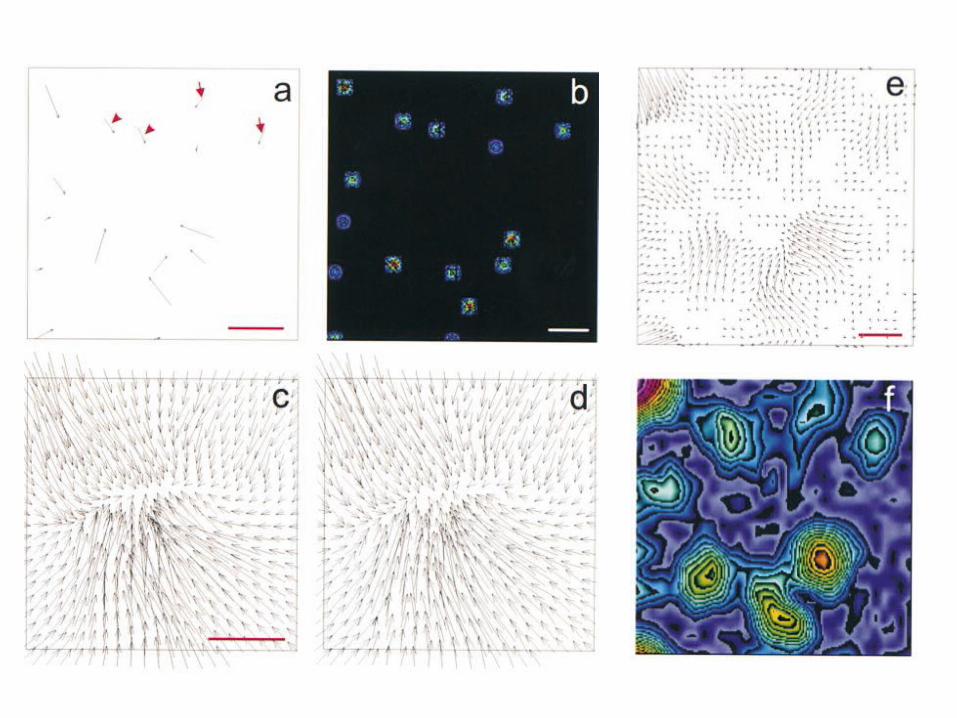
Figure 1. Monte Carlo simulation of traction stressanalysis. Small patches of traction stress were firstassigned at random locations within a square area (aand b). Exact deformation matrix was generatedfrom this map at a finite resolution and density (c).After applying random noise and neighborhood averagingto mimic the resolution limit of the measurements(d), the modified deformation matrix wasused to calculate the original traction stress (e andf). A pair of forces separated by z4.4 mm appears asa single large patch (arrowheads), whereas a pairseparated by z8.0 mm is clearly resolved (arrows).Panels b and f show color rendering of the magnitude,with red corresponding to strong tractionstress and blue corresponding to weak tractionstress

Figure 2. GFP-zyxin as a marker for focal adhesions.Fish fin fibroblasts transfected with GFPzyxinwere plated on collagen-coated coverslips.IRM (b) shows the localization of GFP-zyxin atboth large and small focal adhesions (a). Immunofluorescencestaining of paxillin (d) shows a similarcolocalization with GFP-zyxin (c) at the leadingedge. Bars, 10 mm

Figure 3. Differences between the distribution oftraction stress and focal adhesions.Distributions of traction stress at 0, 6, and 10 minare shown as either vector maps (a, d, and g), or colorimages after converting the magnitude into colors (b, e,and h). The corresponding distributions of GFP-zyxinshow only a limited correlation with traction stress (c, f,and i). Some focal adhesions contain a low concentrationof GFP-zyxin but generate strong forces (open arrow),whereas other focal adhesions show strong GFP-zyxin localization but generate relativelyweak forces (filled arrows). Arrow in g, 105 dyn/cm2. Bar,20 mm.

















