APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Oct. 1978, p. 606-612 0099-2240/78/0036-0606$02.00/0 Copyright ( 1978 American Society for Microbiology Vol. 36, No. 4 Printed in U.S.A. Cellulolytic Enzyme System of Thermoactinomyces sp. Grown on Microcrystalline Cellulose BARBEL G. R. HAGERDAL,t* JOHN D. FERCHAK, AND E. KENDALL PYE Department of Biochemistry and Biophysics, School of Medicine, University of Pennsylvania, Philadelphia, Pennsylvania 19104 Received for publication 15 August 1978 The carboxymethyl-cellulase and Avicelase activities of Thermoactinomyces sp., strain YX, were produced simultaneously with cell growth. Throughout the growth phase these activities were primarily extracellular, with up to 50% ad- sorbed to residual cellulosic substrate at any one time. On the other hand, the ,8-glucosidase activity was associated with the culture solids throughout the entire fermentation and appears to be intracellular. Preparative isoelectric focusing of the culture filtrate, in the pH range 3 to 5, separated three major fractions with cellulolytic activities towards both carboxymethyl-cellulose and Avicel. Based on the carboxymethyl-cellulase and Avicelase activities of these separated fractions, it was not possible to discriminate between endo- and exoglucanases produced by Thermoactinomyces sp. Analytical isoelectric focusing of culture filtrates obtained throughout the growth phase of Thermoactinomyces indicated that all the extracellular cellulolytic enzymes are produced and released into the culture filtrate simultaneously, with no evidence of a sequential appearance of the various enzymes or isoenzymes. When grown on 1% cellulose (Avicel; FMC Corp., Newark, Del.) at 550 C the thermophilic filamentous bacteria Thermoactinomyces de- grade 70% of the substrate within 24 h (11). The fast degradation of cellulosic substrates indicates that the cellulase enzyme system of Thermoac- tinomyces sp. has great potential for the sac- charification of cellulosic materials (19). The extracellular enzyme system of Ther- moactinomyces sp. has so far been studied mainly with respect to "total" cellulase activity measured either as reducing sugar produced from a 20% Avicel slurry (19) or as filter paper activity (13; A. Moreira, Ph.D. thesis, University of Pennsylvania, Philadelphia, 1977). Only re- cently was it discovered that the extracellular enzyme system lacks f-glucosidase and that, at the onset of the stationary growth phase, this enzyme activity is associated almost exclusively with the culture solids (B. Hagerdal, H. Harris, and E. K. Pye, Biotechnol. Bioeng., in press). The present study was undertaken to further investigate the endoglucanase, exoglucanase, and ,8-glucosidase activities throughout an entire growth phase of Thermoactinomyces sp. MATERIALS AND METHODS Culture conditions and fractionation of whole culture broth. A culture of Thermoactinomces sp. t Present address: Biochemistry 2, Chemical Center, S- 22007 Lund, Sweden. (YX strain, obtained from the General Electric Co.) was grown in a well-aerated 14-liter fermentor (New Brunswick Scientific Corp., New Brunswick, N.J.) con- taining medium composed of the following: Avicel (PH 102, FMC Corp.), 95 g; NaCl, 15 g; (NH4)2SO4, 31 g; thiamine hydrochloride, 10 mg; biotin, 10 mg; Na2HPO4, 91 g; KH2PO4, 9.0 g; yeast extract (Difco Laboratories, Detroit, Mich.), 10 g; water, 9.8 liters; and salts solution (2.5 g of disodium ethylenediamine- tetraacetic acid, 10 g of MgS04 7H20, 0.4 g of ZnS04 7H20, 1.0 g of FeSO4 7H20, 0.76 g of MnSO4 H20, 1.0 g of CaCl2, in 1 liter of water), 200 ml. The medium was autoclaved for 45 min, cooled to 55°C, and inoculated with 700 ml of shake flask cul- ture, grown for 48 to 72 h. The temperature was held constant at 55°C. The pH was not controlled and dropped from 7.4 to 6.5 during the fermentation. Samples of the culture were taken throughout the exponential growth phase and into the stationary growth phase. Cell growth was stopped by the addition of sodium azide (Fisher Scientific Co., Pittsburgh, Pa. to a 0.03% final concentration. Each sample (40 ml) was separated in the cold on a glass filter (Whatman GF/F) into culture solids (filamentous cell mass and residual cellulose) and culture filtrate. The culture solids were then washed two successive times on the filter with 10 ml of ice-cold 0.05 M phosphate buffer, pH 5.9, and resuspended in 40 ml of ice-cold 0.05 M phosphate buffer, pH 6.5. Thus, three different frac tions were obtained from each fermentation sample: the whole culture broth, the culture filtrate, and the culture solids. The fractions were assayed separately for hydrolytic activity against carboxymethyl-cellulose (CMC), Avicel, and p-nitrophenol-/3-D-glucoside. Sam- ples of the culture filtrates were also analyzed by 606 Downloaded from https://journals.asm.org/journal/aem on 12 February 2022 by 191.240.112.253.
Transcript
APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Oct. 1978, p. 606-6120099-2240/78/0036-0606$02.00/0Copyright ( 1978 American Society for Microbiology
Vol. 36, No. 4
Printed in U.S.A.
Cellulolytic Enzyme System of Thermoactinomyces sp. Grownon Microcrystalline Cellulose
BARBEL G. R. HAGERDAL,t* JOHN D. FERCHAK, AND E. KENDALL PYEDepartment of Biochemistry and Biophysics, School ofMedicine, University of Pennsylvania, Philadelphia,
Pennsylvania 19104
Received for publication 15 August 1978
The carboxymethyl-cellulase and Avicelase activities of Thermoactinomycessp., strain YX, were produced simultaneously with cell growth. Throughout thegrowth phase these activities were primarily extracellular, with up to 50% ad-sorbed to residual cellulosic substrate at any one time. On the other hand, the,8-glucosidase activity was associated with the culture solids throughout the entirefermentation and appears to be intracellular. Preparative isoelectric focusing ofthe culture filtrate, in the pH range 3 to 5, separated three major fractions withcellulolytic activities towards both carboxymethyl-cellulose and Avicel. Based onthe carboxymethyl-cellulase and Avicelase activities of these separated fractions,it was not possible to discriminate between endo- and exoglucanases produced byThermoactinomyces sp. Analytical isoelectric focusing of culture filtrates obtainedthroughout the growth phase of Thermoactinomyces indicated that all theextracellular cellulolytic enzymes are produced and released into the culturefiltrate simultaneously, with no evidence of a sequential appearance of the variousenzymes or isoenzymes.
When grown on 1% cellulose (Avicel; FMCCorp., Newark, Del.) at 550 C the thermophilicfilamentous bacteria Thermoactinomyces de-grade 70% of the substrate within 24 h (11). Thefast degradation of cellulosic substrates indicatesthat the cellulase enzyme system of Thermoac-tinomyces sp. has great potential for the sac-charification of cellulosic materials (19).The extracellular enzyme system of Ther-
moactinomyces sp. has so far been studiedmainly with respect to "total" cellulase activitymeasured either as reducing sugar producedfrom a 20% Avicel slurry (19) or as filter paperactivity (13; A. Moreira, Ph.D. thesis, Universityof Pennsylvania, Philadelphia, 1977). Only re-cently was it discovered that the extracellularenzyme system lacks f-glucosidase and that, atthe onset of the stationary growth phase, thisenzyme activity is associated almost exclusivelywith the culture solids (B. Hagerdal, H. Harris,and E. K. Pye, Biotechnol. Bioeng., in press).The present study was undertaken to furtherinvestigate the endoglucanase, exoglucanase,and ,8-glucosidase activities throughout an entiregrowth phase of Thermoactinomyces sp.
MATERIALS AND METHODSCulture conditions and fractionation of whole
culture broth. A culture of Thermoactinomces sp.
t Present address: Biochemistry 2, Chemical Center, S-22007 Lund, Sweden.
(YX strain, obtained from the General Electric Co.)was grown in a well-aerated 14-liter fermentor (NewBrunswick Scientific Corp., New Brunswick, N.J.) con-taining medium composed of the following: Avicel (PH102, FMC Corp.), 95 g; NaCl, 15 g; (NH4)2SO4, 31 g;thiamine hydrochloride, 10 mg; biotin, 10 mg;Na2HPO4, 91 g; KH2PO4, 9.0 g; yeast extract (DifcoLaboratories, Detroit, Mich.), 10 g; water, 9.8 liters;and salts solution (2.5 g of disodium ethylenediamine-tetraacetic acid, 10 g of MgS04 7H20, 0.4 g ofZnS04 7H20, 1.0 g of FeSO4 7H20, 0.76 g ofMnSO4 H20, 1.0 g of CaCl2, in 1 liter of water), 200ml. The medium was autoclaved for 45 min, cooled to55°C, and inoculated with 700 ml of shake flask cul-ture, grown for 48 to 72 h. The temperature was heldconstant at 55°C. The pH was not controlled anddropped from 7.4 to 6.5 during the fermentation.
Samples of the culture were taken throughout theexponential growth phase and into the stationarygrowth phase. Cell growth was stopped by the additionof sodium azide (Fisher Scientific Co., Pittsburgh, Pa.to a 0.03% final concentration. Each sample (40 ml)was separated in the cold on a glass filter (WhatmanGF/F) into culture solids (filamentous cell mass andresidual cellulose) and culture filtrate. The culturesolids were then washed two successive times on thefilter with 10 ml of ice-cold 0.05 M phosphate buffer,pH 5.9, and resuspended in 40 ml of ice-cold 0.05 Mphosphate buffer, pH 6.5. Thus, three different fractions were obtained from each fermentation sample:the whole culture broth, the culture filtrate, and theculture solids. The fractions were assayed separatelyfor hydrolytic activity against carboxymethyl-cellulose(CMC), Avicel, and p-nitrophenol-/3-D-glucoside. Sam-ples of the culture filtrates were also analyzed by
606
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 12
Feb
ruar
y 20
22 b
y 19
1.24
0.11
2.25
3.
CELLULOLYTIC ENZYMES OF THERMOACTINOMYCES SP. 607
isoelectric focusing. To minimize prQteolytic degra-tion, these samples were lyophilized immediately.
Cultures of Thermoactinomyces were also grown in500-ml shake flasks, each holding a total of 100 ml ofculture, using 5 ml of frozen inoculum, in a NewBrunswick rotary shaking water bath set at 175 rpm.The cultures were harvested during the exponentialgrowth phase, and samples of 25 ml were separated inthe cold on a glass filter as described above. Theculture solids were washed five successive times with5 ml of ice-cold water, each wash fraction was collectedseparately, and then culture solids were resuspendedto their original volume in ice-cold 0.05 M phosphatebuffer, pH 6.5. The whole broth, the culture filtrate,the culture solids, and the five wash fractions wereassayed for the enzyme activities listed above. Theculture filtrate and the wash fractions were lyophilizedand analyzed by isoelectric focusing.
Protein determination. The protein concentra-tion was either estimated by reading the absorbanceat 280 nm or determined by the method of Lowry etal. (12), using bovine serum albumin, fraction V (SigmaChemical Co., St. Louis, Mo.), as a standard. Reducingsugars (RS) in the culture filtrates never exceeded 0.1mg/ml and thus did not interfere in the protein assay.Enzyme assays. (i) Carboxymethyl (CM)-cel-
lulase (endoglucanase/1,4-,/-glucan glucanohy-drolase [EC 3.2.1.4]) activity. The CMC (4M6SF;Hercules Inc., Wilmington, Del.) had the followingproperties: degree of substitution, 0.38 to 0.48; degreeof polymerization, 1,100; molecular weight, 250,000;medium viscosity, 300- to 600-cP maximum viscosityfor a 2% solution; special solution characteristics; foodgrade. CMC, 0.5%, in 0.05 M phosphate buffer, pH 5.9,was used in a modification of the assay described byBerghem and Pettersson (3). A 2.0-ml amount of thissubstrate solution was incubated with enzyme solutionat 65°C for 10 min. The amount of RS produced wasdetermined according to Miller et al. (15), with glucose(Fisher Scientific Co.) as a standard.
(ii) Avicelase (exoglucanase/1,4-f8-glucan cel-lobiohydrolase [EC 3.2.1.-]) activity. Two differentsubstrates were used in a modification of the assaydescribed by Berghem and Pettersson (3). Avicel PH102 was used to assay the culture filtrate samplesharvested during the fermentation, and Avicel PH 105was used to assay the fractions separated in the pre-parative isoelectric focusing and also the fractionsobtained in the desorption study. For Avicel PH 102the incubation time was 2 h, and for Avicel PH 105 itwas 20 min. A 3.5-ml amount of a 0.5% suspension ofthe substrate in 0.05 M phosphate buffer, pH 7.1, wasincubated with enzyme solution at 65°C under contin-uous shaking. The reaction was stopped by placing thereaction tube in a boiling-water bath for 5 min. Afterthe mixture was filtered (Whatman GF/D), theamount of RS produced in the filtrate was determinedby the Somogyi (18) modification of the method ofNelson (17), with glucose as a standard.
(iii) ,8-Glucosidase (cellobiase [EC 3.2.1.21]) ac-tivity. p-Nitrophenol-,8-D-glucoside (Sigma ChemicalCo.) was used as the substrate for the assay previouslydescribed (Hagerdal et al., in press).
Analytical isoelectric focusing. The lyophilizedculture filtrates and wash fractions were redissolved in
water to a final protein concentration of about 0.5mg/ml and were then desalted in the cold on a PD-10column (Pharmacia Fine Chemicals AB, Uppsala,Sweden). The desalted samples were immediately fro-zen on dry ice and thawed just before applying themto the polyacrylamide gel slabs. The isoelectric focus-ing was carried out with a Multiphor apparatus (LKB-Produkter AB, Bromma, Sweden). Either commercialpolyacrylamide gel plates (Ampholine PAG plates)with a carrier gradient of pH 3.5 to 9.5 or polyacryl-amide gel plates prepared to have a narrow carriergradient were used. The composition of these gels was:total concentration of monomers added, 5%; cross-linking concentration, 3%. Ammonium persulfate wasused as polymerizer. A 1.2-ml portion of Ampholine(LKB-Produkter AB), pH 2.5 to 4, 1.2 ml of Ampho-line, pH 4 to 6, and 1.2 ml of Ampholine, pH 5 to 7, ina total volume of 60 ml of water also holding 6.1 ml of99.6% glycerol (Fisher Scientific Co.) gave a carriergradient of pH 2.5 to 6.The enzyme samples were applied to the surface of
the gel plate with two pieces (0.5 by 1 cm2) of glassfilter paper (Whatman GF/A). The separations weremade at 10°C for 1.5 h in the wide carrier gradient andfor 3 h in the narrow carrier gradient. A constantpower of 1 W/cm was applied. Fixing and stainingwere performed according to Winter et al. (20), using0.1% Coomassie brilliant blue R240 (Sigma ChemicalCo.).
Preparative isoelectric focusing. Culture fil-trates harvested in the late exponential growth phasewere separated by using preparative isoelectric focus-ing. The samples were concentrated 5 to 10 times bylyophilization and desalted on a Sephadex G-25 col-umn. The desalted protein fraction was again lyophi-lized.The preparative flat-bed isoelectric focusing was
carried out in a granulated gel (Ultrodex, LKB-Pro-dukter AB), using the same Multiphor equipment asused for the analytical isoelectric focusing. The sepa-ration was carried out according to Winter et al. (21)in a pH 3 to 5 gradient for 48 h at 10°C. The gradientwas made up with 0.5 ml of Ampholine, pH 2.5 to 4, 4ml of Ampholine, pH 3.5 to 4, and 0.5 ml of Ampholine,pH 5 to 7, in a total volume of 100 ml. The lyophilized,desalted enzyme sample was redissolved in 3 ml of thesame Ampholine solution and applied as a zone afterthe water content of the gel bed had been reduced tothe proper value by evaporation. When the focusingwas completed the gel bed was sectioned into 30fractions with a grid, and the pH of every third fractionwas determined with a surface electrode (Ingold LOT403-30 M8).The separated zones were collected as described by
Fagerstam et al. (7), using ice-cold 0.05 M phosphatebuffer, pH 7.1. The carrier ampholytes were not re-moved before the fractions were assayed for CM-cel-lulase and Avicelase activity.
RESULTSExtracellular protein and pH develop-
ment. When a culture of Thermoactinomycessp. was grown on 1% Avicel at 550C, using a 7%inoculum, the extracellular protein concentra-tion remained at about 0.2 mg/ml through a lag
VOL. 36, 1978
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 12
Feb
ruar
y 20
22 b
y 19
1.24
0.11
2.25
3.
608 HAGERDAL, FERCHAK, AND PYE
phase of approximately 6 h (Fig. 1), then in-creased rapidly over the next 6 h to about 1.2mg/ml, and reached a maximum of about 1.7mg/ml after 24 h.The starting pH during such a fermentation
was 7.4; it dropped to 7.2 during the first 6 h andto 6.7 over the next 6 h. The pH reached aconstant level of around 6.6 after about 24 h(Fig. 1). Thus, the decrease in pH is closelyrelated to the release of extracellular protein anddescribes an initial lag phase, an exponentialphase, and a stationary phase.CM-cellulase activity. The CM-cellulase ac-
tivities of the whole broth, culture filtrate, andculture solids are shown in Fig. 2. The develop-ment of the whole-broth and the culture filtrateactivities is closely related to the release of theextracellular protein. The activity of the culturefiltrate is lower throughout the lag and exponen-tial growth phases but becomes equal to that of
16755
+ ~~~~~~~~~~~~~~~~~~12
7 0
0.8
10 20 30 40 50
TIME (hours)
FIG. 1. pH and extracellular protein developmentin a culture of Thermoactinomyces sp., grown on 1%Avicel (PH 102) at 550C in a 14-liter batch fermentor,using a 79o shake flask inoculum. Symbols: (+) pH;(O) protein.
E.Ea:
E
TIME (hours)
FIG. 2. Development of CM-cellulase activity.Same conditions as for Fig. 1. Symbols: (O) wholeculture broth; (0) culture filtrate; (0) culture solids.
TIME (hours)
FIG. 3. Avicelase activity in culture filtrate. Sameconditions as for Fig. 1.
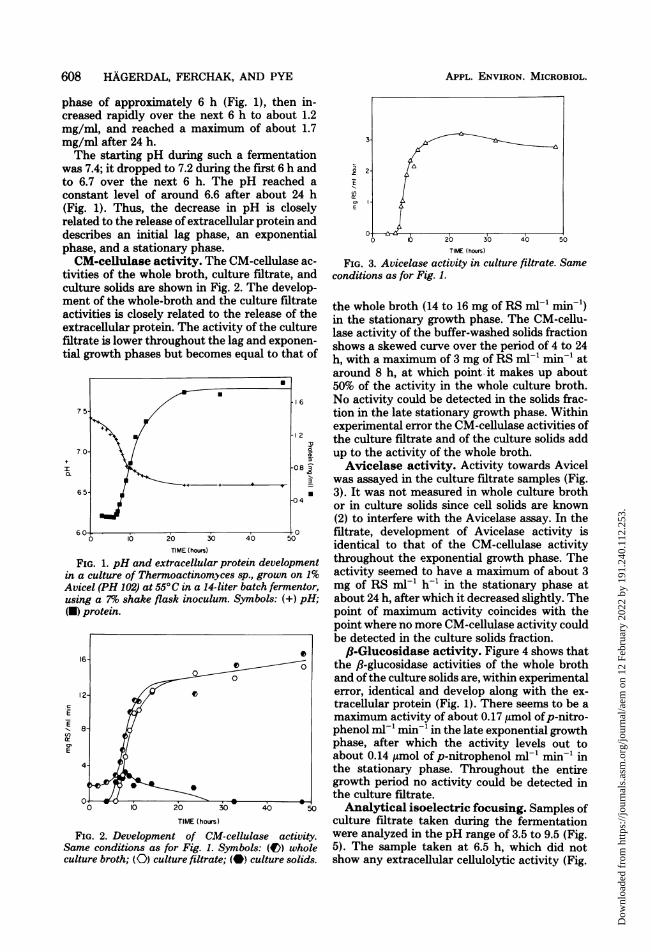
the whole broth (14 to 16 mg of RS mn-l min-x)in the stationary growth phase. The CM-cellu-lase activity of the buffer-washed solids fractionshows a skewed curve over the period of 4 to 24h, with a maximum of 3 mg of RS ml-' min-' ataround 8 h, at which point. it makes up about50% of the activity in the whole culture broth.No activity could be detected in the solids frac-tion in the late stationary growth phase. Withinexperimental error the CM-cellulase activities ofthe culture filtrate and of the culture solids addup to the activity of the whole broth.Avicelase activity. Activity towards Avicel
was assayed in the culture filtrate samples (Fig.3). It was not measured in whole culture brothor in culture solids since cell solids are known(2) to interfere with the Avicelase assay. In thefiltrate, development of Avicelase activity isidentical to that of the CM-cellulase activitythroughout the exponential growth phase. Theactivity seemed to have a maximum of about 3mg of RS ml-' h-' in the stationary phase atabout 24 h, after which it decreased slightly. Thepoint of maximum activity coincides with thepoint where no more CM-cellulase activity couldbe detected in the culture solids fraction.,B-Glucosidase activity. Figure 4 shows that
the 8l-glucosidase activities of the whole brothand of the culture solids are, within experimentalerror, identical and develop along with the ex-tracellular protein (Fig. 1). There seems to be amaximum activity of about 0.17 limol ofp-nitro-phenol ml-' min-' in the late exponential growthphase, after which the activity levels out toabout 0.14 ,umol of p-nitrophenol ml-' min-' inthe stationary phase. Throughout the entiregrowth period no activity could be detected inthe culture filtrate.Analytical isoelectric focusing. Samples of
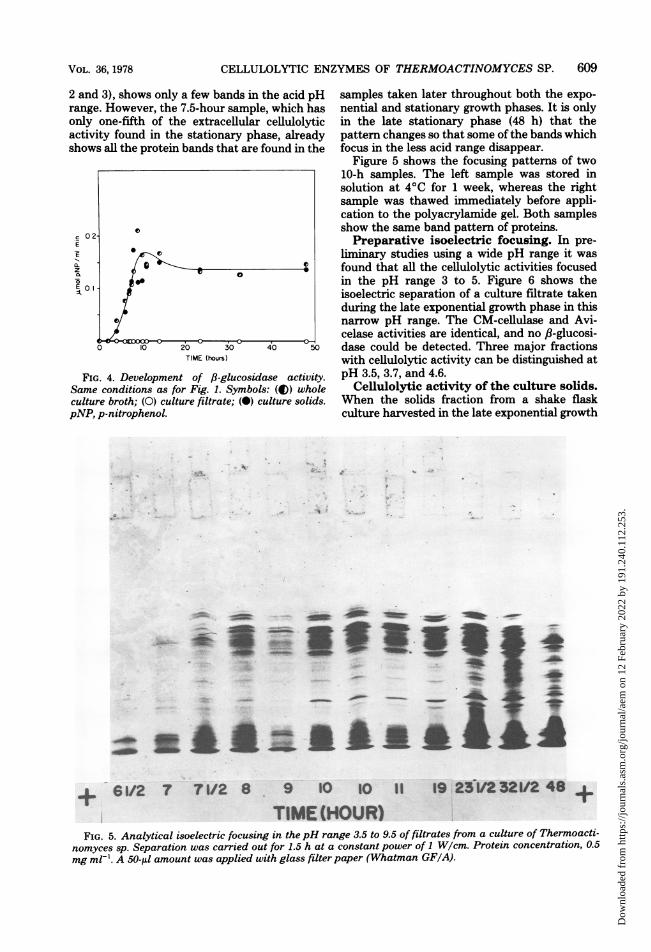
culture filtrate taken during the fermentationwere analyzed in the pH range of 3.5 to 9.5 (Fig.5). The sample taken at 6.5 h, which did notshow any extracellular cellulolytic activity (Fig.
APPL. ENVIRON. MICROBIOL.
'EI
E
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 12
Feb
ruar
y 20
22 b
y 19
1.24
0.11
2.25
3.
CELLULOLYTIC ENZYMES OF THERMOACTINOMYCES SP. 609
2 and 3), shows only a few bands in trange. However, the 7.5-hour sampleonly one-fifth of the extracellularactivity found in the stationary phsshows all the protein bands that are f
IEE
CzE
02
0.1
TIME (hours)
FIG. 4. Development of 83-glucosidcSame conditions as for Fig. 1. Symbolsculture broth; (0) culture filtrate; (0) cipNP, p-nitrophenol.
the acid pH samples taken later throughout both the expo->, which has nential and stationary growth phases. It is onlycellulolytic in the late stationary phase (48 h) that theise, already pattern changes so that some of the bands whichfound in the focus in the less acid range disappear.
Figure 5 shows the focusing pattems of two10-h samples. The left sample was stored insolution at 4°C for 1 week, whereas the rightsample was thawed immediately before appli-cation to the polyacrylamide gel. Both samplesshow the same band pattern of proteins.Preparative isoelectric focusing. In pre-
liminary studies using a wide pH range it was-4 found that all the cellulolytic activities focused
in the pH range 3 to 5. Figure 6 shows theisoelectric separation of a culture filtrate takenduring the late exponential growth phase in thisnarrow pH range. The CM-cellulase and Avi-
0celase activities are identical, and no ,B-glucosi-
40 5O dase could be detected. Three major fractionswith cellulolytic activity can be distinguished at
ase activity. pH 3.5, 3.7, and 4.6.s: (4)) whole Celiulolytic activity of the culture solids.ulture solids. When the solids fraction from a shake flask
culture harvested in the late exponential growth
.I I..
%O.. I.
.- &-..-
' 'Cs
I''g.
FIG. 5. Analytical isoelectric focusing in thepH range 3.5 to 9.5 of filtrates from a culture of Thermoacti-nomyces sp. Separation was carried out for 1.5 h at a constant power of 1 W/cm. Protein concentration, 0.5mg ml-'. A 50-,ul amount was applied with glass filter paper (Whatman GF/A).
0
so
< S0~~~~~
4r In -AC
ii11
A+6122 Va- +
VOL. 36, 1978
0
I
O.6. .i.. I
4 . - AL-71- *1 I
" 11. x
a :r ..3%,A
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 12
Feb
ruar
y 20
22 b
y 19
1.24
0.11
2.25
3.
610 HAGERDAL, FERCHAK, AND PYE
FRACTION
FIG. 6. Preparative isoelectric focusing of a de-salted lyophilized culture filtrate (50 mg of protein)from the late exponential growth phase of Thermoac-tinomyces sp. Separation was carried out for 48 h ata constant power of 8 W. Symbols: (0) CM-cellulaseactivity; (A) Avicelase activity; (+) pH.
TABLE 1. Total activities and specific activities ofCM-cellulase and Avicelase in a culture of
Thermoactinomyces sp., its subfractions, and washfractions of the culture solids
CM-cellulase Avicelase
Culture/fraction Total Totalactiv- actb aci c tbitya c ity'
a Milligrams of RS released per minute.b Milligrams of RS released per milligram of protein per
minute.
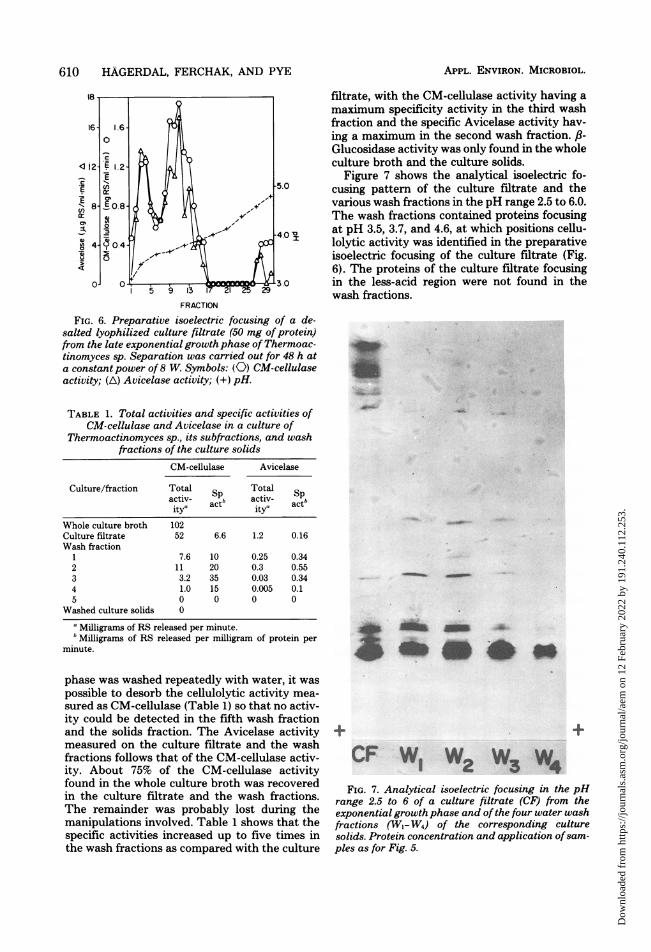
phase was washed repeatedly with water, it waspossible to desorb the cellulolytic activity mea-sured as CM-cellulase (Table 1) so that no activ-ity could be detected in the fifth wash fractionand the solids fraction. The Avicelase activitymeasured on the culture filtrate and the washfractions follows that of the CM-cellulase activ-ity. About 75% of the CM-cellulase activityfound in the whole culture broth was recoveredin the culture filtrate and the wash fractions.The remainder was probably lost during themanipulations involved. Table 1 shows that thespecific activities increased up to five times inthe wash fractions as compared with the culture
filtrate, with the CM-cellulase activity having amaximum specificity activity in the third washfraction and the specific Avicelase activity hav-ing a maximum in the second wash fraction. ,8-Glucosidase activity was only found in the wholeculture broth and the culture solids.
Figure 7 shows the analytical isoelectric fo-cusing pattern of the culture filtrate and thevarious wash fractions in the pH range 2.5 to 6.0.The wash fractions contained proteins focusingat pH 3.5, 3.7, and 4.6, at which positions cellu-lolytic activity was identified in the preparativeisoelectric focusing of the culture filtrate (Fig.6). The proteins of the culture filtrate focusingin the less-acid region were not found in thewash fractions.
FIG. 7. Analytical isoelectric focusing in the pHrange 2.5 to 6 of a culture filtrate (CF) from theexponential growth phase and of the four water washfractions (W,- W4) of the corresponding culturesolids. Protein concentration and application ofsam-ples as for Fig. 5.
APPL. ENVIRON. MICROBIOL.
0, "
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 12
Feb
ruar
y 20
22 b
y 19
1.24
0.11
2.25
3.
CELLULOLYTIC ENZYMES OF THERMOACTINOMYCES SP.
DISCUSSION
When a culture of Thermoactinomyces strainYX is grown on 1% Avicel in a 40-liter batchfermentor at pH 7.2, the cell growth and theextracellular cellulolytic activity, measured asfilter paper activity (13), occur simultaneously(Moreira, Ph.D. thesis, University of Pennsyl-vania, Philadelphia, 1977). The present study, inwhich the fermentation was carried out in asmaller fermentor (14 liters) and with an uncon-trolled pH, confirmed these results. The extra-cellular cellulolytic activity, measured as RSreleased from either CMC or Avicel, was pro-duced rapidly and approached a maximumw,ithin 18 to 24 h of growth, at which point theculture could be harvested to produce enzymesfor saccharification of cellulose. By comparison,Trichoderma viride produces extracellular cel-lulolytic activity only after the substrate hasbeen degraded; the activity reaches a maximumafter about 7 days (14). It has been suggestedthat the enzymes of T. viride are part of the cellwall during the exponential growth phase andare therefore not released into the culture fluiduntil the entire substrate has been degraded (2).A maximnum of 50% of the CM-cellulase activ-
ity of the whole broth was associated with theculture solids in the exponential growth phase.However, in the stationary growth phase, wherethe major part of the substrate has been di-gested, no more cellulolytic activity could bedetected in the culture solids. Similarly, it wasfound for Thermoactinomyces strain MJ0r,grown on 0.5% microcrystalline cellulose, thatthere is a lag before appearance of extracellularcellulolytic activity, as compared with the activ-ity in the whole culture broth (19). The presentstudy shows that this activity could be readilydesorbed when the solids fraction is washed withwater. Thus, the CM-cellulase activity is extra-cellular and not cell wall bound. The fact thatCM-cellulase activity of the solids fraction re-sults in a skewed curve that decreases to zero inthe stationary growth phase indicates that theactivity is probably adsorbed to the substraterather than to the cells. Such a relationship waspredicted by Moreira (Ph.D. thesis, Universityof Pennsylvania, Philadelphia, 1977) in a math-ematical model for the growth of Thermoacti-nomyces on cellulosic substrates.The selective adsorption to microcrystalline
cellulose of cellobiohydrolase (exoglucanase)from T. viride has been used to separate andpurify this enzyme from other cellulolytic activ-ities of the culture filtrate (4, 9). These studiesalso showed that the enzyme could be desorbedfrom the cellulose with water. Similarly, usinghigh salt concentrations, buffer, detergent, and
water, it was found that water desorbed thehighest amount of CM-cellulase and Avicelaseactivity from the culture solids of Thermoacti-nomyces sp. The high (up to fivefold) increaseof specific activity in the wash fractions indicatesa preferential adsorption of cellulolytic enzymesto cellulose, or a selective desorption. However,analytical isoelectric focusing of the wash frac-tions revealed a large number of protein bands,indicating that proteins other than cellulaseswere adsorbed to the culture solids. This wasalso found by Gum and Brown (9), when adsorp-tion to microcrystalline cellulose of the crudeculture filtrate from T. viride was used as a firstpurification step.We have recently shown that the ,B-glucosi-
dase activity from Thermoactinomyces is asso-ciated with the culture solids in samples har-vested at the onset of the stationary growthphase (Hagerdal et al., in press). The results alsoindicated that it is a soluble intracellular en-zyme. The present study shows that the fl-glu-cosidase activity is, in fact, associated with theculture solids throughout the entire growthphase, with no leakage into the culture filtrateeven in the late stationary phase. This supportsthe suggestion (Hagerdal et al., in press) thatthe 8-glucosidase produced by Thermoactino-myces is a truly intracellular enzyme. By com-parison, the ,8-glucosidase activity from T. virideseems to be tightly cell bound, with only about5% of the activity released into the culture fil-trate in the late stationary phase of the culture(2).The measurement of cellulolytic activity as
RS produced from CMC and Avicel has beenused to distinguish endoglucanases (CM-cellu-lases) from exoglucanases/cellobiohydrolases(Avicelases) produced by T. viride (3). However,when a desalted culture filtrate taken at theonset of the stationary phase of a Thermoacti-nomyces culture was separated by preparativeisoelectric focusing, the CM-cellulase and Avi-celase activities were found in the same frac-tions. Furthermore, the proportions of these ac-tivities in the different fractions were the same.Thus, it may be that the extracellular cellulolyticenzymes of Thermoactinomyces are less specificin their function than those produced by T.viride. These findings, however, also indicatethat more specific assays, such as a change inviscosity ofCMC (1) and identification of solublehydrolysis products by high-pressure liquidchromatography (10), should be used to char-acterize the different cellulolytic activities pro-duced by Thermoactinomyces.
Cellulolytic enzymes have been reported asvarying numbers of isoenzymes (4, 6, 10, 22). Arecent study indicated that the multiplicity of
VOL. 36, 1978 611
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 12
Feb
ruar
y 20
22 b
y 19
1.24
0.11
2.25
3.
612 HAGERDAL, FERCHAK, AND PYE
enzymes might in part be due to proteolyticdegradation as a result of the presence of pro-teolytic enzymes in the culture broth (16). Theanalytical isoelectric focusing in this presentstudy indicates that Thermoactinomyces pro-duces and releases all the extracellular proteinsinto the culture fluid simultaneously. Thus, itseems that at least three extracellular cellulo-lytic enzymes are produced by Thermoactino-myces, throughout the exponential growthphase, with no evidence of a sequential appear-ance of the various enzymes. There is also thepossibility that these enzymes originate fromone or more common precursors, which undergodifferent degrees of proteolytic degradation be-fore being released into the culture fluid.
ACKNOWLEDGMENTS
This work was supported by a grant from the Departmentof Energy, contract EY-76-S-02-4070.We thank Harriet Harris and Donna Berner for skillful
technical assistance.
LITERATURE CITED
1. Almin, K.-E., and K.-E. Eriksson. 1967. Enzymaticdegradation of polymers. I. Viscometric method for thedetermination of enzymic activity. Biochim. Biophys.Acta 139:238-247.
2. Berg, B., and G. Pettersson. 1977. Location and for-mation of cellulases in Trichoderma viride. J. Appl.Bacteriol. 42:65-75.
3. Berghem, L. E. R., and L. G. Pettersson. 1973. Themechanism of enzymatic cellulose degradation. Purifi-cation of a cellulolytic enzyme from Trichoderma virideactive on highly ordered cellulose. Eur. J. Biochem.37:21-30.
4. Berghem, L. E. R., L. G. Pettersson, and U.-B. Axio-Fredriksson. 1976. The mechanism of enzymatic cel-lulose degradation. Purification and some properties oftwo different 1,4-,f-glucan glucanohydrolases fromTrichoderma viride. Eur. J. Biochem. 61:621-630.
5. Emert, G. H., E. K. Gum, Jr., J. A. Lang, T. H. Liu,and R. D. Brown, Jr. 1974. Cellulases. Adv. Chem.Ser. 136:79-100.
6. Eriksson, K.-E., and B. Pettersson. 1975. Extracellularenzyme system utilized by the fungus Sporotrichumpulverulentum (Chrysosporium lignorum) for thebreakdown of cellulose. 1. Separation, purification andphysico-chemical characterization of five endo-1,4-,B-glucanases. Eur. J. Biochem. 51:193-206.
7. Fagerstam, L., U. Hakansson, G. Pettersson, and L.Andersson. 1977. Purification of three different cellu-lolytic enzymes from Trichoderma viride QM 9414 on
a large scale, p. 165-178. In T. K. Goose (ed.), Biocon-version of cellulosic substances into energy, chemicalsand microbial protein. Swiss Federal Institute of Tech-nology, Zurich.
8. Gong, C.-S., M. R. Ladisch, and G. T. Tsao. 1977.Cellobiase from Trichoderma viride: purification, prop-erties, kinetics and mechanism. Biotechnol. Bioeng.19:959-981.
9. Gum, E. K., Jr., and R. D. Brown, Jr. 1976. Structuralcharacterization of a glycoprotein cellulase, 1,4-,8-D-glu-can cellobiohydrolase C from Trichoderma viride.Biochim. Biophys. Acta 446:371-386.
10. Gum, E. K., Jr., and R. D. Brown, Jr. 1977. Comparisonof four purified extracellular 1,4-f8-D-glucan cellobio-hydrolase enzymes from Trichoderma viride. Biochim.Biophys. Acta 492:225-231.
11. Humphrey, A. E., A. Moreira, W. Armiger, and D.Zabriskie. 1977. Production of single cell protein fromcellulose wastes. Biotechnol. Bioeng. Symp. 7:45-64.
12. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J.Randall. 1951. Protein measurement with the Folinphenol reagent. J. Biol. Chem. 193:265-275.
13. Mandels, M., R. Andreotti, and C. Roche. 1976. Mea-surement of saccharifying cellulase. Biotechnol. Bioeng.Symp. 6:21-33.
14. Mandels, M., E. R. Andreotti, and C. Roche. 1977.Effect of fermentation variables on growth and enzymeproduction by Trichoderma, p. 249-268. In T. K. Goose(ed.), Bioconversion of cellulosic substances into energy,chemicals and microbial protein. Swiss Federal Instituteof Technology, Zurich.
15. Miller, G. L., R. Blum, W. E. Glennon, and A. L.Burton. 1960. Measurement of carboxymethylcellulaseactivity. Anal. Biochem. 2:127-132.
16. Nakayama, M., Y. Tomita, H. Suzuki, and K. Nisi-zawa. 1976. Partial proteolysis of some cellulase com-ponents from Trichoderma viride and the substratespecificity of the modified products. J. Biochem.79:955-966.
17. Nelson, N. 1944. A photometric adaptation of the Somo-gyi method for the determination of glucose. J. Biol.Chem. 153:375-380.
18. Somogyi, M. 1952. Notes on sugar determination. J. Biol.Chem. 195:19-23.
19. Su, T.-M., and I. Paulavicius. 1975. Enzymatic sacchar-ification of cellulose by thermophilic actinomyces. Appl.Polym. Symp. 28:221-236.
20. Winter, A., K. Ek, and U.-B. Andersson. 1977. Analyt-ical electrofocusing in thin layers of polyacrylamidegels. Application note 250. LKB-Produkter AB,Bromma, Sweden.
21. Winter, A., H. Perlmutter, and H. Davies. 1975. Pre-parative flat-bed electrofocusing in a granulated gelwith the LKB 2117 Multiphor. Application note 198.LKB-Produkter AB, Bromma, Sweden.
22. Wood, T. M., and S. I. McCrae. 1972. The purificationand properties of the C, component of Trichodermakoningii cellulose. Biochem. J. 128:1183-1192.












![Selective Isolation and Characterization of Cellulolytic ... · JIRCAS]ournalNo.5: 79-89 (1997) Selective Isolation and Characterization of Cellulolytic Bacteria by Cellulose Enrichment](https://static.documents.pub/doc/80x56/5c0a681409d3f2501a8b8ffd/selective-isolation-and-characterization-of-cellulolytic-jircasournalno5.jpg)








