ACTA UNIVERSITATIS UPSALIENSIS UPPSALA 2013 Digital Comprehensive Summaries of Uppsala Dissertations from the Faculty of Medicine 902 Effects of Endocrine Disrupting Chemicals on Human Endometrial Endothelial Cells MALIN HELMESTAM ISSN 1651-6206 ISBN 978-91-554-8663-1 urn:nbn:se:uu:diva-196730 In Vitro
Transcript
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2013
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 902
Effects of Endocrine DisruptingChemicals on HumanEndometrial Endothelial Cells
Dissertation presented at Uppsala University to be publicly examined in Auditorium Minus,Akademigatan 3, Uppsala, Wednesday, June 5, 2013 at 09:15 for the degree of Doctor ofPhilosophy. The examination will be conducted in Swedish.
AbstractHelmestam, M. 2013. Effects of Endocrine Disrupting Chemicals on Human EndometrialEndothelial Cells In Vitro. Acta Universitatis Upsaliensis. Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty of Medicine 902. 70 pp. Uppsala.ISBN 978-91-554-8663-1.
Evidence from an abundant number of studies suggests that human female reproductivefunctions have become impaired over the past half century and that there might be a relationshipbetween endocrine disrupting chemicals (EDCs) and reduced fertility. It is, however, notknown by what mechanisms EDCs affect different reproductive functions such as endometrialreceptivity, embryo implantation and placentation.
The endometrium is continuously changing its morphological and functional properties,responding to cyclic changes of oestrogen and progesterone levels during the menstrualcycle. These changes include monthly preparation for embryo implantation through changedendometrial angiogenic activity and consequent changes in endometrial vasculature.
Use of primary human endometrial endothelial cells (HEECs) in this work was evaluated asa possible screening tool for effects caused by EDCs on human endometrial vasculature andsubsequently on various endometrial functions.
In this study HEEC and endometrial stromal cells were isolated. HEECs were grown inmonocultures, and together with stromal cells in co-cultures, and exposed to endocrine activesubstances. These were cadmium, which has oestrogenic properties, tamoxifen, with anti-oestrogenic effects, mifepristone, which is an anti-progestin, and bisphenol A, with oestrogenicproperties. The effects were evaluated by using proliferation and viability assays, migration andtube formation assays, quantitative PCR (qPCR), immunohistochemistry and western blot.
Cadmium affected the expression of angiogenesis-related genes, and caused different effectsin HEECs cultured alone vs. HEECs co-cultured with stromal cells. Tamoxifen alteredthe expression of angiogenesis-related genes and reduced HEEC migration, thus having ananti-angiogenic effect. Mifepristone caused reduced formation of tubular structures in tube-formation assays involving HEECs co-cultured with stromal cells. Bisphenol A promoted tubeformation in co-cultured HEECs which was related to changes in the expression of severalangiogenesis-related genes as well as up-regulated expression of VEGF-D protein.
In conclusion, we showed that EDCs have the ability to induce changes in endometrialangiogenic activity in vitro and may thus disturb normal endometrial functions related to fertilityand pregnancy. HEECs grown in vitro may provide valuable information on the effects of EDCson human endometrial functions. However, this model is not suitable as a large-scale screeningtool.
“In the present circumstances, no one can afford to assume that someone else will solve their problems. Every individual has a responsibility to help and guide ourglobal family in the right direction. Good wishes are notsufficient; we must become actively engaged.”
Dalai Lama
List of Papers
This thesis is based on the following papers, which are referred to in the text by their Roman numerals.
I Helmestam, M., Stavreus-Evers, A., Olovsson, M. (2010) Cadmium
alters mRNA levels of angiogenesis related genes in primary human endometrial endothelial cells grown in vitro. Reproductive Toxicolo-gy. 30(3):370–6
II Helmestam, M., Andersson,H., Stavreus-Evers, A., Brittebo, E., Olovsson, M. (2012) Tamoxifen modulates cell migration and ex-pression of angiogenesis-related genes in human endometrial endo-thelial cells. American Journal of Pathology. 180(6):2527-35
III Helmestam, M., Lindgren, K. E., Stavreus-Evers, A., Olovsson, M. Mifepristone exposure of human endometrial endothelial cells in vitro. Submitted
IV Helmestam, M., Stavreus-Evers, A., Davey, E., Olovsson, M. Effects of bisphenol A on human endometrial endothelial cells in vitro. Manuscript
Reprints were made with permission from the respective publishers.
Contents
Introduction ................................................................................................... 11 The female genital system ........................................................................ 11
The uterus ............................................................................................ 11 The ovaries .......................................................................................... 12 Ovarian steroid hormones and their receptors ..................................... 12 The endometrium and the menstrual cycle .......................................... 16 Endometrial receptivity ....................................................................... 17 Cellular composition of the human endometrium ............................... 17
Material and Methods ................................................................................... 28 Subjects ................................................................................................ 28 Establishment of cell cultures .............................................................. 28 Exposure to test substances ................................................................. 28
Methods .................................................................................................... 29 Viability and proliferation assays (Papers I–IV) ................................. 29 Migration assay (Paper II) ................................................................... 29 Tube-formation assay (Papers III and IV) ........................................... 30 Cell culture for RNA and protein isolation (Papers I–IV) ................... 30 Quantitative PCR (Papers I–IV) .......................................................... 30 Immunohistochemistry (Paper II) ........................................................ 31 VEGF ELISA (Paper IV) ..................................................................... 32 Western Blots (Paper IV) ..................................................................... 32 Statistical analyses ............................................................................... 32
Results and Discussion ................................................................................. 33 Viability and proliferation (Papers I–IV) ................................................. 33 Migration and tube formation (Papers II, III and IV) ............................... 36
Quantitative PCR (Papers I–IV) ............................................................... 40 Expression of metabolizing enzymes in the human endometrium ........... 46 ELISA ...................................................................................................... 48 Western Blot ............................................................................................. 48
Summary of findings ..................................................................................... 51
General Discussion ....................................................................................... 52 Conclusions .............................................................................................. 54
Summary in Swedish .................................................................................... 56 Sammanfattning på svenska ..................................................................... 56
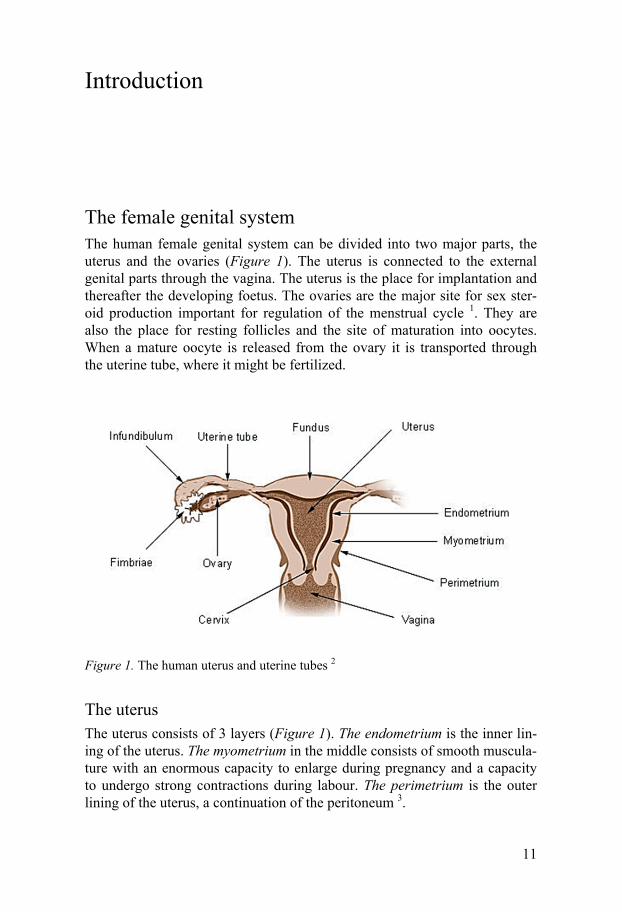
The female genital system The human female genital system can be divided into two major parts, the uterus and the ovaries (Figure 1). The uterus is connected to the external genital parts through the vagina. The uterus is the place for implantation and thereafter the developing foetus. The ovaries are the major site for sex ster-oid production important for regulation of the menstrual cycle 1. They are also the place for resting follicles and the site of maturation into oocytes. When a mature oocyte is released from the ovary it is transported through the uterine tube, where it might be fertilized.
Figure 1. The human uterus and uterine tubes 2
The uterus The uterus consists of 3 layers (Figure 1). The endometrium is the inner lin-ing of the uterus. The myometrium in the middle consists of smooth muscula-ture with an enormous capacity to enlarge during pregnancy and a capacity to undergo strong contractions during labour. The perimetrium is the outer lining of the uterus, a continuation of the peritoneum 3.
12
The ovaries The ovaries have two major functions. One function is the production of sex steroids and the other one is the storage, maturation and release of oocytes. The ovaries produce all three classes of sex steriods, progestogens, androgens and oestrogens, all through the conversion of cholesterol (Figure 2) 1. Cholesterol is a essential building block in steriodogenesis. It can be synthesised from acetate, but the major resource is the bloodstream.
Figure 2. Steroidogenesis
Ovarian steroid hormones and their receptors
Progestogens The progestogens are a group of hormones derived directly from cholesterol, and named after their function in maintaining pregnancy, (pro-gestational). Progesterone (Figure 3) is considered to be the most important progestogen in the reproductive tract. It is produced by the corpus luteum and is the pre-cursor of testosterone, which in turn may be converted to oestrogens. Proges-terone stimulates proliferation and differentiation of stromal and epithelial cells and plays an essential role in initiation and maintenance of pregnancy. During the secretory phase of the menstrual cycle progesterone drives the growth of spiral arterioles 4.
Two subtypes of human progesterone receptors (PRs) have been de-scribed, PRA and PRB. Both are derived from the same gene, but different promoters make PRA 164 amino acids shorter in the N-terminus. Both PRA
Cholesterol
Progestogens
Corticoids Androgens
Estrogens
Testosterone
13
and PRB can bind DNA as either homo- or heterodimers with one hormone molecule attached to each unit of the dimer. PRA and PRB are expressed in varying amounts in different cell types. The two receptor forms can be regu-lated independently; however, they may also affect the activity of the other receptor. In general, PRB is the positive regulator of progesterone-responsive genes and PRA inhibits the activity of PRB. The relative expres-sion levels of the two forms differ in the endometrium during the menstrual cycle. Progesterone may also bind to the glucocorticoid receptor (GR) 1 and the androgen receptor (AR) 1,5.
Most of the circulating progesterone is bound to albumin, some is bound to the corticosteroid-binding globulin transcortin and only a very small frac-tion is bound to sex hormone-binding globulin (SHBG) or is unbound 6.
Figure 3. Chemical structure of progesterone
14
Androgens Androgens in females are important precursors of oestrogens 1. Andros-tenedione and testosterone (Figure 4) are synthesized in the maturing follicle and thereafter aromatized in the granulosa cells into oestrogens. Androgens in the ovarian follicle control aromatase activity and hence oestrogen pro-duction. At low androgen concentrations aromatase activity is enhanced and at high concentrations it is inhibited. Androgens in the developing follicle are important in the regulation of ovarian function and folliculogenesis. In the endometrium there are significant amounts of androgens. However, their function has not yet been fully clarified 1.
Figure 4. Chemical structure of testosterone
Oestrogens Oestrogens are produced through conversion of androgens into testosterone, by the aromatase enzyme. Testosterone is further metabolized to oestradiol, the major oestrogen secreted by the ovary. Of the more than twenty oestro-gens isolated, oestradiol (Figure 5) is the most active. In the blood stream most of the oestradiol is bound to transport proteins, mainly SHBG and al-bumin. Once oestradiol is bound to an oestrogen receptor it is converted into oestriol, which is a less active oestrogen, and is excreted through the kid-neys.
Two major classical oestrogen receptors (ERs) have been identified 1, ER-α and ER-β. Oestrogen receptor-α was discovered in the 1960s and its gene is located at chromosome 6. Oestrogen receptor-β was not discovered until the 1990s and its gene is located at chromosome 14. The receptors bind to DNA either as homo-or heterodimers of ER-α and -β. The two oestrogen receptors share a common structure. However, differences in their NH2-terminal regions make them respond differently to various ligands, especial-ly to selective oestrogen receptor modulators 7,8. Both receptors bind the hormone with similar affinity, although phytoestrogens and synthetic oestro-gens may have different affinities for the two receptors. Oestrogen signalling may also occur via the transmembrane G-protein-coupled receptor (GPR30) 9-12.
15
In the endometrium ER-β is less prominent than ER-α 13,14 and it exhibits less change during the menstrual cycle, except when it becomes the predom-inant oestrogen receptor in the endometrial vasculature in the late secretory phase 1,13,15.
Figure 5. Chemical structure of oestradiol
General receptor mechanism The classical model of sex steroid hormone receptor actions begins with diffusion of the ligand, i.e. the sex steroid hormone, across the cell mem-brane, enabling binding to the receptor 16. Upon binding of the ligand, the receptors are activated and dissociated from protecting heat shock proteins, and a chromatin binding site is revealed. The hormone-receptor complex then interacts with specific nuclear DNA sites, called hormone responsive elements. Transcription of target genes to messenger ribonucleic acid (mRNA) occurs through communication with co-regulators and the general transcription apparatus 16,17. The mRNAs are transported to the cytoplasmic ribosomes, where translation into proteins takes place.
G protein-coupled receptors In general, signalling from G protein-coupled receptors often results in changes in the activity of already existing proteins, even though it has been observed that activation of these receptors may induce changes in gene ex-pression 16. It is also possible that these receptors can initiate signalling via more than one intracellular signalling pathway, leading to various cellular responses. Stimulation of a coupled receptor causes activation of the G pro-tein, which in turn modulates the activity of an associated effector protein. GPR30 is a G protein-coupled receptor activated by oestrogen 18. Its expres-sion in the endometrium follows the cyclic change of sex steroid hormones – it is up-regulated by oestrogen via ER-α during the proliferative phase and suppressed by progesterone via PRA during the secretory phase 19.
16
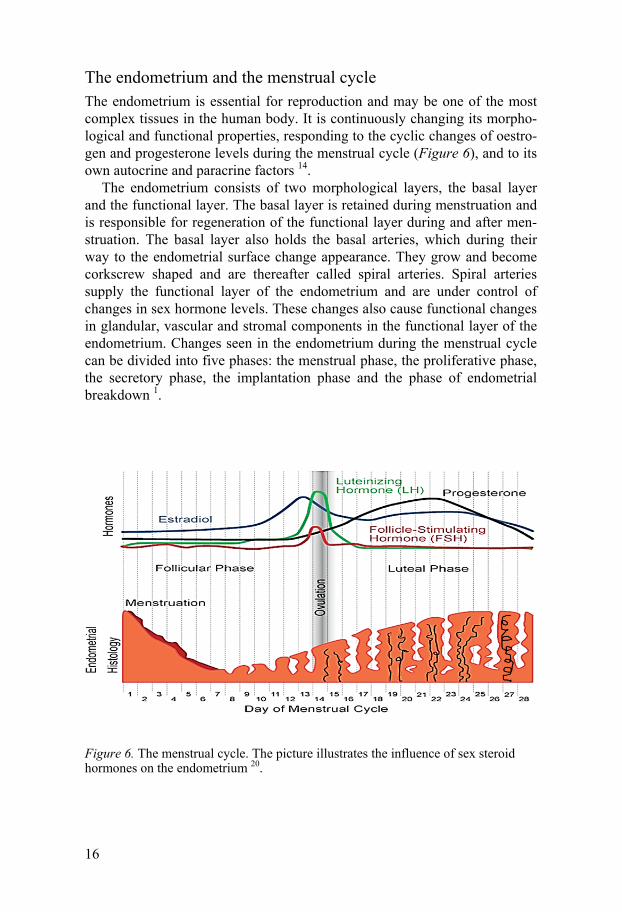
The endometrium and the menstrual cycle The endometrium is essential for reproduction and may be one of the most complex tissues in the human body. It is continuously changing its morpho-logical and functional properties, responding to the cyclic changes of oestro-gen and progesterone levels during the menstrual cycle (Figure 6), and to its own autocrine and paracrine factors 14.
The endometrium consists of two morphological layers, the basal layer and the functional layer. The basal layer is retained during menstruation and is responsible for regeneration of the functional layer during and after men-struation. The basal layer also holds the basal arteries, which during their way to the endometrial surface change appearance. They grow and become corkscrew shaped and are thereafter called spiral arteries. Spiral arteries supply the functional layer of the endometrium and are under control of changes in sex hormone levels. These changes also cause functional changes in glandular, vascular and stromal components in the functional layer of the endometrium. Changes seen in the endometrium during the menstrual cycle can be divided into five phases: the menstrual phase, the proliferative phase, the secretory phase, the implantation phase and the phase of endometrial breakdown 1.
Figure 6. The menstrual cycle. The picture illustrates the influence of sex steroid hormones on the endometrium 20.
17
The menstrual endometrium is dense and thin; it consists of the basal layer and minor residues of the functional layer. At menstruation the functional layer is shed and DNA synthesis begins in the basal layer where the func-tional layer is completely absent. New surface epithelium grows out from the remnants of glands in the basal layer.
The proliferative phase is stimulated by oestrogens secreted by the grow-ing ovarian follicles. The increasing amount of oestrogens induces cell pro-liferation and tissue regeneration in the endometrial glands, spiral arteries and stroma. A few days before ovulation the oestrogen level reaches a peak (Figure 6); this is associated with the highest expression of oestrogen recep-tors and extensive proliferation 1.
The secretory phase starts after ovulation when the corpus luteum is formed and progesterone secretion is initiated. Progesterone terminates epi-thelial proliferation, the mitotic processes and DNA synthesis in the endome-trium. The endometrial glands enlarge and become filled with secretory products, required if implantation occurs. If implantation does not occur, the corpus luteum degenerates and oestrogen and progesterone production is largely inhibited. The phase of endometrial breakdown is initiated, a process involving endometrial vasomotor reactions, apoptosis, tissue loss and finally shedding of the functional layer 14.
Endometrial receptivity The progression of fertilization into normal pregnancy is not possible with-out successful blastocyst implantation. The human endometrium is not re-ceptive to blastocyst implantation at all times. Under the influence of proges-terone the endometrium changes from proliferative to secretory. At the time of blastocyst implantation, the endometrium has reached its peak in secretory activity. This period, when the endometrium is receptive to the blastocyst, is called the implantation window and it lasts only a few days 6. The secretion of nourishing glycogen and lipids is important, since the growing embryo is dependent on the endometrium for nourishment during the first trimester. Thereafter the placenta becomes the major site of exchange of nutrients and waste products between the mother and the foetus. Implantation is coordi-nated between the embryo and the endometrium. This process is influenced by oestrogen and progesterone, growth factors, cytokines and vasoactive factors 21.
Cellular composition of the human endometrium The endometrium can be separated into two hormone-responsive parts, the epithelial part (luminal and glandular epithelium) and the stromal part (endo-thelial cells, leukocytes and fibroblasts). Luminal epithelial cells provide a defensive barrier and also act as the crucial site of embryo attachment. Glan-
18
dular epithelial cells secrete numerous autocrine/paracrine factors that are required for endometrial development and/or embryo implantation.
Endothelial cells are present in the walls of arteries and veins in the stro-ma, playing an active role in endometrial angiogenesis. Leukocytes are in-volved in inflammatory reactions and form part of the immune system. Fi-broblast cells, secreting matrix metalloproteinases, are involved in remodel-ling of the endometrial extracellular matrix. The extracellular matrix is the extracellular part of the endometrium, providing structural support.
Angiogenesis Endothelial cells that line the vasculature are normally quiescent but can be induced to proliferate, migrate and form new blood vessels when stimulated by angiogenic factors. This process is referred to as angiogenesis 22.
New blood vessels may arise either through vasculogenesis, which occurs during embryonic development when angioblasts differentiate and subse-quently form new blood vessels, or through angiogenesis, when new blood vessels are formed out of pre-existing vessels. Angiogenesis may occur by different mechanisms: sprouting, intussusception or elongation/widening.
The first mechanism to be described was sprouting, which involves sever-al steps that include activation of endothelial cells, breakdown of the base-ment membrane, migration and proliferation of endothelial cells, tube for-mation and stabilization of tubes. Intussusception, on the other hand, in-volves partition of the vessel by insertion of extracellular matrix and endo-thelial cells. This vessel-dividing action results in formation of smaller vessels, which may remodel or enlarge. Elongation occurs through circulat-ing progenitor endothelial cells and proliferating existing endothelial cells in the vessel that elongate the vessel lengthways.
The different mechanisms of angiogenesis have consequences as regards functioning of the vessel and the vascular bed. In sprouting, blood cannot begin to flow until a new lumen is formed; in both intussusception and elon-gation blood flows continuously 4.
Growth Factors The angiogenic process is initiated by stimulation of growth factors. The angiogenic factors involved balance the progress of angiogenesis via delicate coordination of expression of positive and negative factors.
Growth factors with promoting effects on angiogenesis include vascular endothelial growth factors (VEGF-A, -B, -C, -D and -E) and their receptors (VEGFR-1, -2 and -3), placental growth factor (PlGF), epidermal growth factor (EGF) and its receptor and fibroblast growth factors (FGFs) and their receptors 22,23.
19
The VEGF family is essential for the control of angiogenesis. VEGFs are also endothelial mitogens and exhibit specific affinity for vascular endotheli-al cells. VEGFs and PlGF and their receptors are important in the develop-ment of the vasculature in a developing and growing embryo. The FGF fami-ly members are effective inducers of endothelial cell proliferation, migration and tube formation 22.
Endometrial angiogenesis Angiogenesis is an essential process in tissue growth and development. In adult life angiogenesis is more or less absent under normal conditions, ex-cept in the female genital tract 24. In the female reproductive tract, the cyclic changes in hormones and subsequent tissue effects lead to periods of rapid growth and regression. Angiogenesis occurs during all stages of the men-strual cycle, the menstrual, the proliferative and the secretory phases. Vessel elongation is a major mechanism during the oestrogenic-driven mid–late proliferative phase. During the progesterone-dominated secretory phase, sprouting and intussusception are probably more dominant, although it has been hard to prove what actually is happening 4.
The endometrium is a major source of angiogenic factors necessary dur-ing the menstrual cycle and pregnancy 25. VEGFs are abundantly expressed in the endometrium, reaching a peak that correlates with the highest angio-genic activity observed, during the secretory phase 26. During the prolifera-tive phase oestrogen stimulates VEGF production. Angiogenesis is also in-fluenced by many other growth factors and substances such as fibronectin and prostaglandins. The fibroblast growth factor family, in particular, is highly mitogenic as regards endothelial and stromal cells. Angiopoietins sustain the endometrium by preventing apoptosis. The endometrium also produces inhibitory proteins, and the final growth of blood vessels reflects the balance between inhibitory and stimulatory factors.
Hormone effects on endothelial cells The endothelium appears to be a direct target for oestrogens. They have been found to be protective of the endothelium in various pathophysiological situ-ations. It is well established that the frequency of coronary heart disease is lower in women than in men and increases after menopause when the secre-tion of sex steroids decreases 27.
Endothelial cells express oestrogen receptors, and depending on the origin of the endothelial cell the expression of ER-α and ER-β is different. Most investigational work has been done on ER-α and endothelial cells 27. Endo-thelial cell proliferation is promoted by ER signalling 28,29. Once activated, ERs undergo a conformational change and regulate transcription of oestro-gen-responsive genes by binding to the oestrogen-responsive elements 30.
20
Both ER-α and -β have important roles as transcription factors, activated by oestrogen binding.
Human endometrial endothelial cells express ER-β but not ER-α 31-33. ER-β is located either on the plasma membrane, in the cytosol, on the mitochon-dria or in the nucleus 27. It may activate different pathways and is involved in protection against cell injury, and it increases mitogen transcription upon activation 27.
Binding of estradiol to endothelial cell membrane receptors may also re-sult in rapid activation of endothelial nitric oxide synthase (eNOS) 34. This pathway has given rise to the concept that ERs on the plasma membrane may bind sex steroids and interact with other proteins and initiate signalling. The activation of eNOS by oestrogen is a well-established pathway. Other sex steroids have also been investigated and it is evident that rapid actions of sex steroids are important in the regulation of a number of long-term endothelial functions such as proliferation, senescence, apoptosis and migration 34.
Progesterone influences the expression of a variety of genes mediated by the progesterone receptor. The increase of circulating progesterone causes a shift from the proliferative phase to the secretory phase in the endometrium. During the proliferative phase and under the influence of elevated oestrogen concentrations the expression of PRs is increased. Subsequently, in the se-cretory phase, rising progesterone levels down-regulate ERs 35. It is not clear whether or not endometrial endothelial cells express PRs. Such expression has been proposed 32,33,35, but other investigators, however, have failed to show constitutive endothelial cell expression of PRs in humans 36,37 or in rhesus macaques 13. Although there are conflicting in vitro results, endome-trial angiogenesis has been observed in response to progesterone in vivo 25. Non-genomic effects of PRs have also been proposed 36.
21
Endocrine disrupting chemicals Reproductive disorders caused by chemicals have been known since Roman times, when lead was recognized as causing miscarriage and infertility in women and men. Throughout times it has been evident that many other chemicals may impair reproduction in different species 38.
The endocrine system is closely regulated by fluctuations in hormone lev-els. Hormones are signalling molecules secreted into the bloodstream, exert-ing their effects on different target organs in the body. Hormones are chemi-cally classified into water-soluble and lipid-soluble. Steroids and thyroid hormones are lipid-soluble and diffuse across cell plasma membranes and bind to intracellular receptors. Activation of these receptors leads to regula-tion of gene expression 16.
Endocrine disrupting chemicals (EDCs) are chemicals or chemical mix-tures that interfere with normal hormone action. They affect the hormone system by interfering with hormones and hormone receptors. There are gen-erally two ways that chemicals may disrupt hormone action: a direct action on a hormone-receptor protein complex or an indirect action on a specific protein that controls some aspect of hormone delivery to the right place at the right time.
Figure 7. Endocrine disruptors are chemicals or chemical mixtures that interfere with normal hormone actions. They may be of any type of chemical substance.
Endocrine-disrupting chemicals, like sex steroid hormones, are usually lipid-soluble, and can easily cross cell membranes. The consequences of endo-crine disruption are predominantly effects on development, growth processes and sexual differentiation 38,39. Exposure in utero to EDCs is considered es-pecially harmful 40,41. An example detected in the 1970s was the oestrogenic drug diethylstilbestrol (DES) given to women during the 1940s to prevent
22
miscarriages. Diethylstilbestrol was associated with benign reproductive tract problems in more than 95% of DES-exposed daughters. Reproductive organ malformation and dysfunction, poor pregnancy outcome, and immune system disorders were just some of the reported effects. Similarly, DES-exposed male offspring demonstrated structural, functional and cellular ab-normalities following prenatal exposure 42. Since then, many other synthetic and natural compounds (Figure 7) have been shown to interfere with the endocrine system via different mechanisms 43. Bisphenol A and other EDCs might contribute to polycystic ovary syndrome (PCOS), polychlorinated biphenyl (PCB) is associated with increasing menstrual cycle length and phthalates and bisphenol A are associated with endometriosis 44.
EDCs may interfere with the synthesis, secretion and/or signalling of hormones. Much of the information regarding adverse effects of EDCs in humans comes from high-dose exposure as a result of environmental catas-trophes. Most important from a human health perspective, however, is low-dose, long-term exposure, where fertility, neurobehaviour, immune function, metabolism and cancer are known to be affected/involved and there may be considerable effects in large populations 44,45.
For many years, when assessing the effects of possible endocrine disrup-tors, toxicologists have acted on the assumption that “the dose makes the poison”, implying that higher doses cause greater harm and assuming that effects not seen at high doses are not expected at low doses. In contrast to the above-mentioned dogma in toxicology, multiple studies have revealed that neither threshold nor linear no-threshold models are applicable to the responses seen in connection with endocrine disruptors 46. Dose-response curves associated with endocrine-disrupting chemicals often appear as U-shaped (non-monotonic) compared with the linear dose-response curves usually displayed in toxicology 46.
Humans are exposed to many different endocrine-disrupting chemicals simultaneously. Each of these chemicals may be determined as being “safe”, but mixtures of them may have both additive and synergistic effects. A mix-ture may result in more severe endocrine disruption than one chemical at a time 43.
Substances investigated in this thesis In this thesis experimental studies of the effects of four substances on human primary endometrial endothelial cells are presented. Two of them are drugs that currently are in clinical use worldwide. One is the breast cancer drug tamoxifen and the other is mifepristone, which is used for medical abortions. The other two are the environmental pollutants cadmium (Cd) and bisphenol A (BPA).
23
Cadmium The heavy metal cadmium is a chemical element, and a xenobiotic agent that is highly toxic and widespread. It has been classified as a human carcinogen 47 and is known to cause adverse reproductive effects 48. Cadmium is mainly associated with occupational exposure, but the general population is also chronically exposed owing to the widespread nature of its occurrence. Cad-mium can be found at low concentrations in almost every kind of food and in drinking water 49. Another important route of exposure is through cigarette smoke 50. An industrial way of living, burning of fossil fuels, the use of phosphate fertilizers and the use of cadmium as an anticorrosion agent con-tribute greatly to exposure. Furthermore, cadmium used in polyvinyl chlo-ride (PVC) products, colour pigments, alloys and batteries add to the cadmi-um load in the environment. In view of its long biological half-life, approx-imately 25 years, it is plausible that cadmium accumulates in biological sys-tems 51.
It is known that cadmium affects human and animal reproductive func-tions 48,52, probably through interference with endocrine mechanisms. Data suggest that cadmium may interfere directly with hormone production and it is therefore classified as an EDC 52,53. It has been shown that exposure to cadmium has an oestrogenic effect both in vivo and in vitro 54,55 and that it affects progesterone synthesis in human placental trophoblast cells 52. Expo-sure of the human breast cancer cell line MCF-7 to cadmium has been found to decrease ER expression and increase PR expression 54. In mammals cad-mium exposure is associated with ovulation failure 56,57, defective steroido-genesis 58, suppressed oocyte maturation 59 and implantation failure 60. Ex-posure of pregnant women to cadmium is related to an increased incidence of premature delivery 61. It has also been demonstrated that cadmium expo-sure is associated with endometrial cancer 62.
Tamoxifen The selective oestrogen receptor modulator (SERM) tamoxifen (Figure 8) is the most widely used agent for prevention and treatment of oestrogen recep-tor-positive breast cancer 63,64. Tamoxifen treatment inhibits breast cancer growth and prevents breast cancer relapse, but it is also associated with be-nign lesions and malignant tumours in the human endometrium 65. The ad-verse effects of tamoxifen on the endometrium include endometrial hyper-plasia and cancer, conditions that are strongly associated with angiogenesis 66.
Depending on tissue and cell type, tamoxifen can function either as an ER antagonist or agonist 7,67. The antagonistic mode of action of tamoxifen is seen in breast tissue, where it inhibits breast cancer cell proliferation. In con-
24
trast, tamoxifen has been reported to function as an ER agonist in the endo-metrium, where it promotes epithelial cell proliferation 68.
Little is known about the effects of tamoxifen on human endometrial vas-culature. Some reports suggest that tamoxifen modulates the expression of angiogenic factors in human endometrial vasculature both in vivo and in vitro, and also that the antitumor effect of tamoxifen in breast tissue may be related to anti-angiogenic tamoxifen action 69-71.
Figure 8. Chemical structure of tamoxifen
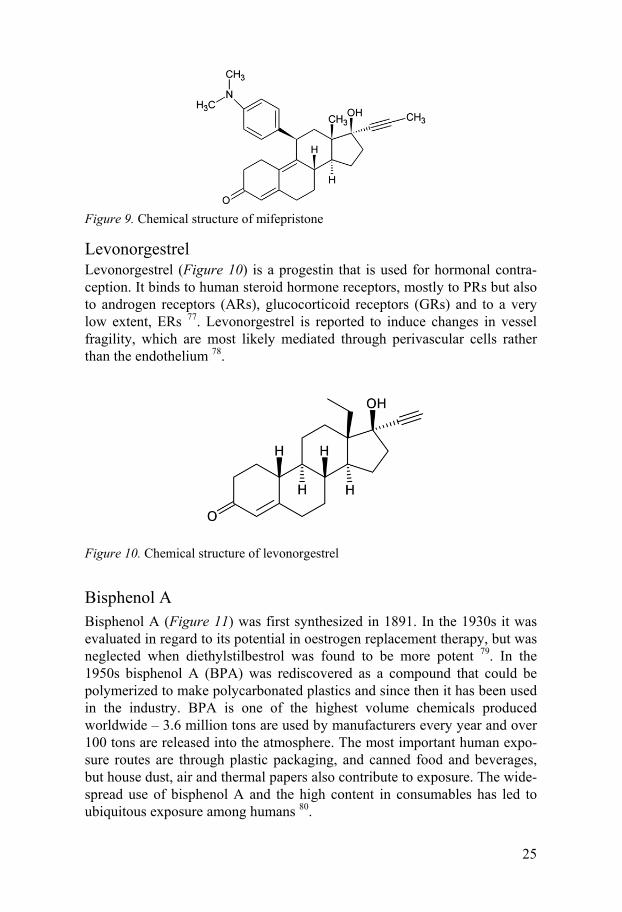
Mifepristone Mifepristone (Figure 9) is a progesterone receptor antagonist or anti-progestin, which act as a fertility regulator by blocking the action of proges-terone 72. In the presence of progesterone, mifepristone is an antagonist, but alone it exhibits endometrial progestational effects at low doses and an anti-proliferative (anti-oestrogenic) effect at higher doses 73. It was the first avail-able anti-progestin and has been used successfully for more than 20 years as a medical alternative for early pregnancy interruption 72. At low doses it is also suitable for emergency contraception, by inhibiting ovulation and alter-ing endometrial functions 74.
Significant changes in endometrial vessels in vivo have been observed af-ter mifepristone administration, seen as cell injury and cell death in capillary endothelial cells 75. This vascular damage has been proposed to compromise embryo survival 72. Mifepristone has also been reported to affect endometrial receptivity markers in a co-culture system of human endometrial epithelial and stromal cells 76.
25
Figure 9. Chemical structure of mifepristone
Levonorgestrel Levonorgestrel (Figure 10) is a progestin that is used for hormonal contra-ception. It binds to human steroid hormone receptors, mostly to PRs but also to androgen receptors (ARs), glucocorticoid receptors (GRs) and to a very low extent, ERs 77. Levonorgestrel is reported to induce changes in vessel fragility, which are most likely mediated through perivascular cells rather than the endothelium 78.
Figure 10. Chemical structure of levonorgestrel
Bisphenol A Bisphenol A (Figure 11) was first synthesized in 1891. In the 1930s it was evaluated in regard to its potential in oestrogen replacement therapy, but was neglected when diethylstilbestrol was found to be more potent 79. In the 1950s bisphenol A (BPA) was rediscovered as a compound that could be polymerized to make polycarbonated plastics and since then it has been used in the industry. BPA is one of the highest volume chemicals produced worldwide – 3.6 million tons are used by manufacturers every year and over 100 tons are released into the atmosphere. The most important human expo-sure routes are through plastic packaging, and canned food and beverages, but house dust, air and thermal papers also contribute to exposure. The wide-spread use of bisphenol A and the high content in consumables has led to ubiquitous exposure among humans 80.
26
Bisphenol A is classified as an EDC with oestrogenic properties. It is classified as a selective oestrogen receptor modulator 81 that has the ability to bind both ER-α and ER-β 80. The proportion of BPA that is bound to serum albumin is very low and therefore BPA has an advantage over oestradiol, which is to a greater extent bound to serum albumin. The binding capacity of BPA to ERs is approximately 1000-fold less compared with oestradiol, which is why BPA is sometimes referred to as a weak oestrogenic com-pound. Others, however, propose BPA to be a strong xenoestrogen 79. When oestrogen is bound to ERs, transcriptional activity depends on the recruit-ment of co-regulators to the oestrogen/ER complex. Even though BPA has a 10-fold greater affinity to ER-β, the BPA/ER-α complex has a 500-fold greater potency in recruitment of co-regulators 81, which may contribute to the diversity of effects caused by BPA. BPA also binds to GPR30, which has been shown to trigger non-genomic steroid actions.
Figure 11. Chemical structure of bisphenol A
27
Aims
Evidence from an abundant number of studies suggests that human female reproductive functions have become impaired over the past half century and that there is a relationship between endocrine disrupting chemicals and re-duced fertility. However, the mechanisms via which EDCs affect different reproductive functions are not known.
The general aim of this thesis was:
• to evaluate whether or not primary human endometrial endothe-lial cells (HEECs) grown in vitro constitute a possible screening tool for determining the effects of EDCs on the human endome-trial vasculature
The specific aims were:
• to study the effects of hormone drugs and EDCs on HEEC viability and proliferation (Papers I–IV)
• to see if the expression of angiogenesis-related genes is af-fected by the investigated chemicals (Papers I–IV)
• to determine if HEEC migration is affected by tamoxifen (Paper II)
• to study CYP expression in the endometrium (Paper II)
• to determine if HEEC tube formation is affected by mifepris-tone or bisphenol A (Papers III and IV)
• to investigate the effect of bisphenol A on the expression of certain proteins in HEECs (Paper IV)
28
Material and Methods
The studies were approved by the Regional Ethics Committee, Uppsala, Sweden. Informed consent was obtained from all women included in the study.
Subjects Endometrial tissue samples were obtained from four to five premenopausal women for each study. The women had regular menstrual cycles and under-went hysterectomy for benign medical conditions at Uppsala University Hospital. None of the participants had received any hormonal treatment dur-ing a period of at least three months prior to surgery and they were all non-smokers.
Establishment of cell cultures All endometrial biopsy samples were immediately placed in saline buffer and transferred from the surgical ward to the laboratory. The tissues were first mechanically minced and then enzymatically digested. Magnetic beads coated with antibodies against the endothelial cell-specific surface antigen CD31 were added to the cell suspension. The endothelial cells were then isolated with the aid of a magnetic holder. The stromal cells in the superna-tant were poured into a separate tube. Both stromal and endothelial cells were suspended and seeded in Endothelial Cell Medium (ECM).
Exposure to test substances Cadmium chloride, tamoxifen, 17β-oestradiol, mifepristone, progesterone, levonorgestrel and BPA stock solutions were diluted in cell culture medium. HEECs were exposed to the test substances for 48 h except for BPA, where the cells were exposed for 24 h.
29
Methods To examine changes in HEECs induced by the investigated substances in this thesis, several well-established methods were used. An overview of the methods is listed in Table 1.
Table 1. Investigated biological endpoints and methods used
Endpoint Method Paper(s)
Cell Viability WST I–IV
Cell Proliferation BrdU I–IV
Cell migration Migration assay II
In vitro angiogenesis Tube-formation assay III, IV
mRNA expression qPCR I–IV
CYP expression Immunohistochemistry, qPCR II
Protein expression
Protein expression
ELISA
Western blot
IV
IV
Viability and proliferation assays (Papers I–IV) When the cells were at sub-confluence in passage one to two, HEECs were transferred to sterile 96-well plates for viability (WST-1) and proliferation (BrdU) assays. For co-cultures, stromal cells were seeded in inserts in the same wells, and the cells were exposed simultaneously to the different test substances. Viability was assessed by using WST-1 kits, where WST-1 solution was added to the test substance-treated HEECs for the last hour of the incubation and absorbance was measured at 450 nm. The viability assay measures met-abolic activity by the ability of mitochondrial enzymes to cleave tetrazolium salt. In the proliferation assay the treated HEECs were exposed to BrdU labelling solution for the last 24 h of the incubation. Labelling was visual-ized using a peroxidase reagent and absorbance was measured at 450 nm. The cell proliferation assay measures the amount of incorporated BrdU (structural analogue of thymidine) into the DNA during the synthesis phase of the cell cycle, thereby serving as a marker of proliferation.
Migration assay (Paper II) In Study II, cell migration was evaluated by using a modified Boyden cham-ber. HEECs were incubated for 48 hours in monoculture or in co-culture with stromal cells in culture media containing 0.01 µM, 0.1 µM, 1 µM or 10 µM tamoxifen or ethanol vehicle. Thereafter, HEECs were transferred to the
30
modified Boyden chamber and grown in the same culture media as before, i.e. containing tamoxifen or vehicle. A filter was coated with collagen and inserted between the chambers. The cells were then placed in the upper chamber, and the lower chamber was filled with endothelial cell media (ECM) (control), ECM supplemented with 10% FCS (positive control) or ECM with VEGF (attractant). Migration was quantified by counting the number of cells that had migrated to the lower chamber.
Tube-formation assay (Papers III and IV) The endothelial tube-formation assay was performed as described previously 82,83. HEECs were cultured either as monocultures or co-cultures and treated with the different substances. The wells of a 96-well cell culture plate were loaded with 70 μL Geltrex reduced growth factor extracellular matrix mem-brane and incubated for 30 min at 37 °C until it gelled. HEECs in monocul-tures or in co-cultures incubated for 24 h (bisphenol A) or 48 h (mifepris-tone) in medium containing test substance or vehicle were detached by tryp-sinization and plated on top of the Geltrex basement membrane and incubat-ed for 6 h at 37 °C. VEGF (mifepristone) or bFGF (bisphenol A) was added as stimulating agent. Tube formation was observed under a microscope. The images were analysed by using the Wimtube formation module assessing the following parameters: Covered Area [%], Total Tube Length, Total Branch-ing Points, Total Loops (loops are defined as the closed areas inside the tub-ular structure), Total Nets (are the different tubular structures on the image that are not connected to each other) and Total Tubes.
Cell culture for RNA and protein isolation (Papers I–IV) At sub-confluence, HEECs seeded in sterile 6-well cell culture plates were exposed to the test substances. Stromal cells were seeded in inserts in the same wells as the HEECs. After exposure, total RNA was isolated according to the RNeasy Mini spin protocol and frozen until cDNA was synthesized. In Study IV RNA and protein isolation was carried out according to the All-Prep® RNA/Protein Kit protocol.
Quantitative PCR (Papers I–IV) Quantitative PCR (qPCR) reactions were carried out in a Step One Plus Re-al-Time PCR System. The efficiency of each run was determined by using LinRegPCR software. Each sample was run in triplicate and the target genes were normalized according to the mean normalized expression equation (see Paper I). The investigated gene expressions are listed in Table 2.
31
Table 2. Genes investigated by qPCR
Gene Assay Paper
VEGF-A SYBR I–III
PlGF SYBR I–III
VEGFR-1 SYBR I–III
VEGFR-2 SYBR I–III
VWF SYBR I–III
HIF-1α SYBR I–III
VE-cadherin SYBR I–III
ER-α SYBR I–III
ER-β SYBR I–III
CYP1A1 SYBR II
CYP1B1 SYBR II
PR A+B SYBR III
GR SYBR III
AR SYBR III
VEGFR-3 (FLT4) TaqMan IV
ANGPLT4 TaqMan IV
PDGFRB TaqMan IV
VEGF-D (FIGF) TaqMan IV
CXCL10 TaqMan IV
TGFα TaqMan IV
Immunohistochemistry (Paper II) Vascular expression of the major tamoxifen metabolizing enzymes CYP1A1, CYP1B1, CYP2A6, CYP2B6, CYP2C8/9/19, CYP2D6, CYP3A4/5 and SULT2A1 was investigated immunohistochemically using human endome-trial biopsy samples. Fixed endometrial samples were embedded in low-temperature paraffin and sectioned (4 μm). Endogenous peroxidase activity was quenched with hydrogen peroxide. The sections were washed and im-mersed in sodium citrate buffer, for antigen retrieval by microwave treat-ment, then incubated overnight with the primary antibody. Thereafter, they were rinsed and incubated with a secondary antibody. Avidin-biotin peroxi-dase complex solution was applied, and the samples stained for 2 to 6 minutes. Immunohistochemical staining was performed on at least two sepa-rate occasions for each primary antibody.
32
VEGF ELISA (Paper IV) Cell culture media were collected after 24 h exposure of the cells to BPA prior to lysing the cells for RNA/protein isolation. The media were collected in glass bottles and frozen at -20 °C until analysed.
Concentrations of VEGF were measured in duplicate using an ELISA ac-cording to the manufacturers' instructions. The assay was a quantitative sandwich ELISA technique. For controls, VEGF concentrations were ana-lysed in fresh media on two different occasions – once when the media were from the refrigerator (+4 °C) and once when the cell culture medium had been in the incubator (5% CO2 in humidified air at 37 ºC) for 24 h in 6-well cell culture plates. This was to confirm that VEGF does not adsorb to the plastic of the cell culture dish.
Western Blots (Paper IV) HEEC protein was pooled group-wise according to exposure to BPA at dif-ferent concentrations. All samples were then diluted and adjusted to the same protein concentration. Protein samples were denatured in sample buffer and reducing agent, thereafter being loaded and separated on a gel.
Proteins were transferred to a PVDF membrane and then blocked in blocking buffer for 1 hour before being incubated at 4 °C overnight with the primary antibodies (β-actin, ER-β1, GPR30, VEGF-D and PR A+B). After incubation with primary antibodies, the membranes were washed and incu-bated for 1 hour with the secondary antibodies. The proteins were then visu-alized by using an Odyssey Infrared Imaging device.
Statistical analyses Mean normalized expression (MNE), based on the ratio between mean Ct values of target and reference genes and the efficiency of the PCR reactions, was calculated as a measurement of target gene transcription 84-86. The data are presented either as MNE or as log2-fold change to illustrate the differ-ence between two data sets.
The Mann–Whitney U test was applied to compare data from the prolifer-ation, viability, tube-formation, migration and qPCR assays. Two-way ANOVA and the Kruskal–Wallis test were used to assess the impact of stromal cells on HEEC proliferation, viability and mRNA expression. Addi-tional post hoc tests were also carried out (Tukey’s HSD test). Values of p less than 0.05 were considered to denote a statistically significant difference.
33
Results and Discussion
Viability and proliferation (Papers I–IV)
Methodological considerations The viability assay measures cellular metabolic activity via NADP-dependent cellular reductive enzymes. Reduction of WST-1 increases with cellular metabolic activity as a result of elevated NADP change. WST-1 reduction is associated with a colour shift which is measured to assess viabil-ity. The proliferation assay measures proliferating cells by incorporation of BrdU, which is a synthetic analogue of thymidine. It is incorporated during DNA replication in dividing cells by substituting for thymidine. BrdU-specific antibodies can then be used to detect the incorporation, thus show-ing cells active in replicating their DNA.
With both assays there are considerations to take into account: Regarding the viability assay, resting cells that are viable but metabolically inactive reduce very little WST-1. In contrast, rapidly dividing cells exhibit high rates of WST-1 reduction. It is important to remember that assay conditions can alter metabolic activity and thus dye reduction without affecting cell viabil-ity. Regarding the proliferation assay it is known that studying incorporation of BrdU in cells might give an overestimation of the total number of live cells, since the method does not ensure the viability of the cells. It is also possible that incorporation of BrdU itself affects proliferation and cell sur-vival. It is important to be aware of these potential methodological short-comings; the combination of these two assays was therefore used to assess the condition of the cells.
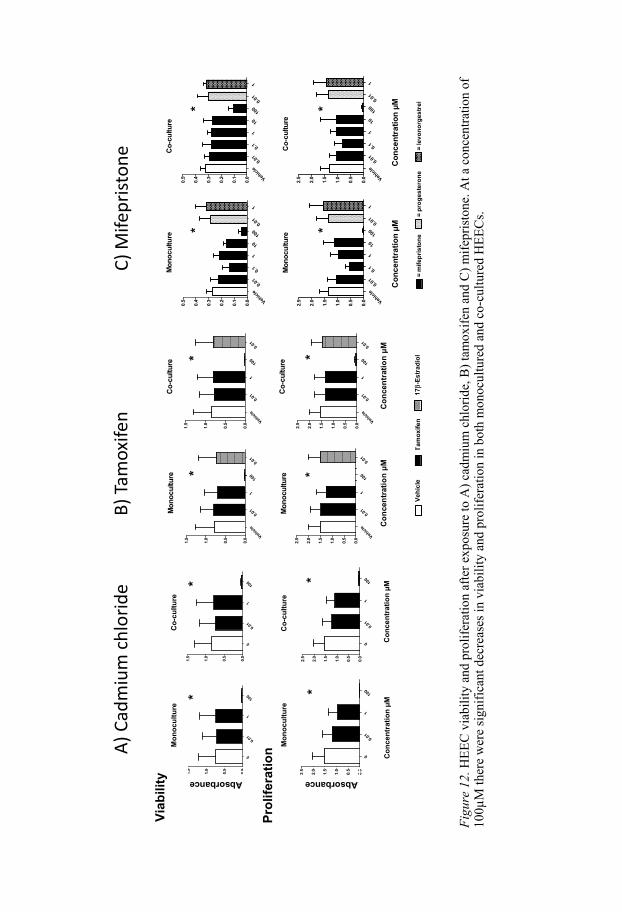
Results of viability and proliferation assays In Studies I–III of this thesis viability and proliferation assays were used to determine the concentrations of test substances at which viability and prolif-eration dropped, and these results were used to keep concentrations below cytotoxic levels. This was important, since the aim was to study endocrine and not cytotoxic effects of the test substances. The results showed that 100 µM cadmium chloride, tamoxifen and mifepristone all decreased both viabil-ity and proliferation considerably (Figure 12)
Pilot studies of the effects of cadmium chloride and tamoxifen on mono-cultured HEECs had previously revealed that there were no effects on cell
34
viability and proliferation at 0.1 and 10µM. Concerning cadmium chloride and tamoxifen we therefore evaluated only three different concentrations, namely 0.01, 1 and 100 µM. We thus do not know if there would be effects on viability and proliferation in co-cultured HEECs with 0.1 and 10 µM cadmium chloride and tamoxifen. Concerning mifepristone, viability and proliferation were evaluated at five concentrations ranging from 0.01–100 µM and there were significant effects only at 100 µM (Figure 12).
F
igur
e 12
. HE
EC
via
bilit
y an
d pr
olif
erat
ion
afte
r ex
posu
re to
A)
cadm
ium
chl
orid
e, B
) ta
mox
ifen
and
C)
mif
epri
ston
e. A
t a c
once
ntra
tion
of
100µ
M th
ere
wer
e si
gnif
ican
t dec
reas
es in
via
bili
ty a
nd p
roli
fera
tion
in b
oth
mon
ocul
ture
d an
d co
-cul
ture
d H
EE
Cs.
0
0.01
1
100
0.0
0.5
1.0
1.5
2.0
2.5
0
0.01
1
100
0.0
0.5
1.0
1.5
0
0.01
1
100
0.0
0.5
1.0
1.5
0
0.01
1
100
0.0
0.5
1.0
1.5
2.0
2.5
Con
cent
ratio
n µM
Con
cent
ratio
n µM
Absorbance Absorbance
** *
*
Viab
ility
Prol
ifera
tionM
onoc
ultu
re
Mon
ocul
ture
Co-
cultu
re
Co-
cultu
re
Vehicl
e0.0
1
0.1
1
1010
00.0
1
1
0.0
0.1
0.2
0.3
0.4
0.5
Vehicl
e0.0
1
0.1
1
1010
00.0
1
1
0.0
0.1
0.2
0.3
0.4
0.5
Vehicl
e0.0
1
0.1
1
1010
00.0
1
1
0.0
0.5
1.0
1.5
2.0
2.5
Vehicl
e0.0
1
0.1
1
1010
00.0
1
1
0.0
0.5
1.0
1.5
2.0
2.5
Con
cent
ratio
n µM
Con
cent
ratio
n µM*
* **
= m
ifepr
isto
ne=
prog
este
rone
= le
vono
rges
trel
Mon
ocul
ture
Co-
cultu
re
Mon
ocul
ture
Co-
cultu
re
Vehicl
e
0.01
1
100
0.01
0.0
0.5
1.0
1.5
Vehicl
e
0.01
1
100
0.01
0.0
0.5
1.0
1.5
2.0
2.5
Vehicl
e
0.01
1
100
0.01
0.0
0.5
1.0
1.5
Vehicl
e
0.01
1
100
0.01
0.0
0.5
1.0
1.5
2.0
2.5
Con
cent
ratio
n µM
Con
cent
ratio
n µM
* *
* *
Mon
ocul
ture
Co-
cultu
re
Vehi
cle
Tam
oxife
n17
β-Es
trad
iol
Mon
ocul
ture
Co-
cultu
re
A) C
adm
ium
chl
orid
eB)
Tam
oxife
n C)
Mife
prist
one
36
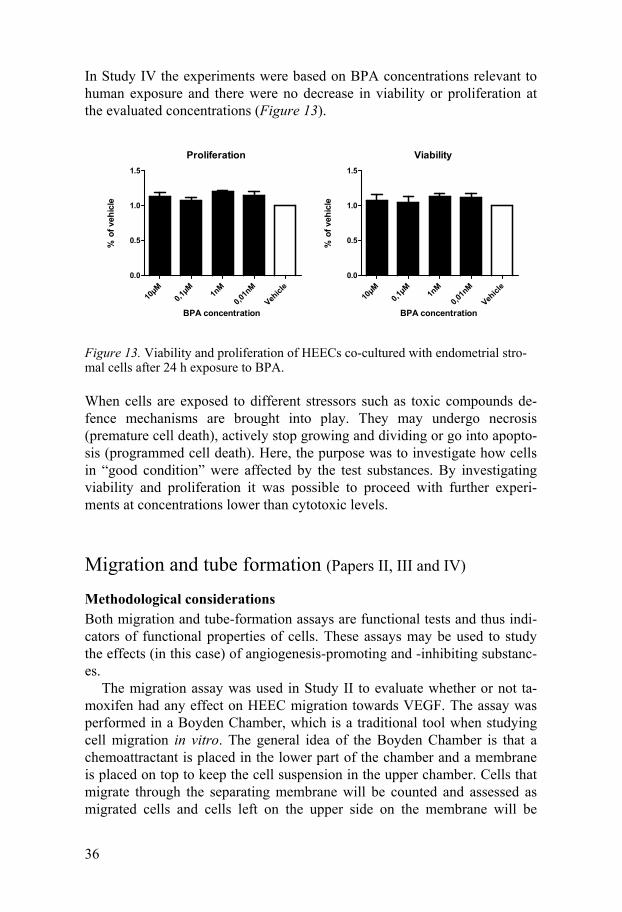
In Study IV the experiments were based on BPA concentrations relevant to human exposure and there were no decrease in viability or proliferation at the evaluated concentrations (Figure 13).
Figure 13. Viability and proliferation of HEECs co-cultured with endometrial stro-mal cells after 24 h exposure to BPA.
When cells are exposed to different stressors such as toxic compounds de-fence mechanisms are brought into play. They may undergo necrosis (premature cell death), actively stop growing and dividing or go into apopto-sis (programmed cell death). Here, the purpose was to investigate how cells in “good condition” were affected by the test substances. By investigating viability and proliferation it was possible to proceed with further experi-ments at concentrations lower than cytotoxic levels.
Migration and tube formation (Papers II, III and IV)
Methodological considerations Both migration and tube-formation assays are functional tests and thus indi-cators of functional properties of cells. These assays may be used to study the effects (in this case) of angiogenesis-promoting and -inhibiting substanc-es.
The migration assay was used in Study II to evaluate whether or not ta-moxifen had any effect on HEEC migration towards VEGF. The assay was performed in a Boyden Chamber, which is a traditional tool when studying cell migration in vitro. The general idea of the Boyden Chamber is that a chemoattractant is placed in the lower part of the chamber and a membrane is placed on top to keep the cell suspension in the upper chamber. Cells that migrate through the separating membrane will be counted and assessed as migrated cells and cells left on the upper side on the membrane will be
Proliferation
10µM
0,1µM 1n
M
0,01n
M
Vehicl
e0.0
0.5
1.0
1.5
BPA concentration
% o
f veh
icle
Viability
10µM
0,1µM 1n
M
0,01n
M
Vehicl
e0.0
0.5
1.0
1.5
BPA concentration%
of v
ehic
le
37
washed away. This is a reliable assay to study whether or not investigated cells are able to react to certain stimuli, and if that reaction can be hampered by an additional factor such as tamoxifen (in this case). A criticism of this assay is that the chemotactic gradient is steep and only transiently estab-lished. Further, there is no control over the rate of cell migration and caution must be taken to prepare a single cell suspension of cells for loading. Moni-toring cell migration in real time is technically complicated. However, it is easy to set up, inexpensive and gives a good picture of functional effects.
The tube-formation assay was used in Studies III and IV to investigate the effects of mifepristone and bisphenol A in this regard. This assay measures the ability of endothelial cells to form capillary-like structures on extracellu-lar matrix membrane. It is an assay widely used to evaluate angiogenic ef-fects of exogenous substances 82,83. The benefits of this assay are that it is relatively easy to set up, the culture period is relatively short, it is easy to quantify, and it is suitable for many samples at a time. A drawback of this assay is that variation among different lots of endothelial cells could affect tube formation and also matrix lot variation could interfere. In the present work only primary endothelial cells were investigated. Primary cells have their pros and cons, but no lot variation is applicable. Lot variation in matrix was excluded by ordering all matrix at the same time and controlling the lot number. The image-processing and quantification method used here was the Wimasis WimTube application. Wimtube has been compared with other quantifying methods to evaluate tube formation, and the conclusion was that it is easy to use and accurately quantifies angiogenesis 87.
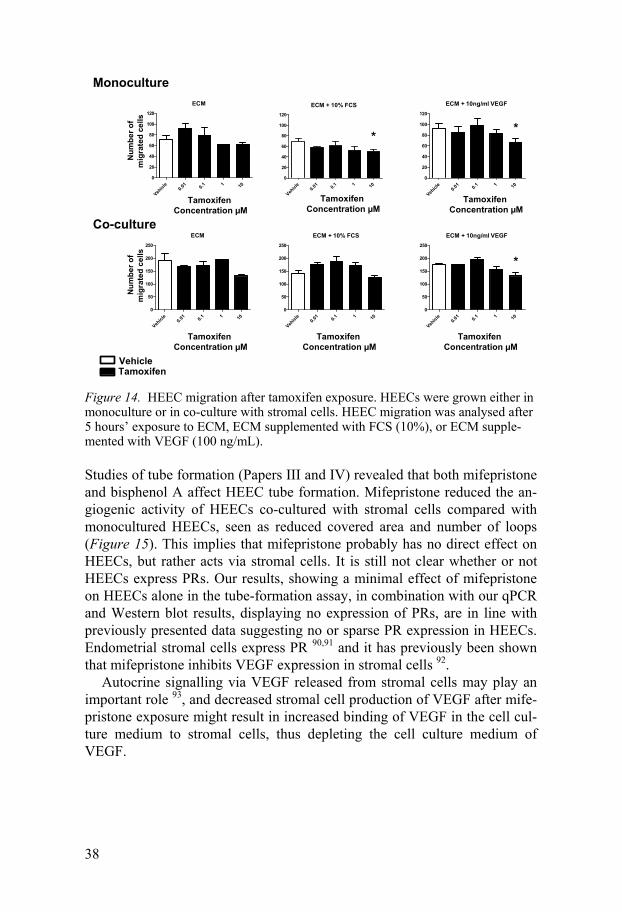
Results of migration and tube-formation assays In Study II it was demonstrated that tamoxifen (10 µM) inhibited VEGF-induced cell migration (Figure 14). It thus seems as if tamoxifen has an in-hibitory effect on angiogenesis, which is in line with results reported by oth-ers 71,88,89. The migration assay demonstrated that 48 hours’ exposure to 10 µM tamoxifen reduced VEGF-stimulated migration of HEECs in both mono- and co-cultures and reduced FCS-stimulated migration of HEECs in monocultures (Figure 14). It seemed as if HEECs in co-cultures migrated to a greater extent than HEECs grown in monocultures, but this difference was not statistically significant. It has previously been shown that tamoxifen increases VEGF production in endometrial stromal cells 89. One might spec-ulate that the tendency towards a greater extent of HEEC migration in co-cultures might be caused by VEGF production and release from stromal cells. It is also possible that the lesser extent of HEEC migration in monocul-tured HEECs compared with co-cultured HEECs is caused by down regula-tion of VEGF-mRNA after tamoxifen exposure.
38
Figure 14. HEEC migration after tamoxifen exposure. HEECs were grown either in monoculture or in co-culture with stromal cells. HEEC migration was analysed after 5 hours’ exposure to ECM, ECM supplemented with FCS (10%), or ECM supple-mented with VEGF (100 ng/mL).
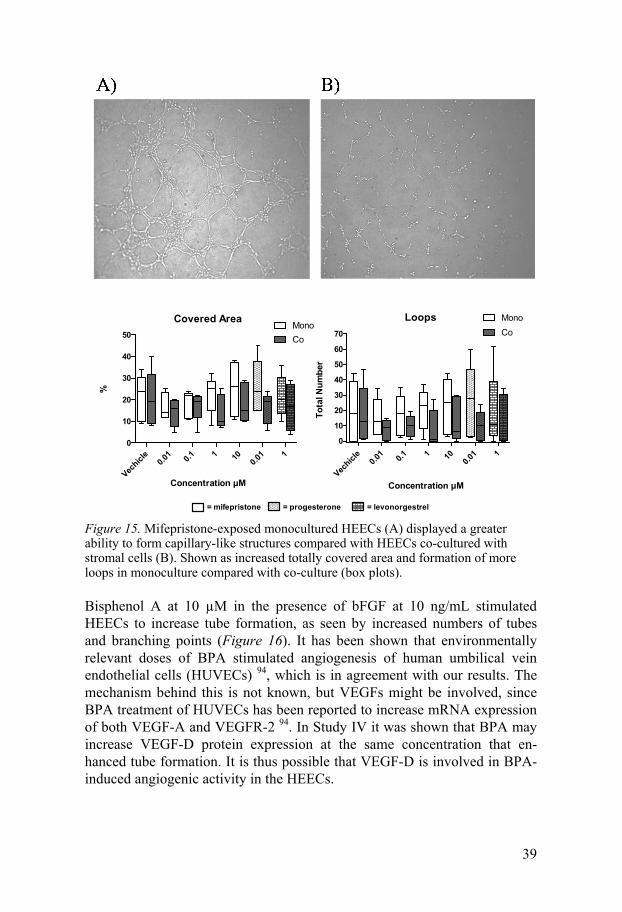
Studies of tube formation (Papers III and IV) revealed that both mifepristone and bisphenol A affect HEEC tube formation. Mifepristone reduced the an-giogenic activity of HEECs co-cultured with stromal cells compared with monocultured HEECs, seen as reduced covered area and number of loops (Figure 15). This implies that mifepristone probably has no direct effect on HEECs, but rather acts via stromal cells. It is still not clear whether or not HEECs express PRs. Our results, showing a minimal effect of mifepristone on HEECs alone in the tube-formation assay, in combination with our qPCR and Western blot results, displaying no expression of PRs, are in line with previously presented data suggesting no or sparse PR expression in HEECs. Endometrial stromal cells express PR 90,91 and it has previously been shown that mifepristone inhibits VEGF expression in stromal cells 92.
Autocrine signalling via VEGF released from stromal cells may play an important role 93, and decreased stromal cell production of VEGF after mife-pristone exposure might result in increased binding of VEGF in the cell cul-ture medium to stromal cells, thus depleting the cell culture medium of VEGF.
ECM
Vehicl
e0.0
1 0.1 1 100
20
40
60
80
100
120 ECM + 10% FCS
Vehicl
e0.0
1 0.1 1 100
20
40
60
80
100
120
ECM + 10ng/ml VEGF
Vehicl
e0.0
1 0.1 1 100
20
40
60
80
100
120
ECM
Vehicl
e0.0
1 0.1 1 100
50
100
150
200
250
ECM + 10% FCS
Vehicl
e0.0
1 0.1 1 100
50
100
150
200
250
ECM + 10ng/ml VEGF
Vehicl
e0.0
1 0.1 1 100
50
100
150
200
250
Tamoxifen Concentration µM
Num
ber o
fm
igra
ted
cells
Num
ber o
fm
igra
ted
cells
*
Monoculture
Co-culture
*
*
VehicleTamoxifen
Tamoxifen Concentration µM
Tamoxifen Concentration µM
Tamoxifen Concentration µM
Tamoxifen Concentration µM
Tamoxifen Concentration µM
39
Figure 15. Mifepristone-exposed monocultured HEECs (A) displayed a greater ability to form capillary-like structures compared with HEECs co-cultured with stromal cells (B). Shown as increased totally covered area and formation of more loops in monoculture compared with co-culture (box plots).
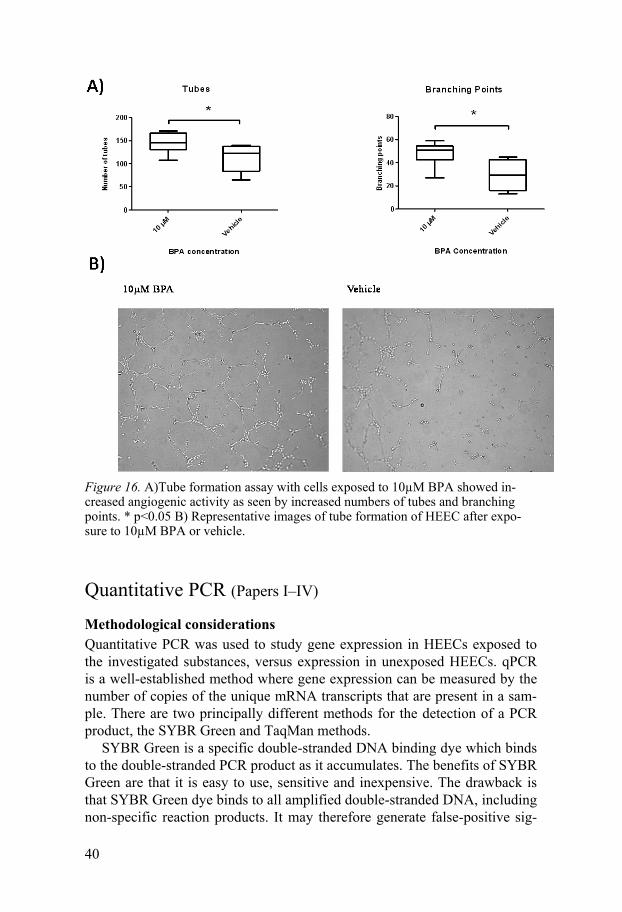
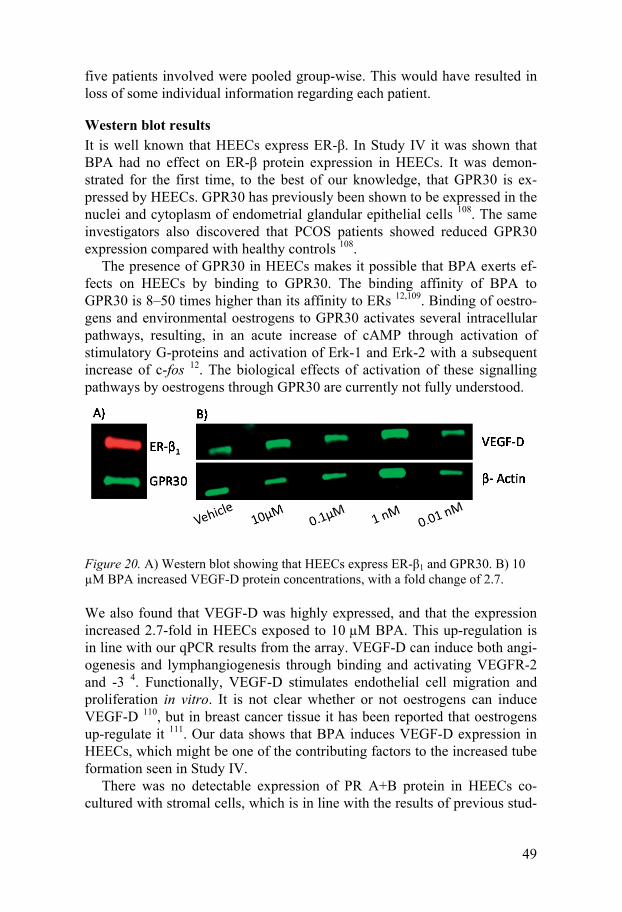
Bisphenol A at 10 µM in the presence of bFGF at 10 ng/mL stimulated HEECs to increase tube formation, as seen by increased numbers of tubes and branching points (Figure 16). It has been shown that environmentally relevant doses of BPA stimulated angiogenesis of human umbilical vein endothelial cells (HUVECs) 94, which is in agreement with our results. The mechanism behind this is not known, but VEGFs might be involved, since BPA treatment of HUVECs has been reported to increase mRNA expression of both VEGF-A and VEGFR-2 94. In Study IV it was shown that BPA may increase VEGF-D protein expression at the same concentration that en-hanced tube formation. It is thus possible that VEGF-D is involved in BPA-induced angiogenic activity in the HEECs.
Covered Area
Vechicl
e0.0
1 0.1 1 10 0.01 1
0
10
20
30
40
50MonoCo
Concentration µM
%
Loops
Vechicl
e0.0
1 0.1 1 10 0.01 1
0
10
20
30
40
50
60
70
MonoCo
Concentration µM
Tota
l Num
ber
= mifepristone = progesterone = levonorgestrel
40
Figure 16. A)Tube formation assay with cells exposed to 10µM BPA showed in-creased angiogenic activity as seen by increased numbers of tubes and branching points. * p<0.05 B) Representative images of tube formation of HEEC after expo-sure to 10µM BPA or vehicle.
Quantitative PCR (Papers I–IV)
Methodological considerations Quantitative PCR was used to study gene expression in HEECs exposed to the investigated substances, versus expression in unexposed HEECs. qPCR is a well-established method where gene expression can be measured by the number of copies of the unique mRNA transcripts that are present in a sam-ple. There are two principally different methods for the detection of a PCR product, the SYBR Green and TaqMan methods.
SYBR Green is a specific double-stranded DNA binding dye which binds to the double-stranded PCR product as it accumulates. The benefits of SYBR Green are that it is easy to use, sensitive and inexpensive. The drawback is that SYBR Green dye binds to all amplified double-stranded DNA, including non-specific reaction products. It may therefore generate false-positive sig-
41
nals and consequently optimization is very important. The TaqMan method involves use of a fluorogenic probe to enable the detection of a specific PCR product as it accumulates during PCR cycles. Specific hybridization between probe and target is required to generate a fluorescent signal, significantly reducing background interference and false-positives. TaqMan probes are very specific and require little optimization; the drawback is that they are expensive to synthesize.
In Studies I–III the genes investigated were selected because they are as-sociated with important endothelial pathways or connected to steroid path-ways. In Study IV an array of 92 genes associated with angiogenesis was used to screen for changes in gene expression when HEECs were exposed to BPA.
Results of qPCRs The oestrogenic compounds cadmium chloride, tamoxifen and 17β-oestradiol induced changes in the expression of several evaluated genes, while mifepristone did not. Among the studied genes it was clear that the expression of VEGF family genes was affected by exposure to both cadmi-um and tamoxifen (Table 3.).
Table 3. General effects of cadmium, tamoxifen and mifepristone on gene expression
Cadmium Tamoxifen Mifepristone
M C M C M C
PlGF = =
VEGF-A = = =
VEGFR-1 = =
= = =
VEGFR-2 = =
= = =
HIF-1α = =
ER-β = = =
- -
PR A+B - - - - / /
(↑) up-regulation (↓) down-regulation (=) = no difference compared with untreated cells (-) = not investigated ( / ) = no detectable expression (M) = monoculture (C) = co-culture
42
In almost all cases the changes seen in the expression of genes were opposite to each other when comparing HEECs in monocultures with those in co-cultures with stromal cells. There is thus some stromal cell effect on HEECs. Cells in monocultures are in an environment that is far from normal. A co-culture system is closer to the in vivo situation, since the studied cells have the opportunity to interact with other cell types such as stromal cells. Data from co-culture experiments may therefore be more valid when trying to understand what is going on in vivo.
PlGF Both cadmium and tamoxifen up-regulated PlGF. Previous studies have shown that PlGF is up-regulated in tissues in several different pathological conditions, although its physiological role is still debated 95. PlGF was the second member of the VEGF family to be discovered, but its importance as an angiogenic factor has been underestimated. Structurally it is very similar to VEGF-A, even though they share only 42% amino acid sequence identity. Despite this similarity, PlGF binds exclusively and with a higher affinity to VEGFR-1 compared with the other members of the VEGF-family. However, it is also known that PlGF has the ability to activate VEGFR-2 indirectly in different ways. One is by binding to VEGFR-1, thereby blocking binding sites for VEGF-A, which then, to a greater extent can bind to and activate VEGFR-2. Another mechanism is seen when PlGF and VEGF-A are pro-duced by the same cell, when they may generate heterodimers that are able to activate VEGFR-1 and they may also induce dimerization of VEGFR-1/VEGFR-2 96. A third alternative is that once VEGFR-1 has bound PlGF, VEGFR-2 may be activated by transphosphorylation. Depending on the situ-ation, PlGF can have either pro- or anti-angiogenic properties. When PlGF is produced outside endothelial cells and binds to VEGFR-1 on endothelial cells it may stimulate angiogenesis. When produced inside endothelial cells PlGF/VEGF dimers may be formed, resulting in an inhibiting effect on angi-ogenesis 95,97. Data from Studies I and II showed up-regulation of PlGF in HEECs co-cultured with stromal cells. This implies that both cadmium and tamoxifen may have an inhibiting effect on angiogenesis.
VEGF-A Cadmium chloride, but not tamoxifen or mifepristone, up-regulated VEGF-A in HEECs co-cultured with stromal cells. Cadmium is reported to be an oestrogenic substance, and oestrogen is known to stimulate the expression of VEGF-A mRNA 4. VEGF-A has previously been shown to be important for normal angiogenesis in the endometrium 98. It has also been suggested that VEGF-A is important for maintenance of differentiated status in quiescent endothelial cells 23. However, the precise role of VEGF-A in the endometri-um is not clear 4,99. It has been proposed that oestrogen regulates endometrial
43
angiogenesis mainly by regulating the production and secretion of angiogen-ic factors such as VEGF-A by endometrial epithelial cells 28.
VEGF-A may bind and signal via both VEGFR-1 and VEGFR-2. VEGFR-2 mediates VEGF-A-stimulated endothelial cell proliferation. VEGFR-1 seems to be more of a negative regulator of VEGF-A activity, but appears to be involved in endothelial cell migration and the immune system. HEECs, in comparison with other endothelial cells, are highly responsive to VEGF 33. This difference in response to VEGF makes it important to study HEECs and not to make assumptions regarding the effects of VEGFs on HEECs from data obtained from studies of other types of endothelial cells. In Studies I and II it was shown that monocultured HEECs exposed to cad-mium and tamoxifen displayed reduced VEGF-A expression. The presence of stromal cells abolished the tamoxifen-induced down-regulation of VEGF-A mRNA. This suggests that HEEC expression of VEGF-A mRNA is also under the control of endometrial stromal cells. Tamoxifen has been reported to increase VEGF production in human endometrial stromal cells 89 and it is possible that VEGF released from stromal cells regulates VEGF-A expres-sion in HEECs via paracrine mechanisms.
VEGFR-1 and VEGFR-2 Tamoxifen, but not cadmium chloride or mifepristone, changed HEEC ex-pression of both VEGFR-1 and VEGFR-2. VEGFR-1, which was up-regulated by tamoxifen, is a strong negative regulator of angiogenesis during embryogenesis – VEGFR-1 knock-out mice die at an early embryonic stage as a result of overgrowth and disorganization of blood vessels 100. In adult-hood the effect of VEGFR-1 on angiogenesis is less clear. Over-expression of VEGFR-1 in endothelial cells has been shown not to result in enhanced proliferative activity 97,100. Over-expression of VEGFR-1 is also associated with pre-eclampsia 101, although the association is more related to soluble VEGFR-1, which acts like a decoy to VEGF-A, and may thereby regulate the amount of VEGF-A that is free to bind to VEGFR-2 97, which is consid-ered to be a more central receptor in the angiogenic cascade. Tamoxifen, as a consequence of the up-regulation seen here, may have an inhibitory effect on endometrial angiogenesis by increasing the expression of VEGFR-1.
VEGFR-2 was slightly down-regulated by tamoxifen in HEECs grown in monoculture. Since this was only seen in monoculture and not in co-cultured HEECs the biological consequence is unclear. VEGFR-2, however, is the crucial receptor for mediating angiogenic activity and vascular permeability.
HIF-1α Both cadmium chloride and tamoxifen, but not mifepristone, changed the expression of HIF-1α which is a central regulatory factor in the control of angiogenesis. In hypoxic conditions HIF-1α binds to the VEGF-A promoter and thereby increases the production of VEGF-A 102. Over-expression of
44
HIF-1α stimulates and promotes endothelial cell migration and tubular or-ganization 102.
Cadmium reduced and tamoxifen up-regulated HIF-1α expression in HEECs co-cultured with endometrial stromal cells. These results are in line with the concept that oestrogen has an inhibitory effect on HIF-1α expres-sion through ER-β 103. These results are also in line with the oestrogenic properties of cadmium and the anti-oestrogenic properties of tamoxifen.
ER-β Tamoxifen up-regulated, and cadmium did not affect, the expression of ER-β in HEECs co-cultured with stromal cells. An increase in ER-β expression has also been shown in the endometrial vasculature when the levels of oes-trogen and progesterone decrease in the late secretory phase of the menstrual cycle 6. The tamoxifen-induced ER-β expression might be related to a feed-back mechanism of anti-estrogenic activity of tamoxifen.
Progestogens As mentioned earlier, it is not clear whether or not HEECs express proges-terone receptors. The results presented in this work support the concept that HEECs do not express PRs. Exposure of HEECs to mifepristone, levonorg-estrel or progesterone resulted in no detectable changes in the investigated genes, including those for the androgen receptor and the glucocorticoid re-ceptor. However, it was shown that HEECs contain mRNA encoding both the androgen receptor and the glucocorticoid receptor, but not PR A+B.
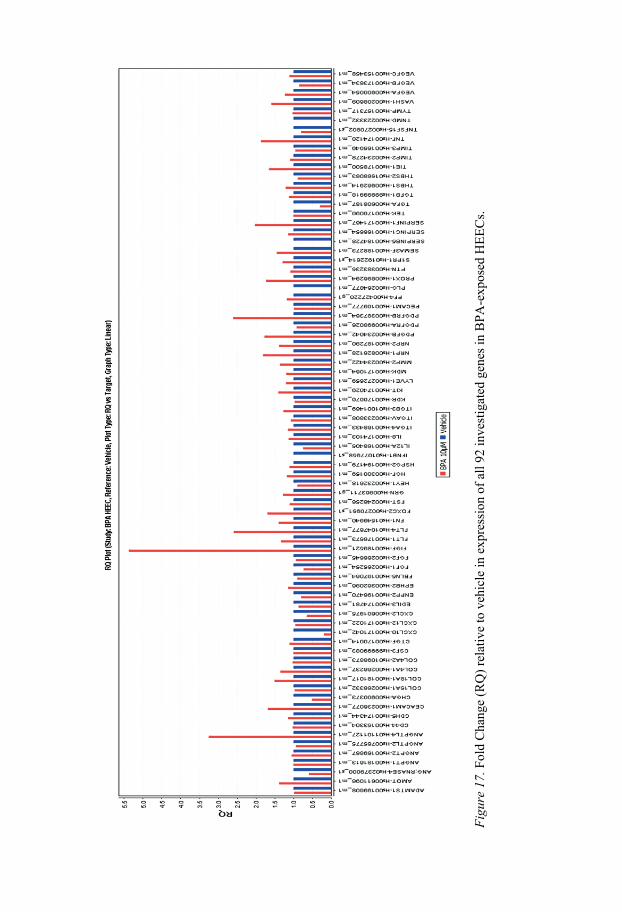
Bisphenol A Bisphenol A showed effects on members of the VEGF family in the qPCR array (Figure 17). These results, however, could not be fully verified in all cell cultures. The reason for this is not known, but there are some possible explanations. It might be that there is no real difference in mRNA levels or that our study population was too small, or that our primary cells were too heterogeneous. We believe that BPA induced increased expression of VEGF-D mRNA, since this is in agreement with increased expression of VEGF-D protein.
F
igur
e 17
. Fol
d C
hang
e (R
Q)
rela
tive
to v
ehic
le in
exp
ress
ion
of a
ll 92
inve
stig
ated
gen
es in
BPA
-exp
osed
HE
EC
s.
46
Expression of metabolizing enzymes in the human endometrium
Methodological considerations Immunohistochemistry was used to determine the presence and cellular dis-tribution of different tamoxifen-metabolizing enzymes. The principal of immunohistochemistry is the detection of antigens in tissue sections by spe-cific antibodies. This technique yields a semi-quantitative measurement of the relative amounts of protein in a tissue section and it is important to re-member that absolute amounts of protein cannot be determined by immuno-histochemistry.
In general, immunohistochemistry enables visualization of antigens in frozen or paraffin-embedded tissues. First, a primary antibody (specific for the antigen) is added, then a secondary biotinylated antibody detecting the primary antibody is added, and subsequently a chromogenic substance. An-tigen-antibody binding is detected by a colour reaction and visualised under a microscope.
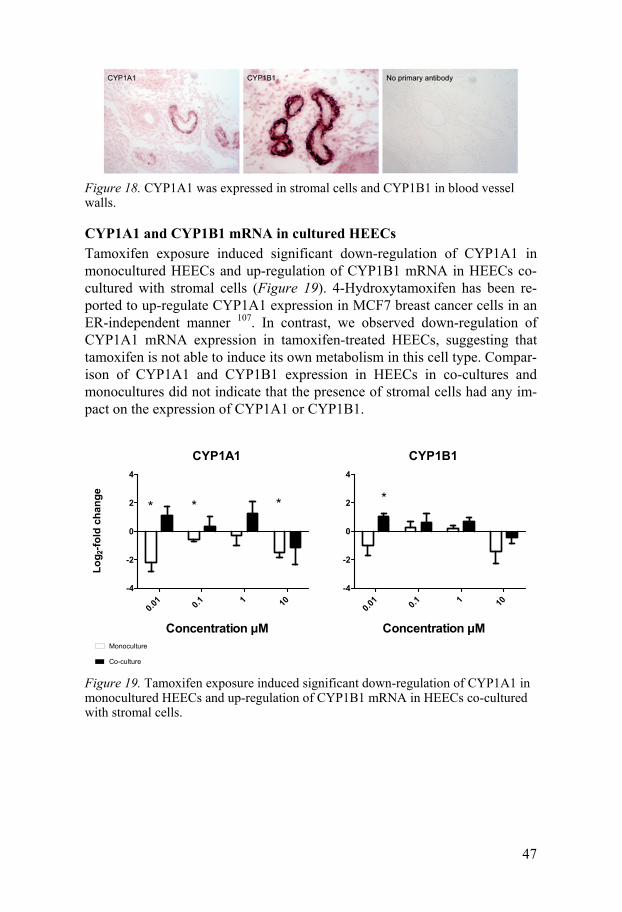
Results of protein expression analysis There is an array of tamoxifen metabolites with mixed agonistic and antago-nistic effects on oestrogen receptors, and 4-hydroxytamoxifen and endoxifen are reported to be the most pharmacologically active 104,105. Several cyto-chrome P (CYP) enzymes can metabolize tamoxifen. We have previously shown selective expression of several tamoxifen-metabolizing CYPs in glandular and surface epithelia, as well as adduct formation in human endo-metrial biopsy samples 106. In agreement with that study of endometrial glands we found no major tamoxifen-metabolizing CYPs in the endometrial vasculature, but there was distinct expression of CYP1A1 in the stroma around endometrial blood vessels and CYP1B1 in the endometrial blood vessel walls (Figure 18).
The results suggest potential bioactivation of tamoxifen by CYP1A1/1B1 in the endometrial vasculature in vivo. However, it is most reasonable to suppose that the major effects of tamoxifen seen in this in vitro study were related to the parent compound, since the expression levels were relatively low in HEECs and stromal cells. It must be emphasized that in vivo, tamoxi-fen undergoes significant hepatic metabolism and forms metabolites that may reach the endometrial vasculature via the systemic circulation.
47
Figure 18. CYP1A1 was expressed in stromal cells and CYP1B1 in blood vessel walls.
CYP1A1 and CYP1B1 mRNA in cultured HEECs Tamoxifen exposure induced significant down-regulation of CYP1A1 in monocultured HEECs and up-regulation of CYP1B1 mRNA in HEECs co-cultured with stromal cells (Figure 19). 4-Hydroxytamoxifen has been re-ported to up-regulate CYP1A1 expression in MCF7 breast cancer cells in an ER-independent manner 107. In contrast, we observed down-regulation of CYP1A1 mRNA expression in tamoxifen-treated HEECs, suggesting that tamoxifen is not able to induce its own metabolism in this cell type. Compar-ison of CYP1A1 and CYP1B1 expression in HEECs in co-cultures and monocultures did not indicate that the presence of stromal cells had any im-pact on the expression of CYP1A1 or CYP1B1.
Figure 19. Tamoxifen exposure induced significant down-regulation of CYP1A1 in monocultured HEECs and up-regulation of CYP1B1 mRNA in HEECs co-cultured with stromal cells.
CYP1A1
0.01 0.1 1 10
-4
-2
0
2
4
* * *
Concentration µM
CYP1B1
0.01 0.1 1 10
-4
-2
0
2
4
*
Concentration µM
Log 2
-fold
cha
nge
Monoculture
Co-culture
48
ELISA
Methodological considerations The amounts of VEGF in the cell culture media were analysed by using a VEGF sandwich ELISA. In the sandwich ELISA technique a capturing anti-body specific for human VEGF165 was pre-coated onto a microplate. Colour intensity is in proportion to the amount VEGF bound. The advantages of ELISA are that it is relatively quick and easy to perform; it is a safe and spe-cific method that is widely used.
ELISA results Secretion of VEGF from HEECs into cell culture media was not changed when the cells were exposed to BPA. The VEGF levels in the media de-clined relatively fast, regardless of treatment, from an average of 1778 pg/mL in fresh media to an average of 30 pg/mL after 24 h. There was no difference in VEGF concentration in fresh media stored at 4 °C overnight or in media incubated in a culture dish at 37 °C overnight. From this it can be concluded that the decline of VEGF concentrations is not caused by nonspe-cific binding to the plastics in the cell culture dish.
The observed decline in the content of VEGF in the culture media might be a result of high binding of VEGF to VEGF receptors on HEECs. In previ-ous studies it has been shown that HEECs have a higher VEGF binding ca-pacity than a number of other endothelial cell types, namely, human coro-nary endothelial cells, human umbilical vein endothelial cells, human dermal endothelial cells, suggesting that HEECs express a high number of VEGF receptors 33. The decline in VEGF might also explain why BPA had the abil-ity to affect HEEC tube formation only in the presence of bFGF, which, like VEGF, also promotes normal angiogenesis. It is known that shortage of VEGF causes endothelial cell dysfunction, which might be the reason why HEECs do not respond to the presence of BPA without bFGF.
Western Blot
Methodological considerations The Western blot technique was used to detect specific proteins in extracts from BPA-treated HEECs co-cultured with stromal cells. Gel electrophoresis was used to separate denatured proteins by their size. The proteins were then transferred to a PVDF membrane and stained with antibodies specific to the target protein. Results were normalized to β-actin, which is one of the three major components in the cytoskeleton and therefore highly conserved. Be-cause of the low protein yield after RNA/protein extraction, samples from all
49
five patients involved were pooled group-wise. This would have resulted in loss of some individual information regarding each patient.
Western blot results It is well known that HEECs express ER-β. In Study IV it was shown that BPA had no effect on ER-β protein expression in HEECs. It was demon-strated for the first time, to the best of our knowledge, that GPR30 is ex-pressed by HEECs. GPR30 has previously been shown to be expressed in the nuclei and cytoplasm of endometrial glandular epithelial cells 108. The same investigators also discovered that PCOS patients showed reduced GPR30 expression compared with healthy controls 108.
The presence of GPR30 in HEECs makes it possible that BPA exerts ef-fects on HEECs by binding to GPR30. The binding affinity of BPA to GPR30 is 8–50 times higher than its affinity to ERs 12,109. Binding of oestro-gens and environmental oestrogens to GPR30 activates several intracellular pathways, resulting, in an acute increase of cAMP through activation of stimulatory G-proteins and activation of Erk-1 and Erk-2 with a subsequent increase of c-fos 12. The biological effects of activation of these signalling pathways by oestrogens through GPR30 are currently not fully understood.
Figure 20. A) Western blot showing that HEECs express ER-β1 and GPR30. B) 10 µM BPA increased VEGF-D protein concentrations, with a fold change of 2.7.
We also found that VEGF-D was highly expressed, and that the expression increased 2.7-fold in HEECs exposed to 10 µM BPA. This up-regulation is in line with our qPCR results from the array. VEGF-D can induce both angi-ogenesis and lymphangiogenesis through binding and activating VEGFR-2 and -3 4. Functionally, VEGF-D stimulates endothelial cell migration and proliferation in vitro. It is not clear whether or not oestrogens can induce VEGF-D 110, but in breast cancer tissue it has been reported that oestrogens up-regulate it 111. Our data shows that BPA induces VEGF-D expression in HEECs, which might be one of the contributing factors to the increased tube formation seen in Study IV.
There was no detectable expression of PR A+B protein in HEECs co-cultured with stromal cells, which is in line with the results of previous stud-
50
ies 36. However, progesterone has been shown to have effects on HEECs, which indicates an alternative non-genomic effect of progesterone in these cells 36.
51
Summary of findings
This cell culture model is not suitable for high-through put screening of EDCs in general. The model, however, does provide valuable information on how chemicals with oestrogenic and progestational characteristics affect HEECs in vitro in mono- and co-cultures with stromal cells.
Paper I Cadmium influences mRNA expression of two important angiogenesis-related genes, namely those encoding VEGF-A and PlGF, and the effect of cadmium exposure on HEECs depends on the presence or absence of endo-metrial stromal cells.
Paper II Tamoxifen decreases HEEC migration and modulates the expression of an-giogenesis-related genes, indicating that tamoxifen affects endometrial angi-ogenesis, most likely by reducing angiogenic activity. Our results also indi-cate that endometrial stromal cells regulate some of the effects of tamoxifen on HEECs.
Paper III Exposure of HEECs to mifepristone resulted in significantly reduced angio-genic activity when the cells were co-cultured with endometrial stromal cells compared with HEECs in monoculture. This suggests that the effect of mife-pristone on HEECs is mediated through stromal cells and that mifepristone seems to induce changes in endometrial angiogenic activity.
Paper IV Bisphenol A increases HEEC tube formation and induces VEGF-D protein and mRNA expression, and may thus disturb normal endometrial vascular development.
52
General Discussion
This thesis presents data obtained from primary human endometrial endothe-lial cells grown in vitro and exposed to different EDCs. The results indicate that HEECs are sensitive to EDCs and it is possible that human exposure could lead to negative effects in the human endometrial vasculature. The widespread use of chemicals and the high content in consumables has led to ubiquitous exposure of humans and animals to EDCs. In recent years nu-merous epidemiological studies have revealed associations between human exposure to EDCs and various diseases 38,46,112.
The amount of published animal data regarding the effects exposure to EDCs has become more comprehensive 46,113. A permanent problem, howev-er, is that extrapolation from animals to humans cannot be done without caution, since significant species and strain differences have been reported 113. There has also been extensive use of tissue and cell cultures during the last century that has generated a great amount of knowledge and understand-ing of regulatory pathways and interactions between cells. One way of link-ing data from animal experiments and experiments on different cell lines or cells of animal origin to humans would be to perform studies on primary cells of human origin. Cell culture experiments using cells of human origin might thus be a feasible way to gain knowledge of possible effects of EDCs on human tissues and organs and subsequent effects on human reproductive functions.
In the present work we have shown that HEECs grown in vitro are sensi-tive to exposure to hormonally active pharmaceutical drugs and endocrine disrupting chemicals by responding with changes in the expression of genes and proteins important for the control of cell migration and angiogenic activ-ity. It has previously been shown that disturbed endometrial vasculature seems to be associated with conditions such as menorrhagia, endometriosis, infertility, implantation failure, and pre-eclampsia 101,114-117. All of these con-ditions are probably related to impaired control of angiogenic factors, espe-cially VEGFs 114. It is possible that the decline in female fertility seen during the last half century could be related to the ability of EDCs to affect angio-genic factors. It has, for instance, been shown that arrest of porcine foetuses is associated with declined transcription of VEGF in endometrial endothelial cells 118. In humans, it has been shown that intrauterine application of human chorionic gonadotrophin (hCG) increases intrauterine VEGF levels, suggest-ing involvement of angiogenesis during early pregnancy, a process essential
53
for the formation of a functional placenta 119. Uterine fluid from women with unexplained infertility has been reported to contain significantly lower levels of VEGF compared with fluid from fertile women 120. Hannan et al. also reported that uterine fluid from fertile women, or pure VEGF itself, en-hanced blastocyst outgrowth, indicating that VEGF may have substantial effects on implantation 120. All this combined emphasizes the importance of VEGF in achieving and maintaining a successful pregnancy.
We now show that the substances investigated have the ability to change the expression of several members of the VEGF family. These substances might thus increase the risk of menstrual bleeding disturbances, endometrio-sis, fertility problems and pregnancy complications. These conditions are not only related to reproduction, but also to everyday life and may have a great impact on the overall wellbeing and quality of life of women. All of the above-mentioned conditions are related to increased healthcare consumption, presenteeism and absence from work, and may thus have a great impact on health economy.
Hormones, which fine-tune the whole endocrine system, including control of our reproductive organs, are powerful substances, and minor changes in serum and tissue levels can cause major functional changes. Chemicals with endocrine-disrupting capacity have the ability to disturb these fine-tuned systems, especially those depending on the sex steroid hormones. Even if the hormonal effect of a certain chemical is known it is hard to predict what biological effects will be seen after exposure. It is therefore important to investigate different cell types and tissues, since similar cell types from dif-ferent tissues have different features. This applies to endothelial cells, which have different properties depending on their organ or tissue of origin in the body.
An advantage of studying the effects of EDCs on primary cells is that they are functionally closer to the in vivo situation compared with cell lines. Disadvantages with primary cells include inter-individual variations and the fact that they are demanding to isolate and culture. For practical reasons it is hard to isolate and culture more than a few cell isolates in each experiment, which is an important fact to consider when interpreting data. Because of these disadvantages, primary HEECs are not suitable for high-throughput screening of EDCs.
Our results indicate that oestrogenic substances act directly on HEECs, while progestational compounds most likely have an indirect effect on HEECs via the stromal cells. The progestational compounds mifepristone, levonorgestrel and progesterone itself did not affect the expression of any of the investigated genes. As mentioned earlier, it is not completely clear whether or not HEECs express progesterone receptors, but by summarizing available data from previous studies and our current data, we suggest that there is no PR expression in HEECs grown in vitro.
54
Most of the previously presented data regarding the effects of sex steroid hormones on endometrial angiogenesis have been focused on oestrogens 4. Both ER-α and -β have been detected in human endometrial vascular smooth muscle cells, and ER-β in HEECs 4. Pro-angiogenic actions of oestrogens on the endometrial vasculature are related to increased VEGF-A expression, and this has been studied both in vitro and in vivo 4.
The effect of progesterone on the endometrial vasculature is less ex-plored. There are several studies on the long-term effects of progestational contraceptives on the endometrial vasculature 121-124. However, the results are not as clear as with oestrogen, since both anti- and proangiogenic actions have been described after progesterone treatment. It is also still not known if progesterone acts directly on the endometrial vasculature or indirectly via other cell types 4.
An interesting finding in our studies was that the effects of the different compounds depended on the presence or absence of endometrial stromal cells, i.e. mono- and co-cultured endothelial cells displayed different results regarding effects on gene expression (Papers I and II) and tube formation (Paper III). It is not surprising that it matters whether the cells are grown alone or together with other cells, and it is of utmost importance to take into account when interpreting and comparing data.
The data presented here show that substances with hormonal activity af-fect the expression of several genes involved in the control of angiogenesis. However, it is important to remember that changes in the expression of genes do not correlate to functional effects per se, since the effects are main-ly related to proteins. There are several steps between a change in mRNA expression and a functional effect. It is therefore pertinent to combine stud-ies of mRNA expression, their corresponding proteins, and functional as-says. In our studies we have seen that the investigated substances have the ability to alter both mRNA and protein expression, and also HEEC migra-tion, and activity in the tube-formation assay. The cell migration and tube-formation assays are functional and provide information about whether or not the investigated substances have any effect on the functionality of HEECs.
Conclusions In summary, human endometrial endothelial cells grown in vitro are sensi-tive to the investigated hormonally active pharmaceutical drugs and EDCs. A culture model based on HEECs in monocultures or co-cultures with stro-mal cells thus constitutes a suitable small-scale test for studying the possible effects on HEECs in vitro. Whether effects seen in this model actually take place in vivo is not easily confirmed. Assume that some of the effects seen in vitro to some extent also occur in vivo. Such effects might cause changes in
55
the endometrial vasculature that subsequently could lead to impaired endo-metrial function with consequences as regards fertility and pregnancy. The connection between laboratory data and what actually happens in vivo in humans is always problematic to confirm, but the identification of potential-ly harmful substances is of utmost importance for future research. One future way to study the impact of identified chemicals would be to create an in vitro implantation model for studies of whether a chemical hampers implan-tation or disturbs the expression of endometrial receptivity markers. Such knowledge may also form a good basis for future regulations concerning the handling of chemicals.
56
Summary in Swedish
Sammanfattning på svenska Blodkärlsnybildning är en process som pågår framförallt under fostertiden medan nybildning av blodkärl hos vuxna generellt sett endast sker vid sår-läkning, i äggstockarna och i livmoderslemhinnan.
I livmoderslemhinnan (endometriet) finns det mycket blodkärl och blod-kärlsnybildning är viktigt under menstruationscykeln, huvudsakligen för att återställa endometriets funktion. Endometriets huvudsakliga funktion är att ta emot och implantera ett befruktat ägg. Vid en implantation är nätverket av blodkärl extremt viktigt eftersom det förser det blivande fostret med den näring som det behöver för att kunna fortsätta utvecklas, mogna och tillslut bli en ny människa. Näringsutbytet mellan fostret och kvinnan sker i mo-derkakan, ett mycket kärlrikt organ som börjar bildas tidigt under gravidite-ten. Om det inte sker någon implantation stöts livmoderslemhinnan ut (men-struation) för att sedan byggas upp och förberedas för en eventuell ny im-plantation.
I uppbyggnadsprocessen bildas det nya blodkärl via förlängning och ut-byggnad från de redan existerande blodkärlen. I denna process är en viss celltyp, endotelcellerna, viktiga eftersom de utgör blodkärlens innersta skikt. Då nya blodkärl ska bildas får endotelcellerna signaler om att vandra ut från det redan existerande blodkärlet och bilda nya.
Endotelcellerna är genom deras funktion vid nybildning av blodkärl vik-tiga för en normalt fungerade livmoderslemhinna och störningar i deras funktion kan leda till påverkan på blodkärlsnybildning och därigenom bidra till blödningsrubbningar, endometrios, missfall, havandeskapsförgiftning och infertilitet. I endometriet styrs endotelcellerna till viss del av svängningar i nivåerna av könshormonerna östrogen och progesteron.
Det finns kemikalier (industrikemikalier, förbränningsprodukter och lä-kemedel) som liknar östrogen och progesteron och som kan störa den natur-liga rytmen och regleringen som sker via hormoner i kroppen. Dessa kemi-kalier benämns gemensamt som ”hormonstörande kemikalier”. Hormonstö-rande kemikalier stör inte enbart regleringen av könshormoner utan även många andra hormoner och kan bland annat öka risken för diabetes, högt blodtryck, hjärt och kärlsjukdomar, ADHD, och depression.
Det övergripande syftet med den här avhandlingen var att undersöka hur några ämnen med hormonstörande egenskaper påverkar endotelceller i en-
57
dometriet. Syftet var även att utvärdera om detta cellsystem skulle kunna fungera som ett screening-verktyg, för att studera andra kemikalier och deras eventuella effekt på endometriets blodkärl. Vid nybildning av blodkärl inte-ragerar flera olika celltyper i livmoderslemhinnan, därför har vi även stude-rat hur endotelcellerna reagerar på de hormonstörande ämnena i närvaron av de celler som normalt finns i deras omgivning, stromaceller.
Det visade sig att alla de kemikalier vi undersökte, kadmium (en tungme-tall med östrogena egenskaper), tamoxifen (ett bröstcancerläkemedel som motverkar östrogen), mifepriston, (ett läkemedel för medicinsk abort och som motverkar progesteron) och bisphenol A (en industrikemikalie med östrogena egenskaper), påverkar den normala funktionen av endotelceller från human livmoderslemhinna. Närvaron av stromaceller visade sig också ha inverkan på hur endotelcellerna reagerade på kemikalierna.
Kadmium och tamoxifen påverkade genuttrycket av PlGF och VEGF, två tillväxtfaktorer som är nyckelmolekyler vid stimulering och välmående hos endotelceller. Tamoxifen hämmade även endotelcellernas förmåga att vandra, vilket tyder på eventuell hämmad förmåga att bilda nya blodkärl. Mifepriston-exponering av cellerna visade sig inte ha någon direkt effekt på endotelcellerna men när stromaceller var närvarande bildade endotelcellerna färre blodkärlsstrukturer. Bisphenol A (BPA) påverkade uttrycket av flera gener som är involverade i blodkärlsnybildning. Den gen som var mest för-ändrad av BPA var en variant av VEGF, VEGF-D, en förändring som även kunde ses på proteinnivå. Bisphenol A ökade även förmågan hos endotelcel-lerna att bilda blodkärlsstrukturer, vilket skulle kunna vara relaterat till det ökade uttrycket av VEGF-D.
Sammanfattningsvis ser vi att dessa kemikalier med hormonstörande egenskaper har förmågan att påverka gener och proteinuttryck som är viktiga för nybildning av blodkärl. Andra studier har visat att påverkan på dessa faktorer kan bidra till blödningsrubbningar, endometrios, reducerad fertilitet och havandeskapsförgiftning, sjukdomar som har stor inverkan på kvinnans liv och livskvalitet. Det är möjligt att en reducering av mängden hormonstö-rande kemikalier i vår omgivande miljö skulle kunna bidra till en minskning av dessa sjukdomar. För att kunna besluta om sådana åtgärder krävs bakom-liggande forskning, såsom studierna som presenteras i den här avhandlingen.
58
Acknowledgements
This work was carried out at the Department of Women’s and Children’s Health, Uppsala University. I would like to thank Jan Gustavsson, professor and head of the Department, and Ove Axelsson, professor and former head of the Department of Women’s and Children’s Health for the opportunity to carry out my PhD studies at this department.
I would also like to express my sincere gratitude to all the women who con-tributed in the studies.
I would especially like to thank:
Professor Matts Olovsson, Department of Women’s and Children’s Health, my main supervisor, for introducing me to this exciting field of research and for taking me on as a PhD student. You have always been supportive and positive about my work, and uplifting at the times when I have come crying .
Associate professor Anneli Stavreus-Evers, Department of Women’s and Children’s Health, my co-supervisor, for all your valuable discussions and advice over the years. I would especially like to express my appreciation and thanks for the always so rapid and excellent comments regarding my work.
Professor Inger Sundström Poromaa, associate professors Helena Åkerud and Alkistis Skalkidou, for interesting and valuable discussions throughout the years.
Professor Eva Brittebo, Department of Pharmaceutical Biosciences, and co-author, for valuable comments on the paper and for all the valuable discus-sions and advice.
Helén Andersson, PhD, Department of Pharmaceutical Biosciences, and co-author, for you much-appreciated comments on the paper and for all the discussions and good advice. Special thanks for being such a nice travel companion in Milan.
59
Elin Bannbers, PhD and Therese Fagerqvist, PhD student, my “ärtor” who I got to know the very first week when I moved to Uppsala 10 years ago, for being such wonderful friends. Thank you both for all the joy and fun we have had, and will have in the future! Elin: you have been in my life more or less daily during the past 10 years; I will miss you! I do hope you don’t end up too far away. But most importantly, as I told you at your dissertation, you mean a lot to me! Therese: I admire your ability to be rational and make things sound easy, even when they are not. I must also compliment you on finding a husband as interested in fishing as mine, lucky for us . But the most important thing is that you mean a lot to me!
Charlotte Hellgren, PhD, office roommate, for all the times you have burst into laughter hearing something funny on the radio or reading a funny name on a paper, keeping the time writing-up our work enjoyable. I really wish you the best in the future, and I will miss seeing you every day.
Karin Lindgren, co-author and fellow PhD student, for all the fun times in the cell-lab. For all the help you have given me, with both qPCR and also for when you have always wanted to know “why” (forcing me to reflect). And for your way of making people around you feel happy.
Theodora Kunovac Kallak, Katja Junus, Helena Kaihola and Tiina Murto, fellow PhD students, for all the chats we have had and for all the support you have given me. Theo: For your constant and tremendous willingness to help, and I really appreciate your company in the early mornings and our “workout dates”. Katja: For all the help and valuable discussions. I will al-ways remember our exciting trip with the night train, but next time let’s stay at a hotel! Helena: for all the help with the Western blots. Tiina: it feels like we have walked this line together, being accepted as PhD students at the same time, being robbed at our wallets at the department at the same day, and giving birth only a few months apart. I have always looked forward to the days you spend in Uppsala.
Eva Davey, laboratory technician and office roommate, for all the help in the lab, counting cells and all the work with the proteins. For the enjoyable chats and for the nice company in our room. I do hope you sleep better now when you don’t have my work keeping you awake.
Lena Moby, research nurse, for sharing breaks, lunches and laughter with me, and for all the nice time at the golf course. Special thanks for introduc-ing me to “surströmming”, which I probably never would have tried if it wasn’t for you.
60
Ulla Geifalk, laboratory technician and former office roommate, for all our conversations and laughs about everything possible. I will never be able to see or hear about Kjell Lönnå without thinking of you and your “fräckisar”. You have the gift of making people around you feel good! Also, thank you for helping me with qPCR and other laboratory matters.
Margareta Nordling, laboratory technician and former colleague, for all that you have taught me in the laboratory. For sharing lunches and breaks with me. Now when I have my own garden it would have been good to have you here for some spontaneous gardening advice during coffee breaks.
Vera Holmgren, administrator and former colleague, for sharing breaks, lunches and laughter with me. I really enjoyed all the times you came back to work.
Roger Olsson, Marie Berglund, Linda Bratteby-Tollerz, Department of Public Health and Caring Sciences. For all the good times, both in and out-side the office, and for sharing office, breaks and laughter with me. Even though you only moved to another part of the building I miss having you guys around.
The administrative staff, especially thanks to Malin, Susanne, Kerstin, Hans and Martin.
Also, all the colleagues in the research group and at the department, par-ticularly Maria Stålhammar, Jocelien, Ragnhiedur, Christian, Helena Volgsten, Signe Altmäe, Anna-Karin Hannerfors and everyone else who have helped, supported and encouraged me during these years.
This thesis was supported by grants from the EU research project Reprotect, Formas Research Council, the Swedish Fund for Research without Animal Experiments and the Family Planning Fund in Uppsala.
61
I would also like to thank my family:
Mamma and Tina, there are no words to describe how much you mean to me, I love you so much! Thank you for all the support and love and for mak-ing me who I am. Mamma: thank you for always being there when I need you; you are the best mum, ever! Tina: I will finally admit it; you are al-lowed to sing ♪♪♪Tinaaaa är bääääääst….♪♪♪ , you are simply the best big sis-ter! A special thanks to Tina and Peter for giving me the most wonderful nephew and niece, Rasmus and Julia, that one could wish for ♥
Pappa, Chrisse, Linus and Elin alias “pappa-familjen” for all the encour-agement, support and love! I know I sometimes forget to tell you, but you are all so important to me! I love you with all my heart; you are fantastic all of you!
Ann-Kristin and Leif, the best parents-in-law that one could wish for, thank you for your help in making this possible! Knowing that Bernard has a great time with you has made it easier to work late and once in a while at the weekends. Linus, Hanna, Eskil and Folke and Pontus, Sandra, Alice and Walle, my family-in-law, for being such a charming, pleasant and sociable family.
Staffan, my beloved husband and my best friend, there are not enough words to describe what you mean to me. Thank you for all your encourage-ment and love! And for all the big and small adventures you come up with that makes our life exciting and fun. I also really appreciate your efforts in making this thesis possible!
Ian, our “firstborn”, thanks for your loyal companionship and all the walks in the woods. I promise you that we will go hunting more often after this.
My precious Bernard, for your tremendous and unconditional love. For making me keep my sanity, and for forcing me to totally leave work at the office. You are my favourite little person in the whole wide world; I love you so much! ♥
62
References
1. Speroff Leon, Marc F. Hormone Biosynthesis, Metabolism, and Mechanism of Action. Clinical Gynecologic Endocrinology and Infertility. 7th ed. Philadelphia: Lippincott Williams &Wilkins; 2005:25-96.
2. SEER Training Modules A, U. S. National Institutes of Health NCI. The human uterus and uterine tubes 2013; http://training.seer.cancer.gov/images/anatomy/reproductive/uterus.jpg.
3. Jones RE, Lopez KH. The female reproductive System. Human Reproductive Biology 2006:31-73.
4. Girling JE, Rogers PA. Endometrial angiogenesis, arteriogenesis, and lymphangiogenesis. In: Aplin JD, Fazleabas AT, Glasser SR, Giudice LC, eds. The endometrium. UK2008:76-92.
5. Brenner RM, Slayden OD, Nayak NR, Baird DT, Critchley HOD. A role for the androgen receptor in the endometrial antiproliferative effects of progesterone antagonists. Steroids. 2003;68(10–13):1033-1039.
6. Speroff L, Fritz MA. Clinical gynecologic endocrinology and infertility. 7th ed. Philadelphia: Lippincott Williams & Wilkins; 2005.
7. Ball LJ, Levy N, Zhao X, et al. Cell type- and estrogen receptor subtype specific regulation of selective estrogen receptor modulator regulatory elements. Mol. Cell. Endocrinol. 2009;299(2):204-211.
8. Nilsson S, Makela S, Treuter E, et al. Mechanisms of Estrogen Action. Physiol. Rev. 2001;81(4):1535-1565.
9. Lindsey SH, Chappell MC. Evidence That the G Protein–Coupled Membrane Receptor GPR30 Contributes to the Cardiovascular Actions of Estrogen. Gender Medicine. 2011;8(6):343-354.
10. Prossnitz ER, Maggiolini M. Mechanisms of estrogen signaling and gene expression via GPR30. Molecular and Cellular Endocrinology. 2009;308(1–2):32-38.
11. Prossnitz ER, Barton M. Signaling, physiological functions and clinical relevance of the G protein-coupled estrogen receptor GPER. Prostaglandins Other Lipid Mediat. 2009;89(3–4):89-97.
12. Thomas P, Dong J. Binding and activation of the seven-transmembrane estrogen receptor GPR30 by environmental estrogens: A potential novel mechanism of endocrine disruption. The Journal of Steroid Biochemistry and Molecular Biology. 2006;102(1–5):175-179.
13. Brenner RM, Slayden OD. Steroid receptors in blood vessels of the rhesus macaque endometrium: a review. Arch. Histol. Cytol. 2004;67(5):411-416.
14. Jabbour HN, Kelly RW, Fraser HM, Critchley HO. Endocrine regulation of menstruation. Endocr. Rev. 2006;27(1):17-46.
63
15. Maggiolini M, Picard D. The unfolding stories of GPR30, a new membrane-bound estrogen receptor. J. Endocrinol. 2010;204(2):105-114.
16. Lodish H, Berk A, Matsudaira P, et al. Molecular cell biology. USA2004. 17. Hall JM, McDonnell DP. Coregulators in nuclear estrogen receptor action:
from concept to therapeutic targeting. Molecular interventions. 2005;5(6):343-357.
18. Nilsson BO, Olde B, Leeb-Lundberg LM. G protein-coupled oestrogen receptor 1 (GPER1)/GPR30: a new player in cardiovascular and metabolic oestrogenic signalling. Br. J. Pharmacol. 2011;163(6):1131-1139.
19. Plante BJ, Lessey BA, Taylor RN, et al. G Protein-Coupled Estrogen Receptor (GPER) Expression in Normal and Abnormal Endometrium. Reproductive Sciences. 2012;19(7):684-693.
20. Chris. This Wikipedia and Wikimedia Commons image is from the user Chris 73 and is freely available at //commons.wikimedia.org/wiki/File:MenstrualCycle.png under the creative commons cc-by-sa 3.0 license.
21. Giudice LC. Potential biochemical markers of uterine receptivity. Human Reproduction. 1999;14(suppl_2):3-16.
22. Carmeliet P, Jain RK. Molecular mechanisms and clinical applications of angiogenesis. Nature. 2011;473(7347):298-307.
23. Ferrara N. The biology of VEGF and its receptors. Nat. Med. 2003;9:669-676.
24. Losordo DW, Isner JM. Estrogen and Angiogenesis: A Review. Arterioscler Thromb Vasc Biol. 2001;21(1):6-12.
25. Demir R, Yaba A, Huppertz B. Vasculogenesis and angiogenesis in the endometrium during menstrual cycle and implantation. Acta Histochem. 2010;112(3):203-214.
26. Girling J, Rogers P. Recent advances in endometrial angiogenesis research. Angiogenesis. 2005;8(2):89-99.
27. Knowlton AA, Lee AR. Estrogen and the cardiovascular system. Pharmacol. Ther. 2012;135(1):54-70.
28. Albrecht ED, Babischkin JS, Lidor Y, Anderson LD, Udoff LC, Pepe GJ. Effect of estrogen on angiogenesis in cocultures of human endometrial cells and microvascular endothelial cells. Hum. Reprod. 2003;18(10):2039-2047.
30. Murdoch FE, Gorski J. The role of ligand in estrogen receptor regulation of gene expression. Mol. Cell. Endocrinol. 1991;78(3):C103-C108.
31. Critchley HOD, Brenner RM, Henderson TA, et al. Estrogen Receptor β, But Not Estrogen Receptor α, Is Present in the Vascular Endothelium of the Human and Nonhuman Primate Endometrium. J. Clin. Endocrinol. Metab. 2001;86(3):1370-1378.
32. Krikun G, Schatz F, Taylor R, et al. Endometrial Endothelial Cell Steroid Receptor Expression and Steroid Effects on Gene Expression. J. Clin. Endocrinol. Metab. 2005;90(3):1812-1818.
64
33. Iruela-Arispe ML, Rodriguez-Manzaneque JC, Abu-Jawdeh G. Endometrial Endothelial Cells Express Estrogen and Progesterone Receptors and Exhibit a Tissue Specific Response to Angiogenic Growth Factors. Microcirculation. 1999;6(2):127 - 140.
34. Chow RWY, Handelsman DJ, Ng MKC. Minireview: Rapid Actions of Sex Steroids in the Endothelium. Endocrinology. 2010;151(6):2411-2422.
35. Rodriguez-Manzaneque JC, Graubert M, Iruela-Arispe ML. Endothelial cell dysfunction following prolonged activation of progesterone receptor. Hum. Reprod. 2000;15 Suppl 3:39-47.
36. Kayisli UA, Luk J, Guzeloglu-Kayisli O, Seval Y, Demir R, Arici A. Regulation of Angiogenic Activity of Human Endometrial Endothelial Cells in Culture by Ovarian Steroids. J. Clin. Endocrinol. Metab. 2004;89(11):5794-5802.
37. Rogers PA, Lederman F, Kooy J, Taylor NH, Healy DL. Endometrial vascular smooth muscle oestrogen and progesterone receptor distribution in women with and without menorrhagia. Hum. Reprod. 1996;11(9):2003-2008.
38. Bergman Å, Heindel JJ, Jobling S, Kidd KA, Zoeller RT. WHO report: State of the science of Endocrine Disrupting Chemicals 20122012.
39. Mantovani A. Hazard identification and risk assessment of endocrine disrupting chemicals with regard to developmental effects. Toxicology. 2002;181-182:367-370.
40. Crain DA, Janssen SJ, Edwards TM, et al. Female reproductive disorders: the roles of endocrine-disrupting compounds and developmental timing. Fertility and Sterility. 2008;90(4):911-940.
41. McLachlan JA, Simpson E, Martin M. Endocrine disrupters and female reproductive health. Best Practice & Research Clinical Endocrinology & Metabolism. 2006;20(1):63-75.
42. Newbold RR. Lessons learned from perinatal exposure to diethylstilbestrol. Toxicol. Appl. Pharmacol. 2004;199(2):142-150.
43. Fowler PA, Bellingham M, Sinclair KD, et al. Impact of endocrine-disrupting compounds (EDCs) on female reproductive health. Molecular and Cellular Endocrinology. 2012;355(2):231-239.
44. De Coster S, van Larebeke N. Endocrine-disrupting chemicals: associated disorders and mechanisms of action. Journal of environmental and public health. 2012;2012:713696.
45. Damstra Terri , Barlow Sue , Bergman Aake , Kavlock Robert , Glen VDK. IPCS Global Assessment - of the State-of-theScience of Endocrine Disruptors. 2002.
46. Vandenberg LN, Colborn T, Hayes TB, et al. Hormones and endocrine-disrupting chemicals: low-dose effects and nonmonotonic dose responses. Endocr. Rev. 2012;33(3):378-455.
47. IARC 1993. Beryllium, Cadmium, Mercury and Exposures in the Glass Manufacturing Industry. IARCMonographs on the Evaluation of Carcinogenic Risk of Chemicals to Humans. Vol 58: France:International Agency for Research on Cancer. 444 pp
65
48. Thompson J, Bannigan J. Cadmium: Toxic effects on the reproductive system and the embryo. Reprod. Toxicol. 2008;25(3):304-315.
49. Järup L, Åkesson A. Current status of cadmium as an environmental health problem. Toxicol. Appl. Pharmacol. 2009;238(3):201-208.
50. Shiverick KT, Salafia C. Cigarette smoking and pregnancy I: ovarian, uterine and placental effects. Placenta. 1999;20(4):265-272.
51. Nordberg M. Environmental exposure and preventive measures in Sweden and EU*. Biometals. 2004;17(5):589-592.
52. Henson MC, Chedrese PJ. Endocrine disruption by cadmium, a common environmental toxicant with paradoxical effects on reproduction. Exp Biol Med (Maywood). 2004;229(5):383-392.
53. Iavicoli I, Fontana L, Bergamaschi A. The Effects of Metals as Endocrine Disruptors. J. Environ. Sci. Health. B. 2009;12(3):206-223.
54. Garcia-Morales P, Saceda M, Kenney N, et al. Effect of cadmium on estrogen receptor levels and estrogen-induced responses in human breast cancer cells. J Biol Chem. 1994;269(24):16896-16901.
55. Johnson MD, Kenney N, Stoica A, et al. Cadmium mimics the in vivo effects of estrogen in the uterus and mammary gland. Nat. Med. 2003;9(8):1081-1084.
56. Lienesch LA, Dumont JN, Bantle JA. The effect of cadmium on oogenesis in Xenopus laevis. Chemosphere. 2000;41(10):1651-1658.
57. Rehm S, Waalkes MP. Cadmium-induced ovarian toxicity in hamsters, mice, and rats. Fundam Appl Toxicol. 1988;10(4):635-647.
58. Piasek M, Laskey J, Kostial K, Blanuša M. Assessment of steroid disruption using cultures of whole ovary and/or placenta in rat and in human placental tissue. Int. Arch. Occup. Environ. Health. 2002;75(0):36-44.
59. Leoni G, Bogliolo L, Deiana G, et al. Influence of cadmium exposure on in vitro ovine gamete dysfunction. Reprod. Toxicol. 2002;16(4):371-377.
60. Yu HS, Tam PPL, Chan STH. Effects of cadmium on preimplantation mouse embryos in vitro with special reference to their implantation capacity and subsequent development. Teratology. 1985;32(3):347-353.
61. Nishijo M, Nakagawa H, Honda R, et al. Effects of maternal exposure to cadmium on pregnancy outcome and breast milk. Occup Environ Med. 2002;59(6):394-396; discussion 397.
62. Akesson A, Julin B, Wolk A. Long-term dietary cadmium intake and postmenopausal endometrial cancer incidence: a population-based prospective cohort study. Cancer Res. 2008;68(15):6435-6441.
63. Barbara KD, Leslie GF. From Adjuvant Therapy to Breast Cancer Prevention: BCPT and STAR. Breast J. 2001;7(3):144-157.
64. Jordan VC. Tamoxifen: a most unlikely pioneering medicine. Nat Rev Drug Discov. 2003;2(3):205-213.
65. Chalas E, Costantino JP, Wickerham DL, et al. Benign gynecologic conditions among participants in the Breast Cancer Prevention Trial. Am. J. Obstet. Gynecol. 2005;192(4):1230-1237.
66. Liao D, Johnson R. Hypoxia: A key regulator of angiogenesis in cancer. Cancer and Metastasis Rev. 2007;26(2):281-290.
66
67. Punyadeera C, Kamps R, Defrère S, et al. Effects of selective oestrogen receptor modulators on proliferation in tissue cultures of pre- and postmenopausal human endometrium. J. Steroid Biochem. Mol. Biol. 2008;112(1-3):102-109.
68. Mourits MJE, Hoor KAT, van der Zee AGJ, Willemse PHB, de Vries EGE, Hollema H. The effects of tamoxifen on proliferation and steroid receptor expression in postmenopausal endometrium. J. Clin. Pathol. 2002;55(7):514-519.
69. Hague S, Manek S, Oehler MK, MacKenzie IZ, Bicknell R, Rees MCP. Tamoxifen induction of angiogenic factor expression in endometrium. Br. J. Cancer. 2002;86(5):761-767.
70. Haran EF, Maretzek AF, Goldberg I, Horowitz A, Degani H. Tamoxifen Enhances Cell Death in Implanted MCF7 Breast Cancer by Inhibiting Endothelium Growth. Cancer Res. 1994;54(21):5511-5514.
71. McNamara DA, Harmey J, Wang JH, Kay E, Walsh TN, Bouchier-Hayes DJ. Tamoxifen inhibits endothelial cell proliferation and attenuates VEGF-mediated angiogenesis and migration in vivo. Eur. J. Surg. Oncol. 2001;27(8):714-718.
72. Spitz IM. Mifepristone: where do we come from and where are we going?: Clinical development over a quarter of a century. Contraception. 2009;82(5):442-452.
73. Wolf JP, Hsiu JG, Anderson TL, Ulmann A, Baulieu EE, GD H. Noncompetitive antiestrogenic effect of RU 486 in blocking the estrogen-stimulated luteinizing hormone surge and the proliferative action of estradiol on endometrium in castrate monkeys. Fertility and Sterility. 1989;52(6):1055-1060.
74. Baird DT, Brown A, Critchley HOD, Williams AR, Lin S, Cheng L. Effect of long-term treatment with low-dose mifepristone on the endometrium. Human Reproduction. 2003;18(1):61-68.
75. Johannisson E, Oberholzer M, Swahn M-L, Bygdeman M. Vascular changes in the human endometrium following the administration of the progesterone antagonist RU 486. Contraception. 1989;39(1):103-117.
76. Meng C-X, Andersson KL, Bentin-Ley U, Gemzell-Danielsson K, Lalitkumar PGL. Effect of levonorgestrel and mifepristone on endometrial receptivity markers in a three-dimensional human endometrial cell culture model. Fertility and Sterility. 2009;91(1):256-264.
77. Sitruk-Ware R. New progestagens for contraceptive use. Hum. Reprod. Update. 2006;12(2):169-178.
78. Mertens HJMM, Heineman MJ, Theunissen PHMH, de Jong FH, Evers JLH. Androgen, estrogen and progesterone receptor expression in the human uterus during the menstrual cycle. Eur. J. Obstet. Gynecol. Reprod. Biol. 2001;98(1):58-65.
79. Alonso-Magdalena P, Ropero AB, Soriano S, et al. Bisphenol-A acts as a potent estrogen via non-classical estrogen triggered pathways. Molecular and Cellular Endocrinology. 2012;355(2):201-207.
67
80. Vandenberg LN, Chahoud I, Heindel JJ, Padmanabhan V, Paumgartten FJ, Schoenfelder G. Urinary, circulating, and tissue biomonitoring studies indicate widespread exposure to bisphenol A. Environ. Health Perspect. 2010;118(8):1055-1070.
81. Welshons WV, Nagel SC, vom Saal FS. Large Effects from Small Exposures. III. Endocrine Mechanisms Mediating Effects of Bisphenol A at Levels of Human Exposure. Endocrinology. 2006;147(6):s56-s69.
82. Arnaoutova I, George J, Kleinman H, Benton G. The endothelial cell tube formation assay on basement membrane turns 20: state of the science and the art. Angiogenesis. 2009;12(3):267-274.
83. Arnaoutova I, Kleinman HK. In vitro angiogenesis: endothelial cell tube formation on gelled basement membrane extract. Nature Protocols. 2010;5(4):628-635.
84. Muller PY, Janovjak H, Miserez AR, Z. D. Processing of gene expression data generated by quantitative real-time RT-PCR. Biotechniques. 2002;32(6):1372-1374, 1376, 1378-1379.
85. Bustin SA, Benes V, Garson JA, et al. The MIQE Guidelines: Minimum Information for Publication of Quantitative Real-Time PCR Experiments. Clin. Chem. 2009;55(4):611 - 622.
86. Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):E45.
87. Khoo CP, Micklem K, Watt SM. A comparison of methods for quantifying angiogenesis in the Matrigel assay in vitro. Tissue engineering. Part C, Methods. 2011;17(9):895-906.
88. Garvin S, Nilsson UW, Dabrosin C. Effects of oestradiol and tamoxifen on VEGF, soluble VEGFR-1, and VEGFR-2 in breast cancer and endothelial cells. Br. J. Cancer. 2005;93(9):1005-1010.
89. Okada H, Tsutsumi A, Imai M, Nakajima T, Yasuda K, Kanzaki H. Estrogen and selective estrogen receptor modulators regulate vascular endothelial growth factor and soluble vascular endothelial growth factor receptor 1 in human endometrial stromal cells. Fertil. Steril. 2010;93(8):2680-2686.
90. Uhlén M, Björling E, Agaton C, et al. A Human Protein Atlas for Normal and Cancer Tissues Based on Antibody Proteomics. Proteomics. 2005;4(12):1920-1932.
91. Yuhki M, Kajitani T, Mizuno T, Aoki Y, Maruyama T. Establishment of an immortalized human endometrial stromal cell line with functional responses to ovarian stimuli. Reprod. Biol. Endocrinol. 2011;9(1):104.
92. Narvekar N, Critchley HOD, Cheng L, Baird DT. Mifepristone-induced amenorrhoea is associated with an increase in microvessel density and glucocorticoid receptor and a decrease in stromal vascular endothelial growth factor. Human Reproduction. 2006;21(9):2312-2318.
93. Duffy AM, Bouchier-Hayes DJ, JH. H. Vascular Endothelial Growth Factor (VEGF) and Its Role in Non-Endothelial Cells: Autocrine Signalling by VEGF. Madame Curie Bioscience Database [Internet]. Austin (TX): Landes Bioscience; 2000-. 2013.
68
94. Andersson H, Brittebo E. Proangiogenic effects of environmentally relevant levels of bisphenol A in human primary endothelial cells. Arch. Toxicol. 2012;86(3):465-474.
95. Ribatti D. The discovery of the placental growth factor and its role in angiogenesis: a historical review. Angiogenesis. 2008;11(3):215-221.
96. De Falco S. The discovery of placenta growth factor and its biological activity. Exp. Mol. Med. 2012;44:1-9.
97. Cao Y. Positive and Negative Modulation of Angiogenesis by VEGFR1 Ligands. Sci Signal. 2009;2(59):re1.
98. Moller B, Rasmussen C, Lindblom B, Olovsson M. Expression of the angiogenic growth factors VEGF, FGF-2, EGF and their receptors in normal human endometrium during the menstrual cycle. Mol. Hum. Reprod. 2001;7(1):65-72.
99. Gargett CE, Lederman FL, Lau TM, Taylor NH, Rogers PAW. Lack of correlation between vascular endothelial growth factor production and endothelial cell proliferation in the human endometrium. Human Reproduction. 1999;14(8):2080-2088.
100. Shibuya M. Differential roles of vascular endothelial growth factor receptor-1 and receptor-2 in angiogenesis. Journal of biochemistry and molecular biology. 2006;39(5):469-478.
101. Maynard SE, Karumanchi SA. Angiogenic Factors and Preeclampsia. Semin. Nephrol. 2011;31(1):33-46.
103. Mukundan H, Kanagy NL, Resta TC. 17-beta estradiol attenuates hypoxic induction of HIF-1alpha and erythropoietin in Hep3B cells. J. Cardiovasc. Pharmacol. 2004;44(1):93-100.
104. Desta Z, Ward BA, Soukhova NV, Flockhart DA. Comprehensive Evaluation of Tamoxifen Sequential Biotransformation by the Human Cytochrome P450 System in Vitro: Prominent Roles for CYP3A and CYP2D6. J. Pharmacol. Exp. Ther. 2004;310(3):1062-1075.
105. Kim SY, Laxmi YRS, Suzuki N, et al. Formation of tamoxifen-DNA adducts via O-sulfonation, not O-acetylation, of alpha-hydroxytamoxifen in rat and human livers. Drug Metab. Dispos. 2005;33(11):1673-1678.
106. Andersson H, Helmestam M, Zebrowska A, Olovsson M, Brittebo E. Tamoxifen-Induced Adduct Formation and Cell Stress in Human Endometrial Glands. Drug Metab. Dispos. 2010;38(1):200-207.
107. DuSell CD, Nelson ER, Wittmann BM, et al. Regulation of Aryl Hydrocarbon Receptor Function by Selective Estrogen Receptor Modulators. Mol. Endocrinol. 24:33-46. 2010;24(1):33-46.
108. Wang A, Ji L, Shang W, et al. Expression of GPR30, ERα and ERβ in endometrium during window of implantation in patients with polycystic ovary syndrome: a pilot study. Gynecol. Endocrinol. 2011;27(4):251-255.
69
109. Matthews J, Celius T, Halgren R, Zacharewski T. Differential estrogen receptor binding of estrogenic substances: a species comparison. The Journal of Steroid Biochemistry and Molecular Biology. 2000;74(4):223-234.
110. Gisterek I, Matkowski R, Lacko A, et al. Serum Vascular Endothelial Growth Factors A, C and D in Human Breast Tumors. Pathology & Oncology Research. 2010;16(3):337-344.
111. Currie MJ, Hanrahan V, Gunningham SP, et al. Expression of vascular endothelial growth factor D is associated with hypoxia inducible factor (HIF-1alpha) and the HIF-1alpha target gene DEC1, but not lymph node metastasis in primary human breast carcinomas. J. Clin. Pathol. 2004;57(8):829-834.
112. Rubin BS. Bisphenol A: an endocrine disruptor with widespread exposure and multiple effects. J. Steroid Biochem. Mol. Biol. 2011;127(1-2):27-34.
113. Vandenberg LN, Maffini MV, Sonnenschein C, Rubin BS, Soto AM. Bisphenol-A and the Great Divide: A Review of Controversies in the Field of Endocrine Disruption. Endocrine Reviews. 2009;30(1):75-95.
114. Andraweera PH, Dekker GA, Roberts CT. The vascular endothelial growth factor family in adverse pregnancy outcomes. Hum. Reprod. Update. 2012;18(4):436-457.
115. Groothuis PG, Nap AW, Winterhager E, Grümmer R. Vascular development in endometriosis. Angiogenesis. 2005;8(2):147-156.
116. Lockwood CJ. Mechanisms of normal and abnormal endometrial bleeding. Menopause. 2011;18(4):408-411.
117. Seo WS, Jee BC, Moon SY. Expression of endometrial protein markers in infertile women and the association with subsequent in vitro fertilization outcome. Fertility and Sterility. 2011;95(8):2707-2710.
118. Tayade C, Fang Y, Croy BA. A Review of Gene Expression in Porcine Endometrial Lymphocytes, Endothelium and Trophoblast During Pregnancy Success and Failure. Journal of Reproduction and Development. 2007;53(3):455-463.
119. Licht P, Fluhr H, Neuwinger J, Wallwiener D, Wildt L. Is human chorionic gonadotropin directly involved in the regulation of human implantation? Molecular and Cellular Endocrinology. 2007;269(1–2):85-92.
120. Hannan NJ, Paiva P, Meehan KL, Rombauts LJF, Gardner DK, Salamonsen LA. Analysis of Fertility-Related Soluble Mediators in Human Uterine Fluid Identifies VEGF as a Key Regulator of Embryo Implantation. Endocrinology. 2011;152(12):4948-4956.
121. Greb RR, Heikinheimo O, Williams RF, Hodgen GD, Goodman AL. Vascular endothelial growth factor in primate endometrium is regulated by oestrogen-receptor and progesterone-receptor ligands in vivo. Human Reproduction. 1997;12(6):1280-1292.
122. Guttinger A, Critchley HOD. Endometrial effects of intrauterine levonorgestrel. Contraception. 2007;75(6, Supplement):S93-S98.
123. Lockwood CJ, Schatz F, Krikun G. Angiogenic factors and the endometrium following long term progestin only contraception. Histol. Histopathol. 2004;19(1):167-172.
70
124. Moller B, Ronnerdag M, Wang G, Odlind V, Olovsson M. Expression of vascular endothelial growth factors and their receptors in human endometrium from women experiencing abnormal bleeding patterns after prolonged use of a levonorgestrel-releasing intrauterine system. Hum. Reprod. 2005;20(5):1410-1417.
Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 902
Editor: The Dean of the Faculty of Medicine
A doctoral dissertation from the Faculty of Medicine, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofMedicine.