Page 1
1
In vitro screening of the open source MMV malaria box reveals novel compounds 1
with profound activities against Theileria annulata schizonts 2
3
4
5
Isabel Hostettler, Joachim Müller, Andrew Hemphill1* 6
7
1Institute for Parasitology, Vetsuisse Faculty, University of Bern, Bern, Switzerland 8
9
* corresponding author: Institute of Parasitology 10
Vetsuisse Faculty 11
University of Bern 12
Länggass-Strasse 122 13
CH-3012 Bern, Switzerland 14
Tel +41 31 6312474 15
Fax +41 31 6312477 16
Email: [email protected] 17
18
19
AAC Accepted Manuscript Posted Online 14 March 2016Antimicrob. Agents Chemother. doi:10.1128/AAC.02801-15Copyright © 2016, American Society for Microbiology. All Rights Reserved.
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 2
2
Abstract 20
21
Intracellular schizonts of the apicomplexans Theileria annulata and Theileria parva 22
immortalize bovine leukocytes and thereby cause fatal diseases. The 23
hydroxynaphthoquinone buparvaquone is currently the only option for the treatment 24
of theileriosis, and resistance formation has been reported. It is therefore tempting to 25
investigate the repurposing of compounds effective against related apicomplexan 26
parasites such as Plasmodium. Here, we present the results of a screen of 400 27
compounds included in the open access Medicines for Malaria Venture (MMV) 28
malaria box on TaC12 cells, a macrophage-derived cell line immortalized by T. 29
annulata schizonts. Using a combination of the classical Alamar blue vitality assay 30
and a recently developed quantitative reverse transcriptase real time PCR method 31
based on the Theileria gene TaSP, we have identified 5 compounds, characterized 32
their effects on the ultrastructure of TaC12 cells, and investigated whether they easily 33
induce resistance formation. Two compounds, the quinolinols MMV666022 and 34
MMV666054, have IC50 values of 0.5 and 0.2 µM on TaC12 cells and 5.3 and 5.2 µM 35
on BoMac cells respectively. Thus, with therapeutic indexes of 11 and 18, they 36
represent promising leads for further development of anti-theilerial 37
chemotherapeutics. 38
39
Key words 40
41
Cellular transformation - drug development – immunoproliferative diseases – 42
Theileria annulata – MMV malaria box - quinolinoles43
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 3
3
1. Introduction 44
45
Apicomplexan parasites such as Cryptosporidium, Eimeria, Plasmodium, Toxoplasma 46
and others are of outstanding veterinary and/or human medical importance. Theileria 47
sporozoites are transmitted by ticks, infect cattle and cause acute and fatal leuko-48
proliferative disease. T. parva is found in East, Central and Southern Africa and 49
causes East Coast fever, while T. annulata, occurs in the Mediterranean and middle 50
East area, Northern Africa, India, and the far East, and is the causative agent of 51
tropical theileriosis (1). Theileria-infected cells share several hallmarks of cancer cells 52
such as resistance to apoptosis, uncontrolled and unlimited proliferation, deregulation 53
of cellular energetics and acquisition of an invasive and metastatic phenotype (2). 54
Currently, there are different strategies for the prevention and treatment of bovine 55
theileriosis, namely (i) targeting the tick vector by acaricide treatment (3) or (ii) by 56
targeting the parasite itself, either by vaccination, chemotherapy, or a combination of 57
both (4, 5). The most efficacious vaccine approach for East Coast fever was 58
developed many years ago, and includes infection of cattle with T. parva sporozoites 59
and immediate treatment with a drug that impairs the establishment of infection such 60
as oxytetracycline (6) , or alternatively buparvaquone (BPQ), which is currently the 61
only effective compound for the treatment of bovine theileriosis (7-9). BPQ is a 62
hydroxynaphtoquinone related to parvaquone (10, 11). To be effective, the compound 63
must be administrated during the early stage of infection, and the drug is not always 64
affordable or available for farmers (12). More recently, first cases of treatment 65
failures were reported due to resistant parasite populations (13). These treatment 66
failures were shown to be associated with mutations in the Theileria cytochrome b 67
gene encoding for the ubiquinone reductase of the respiratory chain (14, 15). Another 68
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 4
4
potential target of BPQ is Theileria prolyl isomerase (TaPIN1) since mutations in the 69
respective gene were demonstrated in BPQ resistant cell lines (16). Using a novel 70
assay based on the differential analysis of transcribed RNA from host and parasite, we 71
have shown that BPQ affects the parasite within few hours thus indicating that – in all 72
likelihood - BPQ acts via a rapid metabolic inhibition (17). 73
The occurrence of BPQ resistance in cattle provides an incentive to search for 74
alternative treatment options. The open access Medicines for Malaria Venture (MMV) 75
malaria box includes 200 drug-like and 200 probe-like compounds, which represent a 76
subset of the 20,000 in vitro anti-malarials identified from the high-throughput 77
screening efforts of St. Jude Children's Research Hospital (TN, USA), Novartis and 78
GlaxoSmithKline (18-20). MMV box compounds have proven to be effective in vitro 79
against the apicomplexans Plasmodium falciparum (blood stage 3D7 and K1 strains) 80
(21), Toxoplasma gondii (22), and Cryptosporidum parvum (23). Apicomplexans 81
share a number of conserved pathways representing potentially druggable targets (24-82
26). For instance, the crossreactivity of inhibitors of calcium-dependent protein 83
kinases (CDPKs) against Plasmodium (27), Toxoplasma (28) and Neospora (29, 30), 84
and the demonstration of profound in vitro and in vivo activity of BPQ against N. 85
caninum infection (31), have underscored a potential role for exploiting the possibility 86
of repurposing drugs among apicomplexans. 87
We here report on the screening of the MMV malaria box against a macrophage cell 88
line infected with the schizont stage of T. annulata, and on the characterization of 5 89
compounds that could potentially serve as a starting point for the development of 90
novel anti-theilerial drugs. 91
92
2. Materials and methods 93
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 5
5
94
2.1. Cell cultures 95
96
TaC12, a bovine macrophage cell line infected with T. annulata, was cultured as 97
previously described (17). BoMac, a bovine macrophage cell line immortalized by the 98
SV40 large T-antigen was cultured as described (32). Human foreskin fibroblasts 99
(HFF) were cultured as described (31). 100
101
2.2. MMV malaria box compounds 102
103
The MMV malaria box was obtained from the Medicines for Malaria Venture (MMV; 104
Geneva, Switzerland). Plate mapping and full data on the Malaria Box were made 105
available under http://www.mmv.org/research-development/malaria-box-supporting-106
information). For more detailed studies, the compounds MMV000760 (1), 107
MMV666022 (2), MMV666023 (3) and MMV666054 (4) were purchased from 108
ChemBridge, and MMV665941 (5) was obtained from AK Scientific, Inc. All 109
compounds were kept as 10 mM stocks in DMSO and diluted in respective culture 110
media for the individual experiments as indicated. 111
112
2.3. Cytotoxicity assays 113
114
For the initial screening of the MMV Malaria box, TaC12 cells (5x103 in 200 µl per 115
well) were seeded into flat-bottomed 96-well plates (Greiner Cellstar, Kremsmünster, 116
Austria) and were allowed to adhere for 2h prior to the addition of 1μM of the malaria 117
box compounds. Some wells received BPQ (0.15 µM; Cross Vet Pharm, Dublin, 118
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 6
6
Ireland) or 0.1% DMSO as positive and solvent controls, respectively. For IC50 119
determinations, cells were incubated in the presence of 0 to 2 μM. 120
BoMac cells (2x103 cells in 200 µl medium per well) were seeded into 96-well plates, 121
and were allowed to adhere for 2h prior to the addition of 0 to 20 μM of the respective 122
compounds. 123
HFF (2x103 cells in 200 µl medium per well) were seeded into 96-well plates, and 124
were allowed to grow to confluent monolayers. Then, the monolayers were exposed to 125
1 μM of selected malaria box compounds. 126
All cells were cultured for 3 days, and cell viability was measured using the Alamar 127
blue assay as described (31). 128
129
2.4. Quantitative reverse transcriptase real time PCR (qRT-PCR) 130
131
Quantitative RT-PCR assays using primers for bovine actin and for TaSP were 132
performed as described (17). Briefly, RNA isolated with the Qiagen (Hilden, 133
Germany) RNeasy kit including DNAseI digestion, and cDNA was synthesized with 2 134
µg of RNA using the Qiagen Omniscript™ kit with random primers according to the 135
manufacturer’s instruction. Quantitative PCR was performed with 10 µL of cDNA 136
(diluted 1:50 in water) using the Quanti TectTM SYBR Green PCR Kit (Roche, 137
Basel, Switzerland) in 20 µL standard reactions containing 0.5 µM of forward and 138
reverse primers (MWG Biotech, Ebersberg, Germany). Real-time PCR was performed 139
using a Corbett cycler (Mortlake, Australia) and expression levels were given as 140
values in arbitrary units relative to the amount of actin RNA as described (17). 141
142
2.5. Transmission electron microscopy (TEM) 143
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 7
7
144
For TEM, 106 TaC12 cells were seeded into T25-tissue- flasks and were cultured for 145
24h. Then, cultures were exposed to 0.5 or 1μM of malaria box compounds, 0.15 µM 146
BPQ, or corresponding amounts of DMSO for 48h. Fixation, postfixation and 147
embedding were carried out as previously described (17). 148
149
2.6. Long term culture of TaC12 cells in the presence of BPQ and selected Malaria 150
box compounds 151
152
For long term cultures, 106 TaC12 cells were seeded into 25cm2 culture flasks, 153
compounds were added after 2h of recovery as indicated. The medium was changed at 154
three-day-intervals. The culture was continued for a maximum of 14 days or until no 155
surviving cells were visible. Then, the cultures were switched to normal medium, and 156
potential regrowth of the parasites was analyzed after one week. 157
158
2.7. Statistics 159
160
Concerning the evaluation of the screen of TaC12 cells, compounds were regarded as 161
potential hits when the Alamar blue values were not higher than the mean values of 162
the positive controls (i.e. BPQ) plus 3 times their standard deviation. Concerning the 163
evaluation of the screen of HFF cells, compounds were regarded as cytotoxic when 164
the Alamar blue values were not higher than the mean values of the negative controls 165
(i.e. DMSO) minus 2 times their standard deviation. IC50 values were calculated as 166
described (33). 167
168
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 8
8
3. Results 169
170
3.1. Identification and first characterization of five lead compounds 171
172
In a primary assay, TaC12 cells were exposed to 1 µM of each malaria box 173
compound, and to 150 nM BPQ as a reference compound, for a period of 3 days and 174
subjected to the Alamar blue viability assay. As expected, BPQ was highly active 175
against TaC12 cells. In parallel, to eliminate compounds that were highly cytotoxic 176
against mammalian cells, the same screen was performed using confluent HFF. 11 177
compounds impaired the viability of TaC12 cells to a similar extent as BPQ without 178
impairing the viability of HFF. Subsequently, IC50 values of these 11 compounds 179
were determined. Furthermore, these compounds were subjected to our previously 180
established assay based on the differential expression of host actin and parasite TaSP 181
(17) in order to eliminate compounds that preferentially affected the host 182
compartment of infected cells. Concerning the RT-PCR assay, in control cells, the 183
TaSP expression levels were in the same order of magnitude as the Act levels. The 184
mean values of the relative expression levels were therefore set as 100 % in the 185
control cells. Compounds causing a drop of these relative TaSP levels to 50% or less 186
of the control levels were regarded as potentially interesting for further studies. Both 187
assays led to the elimination of six compounds (Table 1). Five compounds, henceforth 188
referred to as compounds 1 to 5, inhibited TaC12 at submicromolar concentrations 189
affecting parasite rather than host gene expression. These compounds were retained 190
for subsequent studies. Using the TaSP gene expression assay as described above, we 191
found a clear concentration dependency of relative TaSP mRNA levels with all five 192
compounds, 3 and 5 having the lowest TaSP levels at 0.5 µM (Fig. 1). 193
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 9
9
In order to determine a potential “therapeutic window”, BoMac cells were incubated 194
with concentration series of these five compounds for three days and subjected to 195
Alamar blue viability assays. Compound 5 had the highest toxicity for BoMac cells, 196
compounds 2 and 4 the lowest and therefore the best therapeutic indexes (34), namely 197
11 and 18, of the five compounds (Table 1). 198
199
3.2. Electron microscopy of TaC12 cultures treated with compounds 1 to 5 200
201
TaC12 cells cultured in the absence of any drugs are shown in Fig. 2A and B. T. 202
annulata schizonts were readily detected in the individual cells. They are located 203
freely in the cytoplasm, delineated from the host cell cytoplasm only by the schizont 204
plasma membrane. Typically 2-6 parasite nuclei were visible per section plane. In 205
many instances, electron dense button-like structures, seemingly acting as membrane 206
connectors and structurally exhibiting some resemblance to tight junctions, were 207
found to be abundantly associated with these schizonts. Host cell mitochondria with 208
normal oval-shaped morphology were found near the schizonts. BPQ-treatment of 209
TaC12 cells induced clear alterations within the schizont cytoplasm, while the host 210
cell appeared largely unaffected (Fig. 2C and D). Alterations included extensive 211
accumulation of vacuoles filled with electron dense material of unknown nature, and a 212
general disintegration of the cytoplasmic organization of schizonts. Nuclei were not 213
discernible anymore, and button-like structures had disappeared or only residues were 214
still visible. However, the parasite plasma membrane remained morphologically intact 215
and clearly separated the parasite and host cell cytoplasm. 216
In TaC12 cells treated with compound 1, host cell mitochondria lost their typical oval 217
shape and appeared rather swollen, and in some instances cytoplasmic vacuoles were 218
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 10
10
observed (Fig. 3A). However, the alterations within the schizonts appeared minimal 219
with only few small vacuoles and overall intact structural features. 220
Treatment with compound 5 also led to intra-schizont vacuole formation, but the 221
overall cytoplasmic organization appeared still more intact compared to BPQ treated 222
cells, and nuclei were still clearly discernible. The button-like structures associated 223
with the schizonts appeared normal. On the host cell side, mitochondria morphology 224
was clearly impaired, the organelles appeared less compact and less electron-dense 225
compared to mitochondria in control cells, indicative for effects on the host cell 226
metabolism (Fig. 3A). Schizonts located in compound 2 (Fig. 3B and C) and 3 treated 227
TaC12 cells (Fig. 3D and E) also exhibited extensive cytoplasmic vacuolization, 228
indicating that schizonts were metabolically impaired. In addition, in both instances 229
the button-like structures that were evident in untreated TaC12 cells were not visible 230
anymore, or only residues could be identified. Vacuolization was also observed within 231
the host cell cytoplasm in compound 2 treated cells, but this was less evident in cells 232
treated with 3. Most notably, extensive mitochondrial swelling was evident in TaC12 233
cells treated with compound 3 (Fig. 3) 234
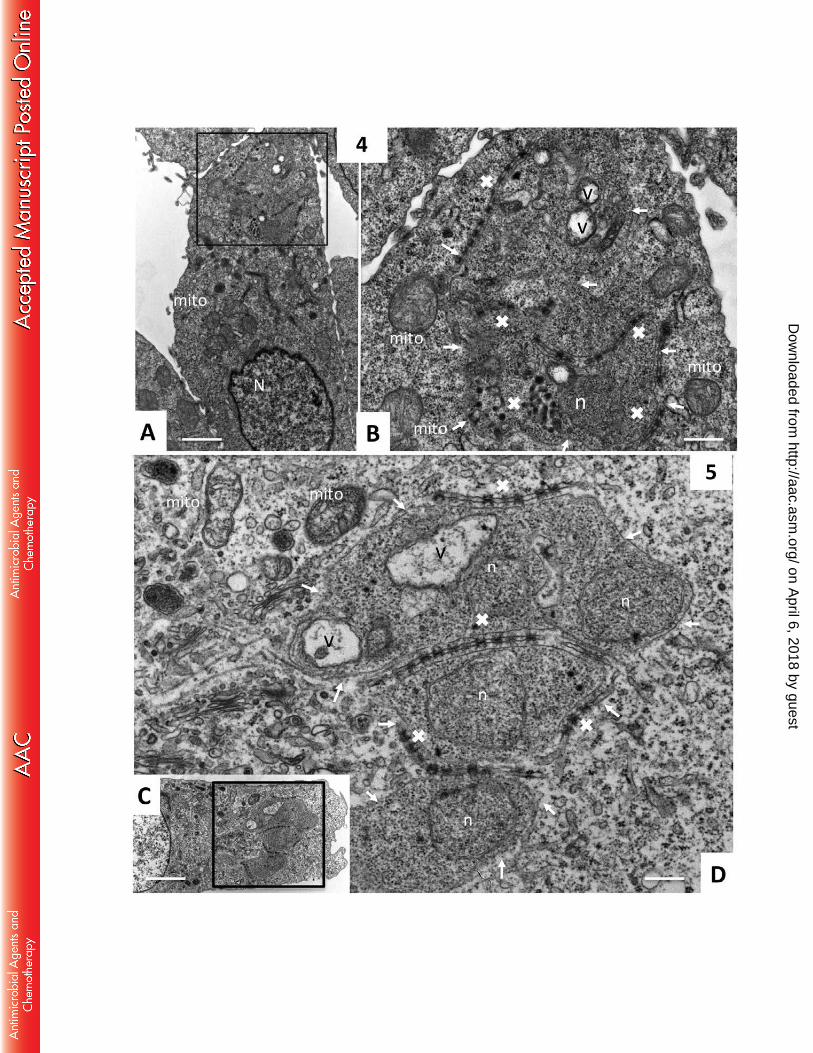
Schizonts located in compound 4 and 5 treated TaC12 cells (Fig. 5) also exhibited 235
extensive cytoplasmic vacuolization. However, the button-shaped structures in those 236
cells were not affected and appeared rather intact. On the other hand, compound 4 237
(Fig. 4 A, B) induced extensive swelling of host cell mitochondria, similar to what 238
was found for compound 1 and 3, and treatment with compound 5 (Fig. 4 C and D) 239
resulted in mitochondria which were not swollen, but rather appeared disintegrated 240
and less electron dense. 241
242
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 11
11
3.3. Long-term treatment of TaC12 cells with BPQ and compounds (1) to (5) reveals 243
the capacity of Theileria annulata to adapt and/or develop resistance to BPQ, but not 244
malaria box compounds 245
246
In order to determine whether TaC12 cells easily adapt to compounds 1 to 5, cells 247
were treated with the concentration that showed maximum effect on TaSP in RT-PCR 248
assay, namely 0.5 µM for compounds 3 and 5 and 1 µM for the others. Moreover, a 249
control with cells treated with 150 nM BPQ was included. We observed an initial 250
dramatic reduction in the number of viable cells within the first 3-5 days of BPQ 251
treatment. However, upon extended culture in the presence of BPQ, small colonies of 252
surviving TaC12 cells have formed, which remained viable, and resumed proliferation 253
at concentrations up to 2 µM (data not shown). On the other hand, when TaC12 cells 254
were exposed to the five malaria box compounds, no viable cells were visible after 14 255
days of culture in medium containing the compounds followed by one week of culture 256
in drug-free medium, indicating that no adaptation to these compounds took place. 257
258
4. Discussion 259
260
Performing a screen with the MMV malaria box, we have identified five compounds 261
inhibiting TaC12 cell proliferation preferentially harming the T. annulata schizont. 262
All five compounds are highly lipophilic (AlogP values above 4). By comparing the 263
IC50 values on TaC12 to IC50 values on BoMac cells, we see that compounds 2 and 4 264
offer convenient therapeutic indexes. One has to keep in mind, however, that both 265
TaC12 and BoMac cells are immortalized, TaC12 by T. annulata, BoMac by the 266
SV40 large T oncogene (32). It is therefore likely, that untransformed blood cells 267
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 12
12
react differently to these compounds, and could potentially be more resistant. 268
Compounds 2 and 4 are highly similar differing by one chlorine group on the 269
quinoline-8-ol scaffold only. Other compounds with this scaffold, namely 9 and 11, 270
also inhibit the proliferation of TaC12 cells, but preferentially affect the host cell. 271
This indicates that anti-theilerial drug design is a tightrope walk between preferential 272
host cell and parasite toxicity. As indicated by their therapeutic indexes, compounds 2 273
and 4 are convenient leads for the design of novel drugs as a backup for 274
buparvaquone. Interestingly, both compounds also exhibited profound activity against 275
Cryptosporidium parvum in vitro (23). None of these 5 compounds was, however, 276
active against T. gondii, another intracellular apicomplexan (22). Compound 5 had not 277
only the lowest IC50 on TaC12 cells, but also the highest toxicity for BoMac cells. 278
This compound is the hydrated precursor of crystal violet, a broad range anti-infective 279
agent used as antimycotic. It is therefore not surprising that it had also been identified 280
as one of the hits against Cryptosporidium. 281
Morphologically, all compounds identified here, except compound 1, induced 282
vacuolization of the schizont cytoplasm, and to different degrees the treatments 283
resulted in morphological alterations that indicated metabolic impairment of the 284
parasite. However, in all cases the schizont cytoplasm appeared still clearly 285
discernible, which indicates that these drugs induce parasite death in a manner that 286
resembles cellular apoptosis rather than necrosis. As shown in a previous study, 287
schizont lobes are connected by button-like structures (17). These structures resemble 288
tight junctions, but their molecular composition and function are unknown. They 289
could be involved in maintaining the structural integrity of the schizont stage, which 290
resides free in the cytoplasm of its host cell. Compounds 2 and 4 affected the integrity 291
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 13
13
of those buttons whereas treatments with the other compounds did not alter these 292
structures. 293
The molecular targets of all five compounds are unknown. In a screen for 294
aminopeptidase inhibitors, compound 3 has been identified as one of the hits (35). We 295
have not detected any inhibitory effect of this compound on aminopeptidase activities 296
in crude extracts neither from TaC12 cells nor from isolated T. annulata schizonts 297
(data not shown). The high lipophilicity of all compounds could be not only 298
instrumental for the multiple membrane passages that are required to reach the 299
schizont, but could also be indicative for membrane-bound targets. Moreover, these 300
compounds could interact with multiple targets in both the parasite and the host cell, 301
since TaC12 cells were not able to adapt to these compounds during long term 302
cultures. Affinity chromatography (see e.g. (33) and references therein) would be a 303
tool to identify those proteins that interact with these compounds, especially 2 and 4, 304
two quinolinols, which are closely related and represent the most interesting leads. 305
Both compounds are probe-like compounds and will certainly been modified prior to 306
in vivo applications in a suitable model. Therefore, there are no preclinical ADME 307
data available. Other quinolinols are effective against intracellular tachyzoites, most 308
likely though a mechanism involving generation of reactive oxygen species (36). This 309
could explain the effects on mitochondrial integrity as observed in our study. 310
311
312
Acknowledgements 313
The authors want to thank Kerry Woods and Sven Rottenberg for crucial logistic and 314
moral support and providing lab space for Theileria cell culture. We also gratefully 315
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 14
14
acknowledge Medicines for Malaria Venture in Geneva for providing the malaria box, 316
and especially Wes Van Voorhis for helpful advice and support. 317
318
Funding 319
This work was supported by the Swiss National Science Foundation (grant No. 320
310030_146162, and the Vetsuisse Faculty of the University of Bern. 321
322
Competing interests 323
None declared 324
325
Ethical approval 326
None required 327
328
References 329
1. Mans BJ, Pienaar R, Latif AA. 2015. A review of Theileria diagnostics and 330 epidemiology. Int J Parasitol Parasites Wildl 4:104-118. 331
2. Tretina K, Gotia HT, Mann DJ, Silva JC. 2015. Theileria-transformed 332 bovine leukocytes have cancer hallmarks. Trends Parasitol 31:306-314. 333
3. Gachohi J, Skilton R, Hansen F, Ngumi P, Kitala P. 2012. Epidemiology of 334 East Coast fever (Theileria parva infection) in Kenya: past, present and the 335 future. Parasit Vectors 5:194. 336
4. McKeever DJ. 2007. Live immunisation against Theileria parva: containing 337 or spreading the disease? Trends Parasitol 23:565-568. 338
5. McKeever DJ. 2009. Bovine immunity - a driver for diversity in Theileria 339 parasites? Trends Parasitol 25:269-276. 340
6. Spooner PR. 1990. The effects of oxytetracycline on Theileria parva in vitro. 341 Parasitology 100 Pt 1:11-17. 342
7. Brown CG. 1990. Control of tropical theileriosis (Theileria annulata 343 infection) of cattle. Parasitologia 32:23-31. 344
8. Mutugi JJ, Young AS, Maritim AC, Linyonyi A, Mbogo SK, Leitch BL. 345 1988. Immunization of cattle using varying infective doses of Theileria parva 346 lawrencei sporozoites derived from an African buffalo (Syncerus caffer) and 347 treatment with buparvaquone. Parasitology 96 ( Pt 2):391-402. 348
9. McHardy N, Morgan DW. 1985. Treatment of Theileria annulata infection 349 in calves with parvaquone. Res Vet Sci 39:1-4. 350
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 15
15
10. McHardy N, Hudson AT, Morgan DW, Rae DG, Dolan TT. 1983. Activity 351 of 10 naphthoquinones, including parvaquone (993C) and menoctone, in cattle 352 artificially infected with Theileria parva. Res Vet Sci 35:347-352. 353
11. Boehm P, Cooper K, Hudson AT, Elphick JP, McHardy N. 1981. In vitro 354 activity of 2-alkyl-3-hydroxy-1,4-naphthoquinones against Theileria parva. J 355 Med Chem 24:295-299. 356
12. Morrison WI, McKeever DJ. 2006. Current status of vaccine development 357 against Theileria parasites. Parasitology 133 Suppl:S169-187. 358
13. Mhadhbi M, Naouach A, Boumiza A, Chaabani MF, BenAbderazzak S, 359 Darghouth MA. 2010. In vivo evidence for the resistance of Theileria 360 annulata to buparvaquone. Vet Parasitol 169:241-247. 361
14. Sharifiyazdi H, Namazi F, Oryan A, Shahriari R, Razavi M. 2012. Point 362 mutations in the Theileria annulata cytochrome b gene is associated with 363 buparvaquone treatment failure. Vet Parasitol 187:431-435. 364
15. Mhadhbi M, Chaouch M, Ajroud K, Darghouth MA, BenAbderrazak S. 365 2015. Sequence Polymorphism of Cytochrome b Gene in Theileria annulata 366 Tunisian Isolates and Its Association with Buparvaquone Treatment Failure. 367 PLoS One 10:e0129678. 368
16. Marsolier J, Perichon M, DeBarry JD, Villoutreix BO, Chluba J, Lopez 369 T, Garrido C, Zhou XZ, Lu KP, Fritsch L, Ait-Si-Ali S, Mhadhbi M, 370 Medjkane S, Weitzman JB. 2015. Theileria parasites secrete a prolyl 371 isomerase to maintain host leukocyte transformation. Nature 520:378-382. 372
17. Hostettler I, Müller J, Stephens CE, Haynes R, Hemphill A. 2014. A 373 quantitative reverse-transcriptase PCR assay for the assessment of drug 374 activities against intracellular Theileria annulata schizonts. . Int J Parasitol 375 Drugs Drug Resist 4:201-209. 376
18. Gamo FJ, Sanz LM, Vidal J, de Cozar C, Alvarez E, Lavandera JL, 377 Vanderwall DE, Green DV, Kumar V, Hasan S, Brown JR, Peishoff CE, 378 Cardon LR, Garcia-Bustos JF. 2010. Thousands of chemical starting points 379 for antimalarial lead identification. Nature 465:305-310. 380
19. Meister S, Plouffe DM, Kuhen KL, Bonamy GM, Wu T, Barnes SW, 381 Bopp SE, Borboa R, Bright AT, Che J, Cohen S, Dharia NV, Gagaring K, 382 Gettayacamin M, Gordon P, Groessl T, Kato N, Lee MC, McNamara 383 CW, Fidock DA, Nagle A, Nam TG, Richmond W, Roland J, Rottmann 384 M, Zhou B, Froissard P, Glynne RJ, Mazier D, Sattabongkot J, Schultz 385 PG, Tuntland T, Walker JR, Zhou Y, Chatterjee A, Diagana TT, 386 Winzeler EA. 2011. Imaging of Plasmodium liver stages to drive next-387 generation antimalarial drug discovery. Science 334:1372-1377. 388
20. Guiguemde WA, Shelat AA, Bouck D, Duffy S, Crowther GJ, Davis PH, 389 Smithson DC, Connelly M, Clark J, Zhu F, Jimenez-Diaz MB, Martinez 390 MS, Wilson EB, Tripathi AK, Gut J, Sharlow ER, Bathurst I, El Mazouni 391 F, Fowble JW, Forquer I, McGinley PL, Castro S, Angulo-Barturen I, 392 Ferrer S, Rosenthal PJ, Derisi JL, Sullivan DJ, Lazo JS, Roos DS, Riscoe 393 MK, Phillips MA, Rathod PK, Van Voorhis WC, Avery VM, Guy RK. 394 2010. Chemical genetics of Plasmodium falciparum. Nature 465:311-315. 395
21. Spangenberg T, Burrows JN, Kowalczyk P, McDonald S, Wells TN, 396 Willis P. 2013. The open access malaria box: a drug discovery catalyst for 397 neglected diseases. PLoS One 8:e62906. 398
22. Boyom FF, Fokou PV, Tchokouaha LR, Spangenberg T, Mfopa AN, 399 Kouipou RM, Mbouna CJ, Donfack VF, Zollo PH. 2014. Repurposing the 400
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 16
16
open access malaria box to discover potent inhibitors of Toxoplasma gondii 401 and Entamoeba histolytica. Antimicrob Agents Chemother 58:5848-5854. 402
23. Bessoff K, Spangenberg T, Foderaro JE, Jumani RS, Ward GE, Huston 403 CD. 2014. Identification of Cryptosporidium parvum active chemical series by 404 Repurposing the open access malaria box. Antimicrob Agents Chemother 405 58:2731-2739. 406
24. Abrahamsen MS, Templeton TJ, Enomoto S, Abrahante JE, Zhu G, 407 Lancto CA, Deng M, Liu C, Widmer G, Tzipori S, Buck GA, Xu P, 408 Bankier AT, Dear PH, Konfortov BA, Spriggs HF, Iyer L, Anantharaman 409 V, Aravind L, Kapur V. 2004. Complete genome sequence of the 410 apicomplexan, Cryptosporidium parvum. Science 304:441-445. 411
25. Xu P, Widmer G, Wang Y, Ozaki LS, Alves JM, Serrano MG, Puiu D, 412 Manque P, Akiyoshi D, Mackey AJ, Pearson WR, Dear PH, Bankier AT, 413 Peterson DL, Abrahamsen MS, Kapur V, Tzipori S, Buck GA. 2004. The 414 genome of Cryptosporidium hominis. Nature 431:1107-1112. 415
26. Wernimont AK, Artz JD, Finerty P, Jr., Lin YH, Amani M, Allali-416 Hassani A, Senisterra G, Vedadi M, Tempel W, Mackenzie F, Chau I, 417 Lourido S, Sibley LD, Hui R. 2010. Structures of apicomplexan calcium-418 dependent protein kinases reveal mechanism of activation by calcium. Nat 419 Struct Mol Biol 17:596-601. 420
27. Ojo KK, Pfander C, Mueller NR, Burstroem C, Larson ET, Bryan CM, 421 Fox AM, Reid MC, Johnson SM, Murphy RC, Kennedy M, Mann H, 422 Leibly DJ, Hewitt SN, Verlinde CL, Kappe S, Merritt EA, Maly DJ, 423 Billker O, Van Voorhis WC. 2012. Transmission of malaria to mosquitoes 424 blocked by bumped kinase inhibitors. J Clin Invest 122:2301-2305. 425
28. Ojo KK, Larson ET, Keyloun KR, Castaneda LJ, Derocher AE, 426 Inampudi KK, Kim JE, Arakaki TL, Murphy RC, Zhang L, Napuli AJ, 427 Maly DJ, Verlinde CL, Buckner FS, Parsons M, Hol WG, Merritt EA, 428 Van Voorhis WC. 2010. Toxoplasma gondii calcium-dependent protein 429 kinase 1 is a target for selective kinase inhibitors. Nat Struct Mol Biol 17:602-430 607. 431
29. Ojo KK, Reid MC, Kallur Siddaramaiah L, Müller J, Winzer P, Zhang Z, 432 Keyloun KR, Vidadala RS, Merritt EA, Hol WG, Maly DJ, Fan E, Van 433 Voorhis WC, Hemphill A. 2014. Neospora caninum calcium-dependent 434 protein kinase 1 is an effective drug target for neosporosis therapy. PLoS One 435 9:e92929. 436
30. Winzer P, Müller J, Aguado-Martínez A, Rahman M, Balmer V, Ortega-437 Mora L, Ojo KK, Fan E, Maly D, Van Voorhis WC, Hemphill A. 2015. In 438 vitro and in vivo effects of the bumped kinase inhibitor 1294 in the related 439 cyst-forming apicomplexans Toxoplasma gondii and Neospora caninum. 440 Antimicrob Agents Chemother 59:6361-6374. 441
31. Müller J, Aguado-Martinez A, Manser V, Balmer V, Winzer P, Ritler D, 442 Hostettler I, Solís D, Ortega-Mora LM, Hemphill A. 2015. Buparvaquone 443 is active against Neospora caninum in vitro and in experimentally infected 444 mice. Int J Parasitol: Drugs Drug Res 5:16-25. 445
32. Stabel JR, Stabel TJ. 1995. Immortalization and characterization of bovine 446 peritoneal macrophages transfected with SV40 plasmid DNA. Vet Immunol 447 Immunopathol 45:211-220. 448
33. Müller J, Hemphill A. 2013. New approaches for the identification of drug 449 targets in protozoan parasites. Int Rev Cell Mol Biol 301:359-401. 450
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 17
17
34. Muller PY, Milton MN. 2012. The determination and interpretation of the 451 therapeutic index in drug development. Nat Rev Drug Discov 11:751-761. 452
35. Paiardini A, Bamert RS, Kannan-Sivaraman K, Drinkwater N, Mistry 453 SN, Scammells PJ, McGowan S. 2015. Screening the Medicines for Malaria 454 Venture "Malaria Box" against the Plasmodium falciparum aminopeptidases, 455 M1, M17 and M18. PLoS One 10:e0115859. 456
36. Strobl JS, Seibert CW, Li Y, Nagarkatti R, Mitchell SM, Rosypal AC, 457 Rathore D, Lindsay DS. 2009. Inhibition of Toxoplasma gondii and 458 Plasmodium falciparum infections in vitro by NSC3852, a redox active 459 antiproliferative and tumor cell differentiation agent. J Parasitol 95:215-223. 460
461 462
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 18
18
TABLES 463
Table 1. Inhibition of TaC12 viability by malaria box compounds. 11 compounds identified in a previous screening were added to TaC12 464
cells previously seeded in 96-well-plates in concentration series ranging from 0 to 2 µM. DMSO (0.1 %) was included as a solvent control, BPQ 465
as a positive control. After three days, viability was assayed using the Alamar blue assay. In a second step, in order to see whether the 466
compounds induced a decrease of T. annulata mRNA levels 107 cells were seeded in the presence of 1 µM of the compounds, BPQ (150 nM) as 467
a positive control, or DMSO as a negative control. After 24 h, cells were harvested, RNA was extracted, and mRNA levels of TaSP were 468
quantified by real-time RT-PCR to host-cell actin. Mean values ± SE are expressed as percentages of the DMSO control and are given for 469
quadruplicates. For cytotoxicity assays on BoMac, cells were seeded into 96-well-plates, treated with concentration series (0 to 20 µM) of 470
compounds 1 to 5 and of BPQ. After three days, viability was assayed using the Alamar blue assay. All assays were performed in quadruplicates. 471
IC50 values with confidence intervals were determined as described. N.d., not determined. Structures were obtained by conversion from SMILES 472
annotations via the ChemSpider website (www.chemspider.com). The five compounds 1-5 that were retained for further studies are highlighted 473
in bold. 474
475
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 19
19
476
Compound MMV code Structure A logP IC50 (µM)
TaC12
TaSP
(rel. mRNA level)
IC50 (µM)
BoMac
BPQ -
6.45 2.0 ± 0.5 x 10-3 5.2 ± 0.5 >20
1 000760
4.431 0.56 ± 0.03 24.0 ± 3.0 1.3 ± 0.1
2 666022
6.55 0.47 ± 0.02 31.7 ± 1.0 5.3 ± 1.5
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 20
20
3
666023
8.504 0.32 ± 0.04 27.5 ± 2.4 1.3 ± 0.1
4 666054
6.154 0.29 ± 0.02 23.6 ± 0.9 5.2 ± 0.4
5 665941
4.763 0.20 ± 0.08 24.2 ± 1.6 0.5 ± 0.2
6 006962 5.38 0.09 ± 0.02 263.3 ± 13.8 n.d.
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 21
21
7 396669
3.23 0.8 ± 0.03 57.0 ± 4.3 n.d.
8 638723
-1.93 0.40 ± 0.16 323.8 ± 22.8 n.d.
9 665814
5.73 750 ± 250 81.2 ± 5.4 n.d. on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 22
22
10 665820
3.59 0.75 ± 0.25 181.7 ± 10.5 n.d.
11 665969
5.88 0.38 ± 0.13 91.3 ± 10.6 n.d.
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 23
23
Figure legends 477
478
Fig. 1. The five most efficient malaria box hits and their respective dose 479
responses with respect to relative levels of TaSP mRNA levels in relation to 480
bovine actin transcripts. TaC12 cells were seeded into 25cm2 culture flasks and 481
treated with 0.1, 0.2, 0.5 and 1 µM of each compound for 24h. Relative TaSP 482
expression levels (in relation to bovine actin) were assessed by the qRT-PCR assay. 483
The experiment was done twice, with essentially identical outcome. Mean values (± 484
SE) for four biological replicates are shown. Low relative TaSP transcipt levels 485
indicated primary action of the drug against T. annulata, higher relative TaSP level 486
indicates that compounds could be acting primarily against the host cell. 487
488
Fig. 2. TEM of non-treated and BPQ treated TaC12 cells. A shows a low 489
magnification view of a non-treated control, and the boxed area is enlarged in B. C is 490
a representative image of a BPQ treated TaC12 cell, and the boxed area is shown at 491
higher magnification in D. Small arrows point towards the schizont-host cell 492
cytoplasm interface, v indicates vacuolization within the schizont cytoplasm, white 493
crosses (x) indicate button-like structures associated with schizonts, n = parasite 494
nuclei, N = host cell nucleus, mito = host cell mitochondria. Bars = 3.4µm in A, 495
0.7µm in B, 1.7µm in C and 0.5 µm in D. 496
497
Fig. 3. TEM of TaC12 cells treated with compounds (1), (2) and (3). A cell treated 498
with (1) is shown in A, B and C shows low and high magnification views, 499
respectively, of compound (2) treated cells, and D an E respective images of a TaC12 500
cell treated with compound (3). Small arrows point towards the schizont-host cell 501
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 24
24
cytoplasm interface, v indicates vacuolization within the schizont cytoplasm, vac 502
indicates the presence of vacuoles within the host cell cytoplasm, white crosses (x) 503
indicate button-like structures or their residues associated with schizonts, n = parasite 504
nuclei, N = host cell nucleus, mito = host cell mitochondria. Bars = 0.9µm in A, 505
3.4µm in B, 0.5µm in C, 3.4µm in D and 0.5µm in E. 506
507
Fig. 4. TEM of TaC12 cells exposed to compounds (4), and (5). A (low 508
magnification) and B (higher magnification view of the boxed area in A) show a 509
TaC12 cell exposed to compound (4). C and D show respective views of a TaC12 cell 510
treated with compound (5). Small arrows indicate the location of the schizont plasma 511
membrane, v indicates vacuolization within the schizont cytoplasm, white crosses (x) 512
indicate button-like structures or their residues associated with schizonts, n = parasite 513
nucleus, N = host cell nucleus, mito = host cell mitochondria. Bars = 2.8µm in A, 514
0.8µm in B, 3.4µm in C, 0.5µm in D. 515
516
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 25
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 26
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 27
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from
Page 28
on April 6, 2018 by guest
http://aac.asm.org/
Dow
nloaded from