Tastsuro Suzuki5 Kwang Hyun Jho6 and Sang Un Park1
1 Department of Crop Science Chungnam National University 99 Daehak-ro Yuseong-gu Daejeon 305-764 Republic of Korea2Department of Bioscience and Biotechnology University of Suwon San 2-2Wauri Bongdameup Hwasung 445-743 Republic of Korea3 Department of Botany and Microbiology Addiriyah Chair for Environmental Studies College of Science King Saud UniversityPO Box 2455 Riyadh 11451 Saudi Arabia
4Department of Bio-Environmental Chemistry Chungnam National University 99 Daehak-ro Yuseong-guDaejeon 305-764 Republic of Korea
5Hokkaido Agricultural Research Center National Agriculture and Food Research Organization Sapporo 062-8555 Japan6Department of Business Administration Sahmyook University Hwarangro 815 Nowon-gu Seoul 139-742 Republic of Korea
Correspondence should be addressed to Kwang Hyun Jho jhokh3679hanmailnet and Sang Un Park suparkcnuackr
Received 24 August 2013 Accepted 13 November 2013 Published 27 January 2014
Academic Editors C Lu M Tadege and Y Wu
Copyright copy 2014 Yeon Bok Kim et al This is an open access article distributed under the Creative Commons Attribution Licensewhich permits unrestricted use distribution and reproduction in any medium provided the original work is properly cited
Anthocyanidin reductase (ANR) and leucoanthocyanidin reductase (LAR) play an important role in the monomeric unitsbiosynthesis of proanthocyanidins (PAs) such as catechin and epicatechin in several plantsThe aim of this study was to clone ANRand LAR genes involved in PAs biosynthesis and examine the expression of these two genes in different organs under differentgrowth conditions in two tartary buckwheat cultivars Hokkai T8 and T10 Gene expression was carried out by quantitative real-time RT-PCR and catechin and epicatechin content was analyzed by high performance liquid chromatography The expressionpattern of ANR and LAR did not match the accumulation pattern of PAs in different organs of two cultivars Epicatechin contentwas the highest in the flowers of both cultivars and it was affected by light in only Hokkai T8 sprouts ANR and LAR levels in tartarybuckwheat might be regulated by different mechanisms for catechin and epicatechin biosynthesis under light and dark conditions
1 Introduction
Proanthocyanidins (PAs also known as condensed tannins)are phenolic oligomers or polymers that result from thepolymerization of flavan-3-ol units they are synthesized fromthe first metabolites via the shikimate and flavonoid pathway[1 2] PAs providemultiple health benefits to humans such asantioxidant anticancer and anticardiovascular effects [3ndash5]PAs are one of the final products of the flavonoid pathwayand contribute to the quality of many important plantproducts such as wine tea and cocoa [6] Winkel-Shirley
[7] reported that PAs are derived from the pathway leadingto anthocyanins a class of flavonoids well understood atboth the biochemical and molecular genetic levels Leucoan-thocyanidin reductase (LAR) and anthocyanidin reductase(ANR) are both key enzymes of the branch pathway ofPAs biosynthesis (Figure 1) ANR is one of two enzymesof the flavonoid biosynthesis pathway that produces flavan-3-ol (epicatechin) monomers producing epicatechin fromanthocyanidin [8] ANR was initially found to be encodedby the BANYULS (BAN) gene from Arabidopsis thaliana andMedicago truncatula [9] ANR enzymatic function has been
Hindawi Publishing Corporatione Scientific World JournalVolume 2014 Article ID 726567 10 pageshttpdxdoiorg1011552014726567
2 The Scientific World Journal
Phenylalanine
4-Coumaroyl-CoA+ 3-Malonyl-CoA
PAL C4H 4CL
Naringenin(flavonoids)
CHS CHI
Dihydrokaemferol
F3H
Dihydroquercetin
F3998400H
Leucoanthocyanidins
DFR
LAR ANR
Catechin Epicatechin
HO
OHOH
OH
OH
O HO
OHOH
OH
OH
O
Figure 1 Flavonoid biosynthesis pathway in F tataricum PAL phenylalanine ammonia lyase C4H cinnamate 4-hydroxylase 4CL 4-coumaroyl CoA ligase CHS chalcone synthase CHI chalcone isomerase F3H flavones 3-hydroxylase F31015840H flavonoid 31015840-hydroxylaseDFR dihydroflavonol-4 reductase ANR anthocyanidin reductase LAR leucoanthocyanidin reductase The red color genes were isolatedfrom F tataricum in this study
identified in various plants such as grape (Vitis vinifera)soybean (Glycine max) tea (Camellia sinensis) and legume(Medicago truncatula) [10ndash13] Bogs et al [10] reported thatLAR catalyzes the conversion of leucocyanidin to catechinclearly establishing its role in PA biosynthesis The function-ality of LARhas been reported in several plants and its activityis correlated with PA accumulation [11 13 14]
Buckwheat (Fagopyrum esculentum and Fagopyrum tatar-icum) a dicotyledonous crop of the Polygonaceae familyhas received attention as health food In particular tartarybuckwheat has also been found to have several beneficialpharmacological and biological effects such as anticancerantidiabetic and antioxidant activities [15ndash17] In additiontartary buckwheat is a richer source of rutin a flavonol glyco-side that prevents ultraviolet light-induced DNA damage anddisease than common buckwheat [16 17] Recent research
on buckwheat has focused on functional food materialparticularly with respect to seed sprouts in Korea and JapanTartary buckwheat sprouts are an excellent dietary source ofphenolic compounds [17] The sprouts of the F tataricumldquoHokkai T10rdquo (T10) cultivar derived from ldquoHokkai T8rdquo (T8)by chemical treatment (ethyl methane sulfonate) have ahigher anthocyanin content than common buckwheat [18]The duration and amount of light exposure that was usedfor sprouting strongly affect the nutritional quality of tartarybuckwheat sprouts [19]
When common buckwheat sprouts (F esculentum) weregrown under light or dark condition flavonoid content wasincreased significantly [20] In present study we cloned ANRand LAR genes involved in PAs biosynthesis and exam-ined the expression of these two genes in different organsunder different growth conditions in two tartary buckwheat
The Scientific World Journal 3
cultivars Hokkai T8 and T10 In addition we analyzedcontent of catechin and epicatechin in different organs andsprouts
2 Materials and Methods
21 Plant Materials and Growth Conditions Two tartarybuckwheat cultivars T8 and T10 were bred by the HokkaidoAgricultural Research Center (Hokkaido Japan) The seedswere surface-sterilized with 70 ethanol for 1min and4 (vv) bleach solution for 20min and then rinsed severaltimes in sterile water Sterilized seeds were germinated on12MS medium in a growth chamber under light condition(16 h light8 h darkness) and dark condition (24 h darkness) at25∘C 60 humidity and 440 120583molesm2s light intensity Forbiological replicates we used 3 plastic boxes for each singletreatment placing 30 seeds per box Sprouts including rootswere harvested at 0 3 6 9 and 12 days after sowing DASTheseeds of both T8 andT10were sown on 10May 2012 and thentransferred into pots filledwith the perlite-mixed soil Tartarybuckwheat plants were grown in the greenhouse (25∘C and50 humidity) at Chungnam National University (DaejeonKorea) Different plant organs (seed stages 1 2 and 3 flowersstems leaves and roots) were collected after 6 weeks Theseed stages were distinguished as described previously [21]All samples were frozen in liquid nitrogen upon collectionand stored at minus80∘C until use
22 Isolation of Genes Encoding ANR and LAR from Ftataricum The full-length cDNA sequence of putative ANRwas obtained from next generation sequencing platforms(NGS) (Roche454 GS FLX+ and IlluminaSolexaHiSeq2000) (unpublished data) of F tataricum of SolGentcompany (Daejeon Korea) LAR partial sequences wereobtained from NGS data and a full-length cDNA wasobtained using rapid amplification of cDNA ends (RACE)PCR Sequence similarities were calculated with the BasicLocal Alignment Search Tool (BLAST) (httpblastncbinlmnihgov) The sequence data were used to design newprimer pairs for RACE PCR qRT-PCR and ORF PCR andthe primes were mentioned in Supplementary Table S1 (seeSupplementary Material available online at httpdxdoiorg1011552014726567)
23 Total RNA Extraction and cDNA Synthesis Total RNAwas isolated from two tartary buckwheat sprouts anddifferentorgans The total RNA of sprouts was isolated using theRNeasy Plant Mini Kit (Qiagen Valencia CA) whereasthat from different organs was extracted by a modifiedCTAB (cetyltrimethylammonium bromide) method becauseof complex polysaccharides [22]The RNA pellet was washedwith 70 EtOH and dissolved in DEPC water RNA quantityand quality were determined by a NanoVue Plus Spec-trophotometer (GE Health Care Life Sciences USA) andassessed by running 1 120583g of total RNA on 12 formalde-hyde RNA agarose gel respectively Subsequently 1 120583g oftotal RNA was reverse-transcribed using the ReverTra Ace-120572-(Toyobo Osaka Japan) Kit and oligo (dT)
20primer
according to the manufacturerrsquos protocol The synthesized
cDNA was used as the template for qRT-PCR and RACEPCR
24 Quantitative Real-Time RT-PCR Analysis For qRT-PCRthe TM calculator program (httpbioinfouteeprimer3-040) was used to compute the PCR annealing temperaturesqRT-PCR assay was carried out in a total volume of 20120583Lcontaining 10 120583L of 2 X SYBR Green Real-time PCR mastermixes (Toyobo Osaka Japan) 05 120583M (each) of specificprimers and 5 120583L of cDNA that was diluted 20-fold Theamplification program consisted of one cycle of 95∘C for3min followed by 40 cycles of 95∘C for 15 s 72∘C for 20 s andannealing temperature 55∘C for 30 s The reaction was per-formed in triplicate on a CFX96 Real-Time PCR System (Bio-Rad Hercules CA USA) The histone H3 gene (GenBanknumber HM628903) was used as a reference gene [19ndash21]
25 Bioinformatic Analysis of FtANR and FtLAR Alignmentof the deduced amino acid sequences of FtANR and FtLARwas carried out using the Biological Sequence AlignmentEditor (BioEdit) software The phylogenetic relationships ofFtANR and FtLAR were analyzed using ClustalX andMEGAversion 40 In the bootstrap the multiple alignment wasresampled 100 times Theoretical molecular weights and pIvalues were calculated by the Compute pIMw tool (httpcaexpasyorgtoolspi toolhtml) The secondary structure waspredicted using SOPMA (httpnpsa-pbilibcpfrcgi-binnpsa automatplpage=NPSAnpsa sopmahtml) The puta-tive target location of the plant was predicted online throughPSORT (httpwolfpsortorg)
26 Estimation of Catechin and Epicatechin Chemical anal-ysis of catechin and epicatechin was carried out by HPLCanalysis and we used a minor modification of a previouslypublished method [23] For this freeze-dried samples ofbuckwheat sprouts and different organs were ground into afine powder using a mortar and pestle Powdered samples(sim100mg) were extracted with 80 (vv) methanol at roomtemperature for 60min Subsequently the extracts werecentrifuged and the supernatant was filtered with a 045 120583mAcrodisc syringe filter (Pall Corp Port Washington NY) forHPLC HPLC analysis was performed with a C18 column(120583Bondapak C18 10 120583m 125 A 39 times 300mm) The mobilephase was a gradient pattern prepared from mixtures ofmethanol and 05 acetic acid The flow rate was maintainedat 08mLmin An injection volume of 20 120583L and wavelengthof 280 nm were used for detection The compounds in thesample were determined using a standard curve All sampleswere analyzed in triplicate
3 Results and Discussion
31 Isolation and Sequence Analysis of FtANR and FtLAR fromTartary Buckwheat Open reading frame (ORF) of FtANR(Genbank Accession number KC404848) obtained fromNGS data for tartary buckwheat was 1011 bp long encodinga protein of 336 amino acids with a theoretical molecularweight of 368 kDa and a pI value of 542Medicago truncatula(MtANR) andArabidopsis thaliana (AtANR) consisted of 338
and 340 amino acids with molecular weights of 369 and379 kDa respectively [24] FromBLAST analysis of theANR-deduced amino acid sequences FtANR was found to share73 77 82 78 and 80 identities with M truncatula(AAN77735) Malus times domestica (JN035299) Gossypiumhirsutum (EF187443) Camellia sinensis (AAT68773) andV vinifera (BAD89742) respectively FtANR was found tohave a conserved motif GXGXXA similar to that found inmonodehydroascorbate reductase (NADH) (SupplementaryFigure S1) [25]
Using RACE technology with partial sequences obtainedfromNGS data we isolated 1581-bp-long FtLAR cDNA (Gen-bank accession number KC404849) from flowers of F tatar-icum T10 which contained a 1176-bp ORF this gene encoded
a protein of 391 amino acids with a theoretical molecularmass of 433 kDa and a pI value of 527 Recently Ma et al[26] cloned and characterized the LAR gene from Fagopyrumdibotrys The amino acid sequences of FtLAR and FdLARwere found to differ by only 12 amino acids The FdLARtarget fusion peptide had a molecular weight of 66 kDa [26]which did not correspond to the theoretical molecular weightof FtLAR The amino acid sequence of FtLAR showed 6870 69 and 66 identity with V vinifera (AAZ82410)Prunus avium (ADY15310) Diospyros kaki (BAH89267) andMalus times domestica (AAX12186) respectively In particularFtLAR showed 97 homology with F dibotrys (JN793953)As reported previously [10 27] FtLAR had specific aminoacid motifs of ICCN and THD (Supplementary Figure S2)
The Scientific World Journal 5
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
6
7
0 3 6 9 12
FtANR
Days after sowing
(a)
0 3 6 9 12
FtANR
Gen
e exp
ress
ion
relat
ive t
o H3
Days after sowing
0
5
10
15
20
25
(b)
0 3 6 9 12
T8-LT10-L
FtLAR
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
(c)
0 3 6 9 12
T8-DT10-D
FtLAR
0
1
2
3
4
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
(d)
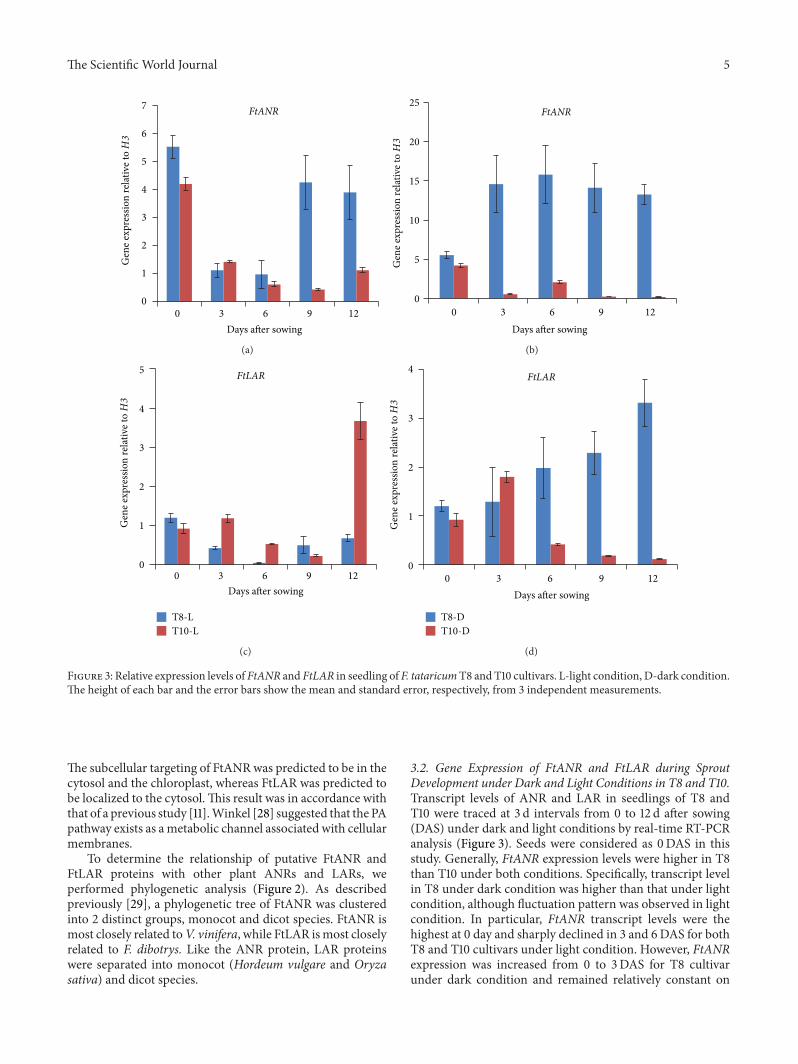
Figure 3 Relative expression levels of FtANR and FtLAR in seedling of F tataricumT8 and T10 cultivars L-light condition D-dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
The subcellular targeting of FtANRwas predicted to be in thecytosol and the chloroplast whereas FtLAR was predicted tobe localized to the cytosolThis result was in accordance withthat of a previous study [11]Winkel [28] suggested that the PApathway exists as ametabolic channel associated with cellularmembranes
To determine the relationship of putative FtANR andFtLAR proteins with other plant ANRs and LARs weperformed phylogenetic analysis (Figure 2) As describedpreviously [29] a phylogenetic tree of FtANR was clusteredinto 2 distinct groups monocot and dicot species FtANR ismost closely related toV vinifera while FtLAR ismost closelyrelated to F dibotrys Like the ANR protein LAR proteinswere separated into monocot (Hordeum vulgare and Oryzasativa) and dicot species
32 Gene Expression of FtANR and FtLAR during SproutDevelopment under Dark and Light Conditions in T8 and T10Transcript levels of ANR and LAR in seedlings of T8 andT10 were traced at 3 d intervals from 0 to 12 d after sowing(DAS) under dark and light conditions by real-time RT-PCRanalysis (Figure 3) Seeds were considered as 0DAS in thisstudy Generally FtANR expression levels were higher in T8than T10 under both conditions Specifically transcript levelin T8 under dark condition was higher than that under lightcondition although fluctuation pattern was observed in lightcondition In particular FtANR transcript levels were thehighest at 0 day and sharply declined in 3 and 6DAS for bothT8 and T10 cultivars under light condition However FtANRexpression was increased from 0 to 3DAS for T8 cultivarunder dark condition and remained relatively constant on
6 The Scientific World Journal
00
02
04
06
08
10
12
F L ST SD1 SD2 SD3 RT
FtANRG
ene e
xpre
ssio
n re
lativ
e to H3
(a)
T8T10
Gen
e exp
ress
ion
relat
ive t
o H3
02
04
FtLAR
00
06
08
F L ST SD1 SD2 SD3 RT
(b)
Figure 4 Relative expression levels of FtANR and FtLAR in different organs of F tataricum T8 and T10 cultivars F flowers ST stems Lleaves SD 1 2 3 seed stage 1 2 3 RT roots The height of each bar and the error bars show the mean and standard error respectively from3 independent measurements
6 9 and 12DAS Unlike FtANR different gene expressionpattern was observed in FtLARThe highest gene expressionlevel was observed at 12DAS in T10 which was nearly 5 timeshigher than that in T8 under light condition In additiona gradual increase in transcript levels was observed in T8under dark condition which is similar as FtANR under darkconditionTherefore in this study the transcription ofFtANRand FtLAR in T8 and T10 sprouts was unaffected by lightconditions
33 Gene Expression of FtANR and FtLAR among DifferentOrgans of T8 andT10 ThemRNA levels ofFtANR andFtLARamong different organs (flower leaf stem root and seeds instages 1 2 and 3) were shown in Figure 4 Unlike transcriptlevels during sprout development FtANR and FtLAR didnot show large variations in their expression patterns amongall the organs in T8 and T10 In some apple species LAR1expression was similar in the cortex and skin while ANRexpression levels were similar in apple skin flesh [30] Therelative levels of P trichocarpa LAR3 transcripts in the rootswere 2- and 3-fold higher than those in the stems andpetioles respectively [31] Bogs et al [10] reported that thetwo LAR genes involved in PA biosynthesis had differentpatterns of expression in grape seeds and skins which affectthe concentration and composition of PAs They suggestedthat V vinifera ANR may have a more significant role inPAs synthesis in grapevine leaves than LAR In additionGlycine max ANR1 is predominantly expressed in the seedcoat whereas GmANR2 is expressed at low levels in allorgans [29] Devic et al [32] reported that the expressionof the BAN gene is limited to the endothelium of immatureseeds in Arabidopsis whereas in Medicago its expressionis seen in young seeds flowers and leaves [9] Recently itwas reported that overexpression of LAR3 in Chinese whitepoplar (Populus tomentosa) leads to plantwide increase in PAlevels that is the highest in the roots [31] In our study we
found that the transcript levels of FtANR and FtLAR weresimilar among different organs According to the transcriptlevels observed in other plants it appears that FtANR andFtLAR are regulated differently compared to other plantspecies It is also possible that FtANR and FtLAR isoformsmay exist and these isoforms may contribute to catechin andepicatechin biosynthesis observed in this study in T8 and T10tartary buckwheat
34 Analysis of Catechin and Epicatechin during SeedlingDevelopment of Tartary Buckwheat The composition of cat-echin and epicatechin during seedling development from T8andT10was determined byHPLC (Figure 5) Epicatechinwasnot detected in T10 sprouts under light conditions whereasT8 sprouts had a high level of epicatechin (Figure 5(a)) Theepicatechin content of T8 was the highest (66mg gminus1 dryweight [DW]) at 12DAS under light conditionsThe oppositepattern was observed under dark conditions where theepicatechin content of T10 was higher than that of T8 Underdark conditions the epicatechin content of T10 was low inseeds and at 3DAS but reached higher levels on 6 9 and12DAS (eg 047mg gminus1DW Day 0 and 29mg gminus1DW on6DAS) while that of T8 increased from 0 (027mg gminus1DW)to 9DAS (279mg gminus1DW) (Figure 5(b)) Under light condi-tions the epicatechin content of T8 increased gradually from6 to 12DAS The epicatechin content of T8 was the highest(66mg gminus1DW) at 12DAS under light conditions while T10exhibited the highest content (31mg gminus1DW) at 6DASunderdark conditions
The catechin content of T8 under light and dark con-ditions was higher than that of T10 (Figures 5(c) and5(d)) The catechin content of T8 was the highest at 6DAS(196mg gminus1DW) and 12DAS (075mg gminus1DW) under darkand light conditions The catechin content of T10 was thehighest at 0DAS (055mg gminus1DW) Under dark conditions
The Scientific World Journal 7
Days after sowing
0
2
4
6
8
0 3 6 9 12
(mg
gminus1
dry
wt)
Epicatechin
(a)
0
1
2
3
4
0 3 6 9 12
(mg
gminus1
dry
wt)
Days after sowing
Epicatechin
(b)
0 3 6 9 12
0
02
04
06
08
1
(mg
gminus1
dry
wt)
Days after sowing
T8-LT10-L
Catechin
(c)
0 3 6 9 12
0
05
1
15
2
25
(mg
gminus1
dry
wt)
Days after sowing
T8-DT10-D
Catechin
(d)
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
Figure 1 Flavonoid biosynthesis pathway in F tataricum PAL phenylalanine ammonia lyase C4H cinnamate 4-hydroxylase 4CL 4-coumaroyl CoA ligase CHS chalcone synthase CHI chalcone isomerase F3H flavones 3-hydroxylase F31015840H flavonoid 31015840-hydroxylaseDFR dihydroflavonol-4 reductase ANR anthocyanidin reductase LAR leucoanthocyanidin reductase The red color genes were isolatedfrom F tataricum in this study
identified in various plants such as grape (Vitis vinifera)soybean (Glycine max) tea (Camellia sinensis) and legume(Medicago truncatula) [10ndash13] Bogs et al [10] reported thatLAR catalyzes the conversion of leucocyanidin to catechinclearly establishing its role in PA biosynthesis The function-ality of LARhas been reported in several plants and its activityis correlated with PA accumulation [11 13 14]
Buckwheat (Fagopyrum esculentum and Fagopyrum tatar-icum) a dicotyledonous crop of the Polygonaceae familyhas received attention as health food In particular tartarybuckwheat has also been found to have several beneficialpharmacological and biological effects such as anticancerantidiabetic and antioxidant activities [15ndash17] In additiontartary buckwheat is a richer source of rutin a flavonol glyco-side that prevents ultraviolet light-induced DNA damage anddisease than common buckwheat [16 17] Recent research
on buckwheat has focused on functional food materialparticularly with respect to seed sprouts in Korea and JapanTartary buckwheat sprouts are an excellent dietary source ofphenolic compounds [17] The sprouts of the F tataricumldquoHokkai T10rdquo (T10) cultivar derived from ldquoHokkai T8rdquo (T8)by chemical treatment (ethyl methane sulfonate) have ahigher anthocyanin content than common buckwheat [18]The duration and amount of light exposure that was usedfor sprouting strongly affect the nutritional quality of tartarybuckwheat sprouts [19]
When common buckwheat sprouts (F esculentum) weregrown under light or dark condition flavonoid content wasincreased significantly [20] In present study we cloned ANRand LAR genes involved in PAs biosynthesis and exam-ined the expression of these two genes in different organsunder different growth conditions in two tartary buckwheat
The Scientific World Journal 3
cultivars Hokkai T8 and T10 In addition we analyzedcontent of catechin and epicatechin in different organs andsprouts
2 Materials and Methods
21 Plant Materials and Growth Conditions Two tartarybuckwheat cultivars T8 and T10 were bred by the HokkaidoAgricultural Research Center (Hokkaido Japan) The seedswere surface-sterilized with 70 ethanol for 1min and4 (vv) bleach solution for 20min and then rinsed severaltimes in sterile water Sterilized seeds were germinated on12MS medium in a growth chamber under light condition(16 h light8 h darkness) and dark condition (24 h darkness) at25∘C 60 humidity and 440 120583molesm2s light intensity Forbiological replicates we used 3 plastic boxes for each singletreatment placing 30 seeds per box Sprouts including rootswere harvested at 0 3 6 9 and 12 days after sowing DASTheseeds of both T8 andT10were sown on 10May 2012 and thentransferred into pots filledwith the perlite-mixed soil Tartarybuckwheat plants were grown in the greenhouse (25∘C and50 humidity) at Chungnam National University (DaejeonKorea) Different plant organs (seed stages 1 2 and 3 flowersstems leaves and roots) were collected after 6 weeks Theseed stages were distinguished as described previously [21]All samples were frozen in liquid nitrogen upon collectionand stored at minus80∘C until use
22 Isolation of Genes Encoding ANR and LAR from Ftataricum The full-length cDNA sequence of putative ANRwas obtained from next generation sequencing platforms(NGS) (Roche454 GS FLX+ and IlluminaSolexaHiSeq2000) (unpublished data) of F tataricum of SolGentcompany (Daejeon Korea) LAR partial sequences wereobtained from NGS data and a full-length cDNA wasobtained using rapid amplification of cDNA ends (RACE)PCR Sequence similarities were calculated with the BasicLocal Alignment Search Tool (BLAST) (httpblastncbinlmnihgov) The sequence data were used to design newprimer pairs for RACE PCR qRT-PCR and ORF PCR andthe primes were mentioned in Supplementary Table S1 (seeSupplementary Material available online at httpdxdoiorg1011552014726567)
23 Total RNA Extraction and cDNA Synthesis Total RNAwas isolated from two tartary buckwheat sprouts anddifferentorgans The total RNA of sprouts was isolated using theRNeasy Plant Mini Kit (Qiagen Valencia CA) whereasthat from different organs was extracted by a modifiedCTAB (cetyltrimethylammonium bromide) method becauseof complex polysaccharides [22]The RNA pellet was washedwith 70 EtOH and dissolved in DEPC water RNA quantityand quality were determined by a NanoVue Plus Spec-trophotometer (GE Health Care Life Sciences USA) andassessed by running 1 120583g of total RNA on 12 formalde-hyde RNA agarose gel respectively Subsequently 1 120583g oftotal RNA was reverse-transcribed using the ReverTra Ace-120572-(Toyobo Osaka Japan) Kit and oligo (dT)
20primer
according to the manufacturerrsquos protocol The synthesized
cDNA was used as the template for qRT-PCR and RACEPCR
24 Quantitative Real-Time RT-PCR Analysis For qRT-PCRthe TM calculator program (httpbioinfouteeprimer3-040) was used to compute the PCR annealing temperaturesqRT-PCR assay was carried out in a total volume of 20120583Lcontaining 10 120583L of 2 X SYBR Green Real-time PCR mastermixes (Toyobo Osaka Japan) 05 120583M (each) of specificprimers and 5 120583L of cDNA that was diluted 20-fold Theamplification program consisted of one cycle of 95∘C for3min followed by 40 cycles of 95∘C for 15 s 72∘C for 20 s andannealing temperature 55∘C for 30 s The reaction was per-formed in triplicate on a CFX96 Real-Time PCR System (Bio-Rad Hercules CA USA) The histone H3 gene (GenBanknumber HM628903) was used as a reference gene [19ndash21]
25 Bioinformatic Analysis of FtANR and FtLAR Alignmentof the deduced amino acid sequences of FtANR and FtLARwas carried out using the Biological Sequence AlignmentEditor (BioEdit) software The phylogenetic relationships ofFtANR and FtLAR were analyzed using ClustalX andMEGAversion 40 In the bootstrap the multiple alignment wasresampled 100 times Theoretical molecular weights and pIvalues were calculated by the Compute pIMw tool (httpcaexpasyorgtoolspi toolhtml) The secondary structure waspredicted using SOPMA (httpnpsa-pbilibcpfrcgi-binnpsa automatplpage=NPSAnpsa sopmahtml) The puta-tive target location of the plant was predicted online throughPSORT (httpwolfpsortorg)
26 Estimation of Catechin and Epicatechin Chemical anal-ysis of catechin and epicatechin was carried out by HPLCanalysis and we used a minor modification of a previouslypublished method [23] For this freeze-dried samples ofbuckwheat sprouts and different organs were ground into afine powder using a mortar and pestle Powdered samples(sim100mg) were extracted with 80 (vv) methanol at roomtemperature for 60min Subsequently the extracts werecentrifuged and the supernatant was filtered with a 045 120583mAcrodisc syringe filter (Pall Corp Port Washington NY) forHPLC HPLC analysis was performed with a C18 column(120583Bondapak C18 10 120583m 125 A 39 times 300mm) The mobilephase was a gradient pattern prepared from mixtures ofmethanol and 05 acetic acid The flow rate was maintainedat 08mLmin An injection volume of 20 120583L and wavelengthof 280 nm were used for detection The compounds in thesample were determined using a standard curve All sampleswere analyzed in triplicate
3 Results and Discussion
31 Isolation and Sequence Analysis of FtANR and FtLAR fromTartary Buckwheat Open reading frame (ORF) of FtANR(Genbank Accession number KC404848) obtained fromNGS data for tartary buckwheat was 1011 bp long encodinga protein of 336 amino acids with a theoretical molecularweight of 368 kDa and a pI value of 542Medicago truncatula(MtANR) andArabidopsis thaliana (AtANR) consisted of 338
and 340 amino acids with molecular weights of 369 and379 kDa respectively [24] FromBLAST analysis of theANR-deduced amino acid sequences FtANR was found to share73 77 82 78 and 80 identities with M truncatula(AAN77735) Malus times domestica (JN035299) Gossypiumhirsutum (EF187443) Camellia sinensis (AAT68773) andV vinifera (BAD89742) respectively FtANR was found tohave a conserved motif GXGXXA similar to that found inmonodehydroascorbate reductase (NADH) (SupplementaryFigure S1) [25]
Using RACE technology with partial sequences obtainedfromNGS data we isolated 1581-bp-long FtLAR cDNA (Gen-bank accession number KC404849) from flowers of F tatar-icum T10 which contained a 1176-bp ORF this gene encoded
a protein of 391 amino acids with a theoretical molecularmass of 433 kDa and a pI value of 527 Recently Ma et al[26] cloned and characterized the LAR gene from Fagopyrumdibotrys The amino acid sequences of FtLAR and FdLARwere found to differ by only 12 amino acids The FdLARtarget fusion peptide had a molecular weight of 66 kDa [26]which did not correspond to the theoretical molecular weightof FtLAR The amino acid sequence of FtLAR showed 6870 69 and 66 identity with V vinifera (AAZ82410)Prunus avium (ADY15310) Diospyros kaki (BAH89267) andMalus times domestica (AAX12186) respectively In particularFtLAR showed 97 homology with F dibotrys (JN793953)As reported previously [10 27] FtLAR had specific aminoacid motifs of ICCN and THD (Supplementary Figure S2)
The Scientific World Journal 5
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
6
7
0 3 6 9 12
FtANR
Days after sowing
(a)
0 3 6 9 12
FtANR
Gen
e exp
ress
ion
relat
ive t
o H3
Days after sowing
0
5
10
15
20
25
(b)
0 3 6 9 12
T8-LT10-L
FtLAR
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
(c)
0 3 6 9 12
T8-DT10-D
FtLAR
0
1
2
3
4
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
(d)
Figure 3 Relative expression levels of FtANR and FtLAR in seedling of F tataricumT8 and T10 cultivars L-light condition D-dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
The subcellular targeting of FtANRwas predicted to be in thecytosol and the chloroplast whereas FtLAR was predicted tobe localized to the cytosolThis result was in accordance withthat of a previous study [11]Winkel [28] suggested that the PApathway exists as ametabolic channel associated with cellularmembranes
To determine the relationship of putative FtANR andFtLAR proteins with other plant ANRs and LARs weperformed phylogenetic analysis (Figure 2) As describedpreviously [29] a phylogenetic tree of FtANR was clusteredinto 2 distinct groups monocot and dicot species FtANR ismost closely related toV vinifera while FtLAR ismost closelyrelated to F dibotrys Like the ANR protein LAR proteinswere separated into monocot (Hordeum vulgare and Oryzasativa) and dicot species
32 Gene Expression of FtANR and FtLAR during SproutDevelopment under Dark and Light Conditions in T8 and T10Transcript levels of ANR and LAR in seedlings of T8 andT10 were traced at 3 d intervals from 0 to 12 d after sowing(DAS) under dark and light conditions by real-time RT-PCRanalysis (Figure 3) Seeds were considered as 0DAS in thisstudy Generally FtANR expression levels were higher in T8than T10 under both conditions Specifically transcript levelin T8 under dark condition was higher than that under lightcondition although fluctuation pattern was observed in lightcondition In particular FtANR transcript levels were thehighest at 0 day and sharply declined in 3 and 6DAS for bothT8 and T10 cultivars under light condition However FtANRexpression was increased from 0 to 3DAS for T8 cultivarunder dark condition and remained relatively constant on
6 The Scientific World Journal
00
02
04
06
08
10
12
F L ST SD1 SD2 SD3 RT
FtANRG
ene e
xpre
ssio
n re
lativ
e to H3
(a)
T8T10
Gen
e exp
ress
ion
relat
ive t
o H3
02
04
FtLAR
00
06
08
F L ST SD1 SD2 SD3 RT
(b)
Figure 4 Relative expression levels of FtANR and FtLAR in different organs of F tataricum T8 and T10 cultivars F flowers ST stems Lleaves SD 1 2 3 seed stage 1 2 3 RT roots The height of each bar and the error bars show the mean and standard error respectively from3 independent measurements
6 9 and 12DAS Unlike FtANR different gene expressionpattern was observed in FtLARThe highest gene expressionlevel was observed at 12DAS in T10 which was nearly 5 timeshigher than that in T8 under light condition In additiona gradual increase in transcript levels was observed in T8under dark condition which is similar as FtANR under darkconditionTherefore in this study the transcription ofFtANRand FtLAR in T8 and T10 sprouts was unaffected by lightconditions
33 Gene Expression of FtANR and FtLAR among DifferentOrgans of T8 andT10 ThemRNA levels ofFtANR andFtLARamong different organs (flower leaf stem root and seeds instages 1 2 and 3) were shown in Figure 4 Unlike transcriptlevels during sprout development FtANR and FtLAR didnot show large variations in their expression patterns amongall the organs in T8 and T10 In some apple species LAR1expression was similar in the cortex and skin while ANRexpression levels were similar in apple skin flesh [30] Therelative levels of P trichocarpa LAR3 transcripts in the rootswere 2- and 3-fold higher than those in the stems andpetioles respectively [31] Bogs et al [10] reported that thetwo LAR genes involved in PA biosynthesis had differentpatterns of expression in grape seeds and skins which affectthe concentration and composition of PAs They suggestedthat V vinifera ANR may have a more significant role inPAs synthesis in grapevine leaves than LAR In additionGlycine max ANR1 is predominantly expressed in the seedcoat whereas GmANR2 is expressed at low levels in allorgans [29] Devic et al [32] reported that the expressionof the BAN gene is limited to the endothelium of immatureseeds in Arabidopsis whereas in Medicago its expressionis seen in young seeds flowers and leaves [9] Recently itwas reported that overexpression of LAR3 in Chinese whitepoplar (Populus tomentosa) leads to plantwide increase in PAlevels that is the highest in the roots [31] In our study we
found that the transcript levels of FtANR and FtLAR weresimilar among different organs According to the transcriptlevels observed in other plants it appears that FtANR andFtLAR are regulated differently compared to other plantspecies It is also possible that FtANR and FtLAR isoformsmay exist and these isoforms may contribute to catechin andepicatechin biosynthesis observed in this study in T8 and T10tartary buckwheat
34 Analysis of Catechin and Epicatechin during SeedlingDevelopment of Tartary Buckwheat The composition of cat-echin and epicatechin during seedling development from T8andT10was determined byHPLC (Figure 5) Epicatechinwasnot detected in T10 sprouts under light conditions whereasT8 sprouts had a high level of epicatechin (Figure 5(a)) Theepicatechin content of T8 was the highest (66mg gminus1 dryweight [DW]) at 12DAS under light conditionsThe oppositepattern was observed under dark conditions where theepicatechin content of T10 was higher than that of T8 Underdark conditions the epicatechin content of T10 was low inseeds and at 3DAS but reached higher levels on 6 9 and12DAS (eg 047mg gminus1DW Day 0 and 29mg gminus1DW on6DAS) while that of T8 increased from 0 (027mg gminus1DW)to 9DAS (279mg gminus1DW) (Figure 5(b)) Under light condi-tions the epicatechin content of T8 increased gradually from6 to 12DAS The epicatechin content of T8 was the highest(66mg gminus1DW) at 12DAS under light conditions while T10exhibited the highest content (31mg gminus1DW) at 6DASunderdark conditions
The catechin content of T8 under light and dark con-ditions was higher than that of T10 (Figures 5(c) and5(d)) The catechin content of T8 was the highest at 6DAS(196mg gminus1DW) and 12DAS (075mg gminus1DW) under darkand light conditions The catechin content of T10 was thehighest at 0DAS (055mg gminus1DW) Under dark conditions
The Scientific World Journal 7
Days after sowing
0
2
4
6
8
0 3 6 9 12
(mg
gminus1
dry
wt)
Epicatechin
(a)
0
1
2
3
4
0 3 6 9 12
(mg
gminus1
dry
wt)
Days after sowing
Epicatechin
(b)
0 3 6 9 12
0
02
04
06
08
1
(mg
gminus1
dry
wt)
Days after sowing
T8-LT10-L
Catechin
(c)
0 3 6 9 12
0
05
1
15
2
25
(mg
gminus1
dry
wt)
Days after sowing
T8-DT10-D
Catechin
(d)
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
cultivars Hokkai T8 and T10 In addition we analyzedcontent of catechin and epicatechin in different organs andsprouts
2 Materials and Methods
21 Plant Materials and Growth Conditions Two tartarybuckwheat cultivars T8 and T10 were bred by the HokkaidoAgricultural Research Center (Hokkaido Japan) The seedswere surface-sterilized with 70 ethanol for 1min and4 (vv) bleach solution for 20min and then rinsed severaltimes in sterile water Sterilized seeds were germinated on12MS medium in a growth chamber under light condition(16 h light8 h darkness) and dark condition (24 h darkness) at25∘C 60 humidity and 440 120583molesm2s light intensity Forbiological replicates we used 3 plastic boxes for each singletreatment placing 30 seeds per box Sprouts including rootswere harvested at 0 3 6 9 and 12 days after sowing DASTheseeds of both T8 andT10were sown on 10May 2012 and thentransferred into pots filledwith the perlite-mixed soil Tartarybuckwheat plants were grown in the greenhouse (25∘C and50 humidity) at Chungnam National University (DaejeonKorea) Different plant organs (seed stages 1 2 and 3 flowersstems leaves and roots) were collected after 6 weeks Theseed stages were distinguished as described previously [21]All samples were frozen in liquid nitrogen upon collectionand stored at minus80∘C until use
22 Isolation of Genes Encoding ANR and LAR from Ftataricum The full-length cDNA sequence of putative ANRwas obtained from next generation sequencing platforms(NGS) (Roche454 GS FLX+ and IlluminaSolexaHiSeq2000) (unpublished data) of F tataricum of SolGentcompany (Daejeon Korea) LAR partial sequences wereobtained from NGS data and a full-length cDNA wasobtained using rapid amplification of cDNA ends (RACE)PCR Sequence similarities were calculated with the BasicLocal Alignment Search Tool (BLAST) (httpblastncbinlmnihgov) The sequence data were used to design newprimer pairs for RACE PCR qRT-PCR and ORF PCR andthe primes were mentioned in Supplementary Table S1 (seeSupplementary Material available online at httpdxdoiorg1011552014726567)
23 Total RNA Extraction and cDNA Synthesis Total RNAwas isolated from two tartary buckwheat sprouts anddifferentorgans The total RNA of sprouts was isolated using theRNeasy Plant Mini Kit (Qiagen Valencia CA) whereasthat from different organs was extracted by a modifiedCTAB (cetyltrimethylammonium bromide) method becauseof complex polysaccharides [22]The RNA pellet was washedwith 70 EtOH and dissolved in DEPC water RNA quantityand quality were determined by a NanoVue Plus Spec-trophotometer (GE Health Care Life Sciences USA) andassessed by running 1 120583g of total RNA on 12 formalde-hyde RNA agarose gel respectively Subsequently 1 120583g oftotal RNA was reverse-transcribed using the ReverTra Ace-120572-(Toyobo Osaka Japan) Kit and oligo (dT)
20primer
according to the manufacturerrsquos protocol The synthesized
cDNA was used as the template for qRT-PCR and RACEPCR
24 Quantitative Real-Time RT-PCR Analysis For qRT-PCRthe TM calculator program (httpbioinfouteeprimer3-040) was used to compute the PCR annealing temperaturesqRT-PCR assay was carried out in a total volume of 20120583Lcontaining 10 120583L of 2 X SYBR Green Real-time PCR mastermixes (Toyobo Osaka Japan) 05 120583M (each) of specificprimers and 5 120583L of cDNA that was diluted 20-fold Theamplification program consisted of one cycle of 95∘C for3min followed by 40 cycles of 95∘C for 15 s 72∘C for 20 s andannealing temperature 55∘C for 30 s The reaction was per-formed in triplicate on a CFX96 Real-Time PCR System (Bio-Rad Hercules CA USA) The histone H3 gene (GenBanknumber HM628903) was used as a reference gene [19ndash21]
25 Bioinformatic Analysis of FtANR and FtLAR Alignmentof the deduced amino acid sequences of FtANR and FtLARwas carried out using the Biological Sequence AlignmentEditor (BioEdit) software The phylogenetic relationships ofFtANR and FtLAR were analyzed using ClustalX andMEGAversion 40 In the bootstrap the multiple alignment wasresampled 100 times Theoretical molecular weights and pIvalues were calculated by the Compute pIMw tool (httpcaexpasyorgtoolspi toolhtml) The secondary structure waspredicted using SOPMA (httpnpsa-pbilibcpfrcgi-binnpsa automatplpage=NPSAnpsa sopmahtml) The puta-tive target location of the plant was predicted online throughPSORT (httpwolfpsortorg)
26 Estimation of Catechin and Epicatechin Chemical anal-ysis of catechin and epicatechin was carried out by HPLCanalysis and we used a minor modification of a previouslypublished method [23] For this freeze-dried samples ofbuckwheat sprouts and different organs were ground into afine powder using a mortar and pestle Powdered samples(sim100mg) were extracted with 80 (vv) methanol at roomtemperature for 60min Subsequently the extracts werecentrifuged and the supernatant was filtered with a 045 120583mAcrodisc syringe filter (Pall Corp Port Washington NY) forHPLC HPLC analysis was performed with a C18 column(120583Bondapak C18 10 120583m 125 A 39 times 300mm) The mobilephase was a gradient pattern prepared from mixtures ofmethanol and 05 acetic acid The flow rate was maintainedat 08mLmin An injection volume of 20 120583L and wavelengthof 280 nm were used for detection The compounds in thesample were determined using a standard curve All sampleswere analyzed in triplicate
3 Results and Discussion
31 Isolation and Sequence Analysis of FtANR and FtLAR fromTartary Buckwheat Open reading frame (ORF) of FtANR(Genbank Accession number KC404848) obtained fromNGS data for tartary buckwheat was 1011 bp long encodinga protein of 336 amino acids with a theoretical molecularweight of 368 kDa and a pI value of 542Medicago truncatula(MtANR) andArabidopsis thaliana (AtANR) consisted of 338
and 340 amino acids with molecular weights of 369 and379 kDa respectively [24] FromBLAST analysis of theANR-deduced amino acid sequences FtANR was found to share73 77 82 78 and 80 identities with M truncatula(AAN77735) Malus times domestica (JN035299) Gossypiumhirsutum (EF187443) Camellia sinensis (AAT68773) andV vinifera (BAD89742) respectively FtANR was found tohave a conserved motif GXGXXA similar to that found inmonodehydroascorbate reductase (NADH) (SupplementaryFigure S1) [25]
Using RACE technology with partial sequences obtainedfromNGS data we isolated 1581-bp-long FtLAR cDNA (Gen-bank accession number KC404849) from flowers of F tatar-icum T10 which contained a 1176-bp ORF this gene encoded
a protein of 391 amino acids with a theoretical molecularmass of 433 kDa and a pI value of 527 Recently Ma et al[26] cloned and characterized the LAR gene from Fagopyrumdibotrys The amino acid sequences of FtLAR and FdLARwere found to differ by only 12 amino acids The FdLARtarget fusion peptide had a molecular weight of 66 kDa [26]which did not correspond to the theoretical molecular weightof FtLAR The amino acid sequence of FtLAR showed 6870 69 and 66 identity with V vinifera (AAZ82410)Prunus avium (ADY15310) Diospyros kaki (BAH89267) andMalus times domestica (AAX12186) respectively In particularFtLAR showed 97 homology with F dibotrys (JN793953)As reported previously [10 27] FtLAR had specific aminoacid motifs of ICCN and THD (Supplementary Figure S2)
The Scientific World Journal 5
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
6
7
0 3 6 9 12
FtANR
Days after sowing
(a)
0 3 6 9 12
FtANR
Gen
e exp
ress
ion
relat
ive t
o H3
Days after sowing
0
5
10
15
20
25
(b)
0 3 6 9 12
T8-LT10-L
FtLAR
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
(c)
0 3 6 9 12
T8-DT10-D
FtLAR
0
1
2
3
4
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
(d)
Figure 3 Relative expression levels of FtANR and FtLAR in seedling of F tataricumT8 and T10 cultivars L-light condition D-dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
The subcellular targeting of FtANRwas predicted to be in thecytosol and the chloroplast whereas FtLAR was predicted tobe localized to the cytosolThis result was in accordance withthat of a previous study [11]Winkel [28] suggested that the PApathway exists as ametabolic channel associated with cellularmembranes
To determine the relationship of putative FtANR andFtLAR proteins with other plant ANRs and LARs weperformed phylogenetic analysis (Figure 2) As describedpreviously [29] a phylogenetic tree of FtANR was clusteredinto 2 distinct groups monocot and dicot species FtANR ismost closely related toV vinifera while FtLAR ismost closelyrelated to F dibotrys Like the ANR protein LAR proteinswere separated into monocot (Hordeum vulgare and Oryzasativa) and dicot species
32 Gene Expression of FtANR and FtLAR during SproutDevelopment under Dark and Light Conditions in T8 and T10Transcript levels of ANR and LAR in seedlings of T8 andT10 were traced at 3 d intervals from 0 to 12 d after sowing(DAS) under dark and light conditions by real-time RT-PCRanalysis (Figure 3) Seeds were considered as 0DAS in thisstudy Generally FtANR expression levels were higher in T8than T10 under both conditions Specifically transcript levelin T8 under dark condition was higher than that under lightcondition although fluctuation pattern was observed in lightcondition In particular FtANR transcript levels were thehighest at 0 day and sharply declined in 3 and 6DAS for bothT8 and T10 cultivars under light condition However FtANRexpression was increased from 0 to 3DAS for T8 cultivarunder dark condition and remained relatively constant on
6 The Scientific World Journal
00
02
04
06
08
10
12
F L ST SD1 SD2 SD3 RT
FtANRG
ene e
xpre
ssio
n re
lativ
e to H3
(a)
T8T10
Gen
e exp
ress
ion
relat
ive t
o H3
02
04
FtLAR
00
06
08
F L ST SD1 SD2 SD3 RT
(b)
Figure 4 Relative expression levels of FtANR and FtLAR in different organs of F tataricum T8 and T10 cultivars F flowers ST stems Lleaves SD 1 2 3 seed stage 1 2 3 RT roots The height of each bar and the error bars show the mean and standard error respectively from3 independent measurements
6 9 and 12DAS Unlike FtANR different gene expressionpattern was observed in FtLARThe highest gene expressionlevel was observed at 12DAS in T10 which was nearly 5 timeshigher than that in T8 under light condition In additiona gradual increase in transcript levels was observed in T8under dark condition which is similar as FtANR under darkconditionTherefore in this study the transcription ofFtANRand FtLAR in T8 and T10 sprouts was unaffected by lightconditions
33 Gene Expression of FtANR and FtLAR among DifferentOrgans of T8 andT10 ThemRNA levels ofFtANR andFtLARamong different organs (flower leaf stem root and seeds instages 1 2 and 3) were shown in Figure 4 Unlike transcriptlevels during sprout development FtANR and FtLAR didnot show large variations in their expression patterns amongall the organs in T8 and T10 In some apple species LAR1expression was similar in the cortex and skin while ANRexpression levels were similar in apple skin flesh [30] Therelative levels of P trichocarpa LAR3 transcripts in the rootswere 2- and 3-fold higher than those in the stems andpetioles respectively [31] Bogs et al [10] reported that thetwo LAR genes involved in PA biosynthesis had differentpatterns of expression in grape seeds and skins which affectthe concentration and composition of PAs They suggestedthat V vinifera ANR may have a more significant role inPAs synthesis in grapevine leaves than LAR In additionGlycine max ANR1 is predominantly expressed in the seedcoat whereas GmANR2 is expressed at low levels in allorgans [29] Devic et al [32] reported that the expressionof the BAN gene is limited to the endothelium of immatureseeds in Arabidopsis whereas in Medicago its expressionis seen in young seeds flowers and leaves [9] Recently itwas reported that overexpression of LAR3 in Chinese whitepoplar (Populus tomentosa) leads to plantwide increase in PAlevels that is the highest in the roots [31] In our study we
found that the transcript levels of FtANR and FtLAR weresimilar among different organs According to the transcriptlevels observed in other plants it appears that FtANR andFtLAR are regulated differently compared to other plantspecies It is also possible that FtANR and FtLAR isoformsmay exist and these isoforms may contribute to catechin andepicatechin biosynthesis observed in this study in T8 and T10tartary buckwheat
34 Analysis of Catechin and Epicatechin during SeedlingDevelopment of Tartary Buckwheat The composition of cat-echin and epicatechin during seedling development from T8andT10was determined byHPLC (Figure 5) Epicatechinwasnot detected in T10 sprouts under light conditions whereasT8 sprouts had a high level of epicatechin (Figure 5(a)) Theepicatechin content of T8 was the highest (66mg gminus1 dryweight [DW]) at 12DAS under light conditionsThe oppositepattern was observed under dark conditions where theepicatechin content of T10 was higher than that of T8 Underdark conditions the epicatechin content of T10 was low inseeds and at 3DAS but reached higher levels on 6 9 and12DAS (eg 047mg gminus1DW Day 0 and 29mg gminus1DW on6DAS) while that of T8 increased from 0 (027mg gminus1DW)to 9DAS (279mg gminus1DW) (Figure 5(b)) Under light condi-tions the epicatechin content of T8 increased gradually from6 to 12DAS The epicatechin content of T8 was the highest(66mg gminus1DW) at 12DAS under light conditions while T10exhibited the highest content (31mg gminus1DW) at 6DASunderdark conditions
The catechin content of T8 under light and dark con-ditions was higher than that of T10 (Figures 5(c) and5(d)) The catechin content of T8 was the highest at 6DAS(196mg gminus1DW) and 12DAS (075mg gminus1DW) under darkand light conditions The catechin content of T10 was thehighest at 0DAS (055mg gminus1DW) Under dark conditions
The Scientific World Journal 7
Days after sowing
0
2
4
6
8
0 3 6 9 12
(mg
gminus1
dry
wt)
Epicatechin
(a)
0
1
2
3
4
0 3 6 9 12
(mg
gminus1
dry
wt)
Days after sowing
Epicatechin
(b)
0 3 6 9 12
0
02
04
06
08
1
(mg
gminus1
dry
wt)
Days after sowing
T8-LT10-L
Catechin
(c)
0 3 6 9 12
0
05
1
15
2
25
(mg
gminus1
dry
wt)
Days after sowing
T8-DT10-D
Catechin
(d)
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
and 340 amino acids with molecular weights of 369 and379 kDa respectively [24] FromBLAST analysis of theANR-deduced amino acid sequences FtANR was found to share73 77 82 78 and 80 identities with M truncatula(AAN77735) Malus times domestica (JN035299) Gossypiumhirsutum (EF187443) Camellia sinensis (AAT68773) andV vinifera (BAD89742) respectively FtANR was found tohave a conserved motif GXGXXA similar to that found inmonodehydroascorbate reductase (NADH) (SupplementaryFigure S1) [25]
Using RACE technology with partial sequences obtainedfromNGS data we isolated 1581-bp-long FtLAR cDNA (Gen-bank accession number KC404849) from flowers of F tatar-icum T10 which contained a 1176-bp ORF this gene encoded
a protein of 391 amino acids with a theoretical molecularmass of 433 kDa and a pI value of 527 Recently Ma et al[26] cloned and characterized the LAR gene from Fagopyrumdibotrys The amino acid sequences of FtLAR and FdLARwere found to differ by only 12 amino acids The FdLARtarget fusion peptide had a molecular weight of 66 kDa [26]which did not correspond to the theoretical molecular weightof FtLAR The amino acid sequence of FtLAR showed 6870 69 and 66 identity with V vinifera (AAZ82410)Prunus avium (ADY15310) Diospyros kaki (BAH89267) andMalus times domestica (AAX12186) respectively In particularFtLAR showed 97 homology with F dibotrys (JN793953)As reported previously [10 27] FtLAR had specific aminoacid motifs of ICCN and THD (Supplementary Figure S2)
The Scientific World Journal 5
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
6
7
0 3 6 9 12
FtANR
Days after sowing
(a)
0 3 6 9 12
FtANR
Gen
e exp
ress
ion
relat
ive t
o H3
Days after sowing
0
5
10
15
20
25
(b)
0 3 6 9 12
T8-LT10-L
FtLAR
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
0
1
2
3
4
5
(c)
0 3 6 9 12
T8-DT10-D
FtLAR
0
1
2
3
4
Days after sowing
Gen
e exp
ress
ion
relat
ive t
o H3
(d)
Figure 3 Relative expression levels of FtANR and FtLAR in seedling of F tataricumT8 and T10 cultivars L-light condition D-dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
The subcellular targeting of FtANRwas predicted to be in thecytosol and the chloroplast whereas FtLAR was predicted tobe localized to the cytosolThis result was in accordance withthat of a previous study [11]Winkel [28] suggested that the PApathway exists as ametabolic channel associated with cellularmembranes
To determine the relationship of putative FtANR andFtLAR proteins with other plant ANRs and LARs weperformed phylogenetic analysis (Figure 2) As describedpreviously [29] a phylogenetic tree of FtANR was clusteredinto 2 distinct groups monocot and dicot species FtANR ismost closely related toV vinifera while FtLAR ismost closelyrelated to F dibotrys Like the ANR protein LAR proteinswere separated into monocot (Hordeum vulgare and Oryzasativa) and dicot species
32 Gene Expression of FtANR and FtLAR during SproutDevelopment under Dark and Light Conditions in T8 and T10Transcript levels of ANR and LAR in seedlings of T8 andT10 were traced at 3 d intervals from 0 to 12 d after sowing(DAS) under dark and light conditions by real-time RT-PCRanalysis (Figure 3) Seeds were considered as 0DAS in thisstudy Generally FtANR expression levels were higher in T8than T10 under both conditions Specifically transcript levelin T8 under dark condition was higher than that under lightcondition although fluctuation pattern was observed in lightcondition In particular FtANR transcript levels were thehighest at 0 day and sharply declined in 3 and 6DAS for bothT8 and T10 cultivars under light condition However FtANRexpression was increased from 0 to 3DAS for T8 cultivarunder dark condition and remained relatively constant on
6 The Scientific World Journal
00
02
04
06
08
10
12
F L ST SD1 SD2 SD3 RT
FtANRG
ene e
xpre
ssio
n re
lativ
e to H3
(a)
T8T10
Gen
e exp
ress
ion
relat
ive t
o H3
02
04
FtLAR
00
06
08
F L ST SD1 SD2 SD3 RT
(b)
Figure 4 Relative expression levels of FtANR and FtLAR in different organs of F tataricum T8 and T10 cultivars F flowers ST stems Lleaves SD 1 2 3 seed stage 1 2 3 RT roots The height of each bar and the error bars show the mean and standard error respectively from3 independent measurements
6 9 and 12DAS Unlike FtANR different gene expressionpattern was observed in FtLARThe highest gene expressionlevel was observed at 12DAS in T10 which was nearly 5 timeshigher than that in T8 under light condition In additiona gradual increase in transcript levels was observed in T8under dark condition which is similar as FtANR under darkconditionTherefore in this study the transcription ofFtANRand FtLAR in T8 and T10 sprouts was unaffected by lightconditions
33 Gene Expression of FtANR and FtLAR among DifferentOrgans of T8 andT10 ThemRNA levels ofFtANR andFtLARamong different organs (flower leaf stem root and seeds instages 1 2 and 3) were shown in Figure 4 Unlike transcriptlevels during sprout development FtANR and FtLAR didnot show large variations in their expression patterns amongall the organs in T8 and T10 In some apple species LAR1expression was similar in the cortex and skin while ANRexpression levels were similar in apple skin flesh [30] Therelative levels of P trichocarpa LAR3 transcripts in the rootswere 2- and 3-fold higher than those in the stems andpetioles respectively [31] Bogs et al [10] reported that thetwo LAR genes involved in PA biosynthesis had differentpatterns of expression in grape seeds and skins which affectthe concentration and composition of PAs They suggestedthat V vinifera ANR may have a more significant role inPAs synthesis in grapevine leaves than LAR In additionGlycine max ANR1 is predominantly expressed in the seedcoat whereas GmANR2 is expressed at low levels in allorgans [29] Devic et al [32] reported that the expressionof the BAN gene is limited to the endothelium of immatureseeds in Arabidopsis whereas in Medicago its expressionis seen in young seeds flowers and leaves [9] Recently itwas reported that overexpression of LAR3 in Chinese whitepoplar (Populus tomentosa) leads to plantwide increase in PAlevels that is the highest in the roots [31] In our study we
found that the transcript levels of FtANR and FtLAR weresimilar among different organs According to the transcriptlevels observed in other plants it appears that FtANR andFtLAR are regulated differently compared to other plantspecies It is also possible that FtANR and FtLAR isoformsmay exist and these isoforms may contribute to catechin andepicatechin biosynthesis observed in this study in T8 and T10tartary buckwheat
34 Analysis of Catechin and Epicatechin during SeedlingDevelopment of Tartary Buckwheat The composition of cat-echin and epicatechin during seedling development from T8andT10was determined byHPLC (Figure 5) Epicatechinwasnot detected in T10 sprouts under light conditions whereasT8 sprouts had a high level of epicatechin (Figure 5(a)) Theepicatechin content of T8 was the highest (66mg gminus1 dryweight [DW]) at 12DAS under light conditionsThe oppositepattern was observed under dark conditions where theepicatechin content of T10 was higher than that of T8 Underdark conditions the epicatechin content of T10 was low inseeds and at 3DAS but reached higher levels on 6 9 and12DAS (eg 047mg gminus1DW Day 0 and 29mg gminus1DW on6DAS) while that of T8 increased from 0 (027mg gminus1DW)to 9DAS (279mg gminus1DW) (Figure 5(b)) Under light condi-tions the epicatechin content of T8 increased gradually from6 to 12DAS The epicatechin content of T8 was the highest(66mg gminus1DW) at 12DAS under light conditions while T10exhibited the highest content (31mg gminus1DW) at 6DASunderdark conditions
The catechin content of T8 under light and dark con-ditions was higher than that of T10 (Figures 5(c) and5(d)) The catechin content of T8 was the highest at 6DAS(196mg gminus1DW) and 12DAS (075mg gminus1DW) under darkand light conditions The catechin content of T10 was thehighest at 0DAS (055mg gminus1DW) Under dark conditions
The Scientific World Journal 7
Days after sowing
0
2
4
6
8
0 3 6 9 12
(mg
gminus1
dry
wt)
Epicatechin
(a)
0
1
2
3
4
0 3 6 9 12
(mg
gminus1
dry
wt)
Days after sowing
Epicatechin
(b)
0 3 6 9 12
0
02
04
06
08
1
(mg
gminus1
dry
wt)
Days after sowing
T8-LT10-L
Catechin
(c)
0 3 6 9 12
0
05
1
15
2
25
(mg
gminus1
dry
wt)
Days after sowing
T8-DT10-D
Catechin
(d)
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
Figure 3 Relative expression levels of FtANR and FtLAR in seedling of F tataricumT8 and T10 cultivars L-light condition D-dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
The subcellular targeting of FtANRwas predicted to be in thecytosol and the chloroplast whereas FtLAR was predicted tobe localized to the cytosolThis result was in accordance withthat of a previous study [11]Winkel [28] suggested that the PApathway exists as ametabolic channel associated with cellularmembranes
To determine the relationship of putative FtANR andFtLAR proteins with other plant ANRs and LARs weperformed phylogenetic analysis (Figure 2) As describedpreviously [29] a phylogenetic tree of FtANR was clusteredinto 2 distinct groups monocot and dicot species FtANR ismost closely related toV vinifera while FtLAR ismost closelyrelated to F dibotrys Like the ANR protein LAR proteinswere separated into monocot (Hordeum vulgare and Oryzasativa) and dicot species
32 Gene Expression of FtANR and FtLAR during SproutDevelopment under Dark and Light Conditions in T8 and T10Transcript levels of ANR and LAR in seedlings of T8 andT10 were traced at 3 d intervals from 0 to 12 d after sowing(DAS) under dark and light conditions by real-time RT-PCRanalysis (Figure 3) Seeds were considered as 0DAS in thisstudy Generally FtANR expression levels were higher in T8than T10 under both conditions Specifically transcript levelin T8 under dark condition was higher than that under lightcondition although fluctuation pattern was observed in lightcondition In particular FtANR transcript levels were thehighest at 0 day and sharply declined in 3 and 6DAS for bothT8 and T10 cultivars under light condition However FtANRexpression was increased from 0 to 3DAS for T8 cultivarunder dark condition and remained relatively constant on
6 The Scientific World Journal
00
02
04
06
08
10
12
F L ST SD1 SD2 SD3 RT
FtANRG
ene e
xpre
ssio
n re
lativ
e to H3
(a)
T8T10
Gen
e exp
ress
ion
relat
ive t
o H3
02
04
FtLAR
00
06
08
F L ST SD1 SD2 SD3 RT
(b)
Figure 4 Relative expression levels of FtANR and FtLAR in different organs of F tataricum T8 and T10 cultivars F flowers ST stems Lleaves SD 1 2 3 seed stage 1 2 3 RT roots The height of each bar and the error bars show the mean and standard error respectively from3 independent measurements
6 9 and 12DAS Unlike FtANR different gene expressionpattern was observed in FtLARThe highest gene expressionlevel was observed at 12DAS in T10 which was nearly 5 timeshigher than that in T8 under light condition In additiona gradual increase in transcript levels was observed in T8under dark condition which is similar as FtANR under darkconditionTherefore in this study the transcription ofFtANRand FtLAR in T8 and T10 sprouts was unaffected by lightconditions
33 Gene Expression of FtANR and FtLAR among DifferentOrgans of T8 andT10 ThemRNA levels ofFtANR andFtLARamong different organs (flower leaf stem root and seeds instages 1 2 and 3) were shown in Figure 4 Unlike transcriptlevels during sprout development FtANR and FtLAR didnot show large variations in their expression patterns amongall the organs in T8 and T10 In some apple species LAR1expression was similar in the cortex and skin while ANRexpression levels were similar in apple skin flesh [30] Therelative levels of P trichocarpa LAR3 transcripts in the rootswere 2- and 3-fold higher than those in the stems andpetioles respectively [31] Bogs et al [10] reported that thetwo LAR genes involved in PA biosynthesis had differentpatterns of expression in grape seeds and skins which affectthe concentration and composition of PAs They suggestedthat V vinifera ANR may have a more significant role inPAs synthesis in grapevine leaves than LAR In additionGlycine max ANR1 is predominantly expressed in the seedcoat whereas GmANR2 is expressed at low levels in allorgans [29] Devic et al [32] reported that the expressionof the BAN gene is limited to the endothelium of immatureseeds in Arabidopsis whereas in Medicago its expressionis seen in young seeds flowers and leaves [9] Recently itwas reported that overexpression of LAR3 in Chinese whitepoplar (Populus tomentosa) leads to plantwide increase in PAlevels that is the highest in the roots [31] In our study we
found that the transcript levels of FtANR and FtLAR weresimilar among different organs According to the transcriptlevels observed in other plants it appears that FtANR andFtLAR are regulated differently compared to other plantspecies It is also possible that FtANR and FtLAR isoformsmay exist and these isoforms may contribute to catechin andepicatechin biosynthesis observed in this study in T8 and T10tartary buckwheat
34 Analysis of Catechin and Epicatechin during SeedlingDevelopment of Tartary Buckwheat The composition of cat-echin and epicatechin during seedling development from T8andT10was determined byHPLC (Figure 5) Epicatechinwasnot detected in T10 sprouts under light conditions whereasT8 sprouts had a high level of epicatechin (Figure 5(a)) Theepicatechin content of T8 was the highest (66mg gminus1 dryweight [DW]) at 12DAS under light conditionsThe oppositepattern was observed under dark conditions where theepicatechin content of T10 was higher than that of T8 Underdark conditions the epicatechin content of T10 was low inseeds and at 3DAS but reached higher levels on 6 9 and12DAS (eg 047mg gminus1DW Day 0 and 29mg gminus1DW on6DAS) while that of T8 increased from 0 (027mg gminus1DW)to 9DAS (279mg gminus1DW) (Figure 5(b)) Under light condi-tions the epicatechin content of T8 increased gradually from6 to 12DAS The epicatechin content of T8 was the highest(66mg gminus1DW) at 12DAS under light conditions while T10exhibited the highest content (31mg gminus1DW) at 6DASunderdark conditions
The catechin content of T8 under light and dark con-ditions was higher than that of T10 (Figures 5(c) and5(d)) The catechin content of T8 was the highest at 6DAS(196mg gminus1DW) and 12DAS (075mg gminus1DW) under darkand light conditions The catechin content of T10 was thehighest at 0DAS (055mg gminus1DW) Under dark conditions
The Scientific World Journal 7
Days after sowing
0
2
4
6
8
0 3 6 9 12
(mg
gminus1
dry
wt)
Epicatechin
(a)
0
1
2
3
4
0 3 6 9 12
(mg
gminus1
dry
wt)
Days after sowing
Epicatechin
(b)
0 3 6 9 12
0
02
04
06
08
1
(mg
gminus1
dry
wt)
Days after sowing
T8-LT10-L
Catechin
(c)
0 3 6 9 12
0
05
1
15
2
25
(mg
gminus1
dry
wt)
Days after sowing
T8-DT10-D
Catechin
(d)
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
Figure 4 Relative expression levels of FtANR and FtLAR in different organs of F tataricum T8 and T10 cultivars F flowers ST stems Lleaves SD 1 2 3 seed stage 1 2 3 RT roots The height of each bar and the error bars show the mean and standard error respectively from3 independent measurements
6 9 and 12DAS Unlike FtANR different gene expressionpattern was observed in FtLARThe highest gene expressionlevel was observed at 12DAS in T10 which was nearly 5 timeshigher than that in T8 under light condition In additiona gradual increase in transcript levels was observed in T8under dark condition which is similar as FtANR under darkconditionTherefore in this study the transcription ofFtANRand FtLAR in T8 and T10 sprouts was unaffected by lightconditions
33 Gene Expression of FtANR and FtLAR among DifferentOrgans of T8 andT10 ThemRNA levels ofFtANR andFtLARamong different organs (flower leaf stem root and seeds instages 1 2 and 3) were shown in Figure 4 Unlike transcriptlevels during sprout development FtANR and FtLAR didnot show large variations in their expression patterns amongall the organs in T8 and T10 In some apple species LAR1expression was similar in the cortex and skin while ANRexpression levels were similar in apple skin flesh [30] Therelative levels of P trichocarpa LAR3 transcripts in the rootswere 2- and 3-fold higher than those in the stems andpetioles respectively [31] Bogs et al [10] reported that thetwo LAR genes involved in PA biosynthesis had differentpatterns of expression in grape seeds and skins which affectthe concentration and composition of PAs They suggestedthat V vinifera ANR may have a more significant role inPAs synthesis in grapevine leaves than LAR In additionGlycine max ANR1 is predominantly expressed in the seedcoat whereas GmANR2 is expressed at low levels in allorgans [29] Devic et al [32] reported that the expressionof the BAN gene is limited to the endothelium of immatureseeds in Arabidopsis whereas in Medicago its expressionis seen in young seeds flowers and leaves [9] Recently itwas reported that overexpression of LAR3 in Chinese whitepoplar (Populus tomentosa) leads to plantwide increase in PAlevels that is the highest in the roots [31] In our study we
found that the transcript levels of FtANR and FtLAR weresimilar among different organs According to the transcriptlevels observed in other plants it appears that FtANR andFtLAR are regulated differently compared to other plantspecies It is also possible that FtANR and FtLAR isoformsmay exist and these isoforms may contribute to catechin andepicatechin biosynthesis observed in this study in T8 and T10tartary buckwheat
34 Analysis of Catechin and Epicatechin during SeedlingDevelopment of Tartary Buckwheat The composition of cat-echin and epicatechin during seedling development from T8andT10was determined byHPLC (Figure 5) Epicatechinwasnot detected in T10 sprouts under light conditions whereasT8 sprouts had a high level of epicatechin (Figure 5(a)) Theepicatechin content of T8 was the highest (66mg gminus1 dryweight [DW]) at 12DAS under light conditionsThe oppositepattern was observed under dark conditions where theepicatechin content of T10 was higher than that of T8 Underdark conditions the epicatechin content of T10 was low inseeds and at 3DAS but reached higher levels on 6 9 and12DAS (eg 047mg gminus1DW Day 0 and 29mg gminus1DW on6DAS) while that of T8 increased from 0 (027mg gminus1DW)to 9DAS (279mg gminus1DW) (Figure 5(b)) Under light condi-tions the epicatechin content of T8 increased gradually from6 to 12DAS The epicatechin content of T8 was the highest(66mg gminus1DW) at 12DAS under light conditions while T10exhibited the highest content (31mg gminus1DW) at 6DASunderdark conditions
The catechin content of T8 under light and dark con-ditions was higher than that of T10 (Figures 5(c) and5(d)) The catechin content of T8 was the highest at 6DAS(196mg gminus1DW) and 12DAS (075mg gminus1DW) under darkand light conditions The catechin content of T10 was thehighest at 0DAS (055mg gminus1DW) Under dark conditions
The Scientific World Journal 7
Days after sowing
0
2
4
6
8
0 3 6 9 12
(mg
gminus1
dry
wt)
Epicatechin
(a)
0
1
2
3
4
0 3 6 9 12
(mg
gminus1
dry
wt)
Days after sowing
Epicatechin
(b)
0 3 6 9 12
0
02
04
06
08
1
(mg
gminus1
dry
wt)
Days after sowing
T8-LT10-L
Catechin
(c)
0 3 6 9 12
0
05
1
15
2
25
(mg
gminus1
dry
wt)
Days after sowing
T8-DT10-D
Catechin
(d)
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
Figure 5 Epicatechin and catechin content in seedling of F tataricum T8 and T10 cultivars (a) (c) light condition (b) (d) dark conditionThe height of each bar and the error bars show the mean and standard error respectively from 3 independent measurements
the catechin content of T8 increased from 0 (017mg gminus1DW)to 6DAS (196mg gminus1DW) and then decreased to the levelssimilar to that of Day 0 On the other hand under lightconditions the catechin content of T10 decreased graduallyfrom0 (055mg gminus1DW) to 6DAS (0015mg gminus1DW)Underlight conditions the catechin content of T8was 37- and 9-foldhigher at 6 and 12DAS respectively than in T10
Unlike catechin analysis epicatechin content was affectedby light in T8 only whereas T10 remained unaffected by lightIn general light acts as an essential stimulus and also modu-lates the intensity of the pigment by affecting the regulatoryand structural genes involved in anthocyanin biosynthesis[33] However Bakhshi and Arakawa [34] described thatthe content of phenolic acids anthocyanin and flavonolsincreased rapidly by irradiation whereas flavanols procyani-dins and dihydrochalcones did not change in either matureor in ripe apple fruits Our results suggest that a response of
catechin and epicatechin content to lightmight depend on thetartary buckwheat cultivar
35 Analysis of Catechin and Epicatechin in Different OrgansCatechin and epicatechin contents were analyzed from differ-ent organs from T8 and T10 (ie flowers stems leave rootsand seeds at stages 1 2 and 3) by HPLC (Figure 6) Similarto the results seen in seedling development epicatechincontent was higher than the catechin content in both T8and T10 In T10 catechin was found mostly in the flowers(268mg gminus1DW) and roots (275mg gminus1DW) whereas inT8 higher amounts were found in leaves (116mg gminus1DW)and seeds at stage 1 (13mg gminus1DW) It is interesting thatcatechin content in T10 was the highest in the roots T10 wildroots in the soil and hairy roots are red in colorThese resultsindicate that catechin content is correlated with color pig-ment The lowest catechin content was found in the
8 The Scientific World Journal
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
Epicatechin
0
3
6
9
12
15
18
(a)
F L ST SD1 SD2 SD3 RT
(mg
gminus1
dry
wt)
T8T10
Catechin
0
05
1
15
2
25
3
(b)
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
Figure 6 Epicatechin and catechin content in different organs of F tataricum T8 and T10 cultivars The height of each bar and the error barsshow the mean and standard error respectively from 3 independent measurements
stems (011mg gminus1DW) of T8 and in seeds at stage 2(04mg gminus1DW) of T10 The catechin content of seedsdecreased gradually from stage 1 (13mg gminus1DW) to stage 3(017mg gminus1DW)
The flowers of T8 and T10 contained the highest levelsof epicatechin (155 and 101mg gminus1DW resp) whereas thelowest amount of epicatechin was in the seeds at stage 3 in T8(03mg gminus1DW) However in all cases T8 exhibited higheramounts of epicatechin content than T10 and in fact epicate-chin was not detected in the stems of T10 The epicatechincontent in T8 flowers was 22-fold higher than that in theirleaves Similarly in T10 flowers the level of epicatechin was14-fold higher than in leaves of the same cultivar Li etal [20] reported that in common buckwheat the catechinhydrate and epicatechin content in flowers is higher thanin other organs The results from the present study indicatethat catechin and epicatechin accumulated abundantly in theorgans of both cultivars and were found more abundantly inthe flowers thus supporting a previous report by Uddin etal [35] wherein the flower of the common buckwheat wasfound to containmanymore phenolic compounds than otherplant parts Watanabe [23] reported that antioxidant activityof catechins isolated from common buckwheat groats wassuperior to that of rutin which is known as an antioxidantin buckwheat at the same concentration Further study isrequired for antioxidant activity of catechins and epicatechinisolated from tartary buckwheat cultivars T8 and T10
4 Conclusion
Our study shows that epicatechin content is affected by lightin both T8 and T10 We observed that catechin and epicate-chin content varies in cultivars The highest amount of epi-catechin was observed in the flowers of both tartary buck-wheat cultivars The highest amount of epicatechin wasobserved in the flowers of both tartary buckwheat culti-vars we studied These data suggest that the two types oftartary buckwheat sprouts may have different mechanisms
for catechin and epicatechin biosynthesis which vary underlight and dark conditions The data obtained in our studyprovides evidence that cultivar differences as well as differ-ences in environmental conditions can differentially regulateflavonoid biosynthesis
The authors declare that there is no conflict of interestsregarding the publication of this paper
Authorsrsquo Contribution
SangUnPark andKwangHyun Jho designed the experimentsand analyzed the data Yeon Bok Kim Aye Aye Thwe YeJiKimXiaohua Li JinWoongCho PhunBumParkMariadhasValan Arasu Naif Abdullah Al-Dhabi Sun-Ju Kim andTastsuro Suzuki wrote the paper performed the experimentand analyzed the data
Acknowledgment
This study was financially supported by research fund ofSahmyook University in 2012
References
[1] R A Dixon D-Y Xie and S B Sharma ldquoProanthocyanidinsmdasha final frontier in flavonoid researchrdquoNew Phytologist vol 165no 1 pp 9ndash28 2005
[2] L Lepiniec I Debeaujon J-M Routaboul et al ldquoGeneticsand biochemistry of seed flavonoidsrdquo Annual Review of PlantBiology vol 57 pp 405ndash430 2006
The Scientific World Journal 9
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
[3] T T C Yang and M W L Koo ldquoInhibitory effect of Chinesegreen tea on endothelial cell-induced LDL oxidationrdquo Athero-sclerosis vol 148 no 1 pp 67ndash73 2000
[4] A Luximon-Ramma V S Neergheen T Bahorun et alldquoAssessment of the polyphenolic composition of the organicextracts of Mauritian black teas a potential contributor to theirantioxidant functionsrdquo BioFactors vol 27 no 1ndash4 pp 79ndash912006
[5] D L Bemis A E Katz andR Buttyan ldquoClinical trials of naturalproducts as chemopreventive agents for prostate cancerrdquo ExpertOpinion on Investigational Drugs vol 15 no 10 pp 1191ndash12002006
[6] P M Aron and J A Kennedy ldquoFlavan-3-ols nature occurrenceand biological activityrdquoMolecular Nutrition and Food Researchvol 52 no 1 pp 79ndash104 2008
[7] B Winkel-Shirley ldquoFlavonoid biosynthesis A colorful modelfor genetics biochemistry cell biology and biotechnologyrdquoPlant Physiology vol 126 no 2 pp 485ndash493 2001
[8] M Gargouri C Manigand C Mauge et al ldquoStructure and epi-merase activity of anthocyanidin reductase from Vitis viniferardquoActa Crystallographica Section D vol 65 no 9 pp 989ndash10002009
[9] D-Y Xie S B SharmaN L Paiva D Ferreira and R ADixonldquoRole of anthocyanidin reductase encoded by BANYULS inplant flavonoid biosynthesisrdquo Science vol 299 no 5605 pp396ndash399 2003
[10] J Bogs M O Downey J S Harvey A R Ashton G J Tannerand S P Robinson ldquoProanthocyanidin synthesis and expres-sion of genes encoding leucoanthocyanidin reductase andanthocyanidin reductase in developing grape berries andgrapevine leavesrdquo Plant Physiology vol 139 no 2 pp 652ndash6632005
[11] Y Pang G J Peel E Wright Z Wang and R A Dixon ldquoEarlysteps in proanthocyanidin biosynthesis in the model legumeMedicago truncatulardquo Plant Physiology vol 145 no 3 pp 601ndash615 2007
[12] N Kovinich A Saleem J T Arnason and B Miki ldquoIdentifica-tion of two anthocyanidin reductase genes and three red-brownsoybean accessions with reduced anthocyanidin reductase 1mRNA activity and seed coat proanthocyanidin amountsrdquoJournal of Agricultural and Food Chemistry vol 60 no 2 pp574ndash584 2012
[13] Y Pang I S Abeysinghe J He et al ldquoFunctional characteriza-tion of proanthocyanidin pathway enzymes from tea and theirapplication for metabolic engineeringrdquo Plant Physiology vol161 no 3 pp 1103ndash1116 2013
[14] S Gagne S Lacampagne O Claisse and L Geny ldquoLeucoantho-cyanidin reductase and anthocyanidin reductase gene expres-sion and activity in flowers young berries and skins of Vitisvinifera L cv Cabernet-Sauvignon during developmentrdquo PlantPhysiology and Biochemistry vol 47 no 4 pp 282ndash290 2009
[15] X Guo K Zhu H Zhang and H Yao ldquoPurification andcharacterization of the antitumor protein from Chinese tar-tary buckwheat (Fagopyrum tataricum Gaertn) water-solubleextractsrdquo Journal of Agricultural and FoodChemistry vol 55 no17 pp 6958ndash6961 2007
[16] Y Yao F Shan J Bian F Chen M Wang and G Ren ldquoD-chiro-inositol-enriched tartary buckwheat bran extract lowersthe blood glucose level in KK-Ay micerdquo Journal of Agriculturaland Food Chemistry vol 56 no 21 pp 10027ndash10031 2008
[17] C-L Liu Y-S Chen J-H Yang and B-H Chiang ldquoAntioxi-dant activity of tartary (Fagopyrum tataricum (L) gaertn) and
common (Fagopyrum esculentummoench) buckwheat sproutsrdquoJournal of Agricultural and Food Chemistry vol 56 no 1 pp173ndash178 2008
[18] S-J Kim T Maeda M Z I Sarker et al ldquoIdentification ofanthocyanins in the sprouts of buckwheatrdquo Journal of Agricul-tural and Food Chemistry vol 55 no 15 pp 6314ndash6318 2007
[19] X Li A A Thwe N I Park T Suzuki S J Kim and SU Park ldquoAccumulation of phenylpropanoids and correlatedgene expression during the development of tatary buckwheatsproutsrdquo Journal of Agricultural and Food Chemistry vol 60 no22 pp 5629ndash5635 2012
[20] X Li N-I Park H Xu S-H Woo C H Park and S U ParkldquoDifferential expression of flavonoid biosynthesis genes andaccumulation of phenolic compounds in common buckwheat(Fagopyrum esculentum)rdquo Journal of Agricultural and FoodChemistry vol 58 no 23 pp 12176ndash12181 2010
[21] N-I Park X Li T Suzuki et al ldquoDifferential expression ofanthocyanin biosynthetic genes and anthocyanin accumulationin tartary buckwheat cultivars lsquoHokkai T8rsquo and lsquoHokkai T10rsquordquoJournal of Agricultural and Food Chemistry vol 59 no 6 pp2356ndash2361 2011
[22] G Gambino I Perrone and I Gribaudo ldquoA rapid and effectivemethod for RNA extraction from different tissues of grapevineand other woody plantsrdquo Phytochemical Analysis vol 19 no 6pp 520ndash525 2008
[23] M Watanabe ldquoCatechins as antioxidants from buckwheat(Fagopyrum esculentum Moench) groatsrdquo Journal of Agricul-tural and Food Chemistry vol 46 no 3 pp 839ndash845 1998
[24] D-Y Xie S B Sharma and R A Dixon ldquoAnthocyanidinreductases fromMedicago truncatula andArabidopsis thalianardquoArchives of Biochemistry and Biophysics vol 422 no 1 pp 91ndash102 2004
[25] S S Murthy and B A Zilinskas ldquoMolecular cloning and char-acterization of a cDNA encoding pea monodehydroascorbatereductaserdquo Journal of Biological Chemistry vol 269 no 49 pp31129ndash31133 1994
[26] J Ma B Wang Y Dai S Z Sui and M Y Li ldquoCloning andexpression analysis of leucoanthocyanidin reductase gene inFagopyrum dibotrysrdquo Acta Pharmacology Sinica vol 47 no 7pp 953ndash961 2012
[27] G J Tanner K T Francki S Abrahams J M Watson P JLarkin and A R Ashton ldquoProanthocyanidin biosynthesis inplants Purification of legume leucoanthocyanidin reductaseandmolecular cloning of its cDNArdquo Journal of Biological Chem-istry vol 278 no 34 pp 31647ndash31656 2003
[28] B S JWinkel ldquoMetabolic channeling in plantsrdquoAnnual Reviewof Plant Biology vol 55 pp 85ndash107 2004
[29] L Yuan L Wang Z Han et al ldquoMolecular cloning and cha-racterization of PtrLAR3 a gene encoding leucoanthocyanidinreductase fromPopulus trichocarpa and its constitutive expres-sion enhances fungal resistance in transgenic plantsrdquo Journal ofExperimental Botany vol 63 no 7 pp 2513ndash2524 2012
[30] R A Henry-Kirk T K McGhie C M Andre R P Hellensand A C Allan ldquoTranscriptional analysis of apple fruit proan-thocaynidin biosynthesisrdquo Journal of Experimental Botany vol63 no 15 pp 5437ndash5450 2012
[31] N Kovinich A Saleem J T Arnason and B Miki ldquoCombinedanalysis of transcriptome and metabolite data reveals extensivedifferences between black and brown nearly-isogenic soybean(Glycine max) seed coats enabling the identification of pigmentisogenesrdquo BMC Genomics vol 12 article 381 2011
10 The Scientific World Journal
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013
[32] M Devic J Guilleminot I Debeaujon et al ldquoThe BANYULSgene encodes a DFR-like protein and is a marker of early seedcoat developmentrdquo Plant Journal vol 19 no 4 pp 387ndash3981999
[33] I S Sheoran T Dumonceaux R Datla and V K SawhneyldquoAnthocyanin accumulation in the hypocotyl of an ABA-over producing male-sterile tomato (Lycopersicon esculentum)mutantrdquo Physiologia Plantarum vol 127 no 4 pp 681ndash6892006
[34] D Bakhshi andOArakawa ldquoInduction of phenolic compoundsbiosynthesis with light irradiation in the flesh of red and yellowapplesrdquo Journal of AppliedHorticulture vol 8 pp 101ndash104 2006
[35] M R Uddin X Li W T Park et al ldquoPhenolic compound con-tent in different organs of Korean common buckwheat culti-varsrdquo Asian Journal of Chemistry vol 25 pp 424ndash426 2013