1 23 Journal of Insect Conservation An international journal devoted to the conservation of insects and related invertebrates ISSN 1366-638X Volume 18 Number 5 J Insect Conserv (2014) 18:757-769 DOI 10.1007/s10841-014-9677-x Diversity of grass-dwelling spiders (Arachnida: Araneae) in calcareous fens of the Coastal Lowland, Latvia Maija Štokmane & Voldemārs Spuņģis
Transcript
1 23
Journal of Insect ConservationAn international journal devoted tothe conservation of insects and relatedinvertebrates ISSN 1366-638XVolume 18Number 5 J Insect Conserv (2014) 18:757-769DOI 10.1007/s10841-014-9677-x
Diversity of grass-dwelling spiders(Arachnida: Araneae) in calcareous fens ofthe Coastal Lowland, Latvia
Maija Štokmane & Voldemārs Spuņģis
1 23
Your article is protected by copyright and
all rights are held exclusively by Springer
International Publishing Switzerland. This e-
offprint is for personal use only and shall not
be self-archived in electronic repositories. If
you wish to self-archive your article, please
use the accepted manuscript version for
posting on your own website. You may
further deposit the accepted manuscript
version in any repository, provided it is only
made publicly available 12 months after
official publication or later and provided
acknowledgement is given to the original
source of publication and a link is inserted
to the published article on Springer's
website. The link must be accompanied by
the following text: "The final publication is
available at link.springer.com”.
ORIGINAL PAPER
Diversity of grass-dwelling spiders (Arachnida: Araneae) incalcareous fens of the Coastal Lowland, Latvia
Maija Štokmane · Voldemārs Spuņģis
Received: 27 August 2013 / Accepted: 26 July 2014 / Published online: 9 August 2014
Abstract Calcareous fens have a high conservation pri-
ority in the European Union. They are very unique, very
sensitive and very rare habitats in Latvia as well as in many
other European countries. Because of their rarity, many
organisms living in calcareous fens are nowadays consid-
ered threatened. The same is applied to spiders. Spiders
have been suggested as good biodiversity indicators
because they have numerous direct and indirect relation-
ships with other organisms. Only few investigations have
been carried out on fen inhabiting spiders. The knowledge
of grass-dwelling spiders is especially lacking. Thus the
aim of this study was to evaluate grass-dwelling spider
diversity in several calcareous fens of Latvia, compare
these fens and to find out the habitat features that might
affect grass-layer inhabiting spiders. The research was
carried out in eight calcareous fens located in the Coastal
Lowland of Latvia. A total of 760 spiders from nine fam-
ilies and 20 species were collected with a sweep net. Two
of the most abundant spider species were Dolomedes fim-briatus (Clerck, 1757) and Tibellus maritimus (Menge,
1875). Diversity indices suggested that the grass-dwelling
spider community consists of few abundant species and
numerous rare species. Correlation analysis as well as DCA
showed that plant species diversity did not significantly
affect spider species richness and diversity, although at the
same time there were a lot of significant associations
between spiders and individual plant species. It was shown
that various spider species responded very differently to the
presence of particular plant species, and thus habitat
structural heterogeneity emerges to be an important factor
influencing the grass-dwelling spider diversity and com-
munity structure. Correlation analysis also revealed that
spider abundance was negatively affected by the vegetation
height and wind speed.
Keywords Araneae · Grass-dwelling spiders ·
Species diversity · Calcareous fens
Introduction
Fen ecosystems were previously common in temperate
Europe (Van Diggelen et al. 2006). Nowadays, however,
they are very rare because during the past few centuries
almost all fens have been selectively drained and changed
into low-productive meadows and pastures (Sefferova et al.
2008). Furthermore, large-scale impacts such as eutrophi-
cation, acidification, habitat fragmentation and climate
change have resulted in the degradation of many fens
(Saunders et al. 1991; Chapman et al. 2003). As a result,
many organisms living in these habitats are nowadays
considered threatened (Koponen 2003).
Calcareous fens are considered a subtype of rich fen
habitats. This group of fens is very unique ecosystems
because they support a number of plant and animal species
that are specially adapted to conditions of high pH and high
calcium concentration—so-called “calcicoles” or calcium-
loving species (Rydin and Jeglum 2006). Fen habitats are
ecologically valuable also for a number of other reasons,
for example, they provide multiple ecosystem services such
as water retention, drought prevention and nutrient removal
as well as they are important carbon sinks and stores
(Gorham 1991; Zedler and Kercher 2005). Fens also have
important scientific value because their peat deposits might
M. Štokmane (&) · V. SpuņģisDepartment of Zoology and Animal Ecology, Faculty of Biology,University of Latvia, Kronvalda Boulevard 4, Riga LV-1586,Latviae-mail: [email protected]
123
J Insect Conserv (2014) 18:757–769
DOI 10.1007/s10841-014-9677-x
Author's personal copy
contain paleoecological information on past vegetation and
climate (Barber 1993). Fens have recreational importance
as well. Because of their rarity, calcareous fens have a high
conservation priority in the European Union—they belong
to natural habitat types of community interest listed in
Annex I of the EU Habitats Directive (EC 1992). Calcar-
eous fens are also very rare in Latvia—they occupy only
about 0.015 % of the whole territory. The largest calcare-
ous fens are situated in the western part of Latvia,
especially in the Coastal Lowland (Aunins et al. 2010).
The ecology of the spider fauna of calcareous fens is
poorly known because, not only in Latvia, but also in many
other countries studies on fen inhabiting spiders are almost
completely lacking. Therefore, taking into account that
calcareous fens are becoming increasingly rare in Europe,
it is important to obtain as much data on fen species as
possible. Besides, such studies are necessary for the
assessment of the conservation value of these unique
habitats. Spiders are among the most abundant insectivo-
rous predators of terrestrial ecosystems (Nyffeler and Benz
1987; Wise 1995) and one of the most diverse arthropod
orders, with over 44,000 species (Platnick 2013). Spiders
are playing an important role in the balance of nature
because they occupy a strategic functional position in ter-
restrial food webs—they act as regulating agents in
terrestrial arthropod communities and are important food
source for higher organisms (Whitcomb 1974; Young and
Edwards 1990; Nyffeler et al. 1994; Wise 1995; Oxbrough
et al. 2005). Spiders have a great potential as biodiversity
indicators because they have high abundances and
numerous direct and indirect relationships with other taxa;
besides they also have the advantage of being efficiently
sampled and relatively easily identified compared to other
invertebrate groups (Churchill 1997; Marc et al. 1999;
Gravesen 2000; Oxbrough et al. 2005; Hore and Uniyal
2008).
The main aim of this study was to evaluate grass-
dwelling spider species richness and diversity in the cal-
careous fens located at the Coastal Lowland of Latvia and
to compare the differences between grass-layer inhabiting
spider communities within several different fens. In addi-
tion, we aimed to find out the habitat features (mainly
associated with vegetation) that might potentially affect
this group of spiders.
Materials and methods
Study sites
The research was carried out in eight calcareous fens of the
Coastal Lowland of Western Latvia (Fig. 1): (1) Apsuciems
fen; (2) fen of the lake Engure; (3) Platene fen; (4) fen of
the lake Kanieris; (5) fen in the marshy depression (called
viga) of Slıtere; (6) fen in the meadow complex of Vıtini;
(7) Kirba fen; and (8) Jeci fen. All these fens are located in
Fig. 1 Map showing the location of the sampled calcareous fens. All of the studied fens are located in the Coastal Lowland at the Western part of
Latvia
758 J Insect Conserv (2014) 18:757–769
123
Author's personal copy
protected areas which are also included in the Natura 2000network of European specially protected nature territories.
Besides, two of the wetlands—lake Engure and lake
Kanieris—are Ramsar sites which means that these are
wetlands of international importance, designated under the
Ramsar Convention (Ramsar Convention 1971).
Sampling
The samples were collected on 16th and 17th July 2011. To
collect spiders, the sweep netting was chosen which is a
semi-quantitative sampling method and appropriate tech-
nique for grass-dwelling arthropod collection. Spiders were
collected along transects (25 m in length each) which were
placed in the way to proportionally represent the diversity
of microhabitats of the particular fen. So the number of
transects was different in each of the fen—more transects
were placed in those fens in which there were more veg-
etation types. One sample consisted of 50 sweeps—25
strokes were performed along one side of transect and the
rest 25—along the other side. The number of transects
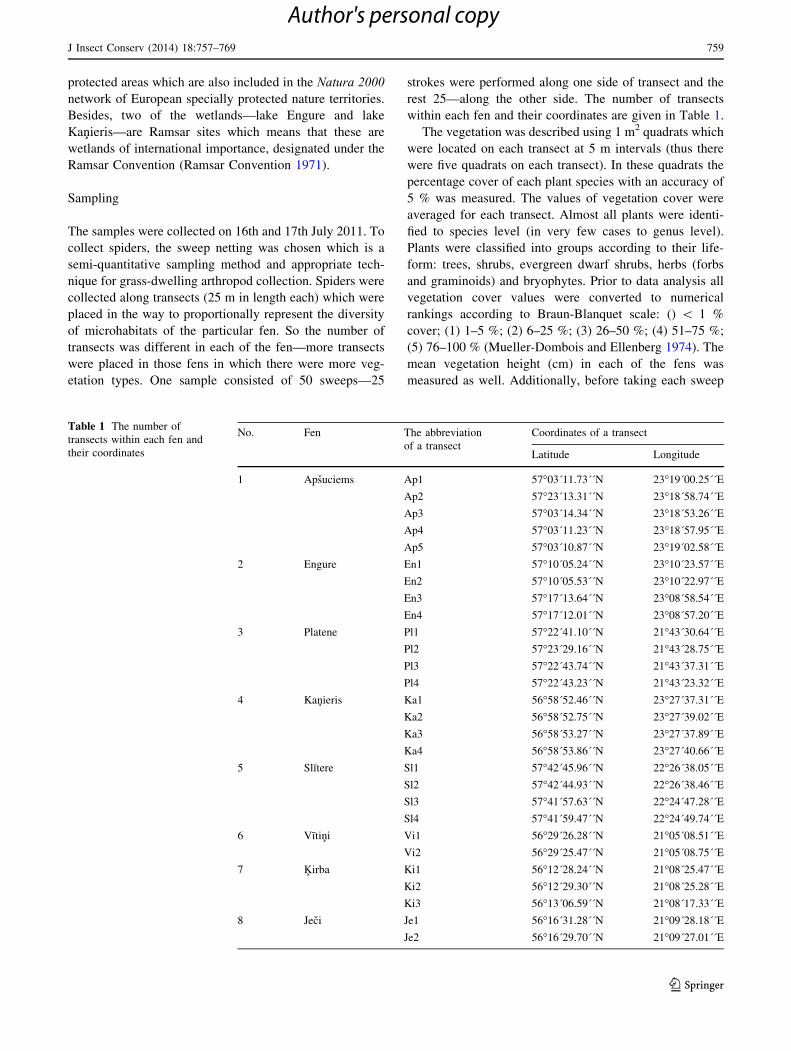
within each fen and their coordinates are given in Table 1.
The vegetation was described using 1 m2 quadrats which
were located on each transect at 5 m intervals (thus there
were five quadrats on each transect). In these quadrats the
percentage cover of each plant species with an accuracy of
5 % was measured. The values of vegetation cover were
averaged for each transect. Almost all plants were identi-
fied to species level (in very few cases to genus level).
Plants were classified into groups according to their life-
Marpissa radiata (Grube, 1859) (5.0 %), Pardosa sphag-nicola (Dahl, 1908) (5.0 %) and Xysticus ulmi (Hahn, 1831)(3.9 %). There were no species that was collected at all
eight fens but ten of the species were collected at only one
fen: Erigone dentipalpis (Wider, 1834) was collected only
in Apsuciems; Dolomedes plantarius (Clerck, 1757), Eri-gone sp. and Araneus sp. only in Kanieris; Gnathonariumdentatum (Wider, 1834) only in Platene; Phylloneta im-pressa (L. Koch, 1881) and Cheiracanthium erraticum(Walckenaer, 1802) only in Vıtini; Larinioides cornutus(Clerck, 1757) only in Slıtere; Enoplognatha ovata (Clerck,1757) only in Kirba; and Kaestneria pullata (O.P.-Cam-
bridge, 1863) only in Jeci. Moreover, two of the species
were collected only in two fens: Araneus diadematus(Clerck, 1757) was collected only in Apsuciems and Kirba
but Microlinyphia pusilla (Sundevall, 1830) – only in
Kanieris and Slıtere. The remaining species found were
Argiope bruennichi (Scopoli, 1772) and Cheiracanthiumsp.
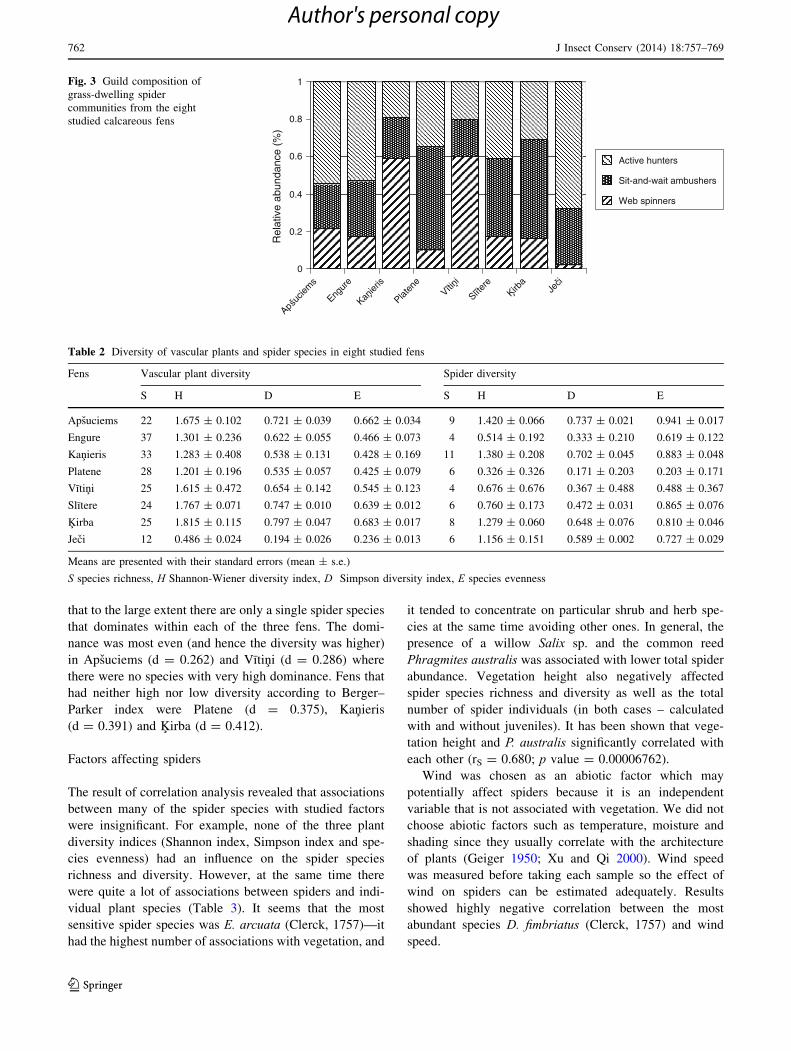
Spider guild composition
The foraging guild composition of spiders was also ana-
lysed. Guilds are groups of species utilizing the same
resource in similar ways. We separated our detected spider
families into three guilds (modified from Uetz 1977;
Canard 1990; Roberts 1996): (1) web spinners (collected
families: Araneidae, Linyphiidae and Theridiidae); (2) sit-
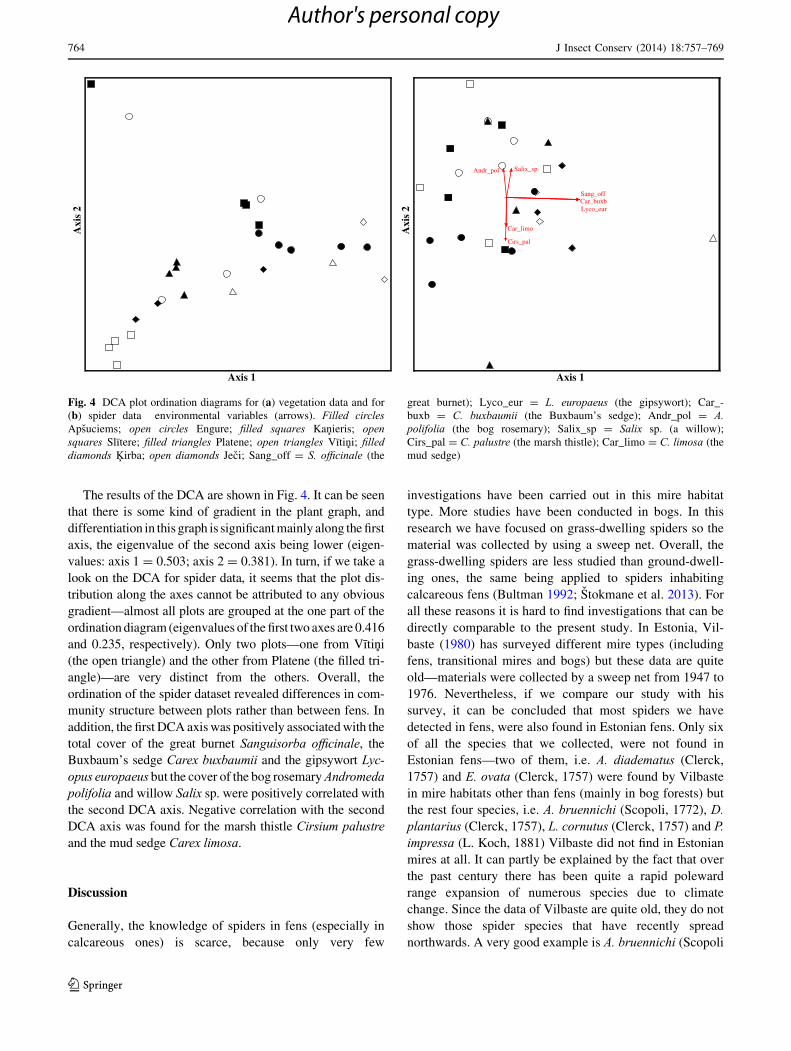
The results of the DCA are shown in Fig. 4. It can be seen
that there is some kind of gradient in the plant graph, and
differentiation in this graph is significantmainly along the first
axis, the eigenvalue of the second axis being lower (eigen-
values: axis 1 = 0.503; axis 2 = 0.381). In turn, if we take a
look on the DCA for spider data, it seems that the plot dis-
tribution along the axes cannot be attributed to any obvious
gradient—almost all plots are grouped at the one part of the
ordination diagram (eigenvalues of the first two axes are 0.416
and 0.235, respectively). Only two plots—one from Vıtini
(the open triangle) and the other from Platene (the filled tri-
angle)—are very distinct from the others. Overall, the
ordination of the spider dataset revealed differences in com-
munity structure between plots rather than between fens. In
addition, the first DCA axiswas positively associatedwith the
total cover of the great burnet Sanguisorba officinale, theBuxbaum’s sedge Carex buxbaumii and the gipsywort Lyc-opus europaeus but the cover of the bog rosemaryAndromedapolifolia and willow Salix sp. were positively correlated withthe second DCA axis. Negative correlation with the second
DCA axis was found for the marsh thistle Cirsium palustreand the mud sedge Carex limosa.
Discussion
Generally, the knowledge of spiders in fens (especially in
calcareous ones) is scarce, because only very few
investigations have been carried out in this mire habitat
type. More studies have been conducted in bogs. In this
research we have focused on grass-dwelling spiders so the
material was collected by using a sweep net. Overall, the
grass-dwelling spiders are less studied than ground-dwell-
ing ones, the same being applied to spiders inhabiting
calcareous fens (Bultman 1992; Stokmane et al. 2013). For
all these reasons it is hard to find investigations that can be
directly comparable to the present study. In Estonia, Vil-
baste (1980) has surveyed different mire types (including
fens, transitional mires and bogs) but these data are quite
old—materials were collected by a sweep net from 1947 to
1976. Nevertheless, if we compare our study with his
survey, it can be concluded that most spiders we have
detected in fens, were also found in Estonian fens. Only six
of all the species that we collected, were not found in
Estonian fens—two of them, i.e. A. diadematus (Clerck,
1757) and E. ovata (Clerck, 1757) were found by Vilbaste
in mire habitats other than fens (mainly in bog forests) but
the rest four species, i.e. A. bruennichi (Scopoli, 1772), D.plantarius (Clerck, 1757), L. cornutus (Clerck, 1757) and P.impressa (L. Koch, 1881) Vilbaste did not find in Estonian
mires at all. It can partly be explained by the fact that over
the past century there has been quite a rapid poleward
range expansion of numerous species due to climate
change. Since the data of Vilbaste are quite old, they do not
show those spider species that have recently spread
northwards. A very good example is A. bruennichi (Scopoli
Axis 1
Axi
s 2
Andr_pol
Car_buxb
Car_limo
Cirs_pal
Lyco_eur
Salix_sp
Sang_off
Axis 1
Axi
s 2
Fig. 4 DCA plot ordination diagrams for (a) vegetation data and for
(b) spider data environmental variables (arrows). Filled circlesApsuciems; open circles Engure; filled squares Kanieris; opensquares Slıtere; filled triangles Platene; open triangles Vıtini; filleddiamonds Kirba; open diamonds Jeci; Sang_off = S. officinale (the
great burnet); Lyco_eur = L. europaeus (the gipsywort); Car_-
buxb = C. buxbaumii (the Buxbaum’s sedge); Andr_pol = A.polifolia (the bog rosemary); Salix_sp = Salix sp. (a willow);
Cirs_pal = C. palustre (the marsh thistle); Car_limo = C. limosa (the
mud sedge)
764 J Insect Conserv (2014) 18:757–769
123
Author's personal copy
1772). It is a thermophilic species that originally occurred
in the Mediterranean part of Europe but during the 20th
century has extended its range northwards (Kumschick
et al. 2011). In 2004 this spider species was detected in
Latvia for the first time (Spungis 2005). The study of Cera
et al. (2010) is another one we need to mention. This study
was conducted in Lake Engure Nature Park from 1997 to
2008, and the grass-dwelling spiders were collected in 12
different habitats including calcareous fens. Eight of spider
species that were collected in our study were also detected
in calcareous fens from Lake Engure Nature Park. These
species were: A. diadematus (Clerck, 1757), E. arcuata(Clerck, 1757), K. pullata (O.P.-Cambridge, 1863), L.cornutus (Clerck, 1757), M. radiata (Grube, 1859), T.maritimus (Menge, 1875) and X. ulmi (Hahn, 1831). Thedominant spider species in the calcareous fens of Lake
Engure Nature Park were T. maritimus (Menge, 1875), M.radiata (Grube, 1859), E. arcuata (Clerck, 1757) and L.cornutus (Clerck, 1757). The former three species were
also abundant in our study but the latter one we collected as
a singleton.
In general, two of the most abundant spider species in
the present study were D. fimbriatus (Clerck, 1757) and T.maritimus (Menge, 1875). It is quite logical for both of
these species to be found in a wetland habitat because they
are typical of moist habitats in general. D. fimbriatus(Clerck, 1757) is usually found in swamps which do not
dry up, because it needs permanent pools of water (Locket
and Millidge 1951). This spider can walk on the surface of
water where it hunts insects, tadpoles or sometimes even
small fish thus also being important predator in semi-
aquatic food webs (Foelix 1996; Roberts 1996). A bit less
abundant in our study was T. maritimus (Menge, 1875).
This species is found on coarse grasses, rushes, heather and
in similar situations. It usually inhabits damp places, for
example, raised bogs (Locket and Millidge 1951; Harvey
et al. 2002). It has also been recorded in calcareous fens
(Cera et al. 2010). The rest of the species accounted for less
than 30 % of all individuals.
In the present study both of the wandering spider guilds
—sit-and-wait ambushers and active hunters—were more
abundant than web builders. This result could suggest that
active wandering might be a more successful hunting
strategy than sedentary lifestyle. There are many reasons
supporting this assumption. Firstly, wandering spiders may
be more likely to find suitable food than web spinners
because while most species of wanderers are capable of
capturing a wide diversity of prey types, web spinners
exhibit considerable specialization on prey (Nentwig 1985,
1986). Secondly, the mobility of actively hunting spiders
provides them with greater flexibility to move readily to
areas with more favorable microclimate and resource val-
ues (Williams 1962; Samu et al. 1999); on the contrary, site
abandonment means a high energetic cost to sedentary
spiders because they must avoid predation during and after
relocation and build a new web (Tanaka 1989; Lubin et al.
1993). And thirdly, wanderers are less sensitive to local
changes in their habitat because they are capable of
exploiting larger areas, i.e. since microhabitat relocations
are part of the foraging strategy of wandering spiders, this
group of spiders uses not only their immediate surrounding
but also suitable habitats in the vicinity of their habitat
patch (Ford 1978; Ehmann and MacMahon 1996).
To evaluate spider diversity in the examined calcareous
fens, several diversity indices were calculated. Diversity
indices are often considered to be a more indicative
diversity measure than mere species richness (Magurran
2004). In all cases the Shannon diversity index of spider
communities was low but the Simpson diversity index—
quite high. It suggests that the grass-dwelling spider com-
munity is characterized by a few abundant species and
numerous rare species. As other authors have shown, this
pattern is actually expected for spider assemblages (Toti
et al. 2000; Bonaldo et al. 2007).
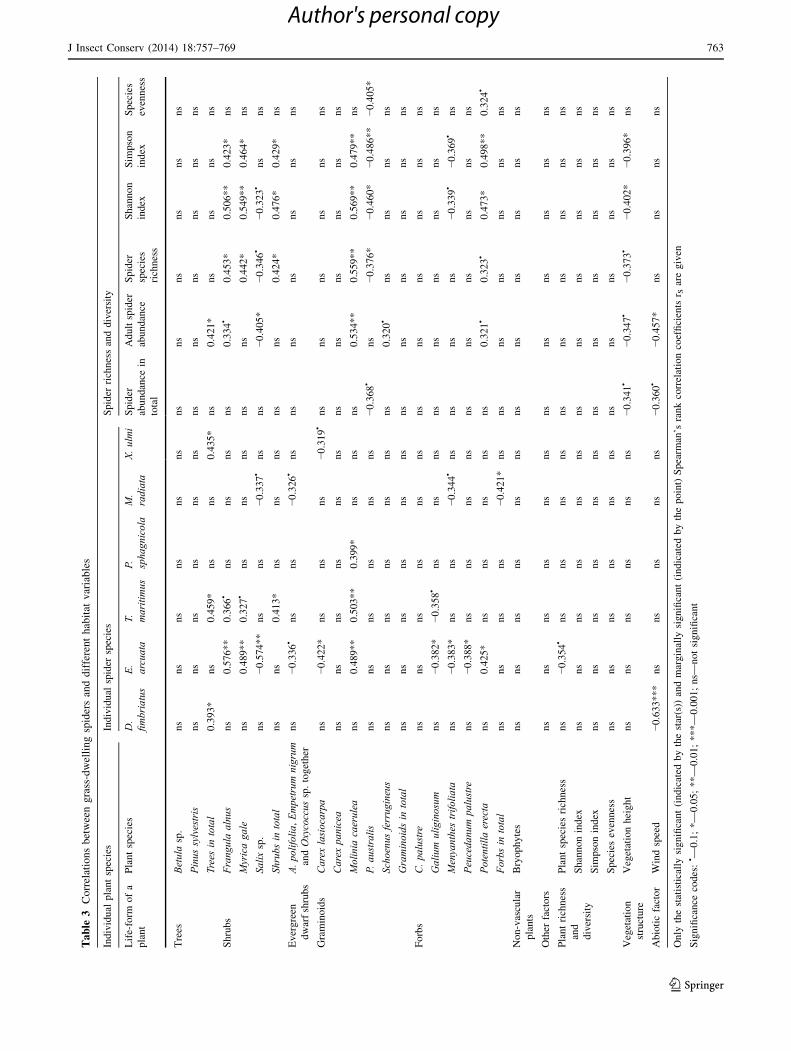
The number of correlations between spiders and differ-
ent habitat features was less than expected. For example,
no consistent relationship between increasing plant species
diversity and patterns of richness and diversity of spider
communities emerged. Nevertheless, there were quite a lot
of associations between spiders and individual plant spe-
cies. The DCA showed quite a similar pattern—some of the
individual plant species were fairly important for spider
communities. These results partly support research by other
authors (Uetz 1991; Pozzi et al. 1998; Jimenez-Valverde
and Lobo 2007; Hore and Uniyal 2008) who have found
that vegetation structure is one of the major habitat features
explaining spider species composition. It has been shown
that greater habitat complexity usually results in a higher
abundance and diversity of spiders (Langellotto and Denno
2004). It is in accordance with the habitat heterogeneity
hypothesis (Tews et al. 2004) which states that structurally
more diverse habitats provide more niches and different
potential ways of resource exploitation. Thus increased
vegetational complexity may provide a lot of benefits for
spiders: more web attachment points (for web spinners),
higher prey density, reduced intraguild predation, access to
alternative food sources and better protection from their
own predators (Uetz 1991; Gunnarsson 1996; McNett and
Rypstra 2000; Langellotto and Denno 2004; Rickers et al.
2006).
The results indicated that spiders were negatively cor-
related with the vegetation height. This observation is
apparently inconsistent with the numerous of other studies
(e.g., Mrzljak and Wiegleb 2000; Dennis et al. 2001; Harris
et al. 2003; Horvath et al. 2009) in which the height of the
vegetation is a habitat feature that positively affects
J Insect Conserv (2014) 18:757–769 765
123
Author's personal copy
spiders. It is associated with the fact that higher vegetation
is usually vertically more structured which increases spider
species richness (Horvath et al. 2009). Probably the main
reason why in the present study vegetation height had a
negative effect on the spider richness and diversity is that
the height of the vegetation in the fens was associated with
a high cover of the common reed P. australis. The corre-
lation analysis revealed that this plant species negatively
influenced spiders. The common reed is a typical expansive
plant which can replace other plant species. Moreover, by
forming very dense stands, this plant can create shading
(Aunins et al. 2010) due to which spider communities
could be affected. Photophilous spider species are espe-
cially negatively influenced. The common reed might have
a high importance for spiders only during the winter
because reed beds have been reported to be substantially
important overwintering places for spiders (Puhringer
1979).
Spiders were also negatively affected by the wind which
is in accordance with several other studies. The negative
effect of the wind is usually associated with the destruction
of spider webs. Spider webs are relatively weak so they can
be easily damaged by wind gusts (LeSar and Unzicker
1978; Hodge 1987; Szinetar 2000). In both the field
observations and in the laboratory experiments there have
been established that spiders tend to build their webs at
places where the influence of wind is smaller (Jocque
1973). Also, Wise (1995) proposed that the abundance of
spiders depends on three niche axes, one of which is wind
(the other two are temperature and moisture).
Calcareous fen conservation is one of the priorities in
the European Community. Although peatland ecosystems
(e.g. fens and bogs) are typical habitats in northern Europe,
nowadays they are endangered even in this region. As a
result many organisms, including spiders, living in peat-
lands are currently threatened. The worst thing is that
spider fauna and ecology are very poorly investigated in
peatland (especially fen) habitats. Spiders have been sug-
gested as good biodiversity indicators which is one of the
main reasons why it is so important to perform systematic
studies of spiders in rare and threatened habitats. In our
previous study we focused on ground-dwelling spiders of
calcareous fens (Stokmane et al. 2013) so this time we
were taking into account the grass-dwelling ones. Consid-
ering all possibilities and limitations, we chose to collect
the data in the mid-July. There are a couple of reasons why
we decided that this period of time would be the best for
the short-term intensive sampling. Firstly, some researchers
have found that spider species diversity and the number of
species positively correlate with mean weekly temperature
and thus reach maximum values during midsummer (Hat-
ley and Macmahon 1980). In Latvia July is the warmest
month of the year (LEGMC 2014), besides our personal
observations also indicate that it is usually the richest
period of the growing season when plant biomass and food
availability are at their peak. Since it is supposed that peaks
of herbivorous insect abundance correlate with flowering
and/or fruiting peaks (Buskirk and Buskirk 1976), it is
expected that the density of spiders will also increase
proportionally—Kiritani et al. 1972, for example, have
discovered that the peak of population density of spiders
coincides with an increase of insect pests. Secondly, it
should be noted that this study was restricted only to grass-
dwelling spiders (including flower-dwellers). If compared
to other spider groups (for instance, ground-dwelling spi-
ders), grass-dwellers is a group of spiders that may be
especially dependent on the presence of different plant
reproductive structures, i.e. buds, flowers and fruits. Grass-
dwellers are indirectly related to these structures, because
they are often dependent on phytophagous insects and
pollinators which usually visit these plant structures (Lo-
uda 1982; Souza and Martins 2004). During the season the
relative amount of plant reproductive elements is varying,
but the midsummer in Latvia is expected to be the hotspot
of the amount and diversity of these structures (personal
observation), so it was hoped that during this period
diversity of grass-dwelling spiders would be at its highest
point or near it. Of course, abundance of spiders varies
seasonally and certain species may dominate at different
times of the season (Robinson et al. 1974; Corey et al.
1998; Sudhikumar et al. 2005), but we need to emphasize
that our aim was to try to evaluate the overall spider
diversity and not the diversity of one or several particular
spider group(s). If our goal, for example, would be to study
a specific spider family, the optimum sampling period
might be very different.
Next, if speaking about the experimental design of this
study, the transect method and the sweep net were used to
collect the grass-dwelling spiders in the chosen fens. The
number of transects differed between fens—it was in
accordance with the vegetation types present in each par-
ticular fen. Since we wanted to focus on the typical
calcareous fen spider species it was supposed that no more
transects are necessary because it has been shown that an
increasing sampling effort results in a higher chance of
finding non-typical species, which have immigrated from
adjacent habitats, and that no typical species are caught
additionally (Bonte et al. 2003). Anyway, sampling over an
entire fen area is recommended to more accurately estimate
spider counts, and more intensive sampling would also be
useful because otherwise it is difficult to assess the habitat
affinities of rarely collected species.
Contrary to our expectations, the total number of adult
spiders collected was much lower than anticipated. In con-
trast, a greater number of juveniles were collected, which
could be explained by the fact that a large number of spider
766 J Insect Conserv (2014) 18:757–769
123
Author's personal copy
species are giving birth to offspring during this time (Cum-
mins 2007). In any case, in the future research it is
recommended to choose some other sampling periods in the
growing season to more completely evaluate the spider
diversity of the calcareous fens. Also, since our sampling
method was restricted only to grass-layer inhabiting spiders,
other collection methods are required to be used in the future
studies to obtain a more complex overview of the spider
fauna of the studied habitats. In general, this research actu-
ally can be considered more like a preliminary assessment,
and the topic deserves future investigations because there is
still a lot to learn about spider fauna and ecology in these rare
and endangered habitats—calcareous fens.
Acknowledgments The authors would like to thank Andris Zie-
melis and Agnese Zukova for their help in collecting spiders. We also
wish to thank Inese Cera for checking spider identifications and for
the help with the identification of difficult spider specimens. This
study was supported by the project (No. 09.1589) “Factors limiting
diversity of animals in terrestrial ecosystems—interaction of natural
and anthropogenic factors” funded by the Latvian Council of Science,
as well as by the European Social Fund project (agreement No. 2009/
0162/1DP/1.1.2.1.1/09/IPIA/VIAA/004) “Support for the implemen-
tation of master studies at University of Latvia”.
References
Aunins A, Bambe B, Engele L, Ikauniece S, Kabucis I, Laime B,
Larmanis V, Reriha I, Rove I, Rusina S, Salmina L, Sniedze R
(2010) European Union protected habitats in Latvia: identifica-
tion handbook. Latvian Fund for Nature, Riga (in Latvian)
Barber KE (1993) Peatlands as scientific archives of past biodiversity.
Biodivers Conserv 2:474–489
Bonaldo AB, Marques MAL, Pinto-da-Rocha R, Gardner T (2007)
Species richness and community structure of arboreal spider
assemblages in fragments of three forest types at Banhado
Grande wet plain, Gravataı River, Rio Grande do Sul, Brazil.
Iheringia Ser Zool 97:143–151
Bonte D, Criel P, Van Thournout I, Maelfait JP (2003) Regional and
local variation of spider assemblages (Araneae) from coastal
grey dunes along the North Sea. J Biogeogr 30:901–911
Bultman TL (1992) Abundance and association of cursorial spiders
from calcareous fens in southern Missouri. J Arachnol 20:165–
172
Buskirk RE, Buskirk WH (1976) Changes in arthropod abundance in
a highland Costa Rican forest. Am Midl Nat 95:288–298
Canard A (1990) Heathland spider communities, a functional group
study. Acta Zool Fenn 190:45–50
Cera I, Spuņģis V, Melecis V (2010) Occurrence of grass-dwelling
spiders in habitats of Lake Engure Nature Park. Environ Exp
Biol 8:59–69
Chapman S, Buttler A, Francez AJ, Laggoun-Defarge F, Vasander H,