Page 1
Effects of Contrasting Temperature on the Developmental Biology of
Bemisia tabaci (Hemiptera, Aleyrodidae) and its transmission of cassava
mosaic and brown streak viruses Tanzania
M. Sc. Thesis
Firaol Taressa Ufga
February 2014
Catania University
Advisors:
Prof. Carmello Rapisarda
Dr. James Legg
Page 2
A Thesis Submitted to the Faculty of the
Department of Agricultural,
School of Graduate Studies
Catania UNIVERSITY
In Partial Fulfillment of the Requirements for the Degree of
MASTER OF SCIENCE IN Sustainable Development in Agriculture
By
Firaol Taressa
February 2014
Catania University
Page 3
iii
DEDICATION
This Thesis manuscript is dedicated to Almighty God, the Omnipotent.
Page 4
iv
STATEMENT OF AUTHOR
I Firaol Taressa Ufga, hereby declare that this dissertation is my original work. It has not
been submitted in any other university. All sources of materials used in this dissertation
have been fully acknowledged.
Name: Firaol Taressa Ufga Signature: …………………
Place: Catania University, Italy
Date of Submission: February 2014
Page 5
v
LIST OF ABBREVIATIONS
ACMD African cassava mosaic disease
ACMV African cassava mosaic virus
ANOVA Analysis of variance
CMD Cassava mosaic disease
CMGs Cassava mosaic geminiviruses
CBSVs Cassava brown streak viruses
CBSD Cassava brown streak disease
CBSV Cassava brown streak virus
CTAB Cetyl trimethylammonium bromide
DNA Deoxyribonucleic acid
EACMV East African cassava mosaic virus
IITA International Institute of Tropical Agriculture
PCR Polymerase chain reaction
RNA Ribonucleic acid
UCBSV Ugandan cassava brown streak virus
Page 6
vi
ACKNOWLEDGMENTS
First thanks to God-the OmniPotent-who gave me this chance to study... I would like to
thank my advisors, Dr. James Legg and Prof. Carmello Rapisarda, as without their
sincere guidance and professional expertise, the completion of this work successful may
not have been possible. Thus, I really thank them for all their support and constructive
comments from the very beginning of proposal write up to the thesis write up.
I feel scanty of words for the boundless love and tireless sacrifice and affection showed
on me by my parents. Therefore, I want to express my unshared thanks to my parents,
who did their best to bring me up and let me to be educated, lacking being educated
themselves.
I would also like to thank Agris Mundus for sponsoring of the whole study and IITA-
Tanzania for the chance; they gave me to conduct my thesis with them and for providing
logistics.
I am extremely grateful to my friends Lensa Sefera and Devid Guastella for their full
cooperation during my thesis work and process.
My special thanks go to IITA-Lab technicians of Dr. James team namely Juma, Digna,
Rudolph, Frank, Simon, and Salehe. I am indebted to all of them for their kindness and
cooperation whenever there was a need. I would like to extend my cordial thanks and
appreciation to my beloved friend Tashome Leta and my aunt Mitike Mosissa for their
generous assistance, moral support, and helpful encouragement during my graduate study
with all their kindness and affection. Finally yet importantly, I would like to thank
Universities of Copenhagen (Denmark) and Catania (Italy) who helped me to acquire the
skills (Sustainable Development in Agriculture) which I believe to help me towards being
an asset of Agricultural Development.
Page 7
vii
TABLE OF CONTENTS
STATEMENT OF AUTHOR iv
LIST OF ABBREVIATIONS v
ACKNOWLEDGMENTS vi
TABLE OF CONTENTS vii
LIST OF TABLES ix
LIST OF FIGURES x
1. INTRODUCTION 1
1.1. Background information and justification 1
1.2. Objectives 3
Main objective 3
1.3. Significance 3
2. REVIEW OF LITERATURE 5
2.1. Origin and constraints to cassava production 5
2.1.1. Whitefly, Bemisia tabaci 5
2.1.1.2. Whitefly as vector of plant viruses 6
2.1.2. Cassava Mosaic Disease (CMD) 7
2.1.2.1. Management of CMD 7
2.1.3. Cassava brown streak disease (CBSD) 8
2.1.3.1. Effect of temperature on CBSD 8
2.1.3.2. Management of CBSD 9
2.1.1.3. Biology 9
Life cycle 9
2.2. Development and growth of insects 10
2.2.1. Effect of temperature on the development of insects 10
2.3. Fecundity 11
2.3.1. Factors governing fecundity 11
2.3.1.1. Temperature and fecundity 11
2.3.1.2. Food and fecundity 12
2.4. Management of B. tabaci 12
2.4.1. Cultural practices 12
2.4.2. Host plant resistance 13
2.4.3. Chemical control of Bemisia tabaci 13
2.4.4. Physical control 14
2.4.5. Biological control 14
Page 8
viii
3. MATERIALS AND METHODS 15
3.1. Experimental site 15
3.2. Methods 16
3.2.1. Host plant production 16
3.2.2. Insect cultures 16
Figure 2. Whitefly, B. tabaci culturing in the screen cages inside screen house 17
3.2.3. Development of immature and its survival period 17
3.2.4. Survival and reproduction of adult female 17
3.2.2. Virus transmission 18
3.2.2.1. Generation of EACMV free-cassava plants (virus receiver) 18
3.2.2.2. Cultivation of virus infected plants (donor plant) 19
3.3.2. Source of test materials 20
3.3.3. Virus detection by PCR and RT-PCR 21
2.5. Whitefly, Bemisia tabaci transmission in screen house 22
3.3. Effect of temperature on CBSV 23
3.3.1. Measuring virus concentration 24
3.3.2 Evaluation of the progression of diseased plants in the growing chambers 25
3.4. Data analysis 25
4. RESULTS AND DISCUSSION 27
4.1. Effect of contrasting temperatures on immature development of Bemisia tabaci 27
4.2. Influence of contrasting temperatures on survival rate of immature stages of B.
tabaci 28
4.4. Effect of contrasting temperatures on female reproductive 29
4.5. Effect of contrasting temperatures on longevity of adult females 32
4.6. Demographic parameters 33
4.7. Transmission of EACMV by B. tabaci 34
4.8. Effect of temperature on CBSV 35
4.8.1. Effect of temperature on CBSV symptom development 35
4.8.2. Effect of temperature on cDNA synthesis 36
4.8.3. Effect of temperature on virus concentration 36
5. CONCLUSIONS AND RECOMMENDATION 38
6. REFERENCES 40
Page 9
ix
LIST OF TABLES
Table page
Table 1. Developmental time of the immature stages of Bemisia tabaci in days (mean
±SE) on cassava at different temperature ……………………………………….28
Table 2. Percentage survival of the immature B. tabaci reared on cassava at three constant
temperatures ……………………………………………………………………..29
Table 3. Survival and fecundity of adult female B. tabaci at different temperatures reared
on cassava ……………………………………………………………………….30
Table 4. Demographic parameters of B. tabaci reared on cassava at three temperatures
.…………………………………………………………………………… ……………..34
Page 10
x
LIST OF FIGURES
Figure page
Figure 1. Map of Kibaha District in coastal region of Tanzania ……………..…………15
Figure 2. Whitefly, B. tabaci culturing in the screen cages inside screen house ………..16
Figure 3. B. tabaci rearing in growth chamber at different temperatures …………..…..17
Figure 4. Study of Bemisia tabaci longevity and fecundity in growth chamber at different
constant temperatures…………………………………………………………….18
Figure 5. Aspirator used in the scollection and transfer of whiteflies…………………...18
Figure 6. Cultivation of CMD infected cassava plant …………………………………...19
Figure 7. PCR products obtained from asymptomatic leaf and healthy leaf. (+) Diseased
from IITA laboratory, (-) Negative control without DNA. M: Molecular marker
(1 Kb plus DNA ladder). ………………………………………………………...21
Figure 8. PCR products obtained from diseased and healthy leaf. (+) from IITA
laboratory, (-) Negative control without DNA and water. M: Molecular marker (1
Kb plus DNA ladder). …………………………………………………………...22
Figure 9. Cassava plant inoculated with CMD and control plants …………………….23
Figure 10 Temperature set up in plant growth chambers for the study of effect of
temperature on CBSV …………………………………………………………...24
Figure 11. Oviposition of adult B. tabaci reared on cassava at three constant
temperatures……………………………………………………………………………32
Figure 12. Longevity of Adult female B. tabaci reared on cassava at three constant
temperatures…………………………………………………………………..33
Figure 13. Effect of temperature on symptom severity of CBSV on cassava………...36
Figure 14. The level of viral RNA accumulation in diseased cassava plants at 18-23
and 33o
C ……………………………………………………………………..37
Page 11
xi
Abstract
Tanzania is one of the leading of cassava-producing countries (Manihot esculenta C.) in
Africa. However, a significant part of the production is lost because of the whitefly,
Bemisia tabaci and its vectored viruses. Temperature is one of the key factors driving
whitefly population development. A study was conducted in Tanzania to determine the
influence of temperature on the developmental biology and survival of B. tabaci and its
transmitted viruses (CBSVs and EACMV) on cassava. This research provides
comprehensive new data about survival and developmental times at four constant
temperatures under laboratory conditions for insects reared on cassava. Temperature
affected each of the immature development stages separately and brood development
from egg to adult. Results showed that the higher the temperature, the shorter the
developmental duration. The survival of each of the immature stages was also differently
affected by temperature. The mean developmental time of adults of B. tabaci ranged from
23 days at 28o C to 35.8days at 18
o C. At 33º C, no adults were produced. In the present
study the percentage of survival of B. tabaci varied from 55.3 at 18 o C to 72.5% at 23
o C.
Fecundity and longevity of adult B. tabaci were significantly affected by the contrasting
temperatures examined. Higher and lower oviposition rates were recorded at 23 and 18o
C correspondingly. Temperature had a significant effect on CBSV symptom development
and viral concentration. At higher temperature (33o
C), the newly developed leaves
appeared less symptomatic. The percentage of plants expressing symptoms varied from
32.0% at 33o C to 59.8% at 18-23
o C. The present study gives basic biological
information which can be used to predict the distribution range of this species and which
will be valuable in anticipating potential future impacts of climate change.
Key words: whitefly, Bemisia tabaci; cassava (Manihot esculenta C.); Tanzania;
Temperature; virus
Page 12
1
1. INTRODUCTION
1.1. Background information and justification
Scientists have anticipated that the mean of global temperature is predicted to augment from 1.4 to 5.80
C (Houghton et al., 2001; IPCC, 2007) at the end of 21th
century because of different reasons. For
Africa, future annual warming ranges from 0.20C to 0.5
0C per decade are predicted under different
scenarios (IPCC, 2007). These changes are projected to influence the population dynamics and the
condition of agricultural insect pests (Cammell and Knight, 1997). Moreover, temperature has a great
effect on their growth, fecundity, and longevity (Bale et al., 2002). During the past few decades, because
of climate change, various shifts in the distribution and abundance of species have already been
produced (Parmesan and Yohe, 2003). Currently, invasive species are considered as one of the
significant environmental challenges facing the world (Ward and Masters, 2007).
Whitefly, Bemisia tabaci is one of the 100 most invasive species of pests and it is the most severe
agricultural pest on cassava and horticultural crops worldwide (Martin et al. 2000 and Oliveira et al.,
2006). Certainly, it has become the most disturbing and prevalent of insect pests of agriculture and
horticulture in the last 3 decades. Several scholars (Olivier et al., 2001 and Oliveira et al., 2006)
reported that damage might be caused directly through feeding and indirectly by vectoring of more
than115 virus species to a wide range of crops. Moreover, it has over 900 host plants (IUCN:
www.issg.org). Cassava is one of the crops most severely affected by B. tabaci and the viruses that it
transmits (Legg et al., 2006; 2011).
Cassava (Manihot esculenta Crantz) is an important crop for food and income generation in Africa,
Latin America, and Asia for at least 800 million people (Alabi et al., 2011). It also serves as a source of
energy (Bruinjn and Fresco, 1989) and livestock feed. In these regions, cassava is considered an
important driver in reducing poverty and enhancing food security. FAOSTAT (2013) showed that from
252.2 mt of global cassava production in 2011, about 55% was produced in Africa. However, in Africa,
its production is limited by several factors that diminish its yield and quality. Nigeria, Congo, and
Tanzania are the leading countries in Africa.
Page 13
2
In Tanzania, cassava is an important staple food crop especially in the rural communities (FAO, 2010;
Nweke, 2002; Mkamilo and Jeremiah, 2005) and ranks second after maize (Zea mays) (Kapinga et al.,
2009). It plays a key role in providing food security, mainly during famine. In 2011, from 739,794 ha of
cassava land 4,600,000 t were harvested (FAOSTAT, 2013), making the yield per unit area 6.2 t/ha.
Nevertheless, cassava production is greatly affected by various pests/diseases, particularly the two main
virus diseases, which cause enormous yield losses in the field (Newke, 2002, Mkamilo and Jeremiah,
2005). Losses have been estimated at 34,000,000 t annually (Legg et al., 2006). Viruses causing both
diseases are vectored by the whitefly, B. tabaci (Dubern, 1994; Maruthi et al., 2005).
Previous studies have shown the importance of various factors that influence the pattern of spread of
virus disease inside and among fields and the factors that restrain or favour such spread. In 1990,
Fauquet and Fargette (1990) reported that disease incidence largely reflects fluctuations in whitefly
populations, which partly depend on climatic factors, including temperature, rainfall, and wind. A
number of studies have suggested high temperature as the primary factor driving the increase in whitefly
populations (Fauquet et al., 1985, David et al., 2006). This causes the rapid spread of cassava mosaic
viruses (Otim-Nape et al., 1997b).
In Tanzania and elsewhere, the management of B. tabaci has been a main challenge for many years. It is
important to know the longevity and reproduction of this pest under different ecological situations to
forecast its population growth and time control strategies. Such knowledge and information are
important in order to know, assess, and forecast the chronological spread of the diseases that B. tabaci
causes or transmits. Thus, the current study aspires to investigate the relationships between temperature
and developmental biology parameters and to determine the influence of contrasting temperatures on the
dynamics of B. tabaci populations and its diseases transmission characteristics.
Climate conditions, particularly temperature, are possibly the key ecological factors affecting life
tables of B. tabaci. Further research (Yamamura and Kiritani, 1998) projected that as
temperature increases by 2oC insects might experience one to five additional life cycles per
season. Similarly, it is one of the important factors, which determines the incidence and severity
of disease in coincidence with the rain, although the influence might be positive or negative
Page 14
3
(Yáñez-López et al., 2012). Studies have shown that the specific impact of climate change on
insects and pathogens can be hard to predict (Petzoldt and Seaman, 2007). Thus, understanding
the prospective influences of temperature in altering pests and diseases on plants is a key subject,
particularly for the invasive pest B. tabaci and its vectored viruses.
In the past, many studies have been conducted on B. tabaci focusing on management aspects
viz., cultural, biological, chemical, host plant resistance, virus transmission, host range, biotype,
and bionomics by many authors (eg. Gerling and Mayer, 1996; Naranjo, 2001; Qiu et al., 2005,
Oliveira, 2006). Despite the widespread occurrence of B. tabaci in Africa, particularly in
Tanzania, there is scarce and limited information on its developmental biology parameters in
relation to temperature when reared on cassava. Moreover, the essential time for whitefly to
complete its development also depends on its host. The proposed study aimed to investigating
the effect of contrasting temperature on developmental biology of B. tabaci and its disease
transmission behaviour for the two main groups of viruses.
1.2. Objectives
Main objective
Explore the consequence of temperature on the life cycle of Bemisia tabaci and on its virus
transmission characteristics.
Specific objectives of the study are to:
1. Assess the influence of different constant temperatures on different stages, survival and
reproductive ability of Bemisia tabaci;
2. Ascertain the effect of temperature on cassava brown streak viruses (CBSVs) and
transmission characteristics of East African cassava mosaic virus by B. tabaci.
1.3. Significance
B. tabaci causes food shortage and famine in many countries of eastern Africa. The effects of
this pest on crop production are diverse and are widely known. However, in this region, little
Page 15
4
evidence is available on how climate change, particularly temperature, affects the developmental
biology and its influence on virus transmission characteristics of cassava mosaic viruses and
cassava brown streak viruses. This study is particularly important in generating essential
information and data for designing and developing models and strategies to manage with the
potential impacts of whitefly. Moreover, it provides the pest adaptation capacity to different
temperatures and its effect on distribution if mean temperatures increase because of global
warming.
Scope and limitation of the study
Farmers in Kibaha District in the coastal region of Tanzania produce both annual and perennial
crops. Cassava is the most important root crop grown in Tanzania but highly damaged by B.
tabaci. The Kibaha-grown field crop of cassava is heavily attacked by Bemisia. In addition, it is
highly affected by diseases such CBSD and CMD which are transmitted by B. tabaci. Therefore,
this study focuses on the effect of contrasting temperatures on the developmental biology of B.
tabaci and its virus transmission behaviour under four constant temperatures which are typical of
those prevailing in different parts Tanzania. The range covered the maximum and minimum
temperatures experienced in coastal Tanzania. The study used data generated from laboratory
and greenhouse to analyze the effect of temperature under the response of climate change.
However, the results and conclusions generated should be useful to research institutes, decision
makers and farmers in ensuring proper pest management of cassava pests and disease.
Page 16
5
2. REVIEW OF LITERATURE
2.1. Origin and constraints to cassava production
Cassava is native to South America more probably to Brazil from where it was introduced into
Africa in the 16th
century by Portuguese traders (Carter et al., 1995). It belongs to the family
Euphorbiaceae (Lebot, 2009). Currently, it is cultivated in more than 80 countries of the world.
Tanzania is of one of leading producer of cassava in Africa, but yields are very low estimated at
6.2 t/ha (FAOSTAT, 2013) as compared to other cassava producing countries such as Nigeria
which produces more than 16 t/ha. Moreover, under optimal conditions cassava can produce up
to 80 t/ha of tubers in a 12 month culture period (Legg and Thresh, 2003). A number of factors
are responsible for the severely reduced yields in Tanzania, and the main threats are virus
diseases, particularly cassava mosaic disease (CMD), cassava brown streak disease (CBSD) as
well as bacterial blight (caused by Xanthomonas axonopodis pv. manihotis) and pests such as B.
tabaci. Other constraints are poor agricultural practices and various other diseases caused by
bacteria, fungi and nematodes (Hillocks and Wydra, 2002), most of which are considered of
minor importance. On the other hand, cassava farmers suffer from inadequate access to disease-
free planting materials, processing facilities, markets, and inconsistent policies (FAO and IFAD,
2005). Both CMD and CBSD are now considered the most disease constraints to cassava
production in Tanzania; both are vectored by B. tabaci.
2.1.1. Whitefly, Bemisia tabaci
2.1.1.1. Economic Importance of Bemisia tabaci
Manzari and Quicke (2006) reported that there are more than 1,450 whitefly species. Among
these species of whitefly, some are economically very important viz., the silverleaf whitefly
(Bemisia tabaci), and the greenhouse whitefly (Trialeurodes vaporariorum). The genus Bemisia
belongs to the order Hemiptera and family Aleyrodidae.
Whiteflies of the Bemisia tabaci (Hemiptera: Aleyrodidae) complex are global pests of a broad
range of plant species of agriculture and horticultural crops (Wang and Tsai, 1996; Simmons and
Page 17
6
Abd-Rabou, 2005; Linjar and Sahito, 2005). It is widely distributed throughout the world
especially in tropical and subtropical regions. Oliveira et al. (2001) reported that the worldwide
economic loss ranges from hundreds to thousands of millions of dollars a year.
B. tabaci is a highly polyphagous pest (Greathead, 1986) which has more than 900 host species
(www.issg.org). It affects hosts by sucking the plant’s sap causing reduced growth, stunting, and
yield reduction. The honeydew secreted from whitefly can result in the development of sooty
mould on produce (Jiang et al., 1999) which stops leaves from functioning efficiently (Jones,
2003). B. tabaci also vectors viruses that result in enormous economic losses in various crops
throughout the tropics.
2.1.1.2. Whitefly as vector of plant viruses
The world’s most significant diseases of field and horticultural crops are spread by whitefly.
From the whitefly genera Bemisia tabaci is an important virus vector on many global crops
(Flores, 2008). More than 115 virus species are transmitted by two principal vectors, Bemisia
tabaci (genera Begomovirus, Ipomovirus, Crinivirus and Torradovirus) and Trialeurodes
vaporariorum (genera Crinivirus and Torradovirus) (Jones, 2003; Stephen, 2010). Ninety
percent of all B. tabaci transmitted viruses are begomoviruses, which are rapidly evolving plant
viruses. Brown and Bird, (1992) reported that by Begomovirus losses of between 20% and 100%.
According to Jones (2003), whitefly instars nymphs and adults feed by inserting their stylets into
the leaf, piercing the phloem, and withdrawing sap. Thus, the plant viruses are acquired during
this feeding process. Adult whiteflies may move and transmit the virus to new plants whilst
feeding. When the plants are infected within 5-6 weeks after germination, the losses range from
40 to 70% (Villas Bôas, 2005).
Among those virus species transmitted by whitefly, cassava mosaic geminiviruses (CMGs) and
cassava brown streak viruses (CBSVs) are exclusively transmitted by Bemisia tabaci (Frederic et
al., 2012). From many biotic factors, which constrain cassava production, these two viruses are
the major threats (Mware et al., 2009).
Page 18
7
2.1.2. Cassava Mosaic Disease (CMD)
Thresh and Cooter (2005) indicated that CMD is caused by a number of begomoviruses and is
the bottleneck of cassava production in Africa. CMGs are naturally vectored by the whitefly B.
tabaci while being widely dispersed by the distribution of stem cuttings. According to Legg
(1999), CMD shows a chlorotic yellow mosaic on the leaves, reduction in size and deformation
of leaves, and general plant stunting which leads to a reduction in tuberous root production.
CMD causes losses of 35,000,000 t annually (Legg and Thresh, 2004; Legg et al., 2006).
Many reports from different corners of the world have shown that the incidence and severity of
CMD are very high. For instance, in India, yield losses of up to 88% in susceptible varieties and
up to 50% in field tolerant varieties (Edison, 2004). Likewise, in Congo, diagnostic surveys
revealed yield losses of up to 95% (Ntawuruhunga et al., 2007). Nevertheless, Thresh et al.,
(1997) suggested that the overall incidence of CMD is currently 50-60% and diseased plants
suffer losses of up to 40%. According to some survey results, the loss due to CMD generally
exceeds 50% in 11 African countries out of 18. For instance, it causes losses up to 79% in
Congo, 84% western Kenya, and 82% in Nigeria (Sseruwagi et al., 2004). In the same way
CBSD also causes a significant yield reduction in cassava. Recent research reported that the two
diseases constitute the greatest disease threat to cassava production in Tanzania. Nevertheless,
the incidence and severities of the problem differ by zone.
2.1.2.1. Management of CMD
CMD has been managed by different methods and approaches. It is mainly focused on the use of
CMD-resistant varieties (Otim-Nape, et al., 2000) and a great success was achieved by this.
However, according to Legg and Fauquet (2004), the pace of the pandemic spread exceeds the
pace of implementation of these measures. This situation creates other alternative means of
management for instance the use of phytosanitation measures, which might avoid undue
dependence on resistant varieties (Thresh, 2003). Selecting clean planting material and
identifying varieties resistant to the whiteflies that spread the disease are some further control
approaches.
Page 19
8
2.1.3. Cassava brown streak disease (CBSD)
CBSD is caused by two species of CBSVs – Cassava brown streak virus (CBSV) and Ugandan
cassava brown streak virus (UCBSV) (family Potyviridae; genus Ipomovirus) (Monger et al.,
2001; Mbanzibwa et al., 2010). The disease is endemic in the coastal lowlands of Eastern Africa
and the lakeshore region of Lake Malawi. The disease is characterized by leaf chlorosis and stem
lesions with can cause complete die back as well as the spoilage of roots due to dry corky
necrotic rot on starchy tissues (Hillocks, 1999; Hillocks et al., 2003). CBSD causes economic
losses resulting from damage to the above ground parts and root rot. CBSD has been reported to
cause up to 70% yield loss by reducing the root sizes, causing pitting, and constriction on roots
(Hillocks et al., 2001). Similarly, a survey has revealed crop losses up to 74% (Muhanna and
Mtunda, 2002) in Tanzania because of this disease. Necrotic lesions and/or discoloration of the
roots due to infection render them unpalatable and unmarketable, and this explains most of the
quantitative and qualitative losses (Nichols, 1950). Due to this reason, the roots of the cassava
plant can become unfit for human consumption so it is of great concern. CBSD is a major threat
to food security particularly where large numbers of people depend on cassava as their staple
food.
The disease is also propagated through the planting of infected cuttings. Lennon et al. (1986)
reported that the majority of attempts to transmit CBSVs with whitefly had failed. Similar
findings were reported from Kenya (Bock, 1994). However, recently Maruthi et al. (2005)
reported successful transmission with B. tabaci. Previously, CBSD was restricted to coastal areas
of East Africa but more recently new spread has been reported from higher altitude areas of East
and Central Africa (Alicai et al., 2007; Legg et al., 2011).
2.1.3.1. Effect of temperature on CBSD
Few scholars have reported that CBSD is affected by temperature (Nichols, 1950). The period
before symptoms appear on indicator a plant is affected by the temperature and amount of
Page 20
9
inoculum applied (Ogwok et al., 2010). It also varies based on cultivar, rainfall, altitude, plant
age, and type of virus (Hillocks et al., 1996). Determining the effect of temperature may
contribute to the management and help to elucidate the effect that climate change will have on
this virus disease.
2.1.3.2. Management of CBSD
The main way of controlling CBSD is the use of clean cassava planting material. The most
sustainable way of controlling CBSD, however, is the use of resistant varieties. Thus, research in
developing or selecting resistant varieties is currently on-going in several countries, with some
promising results in Tanzania, Uganda and Mozambique. Some scholars suggest that integrated
management of CBSD is the best way. These alternatives include: field hygiene, use of disease-
free planting materials, and use of cassava brown streak disease resistant /tolerant cassava
varieties, propagation of clean materials, quarantine and legislation.
2.1.1.3. Biology
Life cycle
Several authors studied the life history of B. tabaci. Some of them reported that optimum
development depends on temperature. The silverleaf whitefly prefers temperatures of 25°C to
30°C for development and rapid generation time (http://entnemdept.ufl.edu/fasulo/whiteflies).
According to their study, the silverleaf whitefly, Bemisia tabaci eggs are attached to the
underside of the leaf surface, usually younger leaves. Eggs hatched in eight to ten days at this
temperature. The immature stages are four namely; crawlers or first instar nymphs, second, third
and fourth instar nymphs. The crawlers move a short distance before settling to feed on plant
tissue and the following nymphal instars are stationary and remain attached to the leaf surface
where they feed until developing into the fourth and final nymph stage. Nymphs stop feeding,
pupate (albeit via an incomplete metamorphosis), and emerge from the pupal case as fully
developed adults. This adult is accountable for virus spread from plant to plant. It takes from 18
to 28 and 30 to 48 days from egg to adult in warm and winter weather respectively
(www.ozanimals.com).
Page 21
10
2.2. Development and growth of insects
Insect development is characterized by a period of time, number of instars, and an increase in
size and weight as the insect passes from immature to adult phase. The increase in size and
weight is often referred to as insect growth as opposed to development (Dent and Walton, 1997;
Wyman and Pelliteri, 1998). Bonne (1951) defined the development time of an insect as the
period between birth and the production of the first offspring by the adult female and hence,
including the pre-oviposition period. Developmental time can be determined either as a total time
from birth to first offspring or as a series of times for each instar, in which case the pre-
oviposition period is defined as a specific stage from the final adult moult to the production of
the first offspring (Dent and Walton, 1997). The number of instars, which constitute the
immature stages of the insect, will often vary according to conditions such as host quality and
temperature. The rate of development of insect eggs and pupae is primarily dependent on
temperature while the development of larval and nymphal stages is dependent both on
temperature and host plant factors (Dent and Walton, 1997).
2.2.1. Effect of temperature on the development of insects
The development of immature stages and the adult maturation of all insects were mainly affected
by temperature (Fletcher, 1989; Hellmann, 2002). Temperature acts on insects in two ways:
directly on survival and development and indirectly through food, humidity, rainfall, wind,
atmospheric pressure and others (Dent and Walton, 1997). The effect of temperature on the
developmental times and survival of insects can profoundly determine their distribution.
Insects have no precise mechanism for regulating the temperature of their bodies because they
are all poikilothermic. Their body temperature, therefore, follows more or less closely that of the
surrounding medium (Gilbert and Raworth, 1996). Due to this fact, estimating insect
development is difficult. Consequently, we need some way to combine time and temperature so
that we can predict insect development. Therefore, there is a need to measure what is called
"Physiological time" (Chinag, 1985).
Page 22
11
Physiological time is the amount of heat required for an insect to achieve a stage of development.
Chinag (1985) defined degree-days as the amount of heat needed for each species to complete
their life cycle or part of it, concerning the temperature to which it is exposed. It is used to
measure physiological time by combining time and temperature. Degree-days are referred by a
number of other terms such as heat units, thermal units, and growing degree-days, but the
concept is the same. In forecasting infestations, monitoring, and timing of insecticide
applications, the thermal constant provides a valuable tool for insect pest control (Zalom et al.,
1983)
In many cases the development of insects determined under constant conditions of temperature
has been shown to differ from the development of those maintained under fluctuating
temperatures, usually with faster development times occurring under fluctuating conditions
(Foley, 1981). It should not always be assumed that rates derived under constant conditions of
temperature would always be applicable (Yazdani and Agarwal, 1997).
2.3. Fecundity
The reproductive output of insects in terms of the total number of eggs produced or laid during
the lifetime of the female is known as fecundity (Jervis and Copland, 1996). In insects, which
mature eggs throughout their adult life, fecundity is measured directly by keeping females under
caged conditions, which assume natural condition as much as possible, and recording the total
number of eggs laid (Southwood, 1978). Fecundity can be influenced by biotic and abiotic
factors (Dent and Walton, 1997). The biotic factors may be classified as intrinsic, for example
insect size, clone and extrinsic such as host plant effects, which may include plant species,
cultivars or growth stage differences. Temperature is the most important factor, which influences
insect reproduction (Fletcher, 1989).
2.3.1. Factors governing fecundity
2.3.1.1. Temperature and fecundity
Similar to the development of immature stages egg production and oviposition are often affected
by the limits of favourable temperature range. In the majority of cases, fecundity is higher
towards a moderately high temperature and declines as the upper and lower limit is reached
Page 23
12
(Yazdani and Agarwal, 1997). The reproductive potential of three fruit flies was measured in
environmental chambers maintained at temperatures of (maximum: minimum) 24:13, 24:24:,
29:18, and 35:24 + 1 o
C (Vargas et al., 2002). According to the authors, at 29:18 o
C all species
attained their highest fecundity. Similarly, in case of cotton stem weevil the maximum number of
eggs was laid at 32.8 o
C but with the increase in temperature, the fecundity decreased (Ayyar et
al., 1981).
2.3.1.2. Food and fecundity
Insects are essentially omnivorous and are influenced by variations in quantity and quality of
food resources. According to Frederic et al. (2012), the preference of nutrients and food plays
significant roles on the development rate and fecundity of insects. Furthermore, Morrison et al.
(1982) revealed the effect of protein on the male was more complex.
2.4. Management of B. tabaci
B. tabaci and the viruses it vectors cause wide losses to cassava in Africa. According to Javed
(2009), control strategies based solely on insecticides have often proved markedly ineffective in
combating whitefly outbreaks. A more predictable outcome has been the development of
insecticide resistance in B. tabaci, diminishing control efficacy still further and promoting even
higher pesticide inputs. Consequently, resistance in B. tabaci is already geographically
widespread and in extreme cases already extends to virtually all available control agents. There
are no simple or universal solutions for managing resistance in B. tabaci (Javed, 2009). Overall,
whitefly management comprises: host plant resistance, cultural control, biological control, and
chemical control. Some of these are currently applied for the control of B. tabaci on cassava in
Africa while few of them are on trials.
2.4.1. Cultural practices
Page 24
13
In integrated pest management, cultural practices can play an important role (Hilje et al., 2001),
due to their preventative nature. For instance, sanitation is one important method (McAuslane,
2000), crop-free periods, altering planting dates, crop rotation (Hilje et al., 2001). In vegetable
production in Israel, cultural control focused on practices such as screens and inert ground covers
(Berlinger and Lebiush-Mordechi, 1996).
2.4.2. Host plant resistance
Russell (1978) states host plant resistance is one of the main basic components of IPM. The
utilization of this has long been considered as one of the most successful mechanisms of insect
and disease control. It may also provide a more bio-rational approach for reducing the impact of
B. tabaci transmitted viruses and plant disorders than reliance on pesticides. Consequently,
research aiming to enhance the use of varieties resistant to pest organisms has been carried out in
many countries (Nombela and Muntz, 2010).
2.4.3. Chemical control of Bemisia tabaci
For many agricultural systems affected by whiteflies and whitefly-transmitted virus, effective
control is dependent on chemicals (Oliviera et al., 2001) that are used in both protected and
unprotected cultivation. It is regarded as a useful and powerful device for the management of B.
tabaci (Muhammad, 2006). Nevertheless, good results have been achieved with the use of more
selective chemistries; use of action thresholds and resistance management. Growers use a
diversity of chemicals however; neonicotinoid-based products are more broadly utilized. The
habit of B. tabaci nymphs developing on the undersides of leaves inhibits the effectiveness of
contact insecticides, and the capacity of B. tabaci to develop resistance to many insecticide
chemistries makes this insect difficulty to control with insecticides (McAuslane, 2000). Studies
have also shown that insect growth regulators (IGRs) can be effective for the control of B. tabaci
(Naranjo et al., 2004), and these may have a more selective action than conventional insecticides.
Pesticides are virtually never used for the control of pests and diseases on cassava, including
whiteflies.
Page 25
14
2.4.4. Physical control
In different parts of the world different physical methods such as a very fine netting; yellow
traps; barriers, UV-absorbing screens or greenhouse plastic films were used to control B. tabaci.
These techniques have been used to stop the adult B. tabaci from reaching the host plants and to
reduce the infestation caused by this pest both in open and protected fields (EPPO, 2002).
2.4.5. Biological control
B. tabaci is also controlled by using parasitoids, predators, and fungal diseases (Gerling, 1996).
Parasitoids have been most widely used, and may be integrated with rational insecticide use
(Manzano et al., 2003). Therefore, management of this pest by using biocontrol is an important
component of IPM. Encarsia formosa is among the best studied biological control agents of B.
tabaci (Mandour et al. (2003). Encarsia adults attack B. tabaci nymphs by repeatedly inserting
their ovipositor into the body of the host and feeding on the fluid exuding from the wound of
their host. The parasitoids Encarsia spp. and Eretmocerus eremicus were found to be the most
important mortality factor in the pupal stage of B. tabaci B biotype (IOBC, 2008). Natural
enemies are an important component of IPM in controlling Bemisia.
Page 26
15
3. MATERIALS AND METHODS
3.1. Experimental site
Laboratory and green house experiments were conducted at the International Institute of Tropical
Agriculture (IITA), Tanzania. The screen houses were found both in Kibaha (-6°46'0.012”S
latitude and 38°55'0.012”E longitude) and Dar es Salaam in Coast Region, Tanzania. Kibaha is
located 42km to the east of Dar-es-Salaam at an altitude of 154 m.a.s.l. It receives an average
annual rainfall of 800 mm and its average annual temperature is 28° C. The soil type of the
center is luvisol/eutric nitosols with a good drainage system. The laboratory experiments were
carried out at IITA’s new science building, Mikocheni, Dar-es-Salaam, Tanzania.
Figure 1. Map of Kibaha District in Coast Region, Tanzania
Page 27
16
3.2. Methods
3.2.1. Host plant production
Host plant production was carried out both in Kibaha and Dar es Salaam. The plant materials
used in studying the effect of contrasting temperature on developmental biology of B. tabaci was
cassava cuttings selected from the Kibaha station. Cassava plants of variety Kiroba were grown
in plastic pots and used in the trials.
3.2.2. Insect cultures
Whiteflies, Bemisia tabaci, were first collected on cassava from the Kibaha station. The stock
population of B. tabaci was reared on potted cassava that was reserved in screened cages (1m
height × 50cm × 50cm) in a screen house at IITA Station, Kibaha, and later shifted to
Mikocheni, Tanzania. For the study of
developmental biology of Bemisia, rearing
containers were made of plastic pots (6cm in top
diameter, 4.8 cm in bottom diameter, and 6.7 cm in
height) containing cassava seedlings (five-eight leaf
stage). Two holes on two sides were cut and covered
with fine cloth particularly at the center of the
screen cage for ventilation. For the developmental
biology study, 3-5 pairs of adult whiteflies were
released using the leaf clip cages into each of a total
five rearing containers for each temperature.
Page 28
17
Figure 2. Whitefly, B. tabaci culturing in the screen cages inside screen house
3.2.3. Development of immature and its survival period
About 30 - 50 couples of B. tabaci adults from the stock colony were released by using leaf clip
cages in to each cassava culture at 270
C for 24hr to lay eggs. After that, the adults were
detached and leaves were observed with a
magnifying glass. The eggs were thoroughly
counted and kept on the abaxial surface of
leaves. Plants were then transferred into the
environmental controlled growth chambers set
up at four different constant temperatures: 18,
23, 28, and 330C with relative humidity of 65 %
adjusted with diurnal photoperiod, in five
replications. A completely randomized design
was used. Observations were carried out daily,
and once the eggs were hatched into first instars
and fixed on the leaf, young nymphs were
identified individually. The exact nymph
development times at each of the four
experimental temperatures were noted.
Figure 3. B.tabaci rearing in growth chamber at different temperatures
3.2.4. Survival and reproduction of adult female
The newly emerged adults of males and females were paired and subsequently placed in
individual a controlled environmental growth chamber set at a different constant temperature. It
was placed in a leaf clip-cage on the under surface of new leaflets. For each temperature, 13-16
adults were placed in growth chambers at a minimum of 65% ± 5% RH. After 48 h, the insects
were moved with the leaf clip-cages to new leaves using aspirators (Figure 6) and then the
Page 29
18
number of eggs laid per
female was counted until
death of the female. The
period of adult survival
was recorded for all
individuals used in this
experiment.
Figure 4. Study of Bemisia tebaci longevity and fecundity in growth chamber at different
constant temperatures
Figure 5. Aspirator used in the scollection and transfer of whiteflies
3.2.2. Virus transmission
3.2.2.1. Generation of EACMV free-cassava plants (virus receiver)
Disease-free plants of cv. Kiroba were collected and planted in plastic pots of 6 - 10cm diameter
and 7 - 11cm height in a sterilized soil medium constituting of gravel, farmyard manure and
forest soil mixed in the ratio of 1:1:3v/v. Plants were grown from cuttings in the screen house at
Kibaha and Dar es Salaam for more than two months in order to see good expression of CMD
Page 30
19
symptoms.
Infection
status was
double-
checked by
PCR. Plants
obtained in this way were used for the EACMV transmission experiment. The experiment was
conducted on 1 July 2013.
3.2.2.2. Cultivation of virus infected plants (donor plant)
The virus (CMD and CBSD) infected hard wood cassava cuttings were collected from Kibaha.
The stem cuttings were planted in plastic planting pots of 10 cm diameter and 11 cm height in a
soil medium containing sterilized soil constituting of gravel, farmyard manure and forest soil
mixed together mixed in the ratio indicated above. The presence of the virus was detected from
both cuttings and seedlings by PCR.
Figure 6. Cultivation of CMD infected cassava plant
Page 31
20
.
3.3.2. Source of test materials
During collection of the cutting for raising cassava seedlings, the two group of cassava disease
were identified based on their symptom from fields at Kibaha. Leaves showing deformed leaves,
mosaic pattern and overall dwarfing of the plant for CMD while leaves showing feathering and
yellowing symptoms for CBSD were collected on two-three diseased plants for each disease
from two varieties namely Kiroba and Karoora. The leaves which showed clear the symptoms
were collected for testing, particularly those from the middle of the shoot. For negative controls,
leaves were collected from symptomless that tested negative for virus with PCR. About fourteen
samples were collected from two sites at Kibaha. Each sample were put in a separate Eppendorf
tube, labeled and stored in plastic bags for preserving moisture and then taken to the lab at IITA
Dar es Salaam for virus detection using PCR and RT-PCR.
DNA was extracted from the cassava leaf tissues using the protocol of Dellaporta et al. (1983)
and RNA extracted using the CTAB (cetyl trimethyl ammonium bromide) (modified from Xu et
al., 2010). About 100 mg of fresh leaf sample was grinded using a Genogrinder and 750 μl
CTAB. These were incubated for fifteen min at 65°C and then it was mixed with 750 μl
chloroform: isoamylalcohol (24:1). This was centrifuged for ten min at 12000 rpm. After
centrifugation, the upper phases were transferred to a new tube, and then mixed with 300μl cold
isopropanol and incubated at -20°C. The supernatant was discarded after the samples were
centrifuged at 13000 rpm for 10 min., and EtOH (70%) was then added to the pellet and
incubated for 10 min at -20°C. Finally, it was centrifuged at 13000 rpm for 5 min. and the
alcohol was removed. The DNA and RNA extract was re-suspended in 100-μl sterile water for
storage and subsequent use in virus detection.
Quantity and purity of DNA
After extraction, the DNA and RNA concentration was measured with a Nanodrop
Spectrophotometer. This helps to check the quantity and purity of the DNA extracted from all
Page 32
21
samples. The absorbance ratios A260/A280 and A260/A230 for protein contamination and the
presence of polyphenolic / polysaccharide compounds were used respectively.
The results of the spectrophotometer analysis for A260/A280 and A260/A230 showed the
extracted DNA was free from proteins and polyphenolic/polysaccharide compounds. The result
found for protein and polysaccharide were >2 and 1.80 to 2.00 respectively. The average final
concentration of DNA ranged from 388 to 1442 ng/μl.
3.3.3. Virus detection by PCR and RT-PCR
The total DNA from plant tissues were extracted based on Dellaporta et al. (1983) for
polymerase chain reaction (PCR) analysis. 2 μl of DNA were applied to PCR in a reaction
volume of 20 μl. The primers used with PCR to amplify partial DNA-B components for all the
samples were EAB555 for both forward and reverse directions (Ndunguru et al., 2005). These
primers amplify PCR products of about 540-560 bp from the DNA-B components of East
African cassava mosaic virus. The thermal profile 3, 1, 1.5 and 10 min for denaturation, primer
annealing and strand extension at 94°C, 94°C, 55°C, 72°C respectively, was used for 42 cycles.
In anticipation, the DNA fragments were electrophoresed in agarose gel. The result shows that
out of fourteen samples subjected to PCR analysis, EACMV was found in three of them. These
were used for virus transmission studies.
PCR analysis before planting the cuttings
Page 33
22
Figure 7. PCR products obtained from asymptomatic leaf and health leaf. (+) Diseased from
IITA laboratory, (-) Negative control without DNA. M: Molecular marker (1 Kb plus DNA
ladder, __).
Figure 8. PCR products obtained from diseased and healthy leaf. (+) from IITA laboratory, (-)
Negative control without DNA and water. M: Molecular marker (1 Kb plus DNA ladder).
For the detection of CBSVs, complementary DNA (cDNA) was generated using RT-PCR. The
cDNA was subjected to PCR as described above with the Strategene MxPro- Mx3000P
Quantitative PCR unit with a programme set at 48oC for 30min for the initial denaturation,
followed by 95oC for 10 min (denaturation), 95
oC for 0.15min (annealing) and 60
oC for 1 min
(strand extension). This was repeated for 52 cycles. The result showed that out of 14 samples
seven were infected by CBSVs; five samples were infected by both UCBSV and CBSV and only
one sample was infected by just UCBSV. It was compared with both positive and negative
controls. As the negative control, water was used.
2.5. Whitefly, Bemisia tabaci transmission in screen house
Whiteflies were collected from colonies (explained under insect culture) and attached to the tip
of the cassava leaf showing typical symptoms of EACMV to feed for 24 hr within a leaf clip-
cage. Viruliferous B. tabaci adults were then shifted onto disease-free cassava plants of cv.
Page 34
23
Kiroba, for 24 hr of virus inoculation. Thus, the object plants were inoculated one time using a
minimum of 13-20 whiteflies.
Thirty days after inoculation of the healthy plant with the viruliferous adult B. tabaci the cassava
symptoms were determined and its profiles by PCR. By using protocol described under 3.2.2 and
3.3.3, the DNA was extracted, and virus was detected from individual infected cassava plants
using the primers and PCR procedures.
Figure 9. Cassava plants inoculated with EACMV and control plants
3.3. Effect of temperature on CBSV
Cassava fields were observed and plants infected by CBSD were selected and cuttings collected
from Kibaha. The cuttings were taken from the same stem. During collection of the cuttings,
samples were taken for virus detection. Complementary DNA (cDNA) was generated using RT-
PCR. The cDNA was subjected to PCR. The result showed that all of the samples were infected
by CBSVs. These cuttings were then used for the study of the effect of temperature on CBSV
severity and virus level of accumulation. They were planted in plastic pots and kept in the green
house for 4 to 5 weeks. After they started to express symptoms, the diseased plants were placed
in the growth chamber set up at a specific temperature (18-23 and 33oC). The plants were then
observed every day and the data on severity was recorded. A month after a plant was placed in
the growth chamber; which was adjusted according to figure 8 PCR was conducted to know the
level of virus accumulation. The result was compared for different temperatures.
Page 35
24
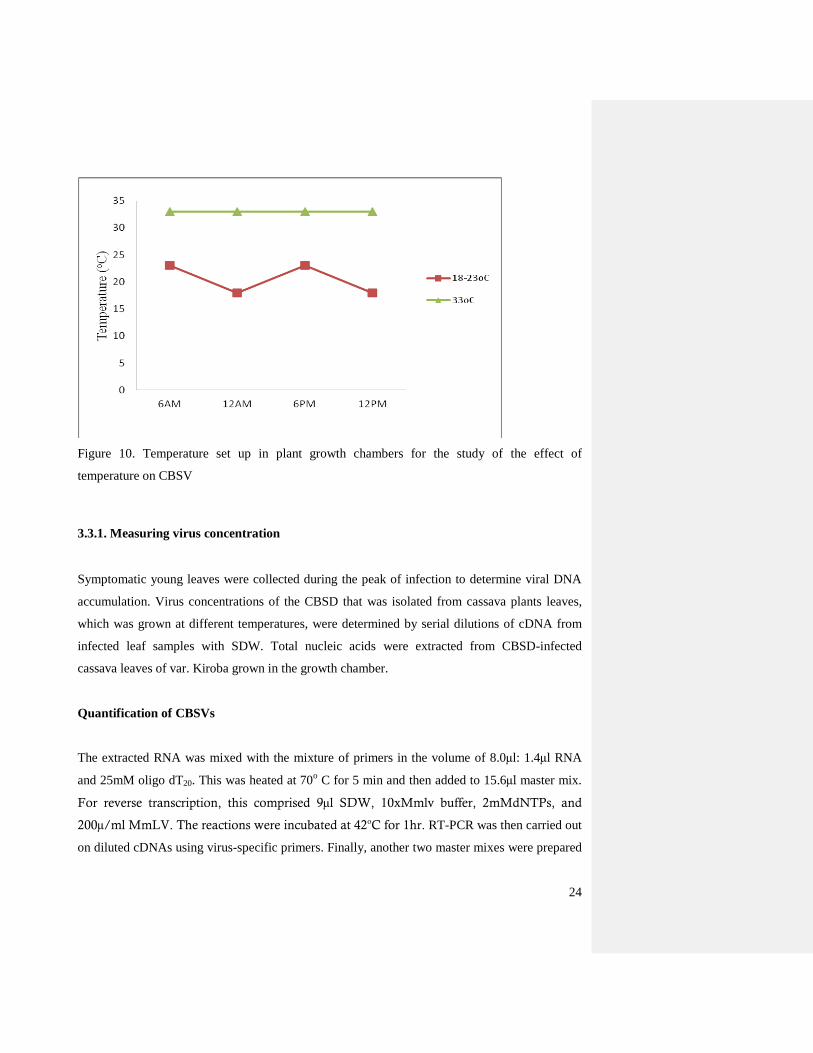
Figure 10. Temperature set up in plant growth chambers for the study of the effect of
temperature on CBSV
3.3.1. Measuring virus concentration
Symptomatic young leaves were collected during the peak of infection to determine viral DNA
accumulation. Virus concentrations of the CBSD that was isolated from cassava plants leaves,
which was grown at different temperatures, were determined by serial dilutions of cDNA from
infected leaf samples with SDW. Total nucleic acids were extracted from CBSD-infected
cassava leaves of var. Kiroba grown in the growth chamber.
Quantification of CBSVs
The extracted RNA was mixed with the mixture of primers in the volume of 8.0µl: 1.4µl RNA
and 25mM oligo dT20. This was heated at 70o C for 5 min and then added to 15.6µl master mix.
For reverse transcription, this comprised 9µl SDW, 10xMmlv buffer, 2mMdNTPs, and
200µ/ml MmLV. The reactions were incubated at 42oC for 1hr. RT-PCR was then carried out
on diluted cDNAs using virus-specific primers. Finally, another two master mixes were prepared
Page 36
25
separately including SYBR green (fluorescing DNA staining dye), ROX solution, specific
primers, and SDW reagents. This was used for virus quantification with 2µl of DNA template.
3.3.2 Evaluation of the progression of diseased plants in the growing chambers
The disease development was assessed weekly and made by visual diagnosis according to
Fargette et al. (1990) and Hillocks et al. (1999). This was based on the observation of the
characteristics of symptoms expressed on the plants such as mosaic, leaf deformation, yellowing
and stunting symptoms appearing on the leaves. The CBSD foliar symptom scoring was rated on
a five-point scale.
1. No symptoms on leaves or stems;
2. Mild/slight vein yellowing or chlorotic blotches on leaves, no brown streaks/lesions on green
stem portions;
3. Mild/slight vein yellowing or chlorotic blotches on leaves mild brown streaks/lesions on green
stem portions;
4. Severe/extensive vein yellowing or chlorotic blotches on leaves, severe brown streaks/lesions
on green stem portions, no defoliation;
5. Severe/extensive vein yellowing or chlorotic blotches on leaves, severe brown streaks/lesions
on green stem portions, defoliation, stem dieback or stunting.
3.4. Data analysis
After data collection, the data was recorded in Excel. The data were then organized and analyzed
using the software SAS. Further, it was organized in to tables and figures and then analyzed
qualitatively and quantitatively. For graphics and tables, Excel was used. Where differences were
observed in developmental parameters and fecundity, these were compared using analysis of
variance (ANOVA). Duncan’s multiple range tests at a significance level of 95% were used to
separate the difference between means. Moreover, the developmental biology parameters were
calculated using the Birch (1948) method.
Page 37
26
Those parameters calculated included intrinsic rate of natural increase (rm), net reproductive rate
(R0) and mean generation time (T). They were calculated from the survival rate (lx) of the
immature and adult stages and from the age-specific oviposition rate (mx). The calculation was
done based on the following equations:
where: x = the age in days. In this, study the female sex ratio was
assumed half for the calculation.
CBSVs quantification was calculated by using the following formula described by Livak and
Schmittgen (2001) and Zhang et al. (2013) that depends on the cycle threshold (Ct) method. Ct =
2 - ΔΔCt
: 2 – [(Ct target gene) – (Ct reference gene)] – [(Ct mean target gene) – (Ct meant
reference gene)]. The quantitative real-time PCR data was used here.
Where: Ct = threshold cycle, ΔΔCt = mean fold change
Page 38
27
4. RESULTS AND DISCUSSION
4.1. Effect of contrasting temperatures on immature development of Bemisia tabaci
The effect of contrasting temperature on the developmental biology of B. tabaci was studied
under laboratory conditions. B. tabaci was reared on cassava at four constant temperatures
(18, 23, 28, and 33o
C) in a plant growth chamber. However, eggs at 33o
C on cassava plants
(with a total of 166 in the first trial and 67 in the second trial) both failed to produce adults
thus all died at different stages. Therefore, this was not included in the analysis of the data. It
was assumed that although these high temperatures are experienced during the hottest period
of the day in coastal Tanzania, they are not sustained throughout the day. Olivier et al. (2006)
in his modeling of B. tabaci Q biotype reported that the optimum immature development is at
32.5o
C. However, in this study the temperature gap was large and optimum mature
development seems more than 28oC.
There was a significant difference between immature developmental times at contrasting
temperatures studied in the present study (Table 1). Developmental time decreased as
temperature increased. For instance, the duration of egg development differed from 12.2 days
at 18o
C to 6.2 days at 28o
C. Comparing between the instars and egg developmental time, the
fourth instars and eggs took longer to develop (egg development F = 123.45; d.f. = 2, 12; P <
0.0001 and 4th
instars development F = 13.62; d.f. = 2, 12; P = 0.0008). Similar results were
reported by other researchers (Sohani et al., 2007; Xie et al., 2010). The newly laid eggs of B.
tabaci are pearly white then turn to pale yellow but when approaching hatching turned to dark
brown at each temperature studied.
Furthermore, the developmental time of the first intar nymphs and adults were different
among the different temperatures examined in the current study. On the other hand, the
developmental period of second instar, third instar and fourth instar nymphs reared on cassava
at 23 and 28o C were not significantly different. It was, however, significantly different
compared with those reared at 18oC.
Page 39
28
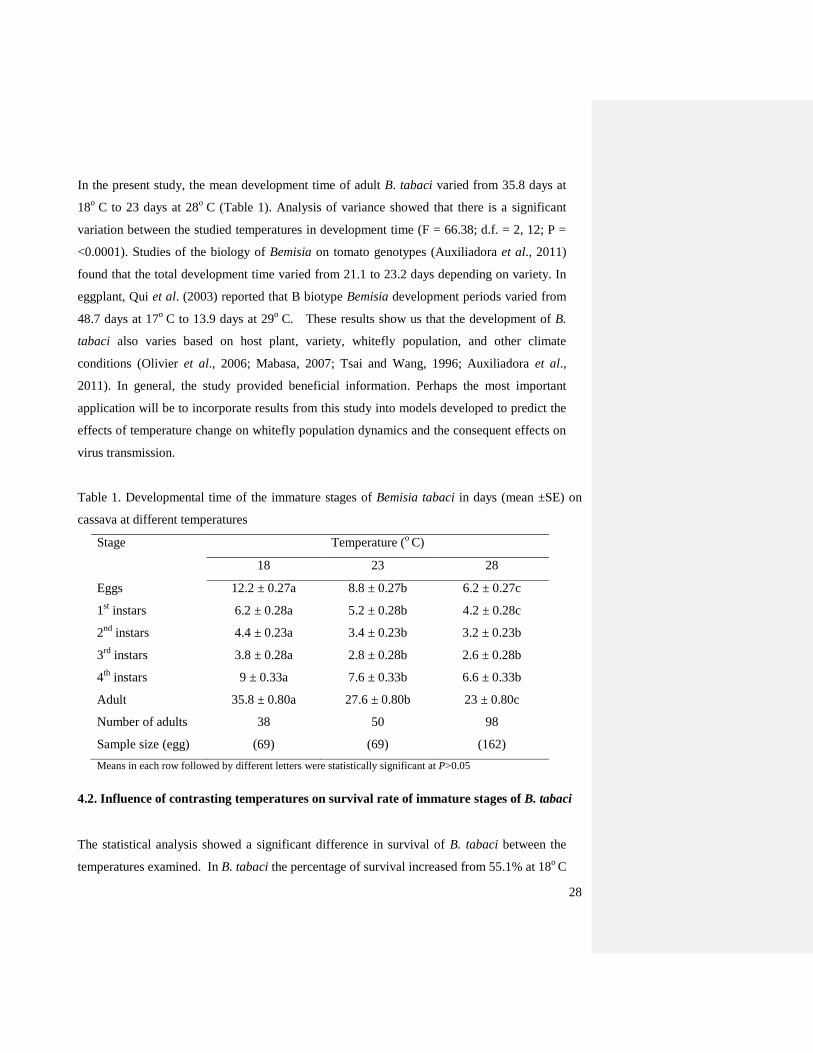
In the present study, the mean development time of adult B. tabaci varied from 35.8 days at
18o
C to 23 days at 28o
C (Table 1). Analysis of variance showed that there is a significant
variation between the studied temperatures in development time (F = 66.38; d.f. = 2, 12; P =
<0.0001). Studies of the biology of Bemisia on tomato genotypes (Auxiliadora et al., 2011)
found that the total development time varied from 21.1 to 23.2 days depending on variety. In
eggplant, Qui et al. (2003) reported that B biotype Bemisia development periods varied from
48.7 days at 17o
C to 13.9 days at 29o
C. These results show us that the development of B.
tabaci also varies based on host plant, variety, whitefly population, and other climate
conditions (Olivier et al., 2006; Mabasa, 2007; Tsai and Wang, 1996; Auxiliadora et al.,
2011). In general, the study provided beneficial information. Perhaps the most important
application will be to incorporate results from this study into models developed to predict the
effects of temperature change on whitefly population dynamics and the consequent effects on
virus transmission.
Table 1. Developmental time of the immature stages of Bemisia tabaci in days (mean ±SE) on
cassava at different temperatures
Stage Temperature (o
C)
18 23 28
Eggs 12.2 ± 0.27a 8.8 ± 0.27b 6.2 ± 0.27c
1st instars 6.2 ± 0.28a 5.2 ± 0.28b 4.2 ± 0.28c
2nd
instars 4.4 ± 0.23a 3.4 ± 0.23b 3.2 ± 0.23b
3rd
instars 3.8 ± 0.28a 2.8 ± 0.28b 2.6 ± 0.28b
4th
instars 9 ± 0.33a 7.6 ± 0.33b 6.6 ± 0.33b
Adult 35.8 ± 0.80a 27.6 ± 0.80b 23 ± 0.80c
Number of adults 38 50 98
Sample size (egg) (69) (69) (162)
Means in each row followed by different letters were statistically significant at P>0.05
4.2. Influence of contrasting temperatures on survival rate of immature stages of B. tabaci
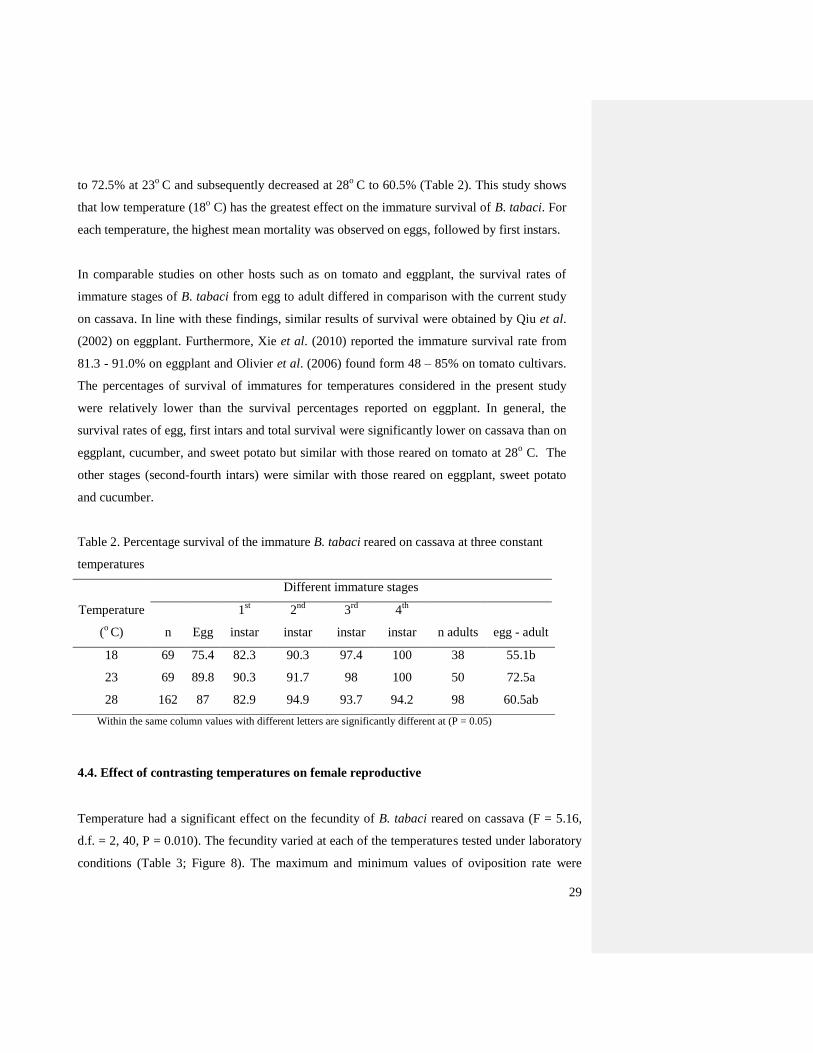
The statistical analysis showed a significant difference in survival of B. tabaci between the
temperatures examined. In B. tabaci the percentage of survival increased from 55.1% at 18o
C
Page 40
29
to 72.5% at 23o
C and subsequently decreased at 28o
C to 60.5% (Table 2). This study shows
that low temperature (18o C) has the greatest effect on the immature survival of B. tabaci. For
each temperature, the highest mean mortality was observed on eggs, followed by first instars.
In comparable studies on other hosts such as on tomato and eggplant, the survival rates of
immature stages of B. tabaci from egg to adult differed in comparison with the current study
on cassava. In line with these findings, similar results of survival were obtained by Qiu et al.
(2002) on eggplant. Furthermore, Xie et al. (2010) reported the immature survival rate from
81.3 - 91.0% on eggplant and Olivier et al. (2006) found form 48 – 85% on tomato cultivars.
The percentages of survival of immatures for temperatures considered in the present study
were relatively lower than the survival percentages reported on eggplant. In general, the
survival rates of egg, first intars and total survival were significantly lower on cassava than on
eggplant, cucumber, and sweet potato but similar with those reared on tomato at 28o C. The
other stages (second-fourth intars) were similar with those reared on eggplant, sweet potato
and cucumber.
Table 2. Percentage survival of the immature B. tabaci reared on cassava at three constant
temperatures
Temperature
(o C)
Different immature stages
n Egg
1st
instar
2nd
instar
3rd
instar
4th
instar n adults egg - adult
18 69 75.4 82.3 90.3 97.4 100 38 55.1b
23 69 89.8 90.3 91.7 98 100 50 72.5a
28 162 87 82.9 94.9 93.7 94.2 98 60.5ab
Within the same column values with different letters are significantly different at (P = 0.05)
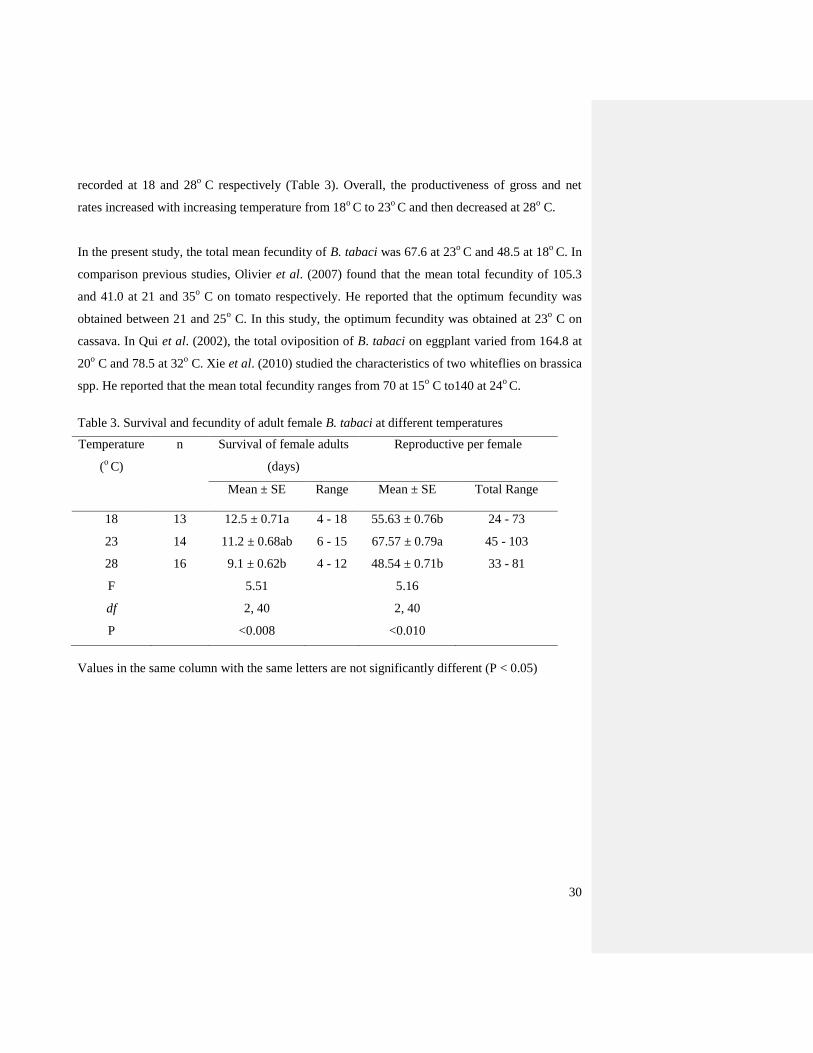
4.4. Effect of contrasting temperatures on female reproductive
Temperature had a significant effect on the fecundity of B. tabaci reared on cassava (F = 5.16,
d.f. = 2, 40, P = 0.010). The fecundity varied at each of the temperatures tested under laboratory
conditions (Table 3; Figure 8). The maximum and minimum values of oviposition rate were
Page 41
30
recorded at 18 and 28o
C respectively (Table 3). Overall, the productiveness of gross and net
rates increased with increasing temperature from 18o
C to 23o
C and then decreased at 28o C.
In the present study, the total mean fecundity of B. tabaci was 67.6 at 23o
C and 48.5 at 18o
C. In
comparison previous studies, Olivier et al. (2007) found that the mean total fecundity of 105.3
and 41.0 at 21 and 35o C on tomato respectively. He reported that the optimum fecundity was
obtained between 21 and 25o C. In this study, the optimum fecundity was obtained at 23
o C on
cassava. In Qui et al. (2002), the total oviposition of B. tabaci on eggplant varied from 164.8 at
20o C and 78.5 at 32
o C. Xie et al. (2010) studied the characteristics of two whiteflies on brassica
spp. He reported that the mean total fecundity ranges from 70 at 15o C to140 at 24
o C.
Table 3. Survival and fecundity of adult female B. tabaci at different temperatures
Values in the same column with the same letters are not significantly different (P < 0.05)
Temperature
(o C)
n Survival of female adults
(days)
Reproductive per female
Mean ± SE Range Mean ± SE Total Range
18 13 12.5 ± 0.71a 4 - 18 55.63 ± 0.76b 24 - 73
23 14 11.2 ± 0.68ab 6 - 15 67.57 ± 0.79a 45 - 103
28 16 9.1 ± 0.62b 4 - 12 48.54 ± 0.71b 33 - 81
F 5.51 5.16
df 2, 40 2, 40
P <0.008 <0.010
Page 43
32
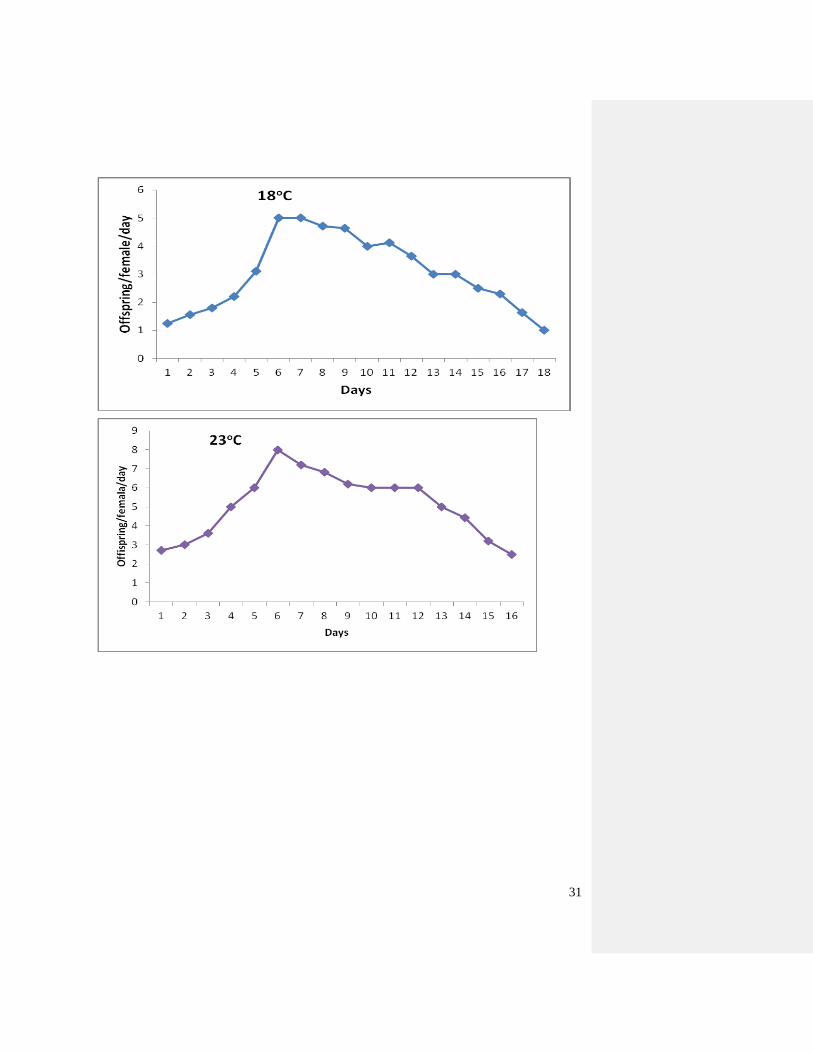
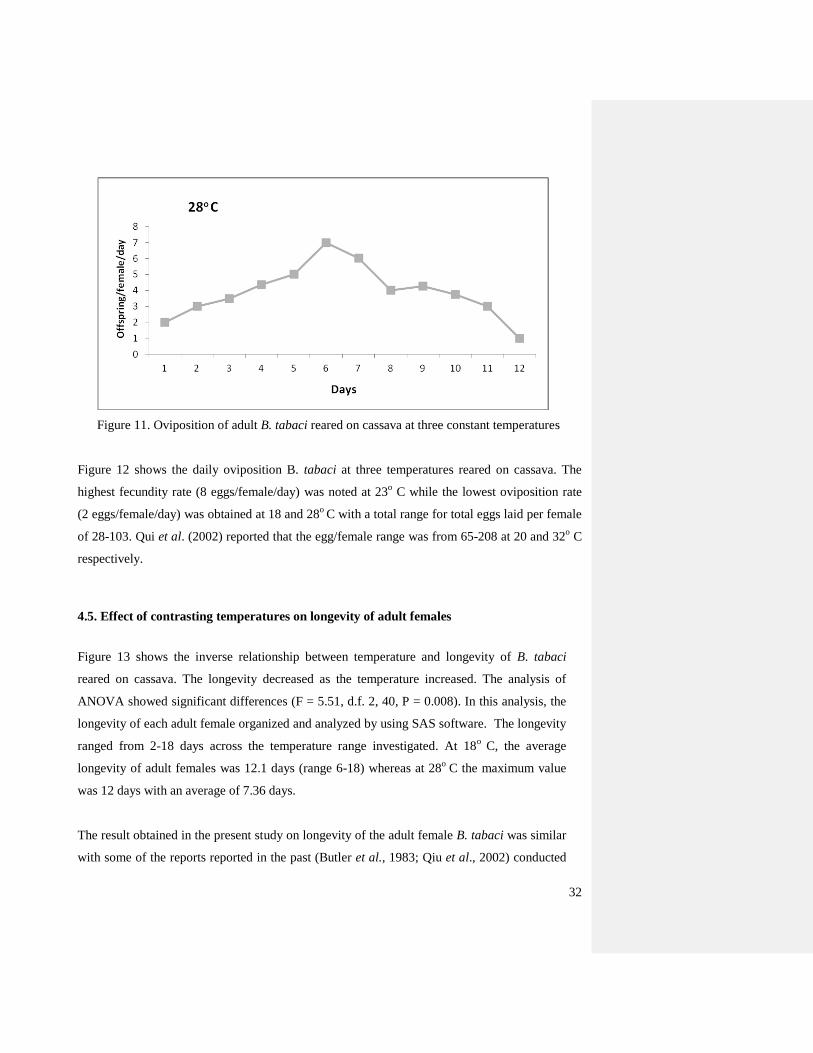
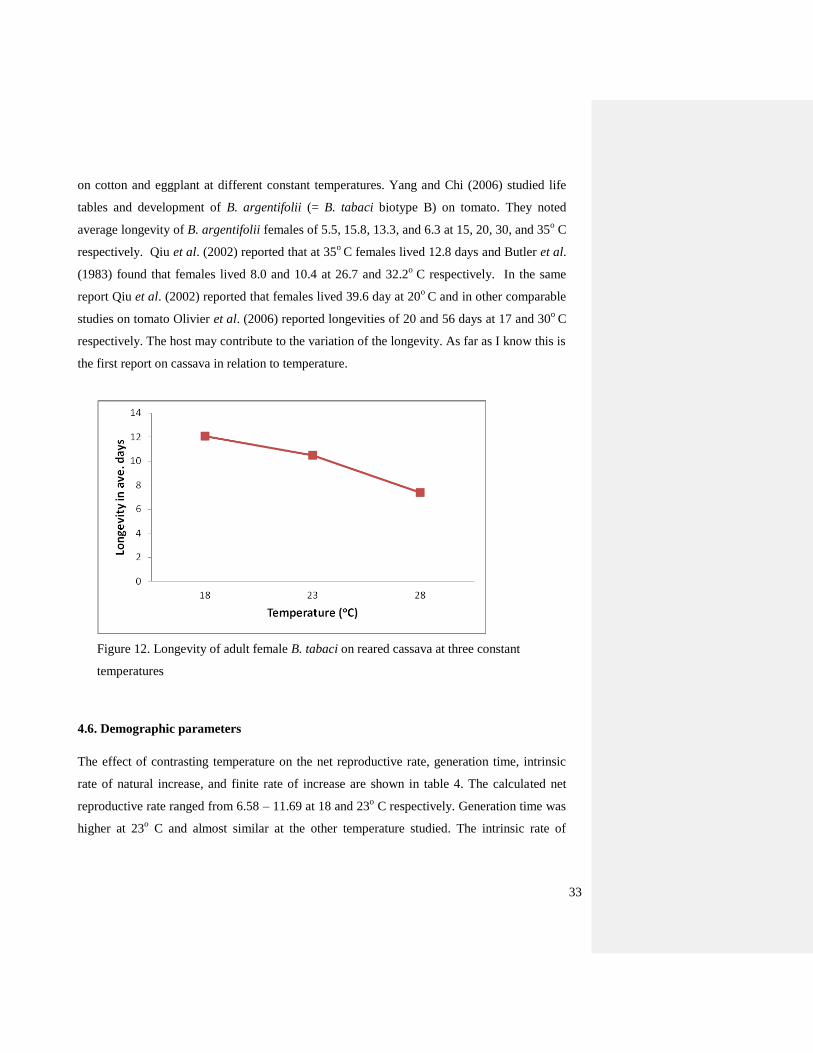
Figure 11. Oviposition of adult B. tabaci reared on cassava at three constant temperatures
Figure 12 shows the daily oviposition B. tabaci at three temperatures reared on cassava. The
highest fecundity rate (8 eggs/female/day) was noted at 23o C while the lowest oviposition rate
(2 eggs/female/day) was obtained at 18 and 28o
C with a total range for total eggs laid per female
of 28-103. Qui et al. (2002) reported that the egg/female range was from 65-208 at 20 and 32o C
respectively.
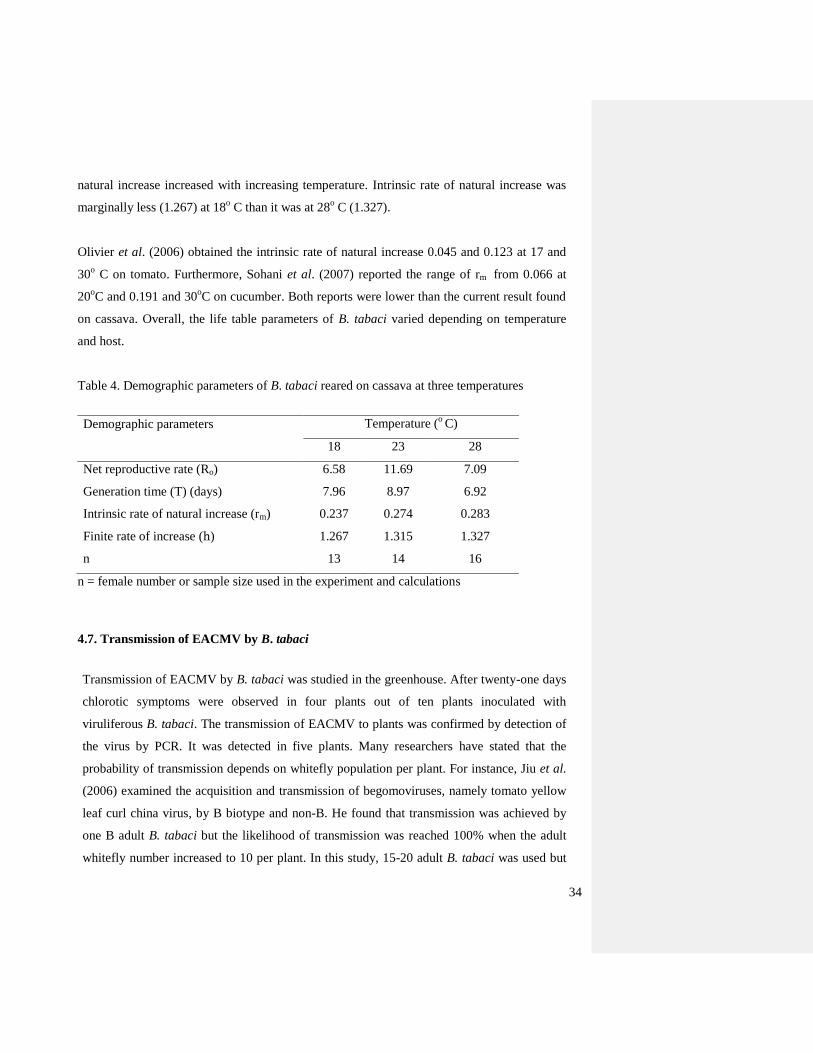
4.5. Effect of contrasting temperatures on longevity of adult females
Figure 13 shows the inverse relationship between temperature and longevity of B. tabaci
reared on cassava. The longevity decreased as the temperature increased. The analysis of
ANOVA showed significant differences (F = 5.51, d.f. 2, 40, P = 0.008). In this analysis, the
longevity of each adult female organized and analyzed by using SAS software. The longevity
ranged from 2-18 days across the temperature range investigated. At 18o
C, the average
longevity of adult females was 12.1 days (range 6-18) whereas at 28o
C the maximum value
was 12 days with an average of 7.36 days.
The result obtained in the present study on longevity of the adult female B. tabaci was similar
with some of the reports reported in the past (Butler et al., 1983; Qiu et al., 2002) conducted
Page 44
33
on cotton and eggplant at different constant temperatures. Yang and Chi (2006) studied life
tables and development of B. argentifolii (= B. tabaci biotype B) on tomato. They noted
average longevity of B. argentifolii females of 5.5, 15.8, 13.3, and 6.3 at 15, 20, 30, and 35o C
respectively. Qiu et al. (2002) reported that at 35o
C females lived 12.8 days and Butler et al.
(1983) found that females lived 8.0 and 10.4 at 26.7 and 32.2o
C respectively. In the same
report Qiu et al. (2002) reported that females lived 39.6 day at 20o
C and in other comparable
studies on tomato Olivier et al. (2006) reported longevities of 20 and 56 days at 17 and 30o
C
respectively. The host may contribute to the variation of the longevity. As far as I know this is
the first report on cassava in relation to temperature.
Figure 12. Longevity of adult female B. tabaci on reared cassava at three constant
temperatures
4.6. Demographic parameters
The effect of contrasting temperature on the net reproductive rate, generation time, intrinsic
rate of natural increase, and finite rate of increase are shown in table 4. The calculated net
reproductive rate ranged from 6.58 – 11.69 at 18 and 23o C respectively. Generation time was
higher at 23o C and almost similar at the other temperature studied. The intrinsic rate of
Page 45
34
natural increase increased with increasing temperature. Intrinsic rate of natural increase was
marginally less (1.267) at 18o C than it was at 28
o C (1.327).
Olivier et al. (2006) obtained the intrinsic rate of natural increase 0.045 and 0.123 at 17 and
30o C on tomato. Furthermore, Sohani et al. (2007) reported the range of rm from 0.066 at
20oC and 0.191 and 30
oC on cucumber. Both reports were lower than the current result found
on cassava. Overall, the life table parameters of B. tabaci varied depending on temperature
and host.
Table 4. Demographic parameters of B. tabaci reared on cassava at three temperatures
Demographic parameters
Temperature (o
C)
18 23 28
Net reproductive rate (Ro) 6.58 11.69 7.09
Generation time (T) (days) 7.96 8.97 6.92
Intrinsic rate of natural increase (rm) 0.237 0.274 0.283
Finite rate of increase (h) 1.267 1.315 1.327
n 13 14 16
n = female number or sample size used in the experiment and calculations
4.7. Transmission of EACMV by B. tabaci
Transmission of EACMV by B. tabaci was studied in the greenhouse. After twenty-one days
chlorotic symptoms were observed in four plants out of ten plants inoculated with
viruliferous B. tabaci. The transmission of EACMV to plants was confirmed by detection of
the virus by PCR. It was detected in five plants. Many researchers have stated that the
probability of transmission depends on whitefly population per plant. For instance, Jiu et al.
(2006) examined the acquisition and transmission of begomoviruses, namely tomato yellow
leaf curl china virus, by B biotype and non-B. He found that transmission was achieved by
one B adult B. tabaci but the likelihood of transmission was reached 100% when the adult
whitefly number increased to 10 per plant. In this study, 15-20 adult B. tabaci was used but
Page 46
35
the disease were observed on some of the plants. These results are similar to those obtained
elsewhere, in which infection rates are typically less than 100% where whiteflies numbers of
between 10 and 50 are used (Dubern, 1994).
4.8. Effect of temperature on CBSV
4.8.1. Effect of temperature on CBSV symptom development
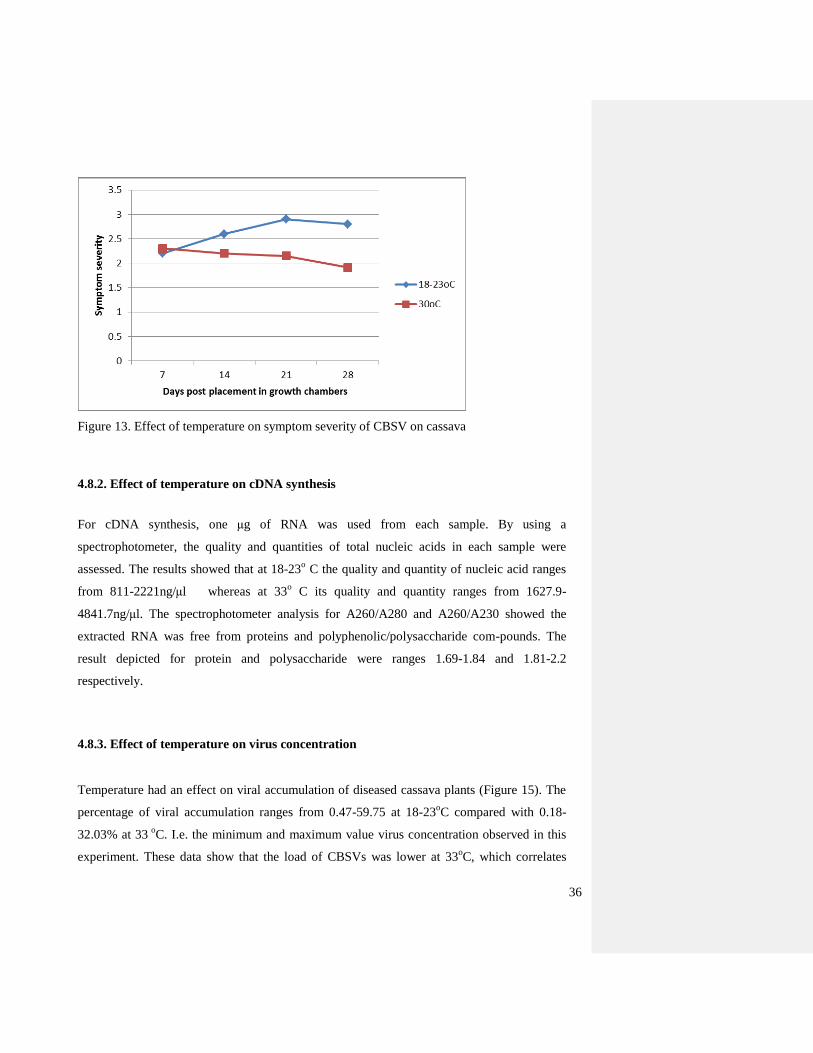
CBSD symptom development varied at different temperatures examined. The symptoms
observed during the period of study included mild/slight vein yellowing, chlorotic blotches on
leaves, mild brown streaks and lesions on green stem portions. The development of disease
was greater at lower temperature, namely from 18 – 23o C, while the disease symptom
development was less at 33o C (Figure 14). After four weeks, at 33
o C the maximum symptom
severity was one third lower than the level of symptom severity observed at 18 – 23o
C. In the
present study, at 33o C most of the newly emerged leaves showed no symptoms at the later
stages.
Studies of the effect of temperature on CBSV have yet to be published elsewhere, but studies on
geminivirus-induced RNA silencing in plants showed that temperature had an effect on the level
of symptom severity (Chellappan et al., 2005). According to these studies when the temperature
rose from 25-30o C the cassava geminivirus –induced RNA silencing was increased, this resulted
in less symptom expression in newly developed leaves. As cited by Mbanzibwa (2011) from
Nicols (1950) low temperature at high altitude (>3500ft) in winter resulted in severe CBSD
symptoms and finally death of plants. This shows that temperature has the ability to change the
effect of infection of viruses such as the CBSVs on cassava plants. The preliminary results
presented here suggest that further research is required on the effect of temperature-induced gene
silencing of CBSVs at a range of temperatures.
Page 47
36
Figure 13. Effect of temperature on symptom severity of CBSV on cassava
4.8.2. Effect of temperature on cDNA synthesis
For cDNA synthesis, one µg of RNA was used from each sample. By using a
spectrophotometer, the quality and quantities of total nucleic acids in each sample were
assessed. The results showed that at 18-23o C the quality and quantity of nucleic acid ranges
from 811-2221ng/µl whereas at 33o C its quality and quantity ranges from 1627.9-
4841.7ng/µl. The spectrophotometer analysis for A260/A280 and A260/A230 showed the
extracted RNA was free from proteins and polyphenolic/polysaccharide com-pounds. The
result depicted for protein and polysaccharide were ranges 1.69-1.84 and 1.81-2.2
respectively.
4.8.3. Effect of temperature on virus concentration
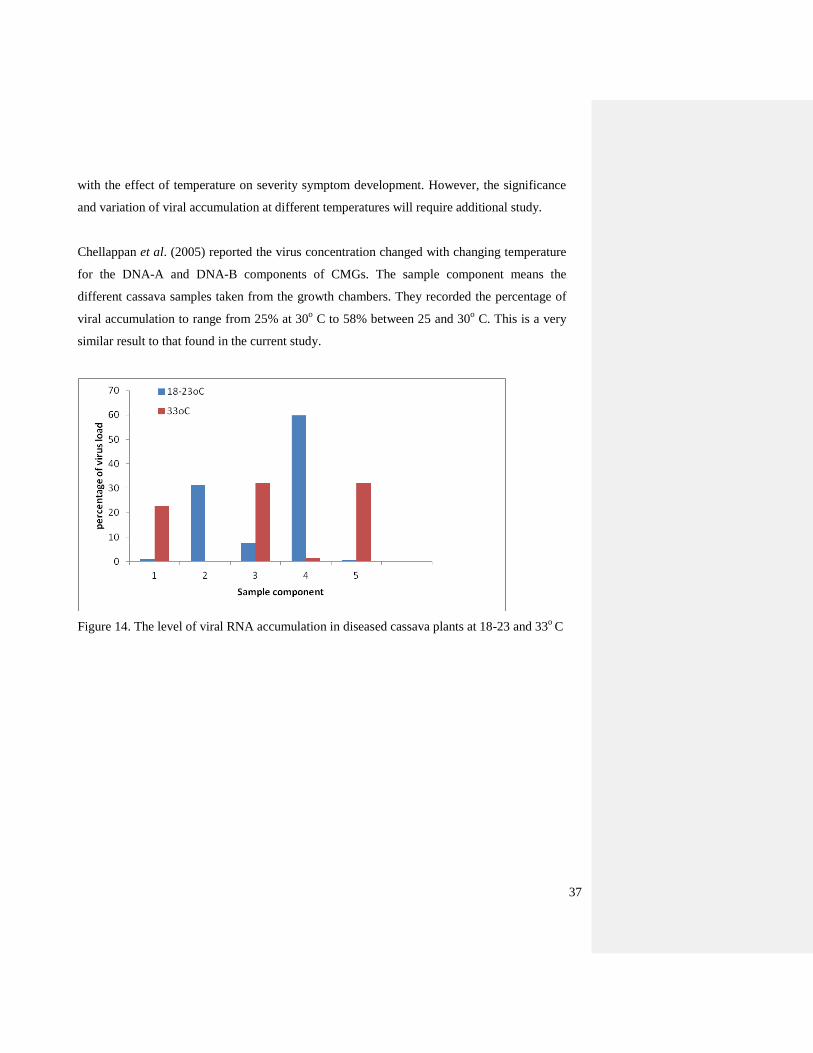
Temperature had an effect on viral accumulation of diseased cassava plants (Figure 15). The
percentage of viral accumulation ranges from 0.47-59.75 at 18-23oC compared with 0.18-
32.03% at 33 o
C. I.e. the minimum and maximum value virus concentration observed in this
experiment. These data show that the load of CBSVs was lower at 33oC, which correlates
Page 48
37
with the effect of temperature on severity symptom development. However, the significance
and variation of viral accumulation at different temperatures will require additional study.
Chellappan et al. (2005) reported the virus concentration changed with changing temperature
for the DNA-A and DNA-B components of CMGs. The sample component means the
different cassava samples taken from the growth chambers. They recorded the percentage of
viral accumulation to range from 25% at 30o C to 58% between 25 and 30
o C. This is a very
similar result to that found in the current study.
Figure 14. The level of viral RNA accumulation in diseased cassava plants at 18-23 and 33o
C
Page 49
38
5. CONCLUSIONS AND RECOMMENDATION
The present study builds our knowledge and gives practical information on the effect of
contrasting constant temperatures on biological parameters of Bemisia tabaci reared on
cassava, which is an important staple food crop in many countries of Africa. Similar studies
have been done on other crops, but not so far on cassava, so this study represents the first of
its kind for this crop.
The findings demonstrated the known effects of temperature on the development time,
survival of immature stages, and longevity/fecundity of adult female B. tabaci. The results
revealed that temperature had significant effects on immature development, survivorship,
fecundity, and longevity of Bemisia tabaci. Further, it affects the population growth and
demographic parameters.
This study showed that temperature had a direct effect on Bemisia tabaci developmental
period, fecundity, and longevity when reared on cassava. The developmental time of adult B.
tabaci varied from 23d at 28o
C to 35.8d at 18o C. Developmental time and percentage of
survivorship differed for each of the immature stages at the temperatures, although these
differences were most significant in the egg and 1st instar stages. The fecundity of Bemisia
tabaci ranges from 18 at 18o C to 103 at 23
o C. Longevity of adult B. tabaci decreased with
the increase of temperature. According to the current findings, the optimal temperature for B.
tabaci is 23o
C. In general, the study revealed that the population growth and fecundity of
Bemisia tabaci was highly affected by lower and higher temperature. The calculated life table
parameters (net reproductive rate and intrinsic rate of natural increase) varied depending on
the temperature examined. Although the finding that B. tabaci performed best at 23o
C is not
greatly surprising, since this is very close to the average temperatures of areas of coastal
Tanzania where these whiteflies live, the values obtained will be of great value when
predicting the likely effects of temperature change. Since temperature increases associated
with global warming are very small, it would be beneficial to extend studies of the type
proposed here to include a much larger range of temperatures, covering a smaller overall
range. For example, it would be useful to look at a range such as 20-25 o
C, using intervals of
Page 50
39
0.5 o
C. Additionally, experiments should be undertaken in which temperatures are varied for
similated day and night periods (e.g. 12h at 25o
C [day] followed by 12h at 18o
C [night]). We
might expect such experiments to provide yet more realistic results.
The EACMV transmission ability of Bemisia tabaci and effect of temperature on CBSV
symptom severity and viral concentration were studied under green house and laboratory
conditions. The healthy plants were inoculated by viruliferous Bemisia tabaci and chlorotic
symptoms were observed after 21 days in four plants. The PCR also showed five plants were
infected by EACMV within the period of study. As the temperature increased from 18-23 to
33oC the CBSV symptom severity development and viral concentration decreased. Moreover,
the newly emerged leaves at 33oC showed fewer symptoms than at 18-23
o C. The symptom
observed under laboratory conditions included mild vein yellowing, chlorotic blotches on leaves,
mild brown streaks, and lesions on green stem portions. Similarly, the percentage of viral
concentration varied at the temperatures studied. It ranged from 32.03% to 59.75% at 33 and 18-
23o C respectively. The severity of disease development and viral load positively correlated in
this study.
The present study was limited to four temperatures due to the limitation of time and growth
chambers. Therefore, further studies are recommended on the effect of temperature on Bemisia
tabaci that will use a more comprehensive set of temperatures with smaller intervals, as
described above. Similarly, to validate the significance of the current results and the effect of
temperature on CBSV concentration and virus transmission ability of Bemisia further studies will
be needed.
Page 51
40
6. REFERENCES
Alabi, O. J., Kumar, P. L., and Naidu, R. A. 2011. Cassava mosaic disease: A curse to food
security in Sub-Saharan Africa. Online. APSnet Features. doi:10.1094/APSnetFeature-2011-
0701.
Atwal, A.S. and Bains, S.S., 1974. Applied animal ecology. Kalyani Publ., New Delhi. 245pp.
Auxiliadora, M., Oriani, D. G., Vendramim, J. D., Jensen, C., Bemisia, B. De, & Hemiptera, B.
B. (2011). Biology of Bemisia tabaci ( Genn .) B biotype ( Hemiptera , Aleyrodidae ) on tomato
genotypes, (February), 37–41.
Ayyar, P. Krishna, N. and Margabandhu, V., 1981. Biology of the cotton stem weevil
Pempherulus affinis under controlled physical condition. Bulletin of Entomological Research,
32: 61-82.
Bale, J., Masters, G.J., Hodkinson, I.D., and Awmack, C., 2002. Herbivory in global climate
change research: direct effects of rising temperature on insect herbivores.
Bale, J. S. and Hayward,S. A. L., 2010. Insect overwintering in a changing climate. The Journal
of Experimental Biology 213, 980-994, Birmingham, UK
Berlinger, M.J. and Lebiush-Mordechi, S., 1996. Physical methods for the control of Bemisia. In:
Gerling, D., Mayer, R.T. (Eds.), Bemisia 1995: Taxonomy, Biology, Damage, Control and
Management. Intercept, UK, pp. 617–634.
Birch, L.C., 1975. The influence of temperature on the development of the different stages of
calandra oryzae L. and Rhizopertha dominica Fab. (Coleoptera). Australian Journal of
Experimental Biology, 23: 29-35.
Bonnemaison, L., 1951. Contribution a l'etude des factews provoquant l'apparition des forms
ailees et Sexuees Chezles Aphidinae. Annals Epiphyties, 2: 1-300
Brown, J. K., Frohlich, D. R. and Rosell, R. C., 1995. The sweet potato or silver leaf whiteflies:
Biotypes of Bemisia tabaci or a species complex? Annual Review of Entomology 40: 511-534.
CAB International, Wallingford, 2002. “Cassava Botany and Physiology,” In: R. J. Hillocks, J.
M. Thresh and A. C. Bellotti, Eds., Cassava: Biology, Production and Utilization, pp. 67-89.
doi:10.1079/9780851995243.0067
Cammell, M, Knight, J., 1997. Effects of climate change on the population dynamics of crop
pests. Adv Ecol Res 22: 117–160.
Page 52
41
Carter, S. E., Fresco, L. O., Jones, P. G., and Fairbairn, J. N. 1995. Introduction and diffusion of
cassava in Africa. Intl. Inst. Trop. Agric. Res. Guide No. 49.
Chellappan, P., Vanitharani, R., Ogbe, F., & Fauquet, C. M. (2005). Effect of Temperature on
Geminivirus-Induced RNA Silencing in Plants 1, 138(August), 1828–1841.
doi:10.1104/pp.105.066563.1828
Chinag, H., 1985. Insects and their environment. pp. 128-161. In: Pfadt, R. E. (eds.).
Fundamentals of applied entomology. MacMillan publishing company, NY, USA.
Colvin, J., Omongo, C.A., Maruthi, M.N., Otim–Nape, G.W. & Thresh, J.M. (2004) Dual
begomovirus infections and high Bemisia tabaci populations: two factors driving the spread of a
cassava mosaic disease pandemic. Plant Pathology, 53, 577–584.
Dent, D.R. and Walton, M.P., 1997. Methods in Ecological and Agricultural Entomology.
Wallingford; UK: International Center for Agriculture and Biosciences (CABI) 478p.
De Bruinjn, G. H. and Fresco, C. O., 1989. “The Importance of Cassava in World Food
Production,” Netherland Journal of Agricultural Sciences, Vol. 37, pp. 21-34.
Dubern, J. (1994). Transmission of African cassava mosaic germinivirus by the whitefly
(Bemisia tabaci). Tropical Science 34, 82-91.
FAO (2010). "Cassava Disease in Africa a major threat to food security: Strategic programme
framework 2010–2015." Food and Agriculture Organization of the United Nations, Rome, Italy.
FAOSTAT, 2013. FAOSTAT. Available at http://faostat.fao.org (accessed 23 June 2013;
verified 25 June 2013). Food and Agriculture Organization (FAO) of the United Nations, Rome,
Italy.
Fargette, D., Fauquet, C. and Thouvenel, J.C., 1985. Field studies on the spread of African
cassava mosaic. Ann. Appl. Biol. 106, 285–294.
Fargette, D., Jeger, M., Fauquet, C. and Fishpool, L.D.C., 1994. Analysis of temporal disease
progress of African cassava mosaic virus. Phytopathology 84, 91–98.
Fauquet, C. and Fargette, D., 1990. African cassava mosaic virus: etiology, epidemiology and
control. Plant Dis. 74, 404–411.
Fishpool, L.D.C. and Burban, C., 1994. Bemisia tabaci: the whitefly vector of African cassava
mosaic geminivirus. Trop. Sci. 34, 55–72.
Fishpool, L.D.C. and Burban, C., 1994. Bemisia tabaci: the whitefly vector of African cassava
mosaic geminivirus. Trop. Sci. 34, 55–72.
Formatted: French (France)
Page 53
42
Fishpool, L.D.C., Fauquet, C., Fargette, D., Thouvenel, J.C., Burban, C. and Colvin, J., 1995.
The phenology of Bemisia tabaci (Homoptera: Aleyrodidae) populations on cassava in southern
Cˆote d’Ivoire. Bull. Entomol. Res. 85, 197–207.
Fletcher, B.S., 1989. Temperature development rate relationships of the immature stages and
adults of Tephritid fruit flies. pp. 273-289. In: A.S. Robinson and G. Hopper (eds.). fruit flies,
their biology, natural enemies and control. World crop pests, 3B. Elsalvador, Amesterdam.
Flores G., Luko H., Gerardo A. M. and Manuel C., (n.d). Antifeedant activity of botanical crude
extracts and their fractions on Bemisia tabaci (Homoptera: Aleyrodidae) adults: III. Quassia
amara (Simaroubaceae)
Frédéric, P., Magali, T., Nathalie B., Pierre L., Bernard R., Hélène D. and Jean-Michel L. (2012).
Biological Invasions of Geminiviruses: Case Study of TYLCV and Bemisia tabaci in Reunion
Island. Viruses 3665-3688; doi:10.3390/v4123665. www.mdpi.com/journal/viruses
Gerling, D. and Mayer, R.T., 1996. Bermisia: 1995 Taxonomy, Biology, Damage, Control, and
Management. Intercept, Andover, U.K.
Gibson, R.W., 1994. Long-term absence of symptoms in heat-treated African cassava mosaic
geminivirus-infected resistant cassava plants. Trop. Sci. 34, 154–158.
Gilbert, N. and Raworth, D.A., 1996. Insect and temperature- a general theory. Canadian
Entomology, 128: 1-13.
Hellmann, J.J., 2002. The effect of an environmental change on mobile butterfly larvae and the
nutritional quality of their hosts. Journal of Animal Ecology, 71: 925-936.
Hilje L., Costab H. S. and Stanslyc P. A., 2001. Cultural practices for managing Bemisia tabaci
and associated viral diseases. Crop Protection 20 (2001) 801–812
Hillocks, R. J. (2002). Cassava in Africa, In: Hillocks, R. J, Thresh, J. M, Bellolli, A. C, eds.
Cassava: Biology, Production and Utilization, CAB International, Wallingford. 41-54
Houghton JT, Ding Y, Griggs DJ, Noguer M., 2001. Climate Change 2001: The Scientific Basis.
(Cambridge University Press, Cambridge). 769 p.
IPCC., 2007. Summary for Policymakers. Cambridge University Press. Cambridge. pp 2–18.
Javed, M., 2009. Integrated management of Bemisia tabaci in Pakistan. PhD thesis, south Asia,
Pakistan.
Jervis, M.A. and Copland, M.J.W., 1996. The life cycle. In: Insect Natural Enemies: Practical
Approach to their Study and Evaluation. Chapman and Hall, London. pp. 63-160.
Jones, D.R., 2003. Plant viruses transmitted by whiteflies. Eur. J. Plant Pathol. 109, 195–219.
Page 54
43
Jones, C.J., Milne, D.E., Patterson, R.S., Schreiber, E.T. and Milio, J.A., 1992. Nectar feeding by
Stomoxys calcitrans (Diptera: Muscidae) effect on reproduction and survival. Environmental
Entomology, 21: 141-147.
Kapinga, R., Mafuru, J., Jeremiah, S. C., Rwiza, E., Kamala, R., Mashamba, F., and Mlingi, N.,
2009. "Status of cassava in Tanzania implications for future Research and Development."
Ministry of Agriculture and Cooperatives, Tanzania, Rome, Italy.
Kasana, A. and AliNiazee, M.T., 1994. Effect of constant temperature on development of the
walnut hsuk fly, Rahgoletis completa. Entomologia Expermentalis et Applicata, 73: 247-254
Lebot, V., 2009. "Tropical Root and Tuber Crops Cassava, Sweet Potato, Yams and Aroids,"
CAB International, Wallingford, UK.
Legg, J.P., 1995. The ecology of Bemisia tabaci (Gennadius) (Homoptera: Aleyrodidae) vector
of African cassava mosaic geminivirus in Uganda. Ph.D. Thesis. University of Reading, UK, 183
pp.
Legg.J.P. (1999). Emergency, spread and strategies for controlling the pandemic of cassava
mosaic virus disease in east and central Africa. Crop protection 18(10), 627-637.
Legg, J. P. and Thresh, J. M., 2003. Cassava virus diseases in Africa. In: Proceedings of the First
International Conference on Plant Virology in Sub-Saharan Africa (4–8 June 2001, Ibadan,
Nigeria), IITA, Ibadan, Nigeria, pp. 517–522.
Legg, J. P. and Fauquet, C. M. (2004). Cassava mosaic geminiviruses in Africa. Plant Mol.
Biol.56, 585–599.
Legg, J. P., Owor, B., Sseruwagi, P. & Ndunguru, J. (2006). Cassava mosaic virus disease in
East and Central Africa: Epidemiology and management of a regional pandemic. Plant Virus
Epidemiology, 67, 355-418.
Legg, J. P., Jeremiah, S. C., Obiero, H. M., Maruthi, M. N., Ndyetabula, I., Okao-Okuja, G.,
Bouwmeester, H., Bigirimana, S., Tata-Hangy, W., Gashaka, G., Mkamilo, G., Alicai, T. &
Kumar, P. L. (2011). Comparing the regional epidemiology of the cassava mosaic and cassava
brown streak virus pandemics in Africa. Virus Research, 159, 161-170.
Lennon, A. M., Aiton, M.M. and Harrison, B.D. (1986). Cassava virus from AfricaScottish
Crops Research Institute Report for 1985. p.168.
Manzari, S. and Quicke, D.L.J. (2006) A cladistic analysis of whiteflies, subfamily Aleyrodinae
(Hemiptera: Sternorrhyncha: Aleyrodidae). Journal of Natural History, 40, 2423–2554.
Martin, J.H., Mifsud, D., and Rapisarda, C., 2000. The whiteflies (Hemiptera: Aleyrodidae) of
Europe and the Mediterranean Basin. Bulletin of Entomological Research 90: 407-448.
Page 55
44
Massala, R., 1987. African cassava mosaic in the Congo, its importance, distribution and
methods of control. In: Proceedings of the International Seminar on African Cassava Mosaic
Disease and its Control, Yamoussoukro, Cˆote d’Ivoire, 4–8 May 1987.
CTA/FAO/ORSTOM/IITA/IAPC, pp. 14–19.
Mkamil,O. S. and Jeremiah ,S.C., 2005. Current status of cassava improvement programme in
Tanzania. African Crop Science Conference Proceedings, Vol. 7. pp. 1311-1314
Muhammad, N., 2006. Management strategy of bemisai tabaci (Gennadius) on cotton in the
Punjab Pakistan. PhD thesis, Bhauddin Zakariya University, Multan.
Naranjo , S. E., 2001. Conservation and evaluation of natural enemies in IPM systems for
Bemisia tabaci. Crop Protection , 20 , 835 – 852 .
Nombela, G., and Muñiz, M., 2010. Host Plant Resistance for the Management of Bemisia
tabaci: A Multi-crop Survey with Emphasis on Tomato. Spain, pp. 357-375.
Ntawuruhunga P., Okao-Okuja, G., Bembe1, A., Obambi, M., Armand, J. C. and Legg, J.P.,
2007. Incidence and Severity of Cassava Mosaic Disease in the Republic of Congo. African Crop Science Journal, Vol. 15, No. 1, pp. 1 – 9 Ohnesorge, B., Sharaf, N. and Allawi, T., 1980. Population studies on the tobacco whitefly
Bemisia tabaci Genn. (Homoptera: Aleyrodidae) during the winter season. I. Spatial distribution
on some host plants. Z. Angew. Entomol. 90, 226–232.
Oliveira, M.R.V., Henneberry, T.J. and Anderson, P., 2001. History, current status, and
collaborative research projects for Bemisia tabaci. Crop Prot. 20, 709–723.
Olivier, B., Amandine, L., Clairey, V. and Jacques, F., 2006. Modelling temperature-dependent
bionomics of Bemisia tabaci ( Q-biotype ). France
Otim-Nape, G.W., Bua, A. and Thresh, J.M. Eds., 1997b. Progress in cassava technology
transfer in Uganda NARO/Gatsby/NRI Publication,Chatham Maritime, U K,39 pp
Parmesan, C. and Yohe, G., 2003. A globally coherent fingerprint of climate change impacts
across natural systems, USA. Nature 421: 37–42.
Perring, T.M., 2001. The Bemisia tabaci species complex. Crop Prot. 20, 725–737.
Petzoldt, C. and Seaman, A., 2007. Climate Change Effects on Insects and Pathogens. Fact
Sheet. http://www.climateandfarming.org/clr-cc.php
Poole, R.W., 1974. An introduction to quantitative ecology. McGraw-Hill, New York.
Page 56
45
QIU, B.L., DE BARRO, P.J. and REN, S.X., 2005. Development, survivorship and reproduction
of Eretmocerus sp. nr. Furuhashii (Hymenoptera: Aphelinidae) parasitizing Bemisia tabaci
(Hemiptera: Aleyrodidae) on glabrous and nonglabrous host plants. Bull. Entomol. Res. 95: 313–
319. Rome, Italy.
Silver whitefly: www.ozanimals.com/Insect/Silverleaf-whitefly/Bemisia/tabaci.htm .
Sohani, Z.N., Shishehbor, P., and Kocheili, F. (2007). Thermal effect on the biology and life
table of Bemisia tabaci Gennadus (Homoptera: Aleyroididae).pakistan journal of biological
sciences 10 (22): 4057 – 4062, Ahwaz, Iran.
Southwood, T.R.E., 1978. Ecological Methods with Particular Reference to the Study of Insect
populations, 2nd edn. University Printing House, Cambridge.
Sseruwagi, P., Sserubombwe,W.S. Legg ,J.P. Ndunguru , J. and Thresh, J.M., 2004. Methods of
surveying the incidence and severity of cassava mosaic disease and whitefly vector populations
on cassava in Africa: a review, Virus Research 100 (2004) 129–142
Sseruwagi, P., Legg, J.P. and Otim-Nape, G.W., 2004. Whiteflies as vectors of plant viruses in
cassava and sweetpotato in Africa: Uganda. In: Anderson and Markham.
Sweetpotato Whitefly Bemisia tabaci (Gennadius):
http://entnemdept.ufl.edu/fasulo/whiteflies/wfly0019.htm [accesssed on: 15/05/2013]
Symrnioudis, A.D. Watt, and Whittaker, J.B., 2002. Herbivory in global climate change
research: direct effects of rising temperatures on insect herbivores. Global Change Biology 8:1-
16.
Van Lenteren J.C. (2008). IOBC Internet Book of Biological Control, version 5
Vargas, R.I., Walsh, W.A. Kaneshisa, D., Stark, J.D. and Nishida, T., 2002. Comparative
demography of three Hawaiian fruit flies (Diptera: Tephritidae) at alternating temperatures.
Annals of the Entomological Society of America, 93 (1): 75-81.
Wang, K. H. and Tsai, J. H., 1996. Temperature effect on development and reproduction of
sliverleaf whitefly ( Homoptera: Aleyrodidae) . 89(3) : 375-384. Ann. Entoml . Sac. Am .
Ward, N.L. and Masters, G.J., 2007. Linking climate change and species invasion: an illustration
using insect herbivores. Glob Change Biol 13: 1605–1615.
Weather forecast [online] 13 June. Available at: < http://www.1blueplanet.net/weather/?wc_location=7647520&wc_language=en [Accessed 13
June 2013]
Page 57
46
Xie, M., Wan, F., Chen, Y., & Wu, G. (2011). Effects of temperature on the growth and
reproduction characteristics of Bemisia tabaci B-biotype and Trialeurodes vaporariorum,
135(Zhou 1949), 252–257. doi:10.1111/j.1439-0418.2010.01524.x
Yamamura, K. and Kiritani, K., 1998. A simple method to estimate the potential increase in the
number of generations under global warming in temperate zones. Appl. Ent. and Zool. 33:289-
298.
Yazdani, S.S. and Agarwal, M.L., 1997. Elements of insect ecology. Narosa publishing house,
New Delhi, India.
Zalom, F. Goodell, P. Wilson, L. Barnett, W. and Bentley, W., 1983. Degree-days: the
calculation and use of heat unit in pest management. Division of agriculatural and natural
resources, University of California, Davis, CA, USA. 10pp.
Zhang, J. D., Ruschhaupt, M., & Biczok, R. (2013). ddCt method for qRT – PCR data analysis
The ddCt Algorithm, 1–8.