Page 1
www.elsevier.com/locate/palaeo
Palaeogeography, Palaeoclimatology, Pa
Neogene palaeoceanographic and palaeoclimatic events
inferred from palynological data: Cape Basin off
South Africa, ODP Leg 175
Chioma U. Udeze1, Francisca E. Oboh-IkuenobeT
Department of Geological Sciences and Engineering, University of Missouri-Rolla, Rolla MO 65409, USA
Received 12 February 2004; received in revised form 14 December 2004; accepted 23 December 2004
Abstract
Sites 1085, 1086 and 1087 were drilled off South Africa during Ocean Drilling Program (ODP) Leg 175 to investigate the
Benguela Current System. While previous studies have focused on reconstructing the Neogene palaeoceanographic and
palaeoclimatic history of these sites, palynology has been largely ignored, except for the Late Pliocene and Quaternary. This
study presents palynological data from the upper Middle Miocene to lower Upper Pliocene sediments in Holes 1085A, 1086A
and 1087C that provide complementary information about the history of the area. Abundant and diverse marine palynomorphs
(mainly dinoflagellate cysts), rare spores and pollen, and dispersed organic matter have been recovered. Multivariate statistical
analysis of dispersed organic matter identified three palynofacies assemblages (A, B, C) in the most continuous hole (1085A),
and they were defined primarily by amorphous organic matter (AOM), and to a lesser extent black debris, structured
phytoclasts, degraded phytoclasts, and marine palynomorphs. Ecostratigraphic interpretation based on dinoflagellate cyst,
spore-pollen and palynofacies data allowed us to identify several palaeoceanographic and palaeoclimatic signals. First, the late
Middle Miocene was subtropical, and sediments contained the highest percentages of land-derived organic matter, even though
they are rich in AOM (palynofacies assemblage A). Second, the Late Miocene was cool-temperate and characterized by periods
of intensified upwelling, increase in productivity, abundant and diverse oceanic dinoflagellate cysts, and the highest percentages
of AOM (palynofacies assemblage C). Third, the Early to early Late Pliocene was warm-temperate with some dry intervals
(increase in grass pollen) and intensified upwelling. Fourth, the Neogene bcarbonate crashQ identified in other southern oceans
was recognized in two palynofacies A samples in Hole 1085A that are nearly barren of dinoflagellate cysts: one Middle
Miocene sample (590 mbsf, 13.62 Ma) and one Upper Miocene sample (355 mbsf, 6.5 Ma). Finally, the extremely low
percentages of pollen suggest sparse vegetation on the adjacent landmass, and Namib desert conditions were already in
existence during the late Middle Miocene.
D 2005 Elsevier B.V. All rights reserved.
Keywords: Palynology; Palynofacies; Neogene; Palaeoceanography; Palaeoclimate; Ocean Drilling Program Leg 175
T Corresponding author. Tel.: +1 573 341 6946; fax: +1 573 341 6935.
0031-0182/$ - s
doi:10.1016/j.pa
E-mail addr1 Present addr
laeoecology 219 (2005) 199–223
ee front matter D 2005 Elsevier B.V. All rights reserved.
laeo.2004.12.026
esses: [email protected] (C.U. Udeze)8 [email protected] (F.E. Oboh-Ikuenobe).
ess: Department of Geology and Geophysics, Texas A&M University, College Station, TX 77843, USA.
Page 2
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223200
1. Introduction
The Cape-Argentine Basin was created as a
precursor to the South Atlantic Ocean by the initial
rifting between Africa and South America, which
began during the Late Jurassic. The Walvis Ridge
divides the South Atlantic Ocean adjacent to Africa
into two major features: the Cape Basin and the
Angola Basin. Totally stagnant euxinic conditions
persisted through much of the Early Cretaceous in
both basins (Bolli et al., 1978). Today, the Angola–
Benguela Current system is one of the great upwell-
ing regions in the world. It extends over a consid-
erable portion of the western margin of South Africa,
and is characterized by organic-rich sediments
containing excellent signals of productivity, which
are closely tied to the regional dynamics of
circulation, mixing, and upwelling (Wefer et al.,
1998).
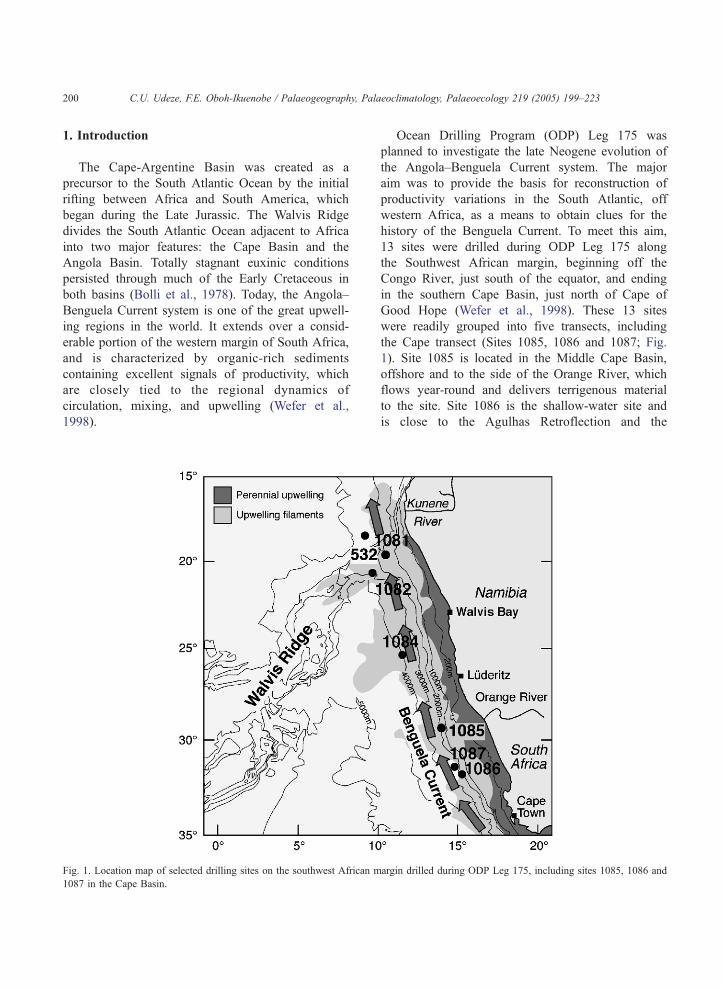
Fig. 1. Location map of selected drilling sites on the southwest African m
1087 in the Cape Basin.
Ocean Drilling Program (ODP) Leg 175 was
planned to investigate the late Neogene evolution of
the Angola–Benguela Current system. The major
aim was to provide the basis for reconstruction of
productivity variations in the South Atlantic, off
western Africa, as a means to obtain clues for the
history of the Benguela Current. To meet this aim,
13 sites were drilled during ODP Leg 175 along
the Southwest African margin, beginning off the
Congo River, just south of the equator, and ending
in the southern Cape Basin, just north of Cape of
Good Hope (Wefer et al., 1998). These 13 sites
were readily grouped into five transects, including
the Cape transect (Sites 1085, 1086 and 1087; Fig.
1). Site 1085 is located in the Middle Cape Basin,
offshore and to the side of the Orange River, which
flows year-round and delivers terrigenous material
to the site. Site 1086 is the shallow-water site and
is close to the Agulhas Retroflection and the
argin drilled during ODP Leg 175, including sites 1085, 1086 and
Page 3
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 201
Subtropical Convergence Zone (Lutjeharms, 1996;
Wefer et al., 1998). Site 1087 is the deep-water site
of the Cape transect. It is located on the continental
rise of southwest Africa and at a crossroad for the
west-wind drift, the Benguela Current and the
Agulhas Retroflection. Sites 1086 and 1087 are
located in the Southern Cape Basin.
The proximity of the Cape sites to the continent
makes them very useful in detecting upwelling
signals and clues to changes in continental climate
(pollen, clay minerals, and terrigenous silt) and sea
level. It is generally believed that variations in
upwelling intensity might lead to changes in
atmospheric pCO2 (Toggweiler, 1999; Bork and
Zonneveld, 2003). However, palynological studies
in the Benguela upwelling region have been limited
to mostly Upper Pliocene and Quaternary sediments
(Davey, 1971; Davey and Rogers, 1975; Bork and
Zonneveld, 2003; Holzwarth et al., 2003). In this
study, 59 samples were obtained from the late
Middle Miocene to early Late Pliocene interval in
Holes 1085A, 1086A and 1087C in order to use
palynomorphs (dinoflagellate cysts, acritarchs,
spores, pollen) and dispersed organic matter (paly-
nofacies) as proxies to (a) interpret biostratigraphy;
(b) infer palaeoclimatic and palaeoceanographic
signals; and (c) detect the Middle to Late Miocene
bcarbonate crashQ (i.e., the depletion of carbonate
material in marine sediments; Lyle et al., 1995)
which has been reported in other southern oceans.
2. Materials and methods
Fifty-nine samples from ODP Leg 175 (34 from
Hole 1085A, 13 from Hole 1086A and 12 from
Hole 1087C) were processed using the standard
palynological techniques of disaggregation and
removal of carbonates and silicates with hydro-
chloric and hydrofluoric acids (Faegri and Iversen,
1989). Centrifugation in ZnBr2 solution was used to
separate the organic matter fraction. After deminer-
alization, the organic residues used for palynomorph
identification were oxidized with Schultze’s solution
and screened using 10 Am sieves. (Sections 3.1 and
4.4.1 discuss how oxidation likely affected the types
of dinoflagellate cysts recovered) Oxidation and
sieving were excluded for kerogen residues used for
palynofacies analyses in order to minimize the
removal of amorphous organic matter (AOM), avoid
the destruction/loss of certain types of organic matter,
and to preserve the colors of the organic material.
Residues were permanently mounted onto slides with
clear casting resin. The slides used for dinoflagellate
cyst analysis were stained with Alizaren Red to
enhance their identification. For routine identification
and description of the palynomorphs, the slides were
carefully examined using transmitted light micro-
scopy. At least 200 specimens were counted in
samples with good recovery of dinoflagellate cysts.
The suprageneric classification of Fensome et al.
(1993) is used and the nomenclature follows Williams
et al. (1998), Strauss et al. (2001), and Head and
Norris (2003).
Transmitted light microscopy was also used for
the identification of dispersed organic matter
(palynodebris) and groups of palynomorphs in
kerogen slides. The following seven types were
identified: AOM, black debris, degraded phyto-
clasts, structured phytoclasts (wood, parenchyma,
cuticle), marine palynomorphs (dinoflagellate cysts,
acritarchs, microforaminiferal inner linings), fungal
remains (spores, hyphae, mycelia), and sporomorphs
(spores and pollen). Our classification scheme is
modified after Tyson (1987, 1995), Boulter (1994),
Batten (1996) and Jaramillo and Oboh-Ikuenobe
(1999). In addition, we were able to refer some
particles to marine palynomorphs by comparison
with Edwards and Andrle (1992) and Head (1997).
Three hundred particles (each with a 5 Am size
cutoff) were point counted per slide, converted to
percentages and subjected to minimum variance
cluster analysis (Q-mode and R-mode) on a data
matrix generated by Euclidean correlation coeffi-
cient (Kovach, 2002). Log transformation (log2)
produced an octave scale that allowed organic
components with low percentage values to play a
bigger role in grouping samples (Beck and Strother,
2003). The Euclidean distance is especially
designed to work with continuous or ratio scales
and the linkage averages all distances between pairs
of objects in different clusters to decide how far
apart they are (Sokal and Michener, 1958). In
addition, data used for cluster analysis were
analyzed using principal components analysis for
comparison purposes. Slides are stored in the
Page 4
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223202
palynological collection in the Palaeontology Labo-
ratory at the University of Missouri-Rolla.
The numbering of sites, holes, cores and samples
follow standard ODP procedures (Wefer et al.,
1998). Age models for the Miocene and Pliocene
sections of ODP Holes 1085A and 1087C were
generated by Westerhold et al. (in press), who used
orbital tuning of an XRFFe intensity to record the
summer insolation astronomical solution of Laskar
(1993). Fine-tuning was done iteratively by correlat-
ing each prominent maximum of the Fe record to the
minima of the target curve. This gave at least one
age control point for each eccentricity cycle. The
shipboard data for biostratigraphic and palaeomag-
netic events (Wefer et al., 1998) provided the
preliminary age model for Sites 1085, 1086 and
1087.
3. Results
3.1. Palynostratigraphy
All 59 samples used for this study were qualita-
tively and quantitatively examined for their palyno-
morph content. None of the samples are barren but the
relative abundance and diversity of the palynomorphs
vary. The most abundant palynomorphs are dinofla-
gellate cysts, which consist of about 80% of the total
palynomorph assemblage and are present in 56
samples. Acritarchs make up about 15% of the total
palynomorph assemblage and are present in 54
samples, while spores and pollen make up about 5%
of the total palynomorph assemblage. The preserva-
tion of the palynomorphs was excellent to occasion-
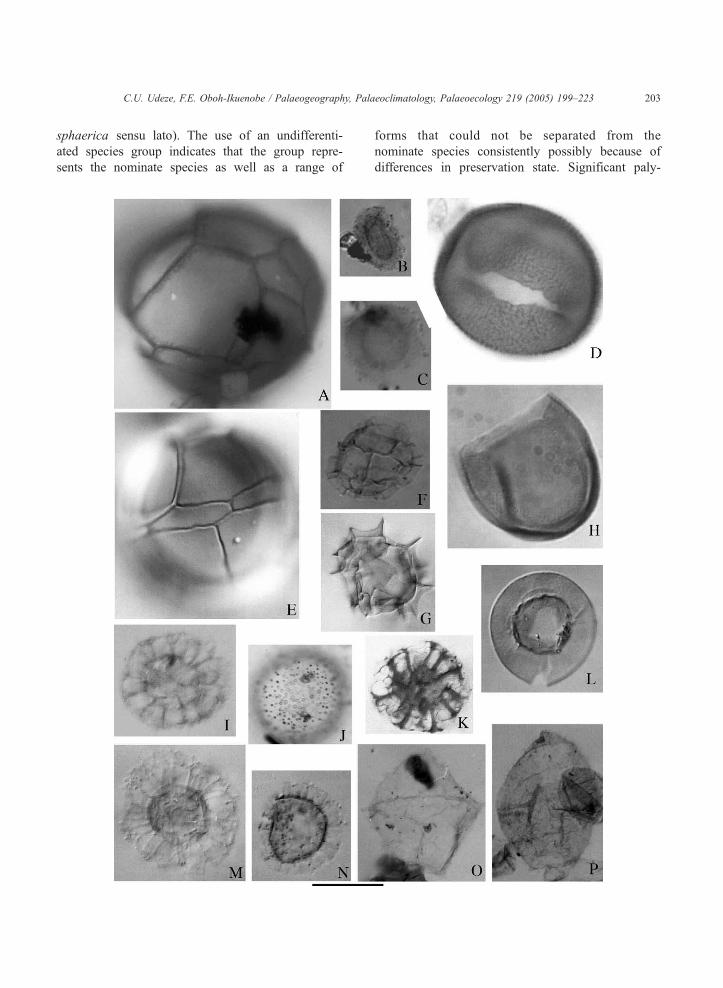
Fig. 2. Photomicrographs of palynomorphs, each identified by sample num
(catalog number 7100); unless noted, specimens are from stained and sieve
and I–P, and 10 Am for figures D and H. (A) Impagidinium sp. A of Wrenn
12 cm, 16N/P. (B) Leiosphaeridia sp. A., uncertain view, mid-focus, 108
rockhallensis, lower focus, 1085A-39X-2, 102 cm, 22M/N. (D) Bitectatodi
23K/L (E) Impagidinium patulum, right lateral view, upper focus, 1086A
view, upper focus, 1085A-15H-6, 26–28 cm, 27K/L. (G) Impagidinium ac
(H) Batiacasphaera sphaerica, uncertain view, mid-focus, 1086A-12H-4
view, mid-focus, 1085A-15H-6, 26–28 cm, 16P/Q. (J) Unidentified sp. A
Reticulatosphaera actinocoronata, uncertain view, mid-focus, 1086A-16H
archeopyle, mid-focus, 1085A-15H-6, 26–28 cm, 22F/G. (M) Nematospha
62 cm, 25M/L. (N) Labyrinthodinium truncatum, dorsal view, mid-focus,
mid-focus, 1085A-31H-4, 52 cm, 20U/V. (P) Trinovantedinium sp., dorsa
ally poor. The appendix lists the taxa identified in this
study.
Gonyaulacalean cysts dominate protoperidinialean
cysts (9:1 ratio), which is in sharp contrast to the
protoperidinialean-dominated assemblages of modern
sediments in this and other upwelling regions (Wall
et al., 1977; Lewis et al., 1990; Powell et al., 1992;
Dale and Fjellsa, 1994; Dale, 1996; Zonneveld et
al., 2001a; Dale et al., 2002). It is very likely that
oxidation of samples with Schultze solution selec-
tively destroyed the majority of the protoperidinia-
lean cysts (Schrank, 1988; Hopkins and McCarthy,
2002); several studies (e.g., Zonneveld et al., 1997,
2001b; Versteegh and Zonneveld, 2002; Esper et al.,
2004) have shown that these cysts are much more
prone to degradation by oxygenated bottom waters
than gonyaulacalean cysts, pollen and spores. It
should be noted that while unoxidized kerogen
slides also contain very few protoperidinialean cysts,
the high amounts of AOM in the sediments
probably obscured some palynomorphs. Notwith-
standing this limitation, the gonyaulacalean cysts
that have been concentrated through the oxidation
process have yielded important biostratigraphic,
palaeoceanographic and palaeoclimatic information
(see Section 4).
Pollen taxa include undifferentiated bisaccate
pollen, Bombacacidites sp. (Bombacaceae), Cheno-
podipolis sp. (Chenopodiaceae), Echitricolporities
spinosus (Compositae), Intratriporopollenites sp.,
Graminidites sp. (Poaceae), Nyssapollenites sp.,
Triatripollenites sp. and Tricolpites sp. The dino-
flagellate cysts and acritarchs are identified as
individual species, undifferentiated taxa and undif-
ferentiated species groups (e.g., Batiacasphaera
ber and location coordinates on the Lovins micro-slide field finder
d slides (#2) unless noted; scale bar is 30 Am for figures A–C, E–G
and Kokinos (1986), uncertain view, mid-focus, 1086A-18H-7, 10–
5A-60X-6, 103 cm, slide 1, 21M/N. (C) Leiosphaeridia sp. cf. L.
nium tepikiense, apical view, lower focus, 1086A-10H-4, 60–62 cm,
-19H-4, 60–62 cm, 29T/U. (F) Impagidinium paradoxum, uncertain
uleatum, uncertain view, upper focus, 1085A-28H-4, 52 cm, 31P/Q.
, 60–62 cm, 33B/C. (I) Nematosphaeropsis labyrinthus, uncertain
., uncertain view, upper focus, 1086A-8H-1, 10–12 cm, 11B/C. (K)
-5, 10–12 cm, 21N/P. (L) Ataxiodinium confusum, dorsal view with
eropsis lemniscata, uncertain view, upper focus, 1086A-10H-4, 60–
1087C-44X-6, 75–77 cm, 16Q/R. (O) Cristadinium sp., dorsal view,
l view, upper focus, 1085A-49X-2, 52 cm, 25R/S.
Page 5
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 203
sphaerica sensu lato). The use of an undifferenti-
ated species group indicates that the group repre-
sents the nominate species as well as a range of
forms that could not be separated from the
nominate species consistently possibly because of
differences in preservation state. Significant paly-
Page 6
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223204
Page 7
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 205
nomorph taxa used for interpretation are illustrated
in Figs. 2–4.
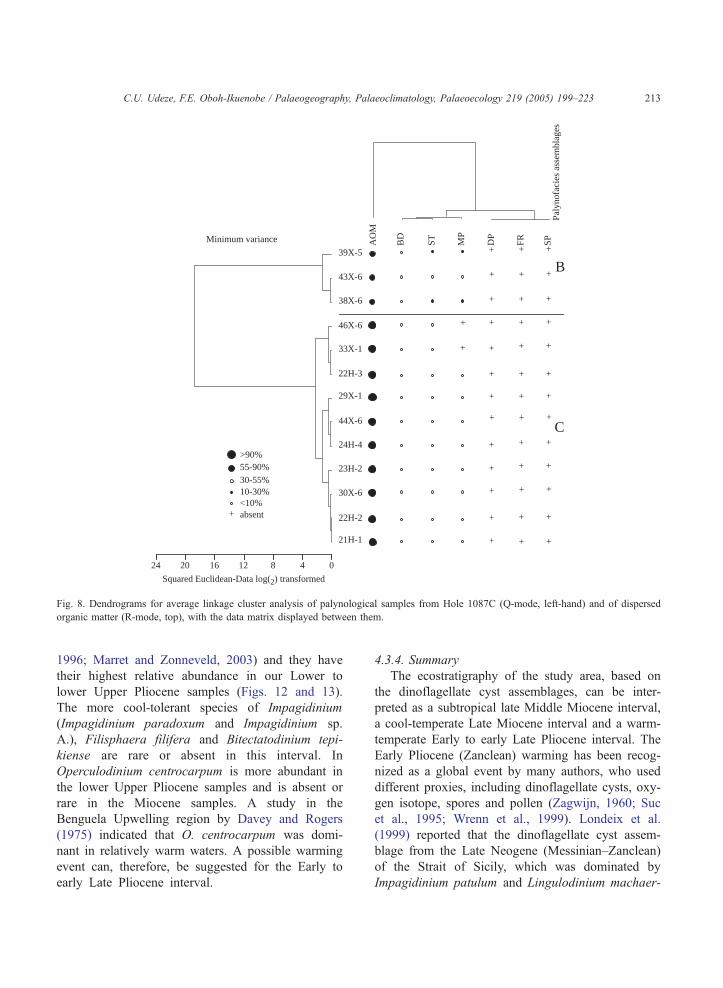
3.2. Palynofacies
Seven types of dispersed organic matter were
identified and they fall into three broad categories,
namely palynomorphs, phytoclasts and unstructured
(amorphous) organic matter (Fig. 5). AOM is the
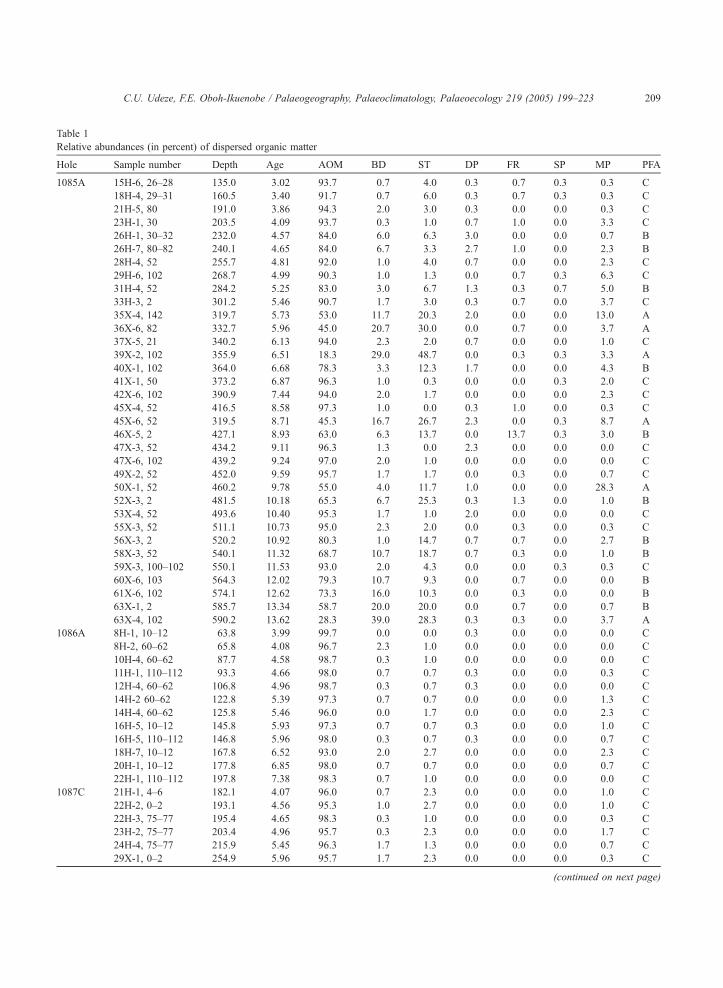
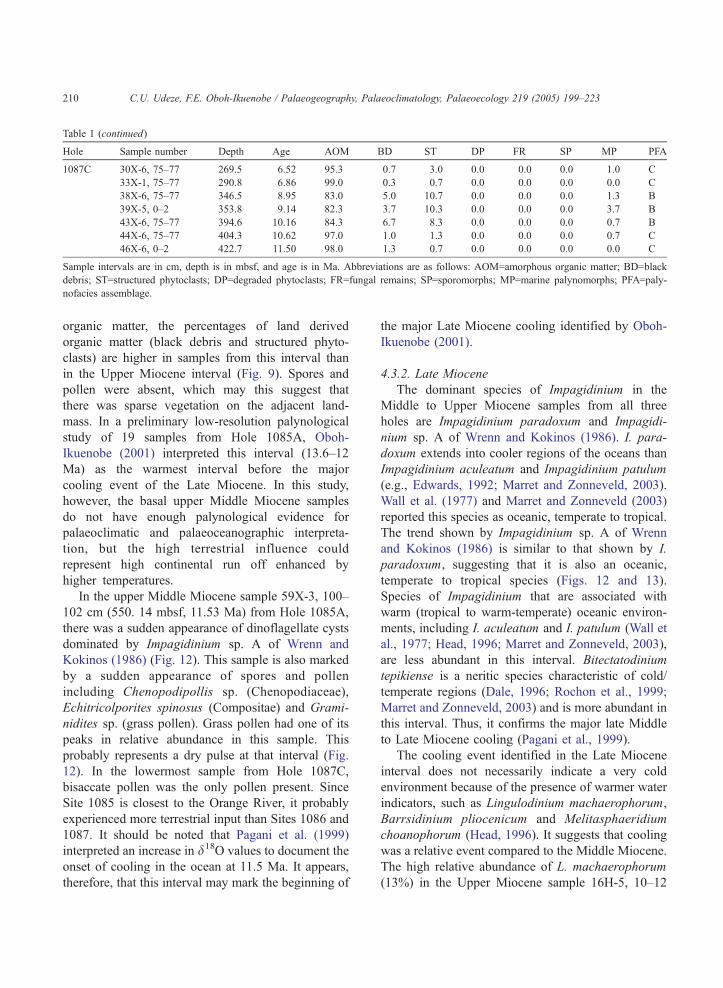
dominant organic material in most samples. Table 1
summarizes the results of the palynofacies analyses
of the samples. Three palynofacies assemblages,
designated A, B and C, are identified in Hole
1085A from cluster analysis (Fig. 6) based on five
statistically important organic components, namely
AOM, black debris, structured phytoclasts, degraded
phytoclasts and marine palynomorphs. The statistical
significance of these components is confirmed by
principal components analysis (Table 2), which
shows that these organic components constitute more
than 80% of the total variance in the sample set on
the first two axes. On the cluster analyses diagrams
in Fig. 6, the top dendrogram (R-mode) groups the
types of palynodebris, the left-hand dendrogram (Q-
mode) is a grouping of the samples, and the matrix is
displayed between them. Hole 1085A assemblages
are used as standards for the other two holes (Figs. 7
and 8). Of the three palynofacies assemblages in
Hole 1085A (Figs. 6 and 9), assemblage C is
represented by 17 samples that are distributed
throughout the hole. Assemblage B is represented
by 11 samples that range from latest Middle Miocene
to Early Pliocene. Assemblage A is represented by
six Middle and Upper Miocene samples. Only two
assemblages (B and C) were identified in Hole
1087C (Fig. 10) while assemblage C was identified
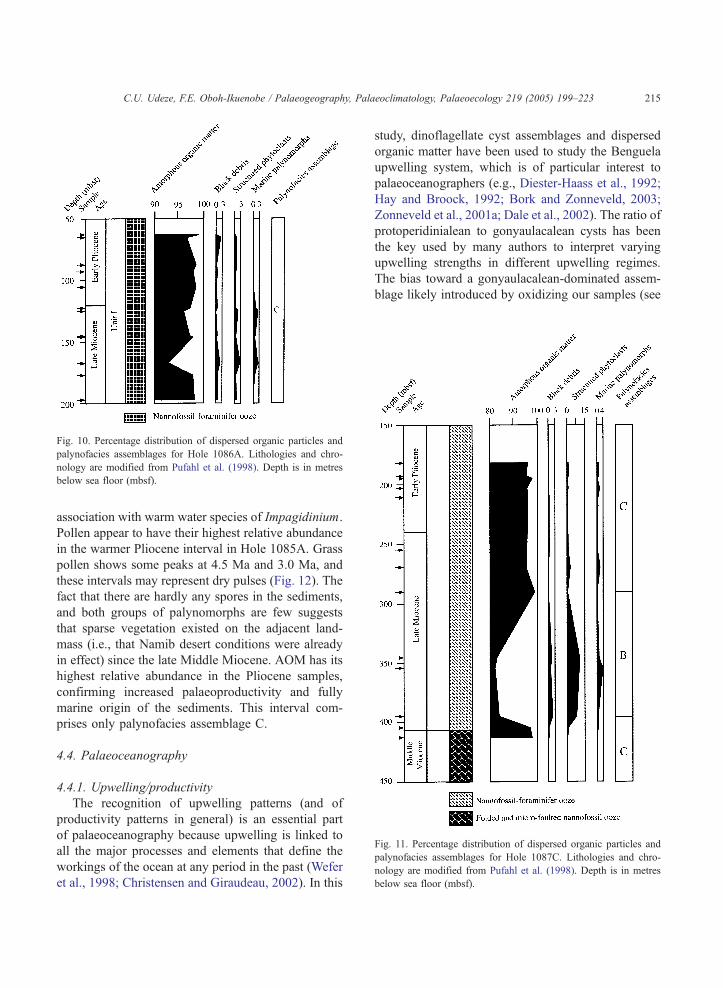
in Hole 1086A (Fig. 11).
Fig. 3. Photomicrographs of palynomorphs, each identified by sample num
(catalog number 7100); unless noted, specimens are from stained and siev
sp. cf. O. israelianum, dorsal view showing operculum, lower focus, 108
dorsal view, lower focus (B), upper focus (C), 1085A-50X-1, 52 cm, 28S/T
lower focus (D), upper focus (E), 1085A-31H-4, 52 cm, 25J/K; uncer
Cymatiosphaera sp., uncertain view, lower focus (G), upper focus (H), 108
section, upper focus, 1087C-39X-5, 0–2 cm, 3P/Q. (J) Spiniferites ramo
Melitasphaeridium choanophorum, dorsal view, upper focus, 1086A-14H-2
view, lower focus, 1086A-16-5, 110–112 cm, 14N/P. (M) Tuberculodinium
20G/A.
4. Discussion
The distribution patterns of dinoflagellate cysts,
acritarchs, spores, pollen and dispersed organic matter
in Hole 1085A, which is the most continuous of the
three holes studied, form the bases for our interpre-
tations. Hole 1086A is the most condensed and does
not show most of the distribution patterns seen in
Holes 1085A and 1087C. Therefore, it is excluded
from the interpretation charts used for this discussion.
4.1. Biostratigraphy
Most of the dinoflagellate cysts and acritarchs
identified are blong-rangingQ and do not show much
biostratigraphic significance in the timeframe of the
study. For biostratigraphic interpretation, only a few
significant species in Holes 1085A and 1087C are
discussed; even fewer dinoflagellate cysts of known
biostratigraphic importance were identified in Hole
1086A. The biostratigraphic framework (composed of
calcareous nannofossils and foraminifers) established
by the Wefer et al. (1998) for Sites 1085, 1086 and
1087 and the age models generated by Westerhold et
al. (in press) have been used as a guide. Of the very
few spores and pollen present in the sediments, only
Echitricolporites spinosus is age diagnostic, with a
base range in the Middle Miocene (Germeraad et al.,
1968); other sporomorphs have not been used for
biostratigraphic interpretation.
Labyrinthodinium truncatum, a latest Early Mio-
cene to latest Late Miocene marker (De Verteuil and
Norris, 1996; Hardenbol et al., 1998), was identified
in samples from the Middle and Late Miocene interval
(Figs. 12 and 13), although its abundance is low.
Operculodinium janduchenei and Impagidinium pat-
ulum with range bases in the Middle Miocene occur in
ber and location coordinates on the Lovins micro-slide field finder
ed slides (#2) unless noted; scale bar is 30 Am. (A) Operculodinium
7C-44X-6, 75–77 cm, 18Q/R. (B, C) Operculodinium janduchenei,
. (D, E, I) Operculodinium centrocarpum, oblique right lateral view,
tain view, upper focus (I), 1085A-21H-5, 80cm, 18M/N. (G, H)
6A-10H-4, 60–62 cm, 23J/K. (H) Operculodinium piaseckii, optical
sus, ventral view, upper focus, 1085A-41X-1, 50 cm, 25E/F. (K)
, 60–62 cm, 16Q/R. (L) Lingulodinium machaerophorum, uncertain
vancampoae, uncertain view, upper focus, 1086A-12H-4, 60–62 cm,
Page 8
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223206
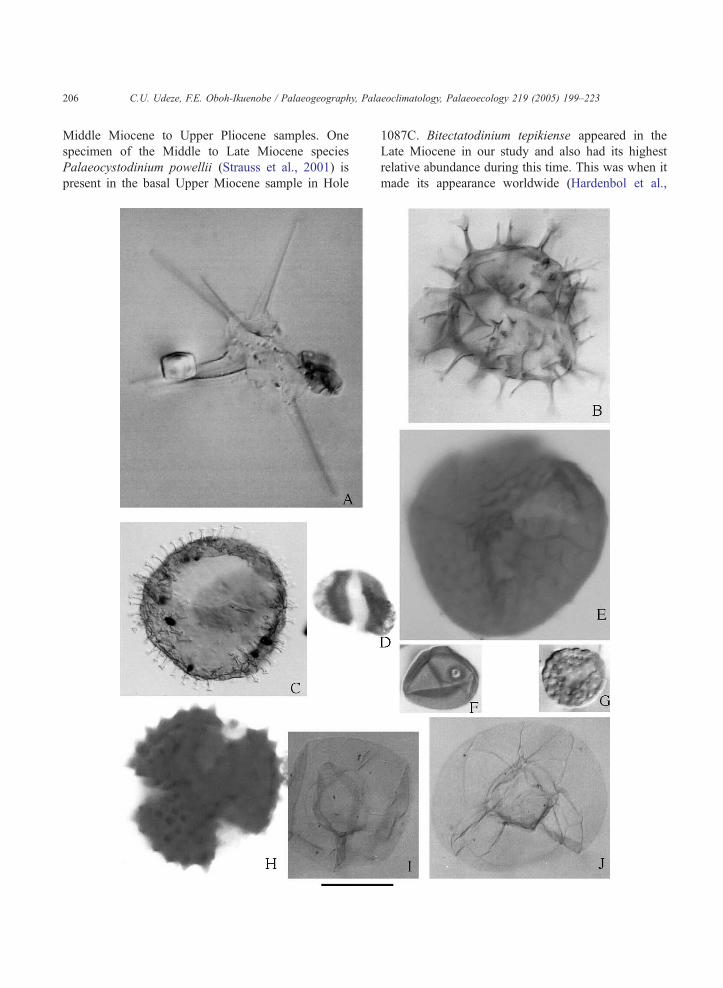
Middle Miocene to Upper Pliocene samples. One
specimen of the Middle to Late Miocene species
Palaeocystodinium powellii (Strauss et al., 2001) is
present in the basal Upper Miocene sample in Hole
1087C. Bitectatodinium tepikiense appeared in the
Late Miocene in our study and also had its highest
relative abundance during this time. This was when it
made its appearance worldwide (Hardenbol et al.,
Page 9
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 207
1998). Therefore, these taxa confirm a Miocene age
for samples from this interval. The acritarch Leios-
phaeridia sp. cf. L. rockhallensis (Fig. 2C) was
present mainly in the least productive samples of the
Middle and Late Miocene interval (Figs. 12 and 13),
although it should be noted that Head and Norris
(2003) recorded the range base of L. rockhallensis as
Early Pliocene. Impagidinium sp. A of Wrenn and
Kokinos (1986) has its highest abundance in the upper
Middle to Upper Miocene samples and is rare or
absent in Pliocene samples from Holes 1085A and
1086A (Fig. 13). Wrenn and Kokinos (1986) recorded
this species in the Middle Miocene interval of the Gulf
of Mexico. Also present in several Upper Miocene to
Upper Pliocene samples are a few taxa that range in
age in from Late Oligocene (Chattian) to Late
Pliocene (Piacenzian–Gelasian) (M.A. Pearce in
Brinkhuis et al., 2004). These taxa include Edward-
siella sexispinosa (Versteegh and Zevenboom, 1995)
and Invertocysta tabulata (Edwards, 1984).
Ataxiodinium confusum and Unidentified sp. A
(possibly copepod eggs) did not appear below the
Pliocene in the three Holes except for two specimens
of Unidentified sp. A in sample 33H-3, 2 cm; Hole
1085A (301.22 mbsf, 5.459 Ma), which is close to the
Miocene–Pliocene boundary. Thus, they may be
Pliocene markers. In the biostratigraphic charts of
Hardenbol et al. (1998), the first appearance datum of
A. confusum was recorded as 5.10 Ma (Early
Pliocene; Zanclean), and extends to Late Placenzian
(2.65 Ma) according to Louwye et al. (2004). This
species, therefore, confirms a Pliocene age for the
samples where it was identified. Furthermore, the
presence of Barssidinium pliocenicum (Late Miocene
to Late Pliocene; De Schepper et al., 2004) in sample
26H-7, 80–82 cm (240.1 mbsf, 4.6 Ma) from Hole
1085A and 22H-3, 75–77 cm (4.6 Ma) from Hole
1087C supports a Pliocene age for these samples.
Fig. 4. Photomicrographs of palynomorphs, each identified by sample num
(catalog number 7100); unless noted, specimens are from stained and sieved
J, and 10 Am for figures E and H. (A) Edwardsiella sexispinosa, un
Achomosphaera sp., uncertain orientation, mid-focus, 1086A-22H-1, 110–
detached operculum, upper focus, 1087C-22H-3, 75–77 cm, 30Q/R. (D) Bi
distal view, mid-focus, 1085A-47X-3, 52 cm, 13L/M. (F) Graminidites s
Chenopodipollis sp., 1087C-44X-6, 0–2 cm, 26Q/R. (H)Echitricolporities sp
dorsal view, mid-focus, 1085A-31H-4, 52 cm, 15P/Q. (J) Invertocysta tabu
4.2. Palynofacies assemblages
The dispersed organic matter content of sediments
from neritic and marginal marine environments
consists of two main components: organic matter
derived from the continent (wood, cuticles, spores,
pollen, fungal remains, etc.) and organic matter
produced in the ocean such as dinoflagellate cysts
and marine amorphous organic matter (Jaramillo and
Oboh-Ikuenobe, 1999). Since the terrestrially derived
organic matter particles behave as clasts in water, their
relative proportions will decrease as the distance from
the source increases (Oboh, 1992; Oboh-Ikuenobe et
al., 1999). In this study, the Orange River appears to
have a strong influence on terrestrial input into the
marine environment.
Palynofacies assemblage A is represented by six
samples in Hole 1085A. It is characterized by an
unusually high input of terrestrially derived phyto-
clasts and near-absence of dinoflagellate cysts,
spores and pollen. The high percentages of
terrestrially derived organic matter particles in this
assemblage appear to be related to high continental
runoff. From offshore Peru, Powell et al. (1990)
reported that high levels of AOM, coupled with the
presence of rich dinoflagellate cyst assemblages
and the absence of accompanying terrestrial paly-
noclasts, indicated an almost exclusive marine
origin. This is similar to that observed in the
samples representing palynofacies assemblages B
and C. Assemblage B samples are characterized by
58–84% of AOM, while assemblage C samples
have more than 90% AOM and a diverse
assemblage of dinoflagellate cysts, including oce-
anic taxa. Deposition of high levels of AOM has
also been attributed to increased productivity in
surface waters (Powell et al., 1990, 1992; Oboh-
Ikuenobe, 2001).
ber and location coordinates on the Lovins micro-slide field finder
slides (#2) unless noted; scale bar is 30 Am for figures A–D, F, G, I and
certain view, upper focus, 1086A-14H-2, 60–62 cm, 29P/Q. (B)
112 cm, 37Q/R. (C) Barrsidinium pliocenicum, dorsal view showing
saccate pollen, 1085A-15H-6, 26–28 cm, 22Q/R. (E) Retitriletes sp.,
p., proximal view, upper focus, 1085A-45X-6, 52 cm, 22K/L. (G)
inosus, 1085A, 15H-6, 26–28cm, 24K/L. (I) Invertocysta lacrymosa,
lata, dorsal view, mid-focus, 1085A-50X-1, 52 cm, 13S/R.
Page 10
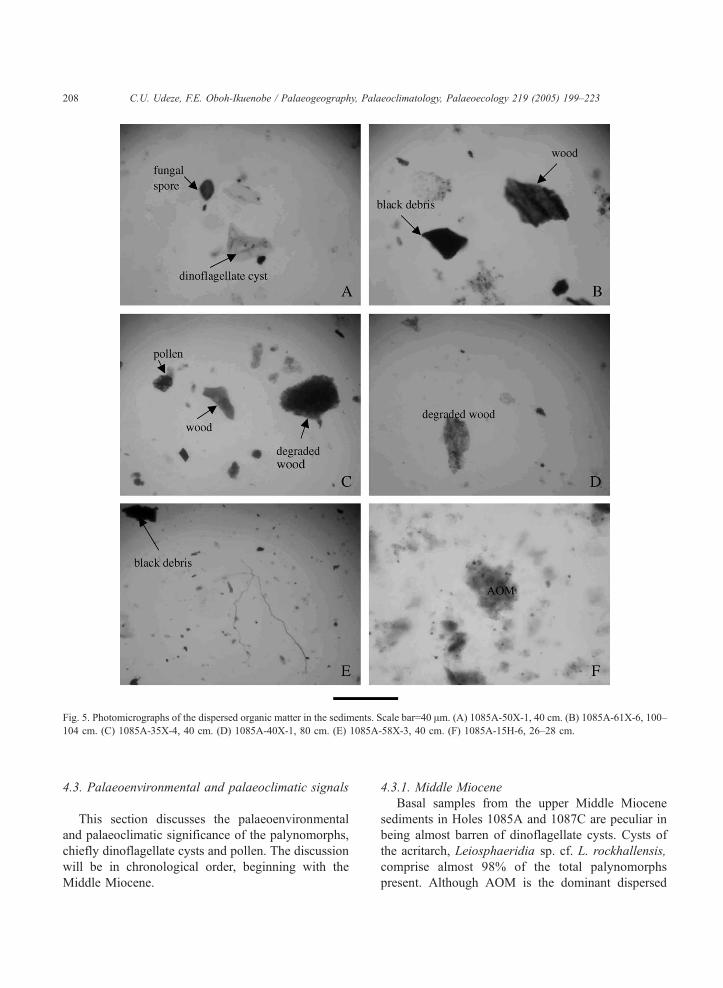
Fig. 5. Photomicrographs of the dispersed organic matter in the sediments. Scale bar=40 Am. (A) 1085A-50X-1, 40 cm. (B) 1085A-61X-6, 100–
104 cm. (C) 1085A-35X-4, 40 cm. (D) 1085A-40X-1, 80 cm. (E) 1085A-58X-3, 40 cm. (F) 1085A-15H-6, 26–28 cm.
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223208
4.3. Palaeoenvironmental and palaeoclimatic signals
This section discusses the palaeoenvironmental
and palaeoclimatic significance of the palynomorphs,
chiefly dinoflagellate cysts and pollen. The discussion
will be in chronological order, beginning with the
Middle Miocene.
4.3.1. Middle Miocene
Basal samples from the upper Middle Miocene
sediments in Holes 1085A and 1087C are peculiar in
being almost barren of dinoflagellate cysts. Cysts of
the acritarch, Leiosphaeridia sp. cf. L. rockhallensis,
comprise almost 98% of the total palynomorphs
present. Although AOM is the dominant dispersed
Page 11
Table 1
Relative abundances (in percent) of dispersed organic matter
Hole Sample number Depth Age AOM BD ST DP FR SP MP PFA
1085A 15H-6, 26–28 135.0 3.02 93.7 0.7 4.0 0.3 0.7 0.3 0.3 C
18H-4, 29–31 160.5 3.40 91.7 0.7 6.0 0.3 0.7 0.3 0.3 C
21H-5, 80 191.0 3.86 94.3 2.0 3.0 0.3 0.0 0.0 0.3 C
23H-1, 30 203.5 4.09 93.7 0.3 1.0 0.7 1.0 0.0 3.3 C
26H-1, 30–32 232.0 4.57 84.0 6.0 6.3 3.0 0.0 0.0 0.7 B
26H-7, 80–82 240.1 4.65 84.0 6.7 3.3 2.7 1.0 0.0 2.3 B
28H-4, 52 255.7 4.81 92.0 1.0 4.0 0.7 0.0 0.0 2.3 C
29H-6, 102 268.7 4.99 90.3 1.0 1.3 0.0 0.7 0.3 6.3 C
31H-4, 52 284.2 5.25 83.0 3.0 6.7 1.3 0.3 0.7 5.0 B
33H-3, 2 301.2 5.46 90.7 1.7 3.0 0.3 0.7 0.0 3.7 C
35X-4, 142 319.7 5.73 53.0 11.7 20.3 2.0 0.0 0.0 13.0 A
36X-6, 82 332.7 5.96 45.0 20.7 30.0 0.0 0.7 0.0 3.7 A
37X-5, 21 340.2 6.13 94.0 2.3 2.0 0.7 0.0 0.0 1.0 C
39X-2, 102 355.9 6.51 18.3 29.0 48.7 0.0 0.3 0.3 3.3 A
40X-1, 102 364.0 6.68 78.3 3.3 12.3 1.7 0.0 0.0 4.3 B
41X-1, 50 373.2 6.87 96.3 1.0 0.3 0.0 0.0 0.3 2.0 C
42X-6, 102 390.9 7.44 94.0 2.0 1.7 0.0 0.0 0.0 2.3 C
45X-4, 52 416.5 8.58 97.3 1.0 0.0 0.3 1.0 0.0 0.3 C
45X-6, 52 319.5 8.71 45.3 16.7 26.7 2.3 0.0 0.3 8.7 A
46X-5, 2 427.1 8.93 63.0 6.3 13.7 0.0 13.7 0.3 3.0 B
47X-3, 52 434.2 9.11 96.3 1.3 0.0 2.3 0.0 0.0 0.0 C
47X-6, 102 439.2 9.24 97.0 2.0 1.0 0.0 0.0 0.0 0.0 C
49X-2, 52 452.0 9.59 95.7 1.7 1.7 0.0 0.3 0.0 0.7 C
50X-1, 52 460.2 9.78 55.0 4.0 11.7 1.0 0.0 0.0 28.3 A
52X-3, 2 481.5 10.18 65.3 6.7 25.3 0.3 1.3 0.0 1.0 B
53X-4, 52 493.6 10.40 95.3 1.7 1.0 2.0 0.0 0.0 0.0 C
55X-3, 52 511.1 10.73 95.0 2.3 2.0 0.0 0.3 0.0 0.3 C
56X-3, 2 520.2 10.92 80.3 1.0 14.7 0.7 0.7 0.0 2.7 B
58X-3, 52 540.1 11.32 68.7 10.7 18.7 0.7 0.3 0.0 1.0 B
59X-3, 100–102 550.1 11.53 93.0 2.0 4.3 0.0 0.0 0.3 0.3 C
60X-6, 103 564.3 12.02 79.3 10.7 9.3 0.0 0.7 0.0 0.0 B
61X-6, 102 574.1 12.62 73.3 16.0 10.3 0.0 0.3 0.0 0.0 B
63X-1, 2 585.7 13.34 58.7 20.0 20.0 0.0 0.7 0.0 0.7 B
63X-4, 102 590.2 13.62 28.3 39.0 28.3 0.3 0.3 0.0 3.7 A
1086A 8H-1, 10–12 63.8 3.99 99.7 0.0 0.0 0.3 0.0 0.0 0.0 C
8H-2, 60–62 65.8 4.08 96.7 2.3 1.0 0.0 0.0 0.0 0.0 C
10H-4, 60–62 87.7 4.58 98.7 0.3 1.0 0.0 0.0 0.0 0.0 C
11H-1, 110–112 93.3 4.66 98.0 0.7 0.7 0.3 0.0 0.0 0.3 C
12H-4, 60–62 106.8 4.96 98.7 0.3 0.7 0.3 0.0 0.0 0.0 C
14H-2 60–62 122.8 5.39 97.3 0.7 0.7 0.0 0.0 0.0 1.3 C
14H-4, 60–62 125.8 5.46 96.0 0.0 1.7 0.0 0.0 0.0 2.3 C
16H-5, 10–12 145.8 5.93 97.3 0.7 0.7 0.3 0.0 0.0 1.0 C
16H-5, 110–112 146.8 5.96 98.0 0.3 0.7 0.3 0.0 0.0 0.7 C
18H-7, 10–12 167.8 6.52 93.0 2.0 2.7 0.0 0.0 0.0 2.3 C
20H-1, 10–12 177.8 6.85 98.0 0.7 0.7 0.0 0.0 0.0 0.7 C
22H-1, 110–112 197.8 7.38 98.3 0.7 1.0 0.0 0.0 0.0 0.0 C
1087C 21H-1, 4–6 182.1 4.07 96.0 0.7 2.3 0.0 0.0 0.0 1.0 C
22H-2, 0–2 193.1 4.56 95.3 1.0 2.7 0.0 0.0 0.0 1.0 C
22H-3, 75–77 195.4 4.65 98.3 0.3 1.0 0.0 0.0 0.0 0.3 C
23H-2, 75–77 203.4 4.96 95.7 0.3 2.3 0.0 0.0 0.0 1.7 C
24H-4, 75–77 215.9 5.45 96.3 1.7 1.3 0.0 0.0 0.0 0.7 C
29X-1, 0–2 254.9 5.96 95.7 1.7 2.3 0.0 0.0 0.0 0.3 C
(continued on next page)
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 209
Page 12
Table 1 (continued)
Hole Sample number Depth Age AOM BD ST DP FR SP MP PFA
1087C 30X-6, 75–77 269.5 6.52 95.3 0.7 3.0 0.0 0.0 0.0 1.0 C
33X-1, 75–77 290.8 6.86 99.0 0.3 0.7 0.0 0.0 0.0 0.0 C
38X-6, 75–77 346.5 8.95 83.0 5.0 10.7 0.0 0.0 0.0 1.3 B
39X-5, 0–2 353.8 9.14 82.3 3.7 10.3 0.0 0.0 0.0 3.7 B
43X-6, 75–77 394.6 10.16 84.3 6.7 8.3 0.0 0.0 0.0 0.7 B
44X-6, 75–77 404.3 10.62 97.0 1.0 1.3 0.0 0.0 0.0 0.7 C
46X-6, 0–2 422.7 11.50 98.0 1.3 0.7 0.0 0.0 0.0 0.0 C
Sample intervals are in cm, depth is in mbsf, and age is in Ma. Abbreviations are as follows: AOM=amorphous organic matter; BD=black
debris; ST=structured phytoclasts; DP=degraded phytoclasts; FR=fungal remains; SP=sporomorphs; MP=marine palynomorphs; PFA=paly-
nofacies assemblage.
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223210
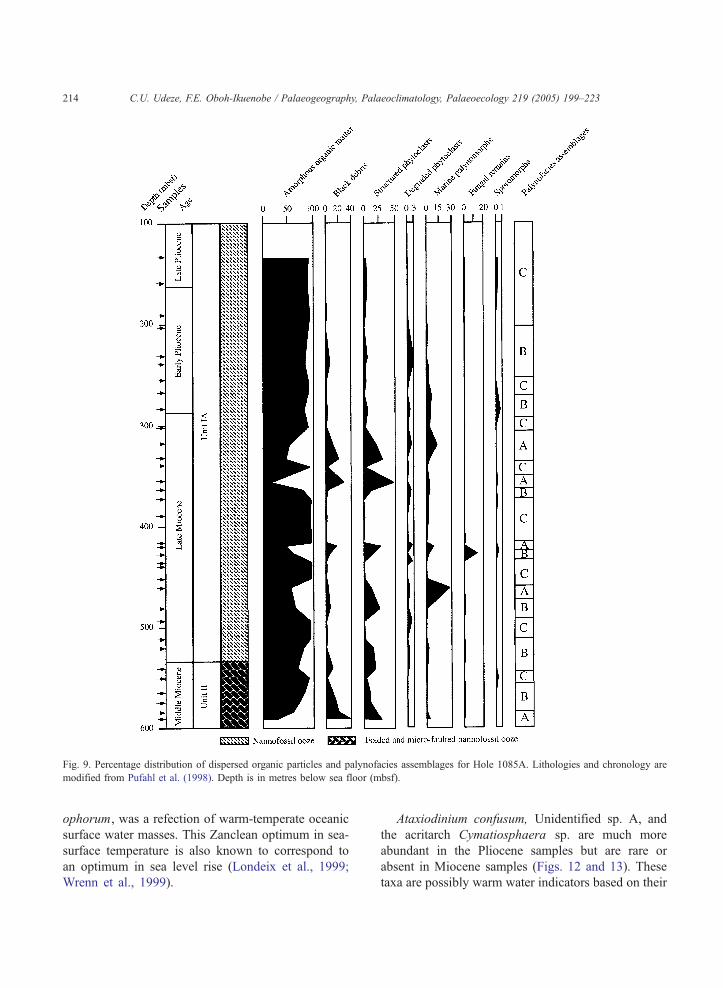
organic matter, the percentages of land derived
organic matter (black debris and structured phyto-
clasts) are higher in samples from this interval than
in the Upper Miocene interval (Fig. 9). Spores and
pollen were absent, which may this suggest that
there was sparse vegetation on the adjacent land-
mass. In a preliminary low-resolution palynological
study of 19 samples from Hole 1085A, Oboh-
Ikuenobe (2001) interpreted this interval (13.6–12
Ma) as the warmest interval before the major
cooling event of the Late Miocene. In this study,
however, the basal upper Middle Miocene samples
do not have enough palynological evidence for
palaeoclimatic and palaeoceanographic interpreta-
tion, but the high terrestrial influence could
represent high continental run off enhanced by
higher temperatures.
In the upper Middle Miocene sample 59X-3, 100–
102 cm (550. 14 mbsf, 11.53 Ma) from Hole 1085A,
there was a sudden appearance of dinoflagellate cysts
dominated by Impagidinium sp. A of Wrenn and
Kokinos (1986) (Fig. 12). This sample is also marked
by a sudden appearance of spores and pollen
including Chenopodipollis sp. (Chenopodiaceae),
Echitricolporites spinosus (Compositae) and Grami-
nidites sp. (grass pollen). Grass pollen had one of its
peaks in relative abundance in this sample. This
probably represents a dry pulse at that interval (Fig.
12). In the lowermost sample from Hole 1087C,
bisaccate pollen was the only pollen present. Since
Site 1085 is closest to the Orange River, it probably
experienced more terrestrial input than Sites 1086 and
1087. It should be noted that Pagani et al. (1999)
interpreted an increase in d18O values to document the
onset of cooling in the ocean at 11.5 Ma. It appears,
therefore, that this interval may mark the beginning of
the major Late Miocene cooling identified by Oboh-
Ikuenobe (2001).
4.3.2. Late Miocene
The dominant species of Impagidinium in the
Middle to Upper Miocene samples from all three
holes are Impagidinium paradoxum and Impagidi-
nium sp. A of Wrenn and Kokinos (1986). I. para-
doxum extends into cooler regions of the oceans than
Impagidinium aculeatum and Impagidinium patulum
(e.g., Edwards, 1992; Marret and Zonneveld, 2003).
Wall et al. (1977) and Marret and Zonneveld (2003)
reported this species as oceanic, temperate to tropical.
The trend shown by Impagidinium sp. A of Wrenn
and Kokinos (1986) is similar to that shown by I.
paradoxum, suggesting that it is also an oceanic,
temperate to tropical species (Figs. 12 and 13).
Species of Impagidinium that are associated with
warm (tropical to warm-temperate) oceanic environ-
ments, including I. aculeatum and I. patulum (Wall et
al., 1977; Head, 1996; Marret and Zonneveld, 2003),
are less abundant in this interval. Bitectatodinium
tepikiense is a neritic species characteristic of cold/
temperate regions (Dale, 1996; Rochon et al., 1999;
Marret and Zonneveld, 2003) and is more abundant in
this interval. Thus, it confirms the major late Middle
to Late Miocene cooling (Pagani et al., 1999).
The cooling event identified in the Late Miocene
interval does not necessarily indicate a very cold
environment because of the presence of warmer water
indicators, such as Lingulodinium machaerophorum,
Barrsidinium pliocenicum and Melitasphaeridium
choanophorum (Head, 1996). It suggests that cooling
was a relative event compared to the Middle Miocene.
The high relative abundance of L. machaerophorum
(13%) in the Upper Miocene sample 16H-5, 10–12
Page 13
15H-6
21H-518H-4
59X-3
49X-655X-341X-145X-4
47X-6
47X-353X-423H-129H-628H-433H-437X-542X-6
63X-439X-2
26H-126H-731H-440X-156X-346X-552X-358X-363X-160X-661X-635X-445X-650X-136X-6
C
B
A
120 100 80 60 40 20 0
Minimum variance
55-90%30-55%10-30%<10%
>90%
absent+
+
++
+
+++
+
+
+
++
+
++
+++
+
+++
+
++++
+++
++
+++
+
+
+
++
++
++
+
++
+
++
+
++
++
+
++
Paly
nofa
cies
ass
embl
ages
AO
M
BD
ST MP
DP
SPFR
Squared Euclidean-Data log(2) transformed
Fig. 6. Dendrograms for average linkage cluster analysis of palynological samples from Hole 1085A (Q-mode, left-hand) and of dispersed
organic matter (R-mode, top), with the data matrix displayed between them.
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 211
cm (5.933 Ma) from Hole 1086A (Udeze, 2003) may
also be controlled by elevated nutrient input (Rochon
et al., 1999).
Nematosphaeropsis labyrinthus and Nematos-
phaeropsis lemniscata (which may be synonymous
according to Rochon et al., 1999) show similarities in
their relative abundance distribution in Holes 1085A
and 1087C (Figs. 12 and 13). Nematosphaeropsis
labyrinthus is a cosmopolitan species that is able to
tolerate a wide variety of environments (Marret and
Zonneveld, 2003). It has been shown to increase with
water depth (Wall et al., 1977; Harland, 1983; Turon,
1984) and has, therefore, been regarded as a character-
istic species for outer neritic to oceanic environments.
According to Wall et al. (1977), this species domi-
nates tropical to subtropical open ocean environments,
while Head (1998) reported its presence to indicate
oceanic influence. N. labyrinthus and N. lemniscata
show no particular trend in this study but their high
relative abundance in the samples, together with
Page 14
Table 2
Principal component loadings for statistically significant types of
phytoclasts and palynomorphs
Organic matter component Axis 1 Axis 2
AOM �0.225* 0.001
Black debris 0.589* �0.349*
Structured phytoclasts 0.013 0.227*
Degraded phytoclasts 0.719* �0.073
Marine palynomorphs 0.287* 0.903*
The asterisk indicates absolute values greater than 0.10; eigenvalue
(Axis) 1=4.319 (61.86% of total variance); Axis 2=1.296 (18.56%).
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223212
several species of Impagidinium, clearly indicates an
oceanic environment. This observation confirms the
conclusions of Oboh-Ikuenobe (2001).
A
OM
8H-1
8H-2
22H-1
12H-4
11H-1
16H-5
16H-5
14H-2
20H-1
14H-4
18H-7
0.0
Squared Euclidean-Data log(2) transformed
10H-455-90%30-55%10-30%<10%
>90%
absent+
Minimum variance
0.81.62.43.24.04.8
Fig. 7. Dendrograms for average linkage cluster analysis of palynologica
organic matter (R-mode, top), with the data matrix displayed between the
The pollen identified in this interval include,
grass pollen, Chenopodipollis sp. and Echitricolpor-
ites spinosus. They do not show any particular
trend but they appear to be more abundant in the
Upper Miocene samples than in the Middle
Miocene samples. The dispersed organic matter
components are dominantly AOM; black debris
and structured phytoclasts are few. There is also
an increase in the relative abundance of marine
palynomorphs.
4.3.3. Early to Late Pliocene
Impagidinium aculeatum and Impagidinium pat-
ulum are considered warm (tropical to warm-
temperate) oceanic species (Wall et al., 1977; Head,
BD
ST SPFRMP
DP
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
C
Paly
nofa
cies
ass
embl
age
l samples from Hole 1086A (Q-mode, left-hand) and of dispersed
m.
Page 15
21H-1
22H-2
30X-6
23H-2
24H-4
44X-6
29X-1
22H-3
33X-1
46X-6
38X-6
43X-6
39X-5
04812162024
Squared Euclidean-Data log(2) transformed
Minimum variance AO
M
BD
ST SPFRMP
+
+ +
+
+
+
+
+
+
+
+
+
+
+
DP
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
+
55-90%30-55%10-30%<10%
>90%
absent+
B
C
Paly
nofa
cies
ass
embl
ages
Fig. 8. Dendrograms for average linkage cluster analysis of palynological samples from Hole 1087C (Q-mode, left-hand) and of dispersed
organic matter (R-mode, top), with the data matrix displayed between them.
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 213
1996; Marret and Zonneveld, 2003) and they have
their highest relative abundance in our Lower to
lower Upper Pliocene samples (Figs. 12 and 13).
The more cool-tolerant species of Impagidinium
(Impagidinium paradoxum and Impagidinium sp.
A.), Filisphaera filifera and Bitectatodinium tepi-
kiense are rare or absent in this interval. In
Operculodinium centrocarpum is more abundant in
the lower Upper Pliocene samples and is absent or
rare in the Miocene samples. A study in the
Benguela Upwelling region by Davey and Rogers
(1975) indicated that O. centrocarpum was domi-
nant in relatively warm waters. A possible warming
event can, therefore, be suggested for the Early to
early Late Pliocene interval.
4.3.4. Summary
The ecostratigraphy of the study area, based on
the dinoflagellate cyst assemblages, can be inter-
preted as a subtropical late Middle Miocene interval,
a cool-temperate Late Miocene interval and a warm-
temperate Early to early Late Pliocene interval. The
Early Pliocene (Zanclean) warming has been recog-
nized as a global event by many authors, who used
different proxies, including dinoflagellate cysts, oxy-
gen isotope, spores and pollen (Zagwijn, 1960; Suc
et al., 1995; Wrenn et al., 1999). Londeix et al.
(1999) reported that the dinoflagellate cyst assem-
blage from the Late Neogene (Messinian–Zanclean)
of the Strait of Sicily, which was dominated by
Impagidinium patulum and Lingulodinium machaer-
Page 16
Fig. 9. Percentage distribution of dispersed organic particles and palynofacies assemblages for Hole 1085A. Lithologies and chronology are
modified from Pufahl et al. (1998). Depth is in metres below sea floor (mbsf).
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223214
ophorum, was a refection of warm-temperate oceanic
surface water masses. This Zanclean optimum in sea-
surface temperature is also known to correspond to
an optimum in sea level rise (Londeix et al., 1999;
Wrenn et al., 1999).
Ataxiodinium confusum, Unidentified sp. A, and
the acritarch Cymatiosphaera sp. are much more
abundant in the Pliocene samples but are rare or
absent in Miocene samples (Figs. 12 and 13). These
taxa are possibly warm water indicators based on their
Page 17
Fig. 10. Percentage distribution of dispersed organic particles and
palynofacies assemblages for Hole 1086A. Lithologies and chro-
nology are modified from Pufahl et al. (1998). Depth is in metres
below sea floor (mbsf).
Fig. 11. Percentage distribution of dispersed organic particles and
palynofacies assemblages for Hole 1087C. Lithologies and chro
nology are modified from Pufahl et al. (1998). Depth is in metres
below sea floor (mbsf).
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 215
association with warm water species of Impagidinium.
Pollen appear to have their highest relative abundance
in the warmer Pliocene interval in Hole 1085A. Grass
pollen shows some peaks at 4.5 Ma and 3.0 Ma, and
these intervals may represent dry pulses (Fig. 12). The
fact that there are hardly any spores in the sediments,
and both groups of palynomorphs are few suggests
that sparse vegetation existed on the adjacent land-
mass (i.e., that Namib desert conditions were already
in effect) since the late Middle Miocene. AOM has its
highest relative abundance in the Pliocene samples,
confirming increased palaeoproductivity and fully
marine origin of the sediments. This interval com-
prises only palynofacies assemblage C.
4.4. Palaeoceanography
4.4.1. Upwelling/productivity
The recognition of upwelling patterns (and of
productivity patterns in general) is an essential part
of palaeoceanography because upwelling is linked to
all the major processes and elements that define the
workings of the ocean at any period in the past (Wefer
et al., 1998; Christensen and Giraudeau, 2002). In this
study, dinoflagellate cyst assemblages and dispersed
organic matter have been used to study the Benguela
upwelling system, which is of particular interest to
palaeoceanographers (e.g., Diester-Haass et al., 1992;
Hay and Broock, 1992; Bork and Zonneveld, 2003;
Zonneveld et al., 2001a; Dale et al., 2002). The ratio of
protoperidinialean to gonyaulacalean cysts has been
the key used by many authors to interpret varying
upwelling strengths in different upwelling regimes.
The bias toward a gonyaulacalean-dominated assem-
blage likely introduced by oxidizing our samples (see
-
Page 18
400
200
300
500
600
100
AgeSample
s
Mid
dle
Mio
cene
Lat
e M
ioce
neE
arly
Plio
cene
Lat
e Pl
ioce
ne
Depth
(mbs
f)
200
Impa
gidini
um ac
uleatu
m
an
d I. p
atulum
700 35
Impa
gidini
um sp
. A of
Wren
n and
Kok
inos (
1986
) and
I. pa
rado
xum
2 40
Ataxiod
inium
confu
sum
35 700
Cymati
osph
aera
sp.
0 20
Operc
ulodin
ium ce
ntroc
arpu
m
60300
Spini
ferite
s ram
osus
30150
Uniden
tified
sp. A
200
Nemato
spha
erop
sis sp
p.
Subt
ropi
cal
clim
ate
War
m-t
empe
rate
clim
ate
Coo
ling
even
t and
tem
pera
te c
limat
e
Dry pulse
Dry pulse
Dry pulsePala
eocli
matic
in
terpr
etatio
ns
Carbonate crash?
Carbonate crash
Incr
ease
d pr
oduc
tivity
and
poss
ible
ons
et o
f up
wel
ling
Incr
ease
d pr
oduc
tivity
Palaeo
cean
ograp
hic
in
terpr
etatio
ns
70
Cheno
podip
ollis
sp.
40200
Echitr
icolpo
rites
spino
sus
100
Gramini
dites
sp.
Dinoflagellate cysts and acritarchs Pollen
C
B
C
B
C
A
CAB
C
B
C
B
C
B
C
B
A
A
A
Palyno
facies
asse
mblage
s
40200
Bitecta
todini
um te
pikien
se
R
R
T
T
Seque
nce
ch
rono
strati
grap
hy
SequencesT-RFaciesCycles
0 100
50
Leiosp
haer
idia s
p. cf.
L. roc
khall
ensis
Ser 4/Tor 1
Ser 3
Ser 2
Tor 2
Tor 3/Me 1
Me 2
Za 1
Za 2
Pla 1
Fig. 12. Palaeoclimatic and palaeoceanographic interpretations of Hole 1085A based on significant palynomorph data and palynofacies assemblages. Sequence chronostratigraphy is
based on Hardenbol et al. (1998); T=transgression; R=regression. Depth is in metres below sea floor (mbsf).
C.U.Udeze,
F.E.Oboh-Iku
enobe/Palaeogeography,Palaeoclim
atology,Palaeoeco
logy219(2005)199–223
216
Page 19
200
Impa
gidini
um ac
uleatu
m
and I
. patu
lum
40200
Labyr
intho
dinium
trun
catum
80400
Leiosp
haer
idia s
p. cf.
L. r
ockh
allen
sis
40200
Operc
ulodin
ium ce
ntroc
arpu
m
100
Barssi
dinium
plioc
enicu
m
30
Gramini
dites
sp.
0 22
Bitecta
todini
um te
pikien
se
0 10
Echitr
icolpo
rites
spino
sus
70350Im
pagid
inium
para
doxu
m and
I. sp
A of
Wren
n and
Kok
inos (
1986
)
Dinoflagellate cysts and acritarchs Pollen
150
Nemato
spha
erop
sis le
mnisca
ta
C
B
C
Palyno
facies
asse
mblage
s
War
m-t
empe
rate
clim
ate
Coo
ling
even
t and
tem
pera
te c
limat
e
Palaeo
climati
c inte
rpret
ation
s
Carbonate crash?
Incr
ease
d pr
oduc
tivity
Palaeo
cean
ograp
hic
interp
retati
ons
AgeSample
150
200
250
300
350
400
450
Lat
e M
ioce
ne M
iddl
eM
ioce
ne
Depth
(mbs
f)
Ear
ly P
lioce
ne
T
T
R
SequencesT-RFaciesCycles
Seque
nce
chro
nostr
atigr
aphy
Ser 4/Tor 1
Tor 2
Tor 3/Me 1
Me 2
Za 1
Za 2
Fig. 13. Palaeoclimatic and palaeoceanographic interpretations of Hole 1087C based on significant palynomorph data and palynofacies
assemblages. Sequence chronostratigraphy is based on Hardenbol et al. (1998); T=transgression; R=regression. Depth is in metres below sea
floor (mbsf).
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 217
Section 3.1) precludes us from using this approach for
interpretation. However, by examining the gonyaula-
calean cysts dominating our palynomorph assemb-
lages, especially Batiacasphaera spp., Impagidinium
spp., Nematosphaeropsis spp., Spiniferites ramosus
and Operculodinium spp., we can make inferences
about palaeoceanographic events in the region.
Margalef (1978) described the Mandala model in
which diatoms dominated areas with turbulent con-
ditions and good nutrient supply, while dinoflagellates
dominated non-turbulent conditions with poor nutrient
supply. Protoperidinialeans are heterotrophic and
typically feed on diatoms and other phytoplankton,
they will favour areas of strong upwelling, unlike
autotrophic gonyaulacaleans. Margalef’s model fur-
ther suggests that red tide-forming dinoflagellates or
harmful algal blooms (HAB; notably some gonyaula-
calean species) dominate in regions of high nutrients
but low turbulence. However, Smayda and Reynolds
(2001) re-examined Margalef’s model and used it
with Reynolds’ C-S-R model (Reynolds, 1987) to
show that for HAB dinoflagellates, distinctive asso-
ciations reflect life-form properties, habitat preference
and stochastic selection of species. All these charac-
teristics will determine the composition of such
communities.
Prolific primary production may occur in surface
waters in areas of oceanic upwelling, resulting in the
accumulation of organic matter both in the water
column and on the sea floor (Batten, 1996). Siesser
Page 20
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223218
(1980) suggested that major, sustained upwelling in
the Benguela upwelling system started in the Early to
Late Miocene based on multiple proxies including
sediment accumulation rates, diatom abundances,
microfossil temperature preferences, and productivity.
Later studies by Diester-Haass et al. (2001, 2002)
using multiple productivity proxies, including benthic
foraminiferal accumulation rate (BFAR) and total
organic carbon (TOC), identified a Late Miocene
(6.5–6.7 Ma) onset of productivity at Site 1085. They
associated this onset of productivity with colder
periods and very low terrigenous input. In this study,
the cooler latest Middle to Late Miocene interval in
Hole 1085A appears to have fewer sporomorphs and
other terrestrial organic material and higher propor-
tions of AOM than the warmer Early Pliocene.
Furthermore, intervals with elevated AOM (palynof-
acies assemblage C) coincide with increases in sea
level (Fig. 12). Since deposition of such massive
amounts of AOM dominates post-Palaeogene upwell-
ing regimes with high productivity (Powell et al.,
1990, 1992), increase in AOM may be an indicator of
increased palaeoproductivity. The productivity pattern
at Site 1085 is similar to the patterns identified in the
Pacific and Indian Oceans, and suggests that the
productivity increase off southwest Africa is part of a
global response to palaeoceanographic changes (Die-
ster-Haass et al., 2002). In this region and areas to the
east, studies characterizing upwelling systems and
productivity based on dinoflagellate cyst assemblages
(or particular species) have focused on Late Pliocene
and Quaternary sediments (Dale and Dale, 2002; Dale
et al., 2002; Esper and Zonneveld, 2002; Holzwarth et
al., 2003; Esper et al., 2004).
Melia (1984) showed that a decrease in the level
of foraminiferal test linings could be correlated with
increasing water depth. Cross et al. (1966) related
their relative abundance to coincide with upwelling
of nutrient rich waters. Powell et al. (1992) found
that samples with high abundance of foraminiferal
test linings tend to be from laminated sedimentary
units (deposited under the influence of upwelling
with anoxic bottom conditions). They suggested
that the strength of upwelling (as measured by
relatively low surface water palaeotemperatures)
was the most significant factor controlling the
levels of foraminiferal linings recorded. In this
study, foraminiferal linings were present but rare
(less than 2% of the total organic matter) and did
not show any particular trend in their distribution. It
is suggested that their rarity could be because of
the individuals present not being mature enough to
have thick chambers that could withstand the rigors
of microbial attack and sample preparation (Powell
et al., 1992).
4.5. The bcarbonate crashQ
The term bcarbonate crashQ was first used by
Lyle et al. (1995) to refer to the interval between
11.2 and 8.6 Ma cored during ODP Leg 138 and
other DSDP sites in the eastern equatorial Pacific,
which are characterized by very low carbonate mass
accumulation rates (MARs) and poorer preservation
of calcium carbonate microfossils. Similar occur-
rences of carbonate dissolution at the middle to late
Miocene transition have been recorded in other
parts of the world, including the Caribbean Sea
(Roth et al., 2000) and South Atlantic (Dean and
Gardner, 1985). Changes in deep water circulation,
shoaling of the carbonate compensation depth, and
shallow–deep fractionation of carbonate material are
considered as possible causes. This phenomenon
has also been identified in ODP Sites 1085 and
1087 in the Cape Basin (Diester-Haass et al., 2001,
2002). These episodes of significant drops in
concentration and mass accumulation rates of
CaCO3 are followed by a strong increase in
biogenous sedimentation (Lyle et al., 1995).
Diester-Haass et al. (2002) associated the cooler
intervals in Hole 1085A with having high CaCO3
concentrations except for a cold period at 6.5 Ma,
which showed a depression in CaCO3 concentra-
tion and low productivity. They interpreted the
level at 6.5 Ma as representing the global Middle
to Late Miocene bcarbonate crash.Q In this study,
samples 63X-4, 102 cm (13.624 Ma) and 39X-2,
102 cm (6.514 Ma) in Hole 1085A are nearly
monospecific, comprising the cysts of Leiosphaer-
idia sp. cf. L. rockhallensis. This is probably due
to poor preservation of the dinoflagellate cysts at
these intervals, which are possibly among the
several intervals of CaCO3 depressions during the
Middle to Late Miocene. These two samples (39X-
2, 102 cm and 63X-4, 102 cm) are among the six
samples that represent palynofacies assemblage A
Page 21
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 219
in Hole 1085A (i.e., with fewer amounts of AOM
and more terrestrial organic debris). They indicate
possible depression in CaCO3 and are interpreted
as possibly indicative of the global bcarbonatecrash.Q
1. Dinoflagellate cysts
Division DINOFLAGELLATA (Bqtschli 1885)Fensome et al. 1993
Subdivision DINOKARYOTA Fensome et al. 1993
Class DINOPHYCEAE Pascher 1914
Subclass PERIDINIPHYCIDAE Fensome et al. 1993
Order GONYAULACALES Taylor 1980
Suborder GONYAULACINEAE (autonym)
Family GONYAULACACEAE Lindemann 1928
Subfamily CRIBROPERIDINIOIDEAE
Fensome et al. 1993
Cordosphaeridium minimum (Morgenroth 1966)
Bebedek 1972
Lingulodinium machaerophorum Deflandre
and Cookson 1955
Operculodinium centrocarpum (Deflandre and
Cookson 1955) Wall 1967
Operculodinium eirikianum Head et al. 1989
Operculodinium sp. cf. O. israelianum
(Rossignol 1962) Wall 1967
Operculodinium janduchenei Head et al. 1989
Operculodinium piaseckii Strauss and Lund 1992
Subfamily GONYAULACOIDEAE (autonym)
Achomosphaera sp.
Ataxiodinium confusum Versteegh and
Zevenboom in Versteegh 1995
Bitectatodinium tepikiense Wilson 1973
Corrudinium sp.
Edwardsiella sexispinosum Versteegh and
Zevenboom in Versteegh 1995
Filisphaera filifera Bujak 1984
Impagidinium aculeatum Wall 1967
Impagidinium paradoxum Wall 1967
Impagidinium patulum Wall 1967
Impagidinium sp. A of Wrenn and Kokinos (1986)
Impagidinium velorum Bujak 1984
5. Conclusions
Palynological analyses of Holes 1085A, 1086A
and 1087C have permitted the palaeoclimatic and
palaeoceanographic interpretations of the Middle
Miocene to early Late Pliocene interval of the Cape
Basin. The Middle Miocene interval is interpreted as
the warmest interval before the onset of the major
Miocene cooling due to the absence of cool water
dinoflagellate species. The presence of cool-tolerant
species in the latest Middle to Late Miocene interval,
such as Bitectatodinium tepikiense, suggests a cooler
interval. The Early to early Late Pliocene interval is
dominated by warmer water species (Impagidinium
aculeatum and Impagidinium patulum) and, therefore,
is consistent with a warming event at the interval
correlated with the Zanclean event reported by other
researchers in different parts of the world. Intervals
with increased percentages of grass pollen have been
interpreted as representing dry pulses. Of the three
palynofacies assemblages identified in Hole 1085A,
assemblage C is characterized by the highest percen-
tages of AOM, most abundant and diverse dinofla-
gellate cysts, and the lowest percentages of land
derived organic material. It represents intervals during
the Late Miocene and Pliocene when palaeoproduc-
tivity was very high. Palynofacies assemblage A
represents periods of high terrestrial input. The
Neogene carbonate crash (Lyle et al., 1995) was
recognized in two Miocene samples in Hole 1085A
(590 mbsf, 13.62 Ma and 355 mbsf, 6.5 Ma) that are
nearly barren of dinoflagellate cysts. Diester-Haass et
al. (2002) recognized the interval at 6.5 Ma as
representing the Late Miocene carbonate crash using
multiple proxies for interpretation.
Nematosphaeropsis labyrinthus (Ostenfield 1903) Reid 1974
Nematosphaeropsis lemniscata Bujak 1984
Nematosphaeropsis oblonga Mudie 1987
Spiniferites ramosus (Ehrenberg 1838) Mantell 1854
Subfamily Uncertain
Invertocysta tabulata Edwards 1984
Invertocysta lacrymosa Edwards 1984
Acknowledgements
We thank Dr. Lisolette Haass for encouraging us to
undertake this study and providing us valuable data to
support our results, Drs. L.E. Edwards and M.J. Head
for their help with taxonomic clarification, the Ocean
Drilling Program for samples, and the University of
Missouri Research Board and Josephine Husband
Radcliffe Scholarship for funding this study. Con-
structive reviews by M.J Head and G.J.M. Versteegh
greatly improved the manuscript.
Appendix A. List of palynomorphs
Page 22
Melitasphaeridium choanophorum Deflandre and
Cookson 1955
Suborder Uncertain
Family Uncertain (order Gonyaulacales)
Batiacasphaera hirsuta Stover 1977
Batiacasphaera sphaerica Stover 1977 sensu lato
Dapsilidinium pseudocolligerum Stover 1977
Labyrinthodium truncatum Piasecki 1980
Reticulatosphaera actinocoronata (Benedek 1972) Bujak
and Matsuoka 1986
Suborder GONIODOMINEAE Fensome et al. 1993
Family GONIODOMACEAE (Autonym)
Subfamily HELGOLANDINIODEAE Fensome et al. 1993
Tuberculodinium vancampoae (Rossignol 1962) Wall 1967
Order PERIDINIALES Haeckel 1894
Family PERIDINIACEAE Ehrenberg 1831
Palaeocystodinium powellii Strauss et al. 2001
Family PROTOPERIDINIACEAE Bujak and Davies 1998
Barssidinium pliocenicum Head 1993
Lejeunecysta sp.
Selenopemphix sp.
2. Acritarchs
Cymatiosphaera sp.
Leiosphaeridia sp. cf. L. rockhallensis Head and Norris 2003
Leiosphaeridia sp. A
3. Angiosperm pollen
Bombacacidites sp.
Chenopodipollis sp.
Echitricolporites spinosus (Van der Hammen 1956) Germeraad
et al. 1968
Intratriporopollenities sp.
Graminidites sp.
Quercoidites sp.
Triatripollenites sp.
4. Gymnosperm pollen
Undifferentiated bissacate pollen
5. Fern spores
Retitriletes sp.
6. Miscellaneous palynomorphs
Microforaminiferal wall linings
Unidentified sp. A
Unidentified sp. B
Unidentified sp. C
Unidentified sp. D
Appendix A (continued)
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223220
References
Batten, D.J., 1996. Palynofacies and paleoenvironmental interpre-
tation. In: Jansonius, J., McGregor, D.C. (Eds.), Palynology:
Principles and Applications. American Association of Strati-
graphic Palynologists Foundation, Dallas, pp. 1011–1064.
Beck, J.H., Strother, P.K., 2003. A method of palynofacies analysis
using cluster analysis of transformed relative abundance data.
36th Annual Meeting of the American Association of Strati-
graphic Palynologists, Program with Abstracts, unpaginated.
Bolli, H.M., Ryan, W.B.F., et al., 1978. Initial Reports Deep Sea
Drilling Project 40. U.S. Government Printing Office, Wash-
ington DC.
Bork, M., Zonneveld, K.A.F., 2003. Palaeoceanographic varia-
bility of the Benguela upwelling system depending on the
northern hemisphere glaciation (NHG)—indicated by organic-
walled dinoflagellates. Seventh International Conference on
Modern and Fossil Dinoflagellates, Program and Abstracts,
pp. 27.
Boulter, M.C., 1994. An approach to a standard terminology for
palynodebris. In: Traverse, A. (Ed.), Sedimentation of Organic
Particles. Cambridge University Press, pp. 199–216.
Brinkhuis, H., Pross, J., Riding, J.B., 2004. Jurassic–Cretaceous–
Tertiary dinoflagellate cyst course: morphology, stratigraphy and
(paleo)ecology. Short Course, 24–28 May, Tqbingen, Germany
(unpublished manual).
Christensen, B., Giraudeau, J., 2002. Neogene and Quaternary
evolution of the Benguela coastal upwelling system. Marine
Geology 180, 1–274.
Cross, A.T., Thompson, G.G., Zaitzeff, J.B., 1966. Source and
distribution of palynomorphs in bottom sediments, southern part
of Gulf of California. Marine Geology 4, 467–524.
Dale, B., 1996. Dinoflagellates cyst ecology: modeling and
geological implications. In: Jansonius, J., MacGregor, D.C.
(Eds.), Palynology: Principles and Applications. American
Association of Stratigraphic Palynologists Foundation, Dallas,
pp. 1249–1275.
Dale, B., Dale, A., 2002. Environmental applications of dino-
flagellate cysts and acritarchs. In: Haslett, S.K. (Ed.), Quater-
nary Environmental Micropalaeontology. Arnold, London,
pp. 207–240.
Dale, B., Fjells3, A., 1994. Dinoflagellate cysts as paleoproduc-
tivity indicators: state of the art, potential, and limits. In:
Zahn, R., et al., (Eds.), Carbon Cycling in the Glacial Ocean:
Constraints on the Ocean’s Role in Global Change. Springer-
Verlag, Berlin, pp. 521–537.
Dale, B., Dale, A.L., Jansen, J.H.F., 2002. Dinoflagellate cysts as
environmental indicators in surface sediments from the Congo
deep-sea fan and adjacent regions. Palaeogeography, Palae-
oclimatology, Palaeoecology 185, 309–338.
Davey, R.J., 1971. Palynology and paleo-environmental studies
with special reference to the continental shelf sediments of
South Africa. In: Farinacci, A. (Ed.), Proceedings of the Second
Planktonic Conference, Roma. Tecnoscienza, vol. 1,
pp. 331–347.
Davey, R.J., Rogers, J., 1975. Palynomorph distribution in Recent
offshore sediments along two traverses off South West Africa.
Marine Geology 18, 213–225.
Dean, W., Gardner, J., 1985. Cyclic variations in calcium carbonate
and organic carbon in Miocene to Holocene sediments, Walvis
Ridge, South Atlantic Ocean. In: Hsu, K.J., Weissert, H.J.
Page 23
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 221
(Eds.), South Atlantic Paleoceanography. Cambridge University
Press, pp. 61–78.
De Schepper, S., Head, M.J., Louwye, S., 2004. New dinoflagellate
cyst and incertae sedis taxa from the Pliocene of northern
Belgium, southern North Sea Basin. Journal of Paleontology 78,
625–644.
De Verteuil, L., Norris, G., 1996. Miocene dinoflagellate stratig-
raphy and systematics of Maryland and Virginia. Micropaleon-
tology 42, 1–172 (Supplement).
Diester-Haass, L., Meyers, P.A., Rothe, P., 1992. The Benguela
current and associated upwelling on the southwest African
margin: a synthesis of the Neogene–Quaternary sedimentary
record at DSDP sites 362 and 532. In: Summerhayes, C.P., Prell,
W.L., Emeis, K.C. (Eds.), Upwelling Systems: Evolution Since
the Early Miocene. Geological Society (London) Special
Publication, vol. 64, pp. 331–342.
Diester-Haass, L., Meyers, P.A., Vidal, L., Wefer, G., 2001. Data
report: sand fraction, carbonate, and organic carbon contents of
Late Miocene sediments from Site 1085, middle Cape Basin. In:
Wefer, G., Berger, W.H., Richter, C. (Eds.), Proceedings of the
Ocean Drilling Program. Scientific Results, vol. 175, pp. 1–23.
Diester-Haass, L., Meyers, P.A., Vidal, L., 2002. The Late
Miocene onset of high productivity in the Benguela current
upwelling system as part of a global pattern. In: Chris-
tensen, B., Giraudeau, J. (Eds.), Neogene and quaternary
evolution of the Benguela coastal upwelling system. Marine
Geology, vol. 180, pp. 87–103.
Edwards, L.E., 1984. Miocene dinoflagellate cysts from Deep
Sea Drilling Project Leg 181, Rockall Plateau, eastern
North Atlantic Ocean. In: Roberts, D.G., Schnitker, D. (Eds.),
Initial Reports of the Deep Sea Drilling Project, vol. 81,
pp. 581–594.
Edwards, L.E., 1992. New semiquantitative (paleo) temperature
estimates using dinoflagellate cysts, an example from the North
Atlantic Ocean. In: Head, M.J., Wrenn, J.H., et al., (Eds.),
Neogene and Quaternary Dinoflagellate Cysts and Acritarchs.
American Association of Stratigraphic Palynologists Founda-
tion, Dallas, pp. 69–87.
Edwards, L.E., Andrle, V.A.S., 1992. Distribution of selected
dinoflagellate cysts in modern marine sediments. In: Head,
M.J., Wrenn, J.H. (Eds.), Neogene and Quaternary Dinoflagel-
late Cysts and Acritarchs. American Association of Stratigraphic
Palynologists Foundation, Dallas, pp. 259–288.
Esper, O., Zonneveld, K.A.F., 2002. Distribution of organic-walled
dinoflagellate cysts in surface sediments of the southern ocean
(eastern Atlantic sector) between the Subtropical Front and
Weddell Gyre. Marine Micropaleontology 46, 177–208.
Esper, O., Versteegh, G.J.M., Zonneveld, K.A.F., Willems, H.,
2004. A palynological reconstruction of the Agulhas Retro-
flection (South Atlantic Ocean) during the Late Quaternary.
Global and Planetary Change 41, 31–42.
Faegri, K., Iversen, J., 1989. Textbook of Pollen Analysis. fourth
edition. John Wiley and Sons, Chichester. 328 pp.
Fensome, R.A., Taylor, F.J.R., Norris, G., Sarjeant, W.A.S.,
Wharton, D.I., Williams, G.L., 1993. A classification of fossil
and living dinoflagellates. Special Paper, vol. 7. Micropaleon-
tology Press. 351 pp.
Germeraad, J.H., Hopping, C.A., Muller, J., 1968. Palynology of
Tertiary sediments from tropical areas. Review of Palaeobotany
and Palynology 6, 189–348.
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., de Graciansky,
P.-C., Vail, P.R., 1998. Mesozoic and Cenozoic chronostrati-
graphic framework of European basins. In: de Graciansky, P.-C.,
Hardenbol, J., Jacquin, T., Vail, P.R. (Eds.), Mesozoic and
Cenozoic Sequence Stratigraphy of European Basins. SEPM
Special Publication, vol. 60, pp. 3–13.
Harland, R., 1983. Distribution maps of Recent dinoflagellate cysts
in bottom sediments from the North Atlantic Ocean and adjacent
seas. Palaeontology 26, 321–387.
Hay, W.W., Broock, J.C., 1992. Temporal variation in intensity of
upwelling off southwest Africa. In: Summerhayes, C.P., Prell,
W.L., Emeis, K.C. (Eds.), Upwelling Systems: Evolution Since
the Early Miocene. Geological Society (London) Special
Publication, vol. 64, pp. 463–497.
Head, M.J., 1996. Late Cenozoic dinoflagellates from the Royal
Society borehole at Ludham, Norfolk, eastern England. Journal
of Paleontology 70, 543–570.
Head, M.J., 1997. Thermophilic dinoflagellate assemblages from
the mid Pliocene of eastern England. Journal of Paleontology
71, 165–193.
Head, M.J., 1998. Marine environmental change in the Pliocene and
early Pleistocene of eastern England: the dinoflagellate evidence
reviewed. Mededelingen Nederlands Instituut voor Toegepaste
Geowetenschappen 60, 199–226.
Head, M.J., Norris, G., 2003. New species of dinoflagellate cysts
and other palynomorphs from the latest Miocene and Pliocene
of DSDP Hole 603C, western North Atlantic. Journal of
Paleontology 77, 1–15.
Holzwarth, U., Esper, O., Zonneveld, K., Willems, H., 2003. Spatial
distribution of organic-walled dinoflagellate cysts in surface
sediments of the Benguela upwelling system and their relation-
ship to environmental parameters. Seventh International Confer-
ence on Modern and Fossil Dinoflagellates, Program and
Abstracts, p. 51.
Hopkins, J.A., McCarthy, F.M.G., 2002. Post-depositional palyno-
morph degradation in Quaternary shelf sediments: a laboratory
experiment studying the effects of progressive oxidation.
Palynology 26, 167–184.
Jaramillo, C.A., Oboh-Ikuenobe, F.E., 1999. Sequence stratigraphic
interpretations from palynofacies, dinocyst and lithological data
of Upper Eocene–Lower Oligocene strata in southern Mis-
sissippi and Alabama, U.S. Gulf Coast. Palaeogeography,
Palaeoclimatology, Palaeoecology 145, 259–302.
Kovach, W.L., 2002. Multivariate statistical package plus. Version
3.1 Users’ Manual. Kovach Computing Services, Pentraeth,
Wales. 127 pp.
Laskar, J., 1993. Der Mond und die Stabilitaet des Erdklimas.
Spektrum-der-Wissenschaft-Verlagsgesellschaft. Federal Repub-
lic of Germany, Weinheim.
Lewis, J., Dodge, J.D., Powell, A.J., 1990. Quaternary dinoflagellate
cysts from the upwelling system offshore Peru, Hole 686B, ODP
Leg 112. In: Suess, E., Von Huene, R., et al., (Eds.), Proceedings
of the Ocean Drilling Program. Scientific Results, vol. 112,
pp. 547–553.
Page 24
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223222
Londeix, L., Benzakour, M., de Vernal, A., Turon, J.L., Suc, J.-P.,
1999. Late Neogene dinoflagellate cyst assemblages from the
Strait of Sicily, Central Mediterranean Sea: paleoecological and
biostratigraphical implications. In: Wrenn, J.H., Suc, J.P., Leroy,
S.A.G. (Eds.), The Pliocene: Time of Change. American
Association of Stratigraphic Palynologists Foundation, Dallas.
250 pp.
Louwye, S., Head, M.J., De Schepper, S., 2004. Dinoflagellate cyst
stratigraphy and palaeoecology of the Pliocene in northern
Belgium, southern North Sea Basin. Geological Magazine 141,
353–378.
Lutjeharms, J.R.E., 1996. The exchange of water between the South
Indian and South Atlantic Ocean. In: Wefer, G., Berger, W.H,
Siedler, G., Webb, D.J. (Eds.), The South Atlantic: Present and
Past Circulation. Springer-Verlag, Berlin, pp. 125–162.
Lyle, M., Dadey, K.A., Farrell, J.W., 1995. The late Miocene (11–8
Ma) eastern Pacific carbonate crash: evidence for reorganization
of deep-water circulation by the closure of the Panama Gateway.
In: Pisias, N.G., Mayer, L.A., Janecek, T.R., Palmer-Julson, A.,
van Andel, T.H. (Eds.), Proceedings of the Ocean Drilling
Program. Scientific Results, vol. 138, pp. 821–838.
Margalef, R., 1978. Life-forms of phytoplankton as survival
alternatives in an unstable environment. Oceanologica Acta 1,
493–509.
Marret, J., Zonneveld, K.A.F., 2003. Atlas of modern organic-
walled dinoflagellate cyst distribution. Review of Palaeobotany
and Palynology 125, 1–200.
Melia, M.B., 1984. The distribution and relationship between
palynomorphs in aerosols and deep-sea sediments off the coast
of northwest Africa. Marine Geology 58, 345–371.
Oboh, F.E., 1992. Multivariate statistical analyses of palynodebris
from the Middle Miocene of the Niger delta and their environ-
mental significance. Palaios 7, 559–573.
Oboh-Ikuenobe, F.E., 2001. A palynological record of
Middle Miocene to Lower Pliocene sedimentary rocks,
ODP Hole 1085A (Cape Basin, southwestern Africa). EOS
Transactions-AGU 82 (20), S228 (Spring Meeting Supplement,
OS42A-05).
Oboh-Ikuenobe, F.E., Hoffmeister, A.P., Chrisfield, R.A., 1999.
Cyclical distribution of dispersed organic matter and dinocysts,
ODP Site 959 (early Oligocene–early Miocene, Cote d’Ivoire–
Ghana Transform Margin). Palynology 23, 87–96.
Pagani, M., Arthur, M., Freeman, K.H., 1999. Miocene
evolution of atmospheric carbon dioxide. Paleoceanography
14, 273–292.
Powell, A.J., Lewis, J., Dodge, J.D., 1990. Late Neogene to
Pleistocene palynological facies of the Peruvian continental
margin upwelling, Leg 112. In: Suess, E., Von Huene, R., et al.,
(Eds.), Proceedings of the Ocean Drilling Program. Scientific
Results, vol. 112, pp. 297–321.
Powell, A.J., Lewis, J., Dodge, J.D., 1992. The palynological
expressions of post-Paleogene upwelling: a review. In: Sum-
merhayes, C.P., Prell, W.L., Emeis, K.C. (Eds.), Upwelling
Systems: Evolution Since the Early Miocene. Geological
Society Special Publication, vol. 64, pp. 215–226.
Pufahl, P.K., Maslin, M.A., Anderson, L., Brqchert, V., Jansen, F.,Lin, H., Perez, M., Shipboard Scientific Party, 1998. Lithostrati-
graphic summary for Leg 175: Angola–Benguela upwelling
system. In: Wefer, G., Berger, W.H., Richter, C., et al., (Eds.),
Proceedings of the Ocean Drilling Program. Initial Reports, vol.
175, pp. 533–542.
Reynolds, C.S., 1987. Community organization in the freshwater
plankton. Symposium of the British Ecological Society 27,
297–325.
Rochon, A., de Vernal, A., Turon, J.-L., Matthiesen, J., Head, M.J.,
1999. Distribution of recent dinoflagellate cysts in surface
sediments from the North Atlantic and adjacent seas in relation
to sea-surface parameters. Contributions Series-American Asso-
ciation of Stratigraphic Palynologists 35, 1–152.
Roth, J.M., Droxler, A.W., Kameo, K., 2000. The Caribbean
carbonate crash at the middle to late Miocene transition; linkage
to the establishment of the modern global ocean conveyor.
Proceedings of the Ocean Drilling Program. Scientific Results
165, 249–273.
Schrank, E., 1988. Effects of chemical processing on the preserva-
tion of peridinoid dinoflagellate cysts: a case from the Late
Cretaceous of NE Africa. Review of Palaeobotany and
Palynology 56, 123–140.
Siesser, W.G., 1980. Late Miocene origin of the Benguela upwelling
system off northern Namibia. Science 208, 283–285.
Smayda, T.J., Reynolds, C.S., 2001. Community assembly in
marine phytoplankton: application of recent models to
harmful dinoflagellate blooms. Journal of Plankton Research
23, 447–461.
Sokal, R.R., Michener, C.D., 1958. A statistical method for
evaluating systematic relationships. University of Kansas
Bulletin 38, 109–1438.
Strauss, C., Lund, J.J., Lund-Christensen, J., 2001. Miocene
dinoflagellate cyst biostratigraphy of the Nieder Ochtenhausen
Research Borehole (NW Germany). Geologische Jahrbuch A
152, 395–447.
Suc, J.P., Diniz, F., Leroy, F., Poumot, C., Bertini, A., Dupont, L.,
Clet, M., Bessais, E., Zheng, Z., Fauquette, S., Ferrier, J., 1995.
Zanclean (Brunssumian) to Early Piacenzian (Early–Middle
Reuverian) climate from 48 to 548 north latitude (West Africa,
West Europe and West Mediterranean areas). Mededelingen-
Rijks Geologische Dienst 52, 43–56.
Toggweiler, J.R., 1999. Variation of atmospheric CO2 by ven-
tilation of the ocean’s deepest water. Paleoceanography 14,
571–588.
Turon, J.L., 1984. Le palynoplankton dans l’environement de
l’Atlantique nord-oriental. Evolution climatique et hydrologique
depuis le dernier maximum glaciar. Memoires de l’institut de
Geologie du Bassin d’Aquitaine 17, 1–313.
Tyson, R.V., 1987. The genesis and palynofacies characteristics of
marine petroleum source rocks. In: Brooks, J., Fleet, A.J. (Eds.),
Marine Petroleum Source Rocks. Special Geological Society,
London, Publication, vol. 26, pp. 46–47.
Tyson, R.V., 1995. Sedimentary Organic Matter: Organic Facies and
Palynofacies. Chapman and Hall, London. 615 pp.
Udeze, C.U., 2003. Neogene paleoceanographic and paleoclimatic
signals from palynomorphs and dispersed organic matter in the
Cape Basin, ODP Leg 175. MS Thesis, University of Missouri-
Rolla, 91 pp.
Page 25
C.U. Udeze, F.E. Oboh-Ikuenobe / Palaeogeography, Palaeoclimatology, Palaeoecology 219 (2005) 199–223 223
Versteegh, G.J.M., Zevenboom, D., 1995. New genera and species
of dinoflagellate cysts from the Mediterranean Neogene. Review
of Palaeobotany and Palynology 85, 213–229.
Versteegh, G.J.M., Zonneveld, K.A.F., 2002. Use of selective
degradation to separate preservation from productivity. Geology
30, 615–618.
Wall, D., Dale, B., Lohmann, G.P., Smith, W.K., 1977. The
environmental and climatic distribution of dinoflagellate cysts
in modern marine sediments from regions in the North and
South Atlantic. Marine Micropaleontology 2, 121–200.
Wefer, G., Berger, W.H., Richter, C., et al., 1998. Proceedings of the
Ocean Drilling Program. Initial Reports 175, 1–1563.
Westerhold, T., Bockert, T., Rfhl, U. Middle to late Miocene
oxygen isotope stratigraphy of ODP site 1085 (SE Atlantic):
new constrains on Miocene climate variability and sea-level
fluctuations. Palaeogeography, Palaeoceanography, Paleaoeco-
logy, 217, in press.
Williams, G.L., Lentin, J.K., Fensome, R.A., 1998. The Lentin
and Williams index of fossil dinoflagellates. Contributions
Series-American Association of Stratigraphic Palynologists 34,
1–817.
Wrenn, J.H., Kokinos, J.P., 1986. Preliminary comments on
Miocene through Pleistocene dinoflagellate cysts from De
Soto Canyon, Gulf of Mexico. In: Wrenn, J.H., Duffield,
S.L., Stein, J.A. (Eds.), Papers from the First Symposium on
Neogene Dinoflagellate Cyst Biostratigraphy. Contributions
Series-American Association of Stratigraphic Palynologists,
vol. 17, pp. 169–225.
Wrenn, J.H., Suc, J.P., Leroy, S.A.G., 1999. The Pliocene: time of
change. American Association of Stratigraphic Palynologists
Foundation, Dallas. 250 pp.
Zagwijn, W.H., 1960. Aspects of the Pliocene and Early Pleistocene
vegetation in the Netherlands. Mededelingen van de Geo-
logische Stichting C III (I) 5, 1–78.
Zonneveld, K.A.F., Versteegh, G.J.M., De Lange, G.J., 1997.
Preservation of organic-walled dinoflagellate cysts in different
oxygen regimes: a 10,000 year natural experiment. Marine
Micropaleontology 29, 393–405.
Zonneveld, K.A.F., Hoek, R., Brinkhuis, H., Willems, H., 2001a.
Lateral distribution of organic-walled dinoflagellates in surface
sediments of the Benguela upwelling region. Progress in
Oceanography 46, 25–72.
Zonneveld, K.A.F., Versteegh, G.J.M., De Lange, G.J., 2001b.
Palaeoproductivity and post-depositional aerobic organic matter
decay reflected by dinoflagellate cyst assemblages of the Eastern
Mediterranean S1 sapropel. Marine Geology 172, 181–195.