Citation: LI Y Y, LI Z F, HE Y F, et al. Phylogeographic analysis of human papillomavirus 58. Sci China Ser C-Life Sci, 2009, 52(12): 1164-1172 doi:10.1007/s11427-009-0149-6
www.scichina.com life.scichina.com
www.springer.com/scpwww.springerlink.com
Phylogeographic analysis of human papillomavirus 58
LI YanYun1, LI ZuoFeng2, HE YiFeng1, KANG Yu1, ZHANG XiaoYan1, CHENG MingJun1, ZHONG Yang2, 3† & XU CongJian1† 1 Obstetrics and Gynecology Hospital of Fudan University, Shanghai 200011, China; 2 Shanghai Bio-information Research Center, Shanghai 200235, China; 3 School of Life Sciences, Fudan University, Shanghai 200433, China
Human papillomavirus 58 (HPV58) is one type of HPV with high risk of causing cervical cancer. Unusually high prevalence of HPV58 has been reported in Asia, Africa and some other areas. However, due to the scattered distribution of global data, in addition to the lack of data of some HPV58 high-incidence nations and regions, like Mainland China, a comprehensive analysis of the global geographical distribution of HPV58 remains blank so far. In this study, HPV58 from the human cervical cancer tissue was detected in Mainland China, and 14 new HPV58-E6/L1 gene sequences were obtained. Moreover, phylogeographic analysis has been conducted combining the HPV58 sequences that have been deposited in GenBank since 1985. The study result shows that the sequences detected from the Shanghai, Jiangsu and Sichuan areas are homologous with those found in the past from Hong Kong and Xi’an, China, as well as Japan and other Southeast Asian areas. Furthermore, Western Africa is considered to be the “root” source of the HPV58 variant, while Mainland China and Southeast Asia are “transit points” and the new sources of HPV58 after receiving the isolates from the “root” source; like HPV16 and HPV18, the HPV58 might also be one of the major HPV types associated with the development and spread of cervical cancer.
human papillomavirus 58 (HPV58), cervical cancer, phylogeography, molecular evolution
Cervical cancer is one of the major diseases threaten- ing women’s health all over the world. Human papillo- mavirus (HPV) infection is recognized as the direct cause of cervical cancer and the primary factor inducing this cancer[1−3]. Before this study, eighteen types of HPV had been classified as high-risk due to their frequent association with anogenital cancer. The most common type among them are HPV16 and HPV18, which are only remotely related to one another and form two separate phylogenetic groups, each together with 6 other HPV types. These groups are now called HPV species 9 and 7, respectively[4]. As a member of the HPV16 phylogenetic group, HPV58 is high-risk but much rarer than HPV16 and HPV18, though it is gradually attrac- ting more concern because of its special geographic distribution and regional high-risk of causing cervical cancer. Although HPV58 infection is not very common worldwide (the positive infection rate is only 2%
worldwide[4], ranked 7th of all HPV types), its unusually high prevalence has been reported in Asia, Africa and some other areas (the positive infection rate has been reported to be up to 6%—30%, ranked 2nd just following HPV16[5,
6]). Mainland China is an HPV58
high incidence area. From 1996 to 2007, it has been frequently reported that the positive rate of HPV58 infection in patients with cervical cancer was second only to HPV16, and in some areas of Mainland China, even higher than HPV16 and HPV18. These figures are far higher than those reported in literature from abroad
Received October 17, 2008; accepted June 9, 2009 doi: 10.1007/s11427-009-0149-6 †Corresponding author (email: [email protected]; [email protected]) Supported by the National Science and Technology Pillar Program of China (Grant No. 2007BAI24B01) and the National High Technology Research and Development Program of China (Grant No. 2006AA02Z342) the Shanghai Leading Academic Discipline Project (B111) and Shanghai Science and Technology Committee (Grant No. 07XD14025).
Article
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1165
during that time. Especially in Shanghai, Jiangsu, Sichuan, Guangdong, etc., the situation was particularly evident[6−13].
The unique geographic diffusion of HPV58 can be largely traced from the perspective of the virus’s victim (women), whose infection has been identified by clinical tests. Another approach tracks the migration from the perspective of the virus alone, by way of a phylogeo- graphy of HPV58 genetic sequences. Genetic phylogeo- graphy not only represents the “gold standard” in characterizing viral genealogy, transmission, and ancient spread, but also provides a powerful epidemiological tool to reveal the association of HPV 58 with cervical cancer. A similar approach has been successfully used for the analysis of avian influenza virus[14]. However, due to the scattered distribution of global data, in addition to the lack of data for some HPV58 high-inci- dence nations and regions, the phylogeographical analysis of the global geographical distribution of HPV58 has not been reported so far. None of literatures reviewed[4,15,16] has analytically explored global phylo- geography of HPV58 comprehenively and the history of the virus’s migration. It is worth noting that Mainland China, as an exceptionally high incidence area of HPV58, has little existing sequence data of HPV58 for analysis. Thus, the analysis based on existing data cannot cover certain regions of HPV58’s distribution such as Mainland China.
In this study,we collected HPV-positive human
cervical cancer tissue samples across localities of Mainland China, and then completed typing and gene sequencing. In order to reconstruct HPV58’s global evolution path and to explore the important role Mainland China played in the process of worldwide transmition and migration, we conducted a phylogeo- graphic analysis by combining the newly obtained HPV58 sequences with those that have been deposited in GenBank database since 1985.
1 Material and methods
1.1 Sample source
Tissue samples of cervical cancer embedded in paraffin were collected in Jiangsu Province, Sichuan Province, and Shanghai City of Mainland China from 1996 to 2007 (50 cases in each region). Each sample was cut into 10—20 μm thick sections, which were checked by pathologists. After DNA extraction, PCR amplification and sequencing, 7 HPV58-E6 and 7 HPV58-L1 nucleotide sequences were obtained, all of which were submitted to GenBank in 2008 (Table 1).
1.2 Detection of sequences
After xylene dewaxing, alcohol dehydration and other steps, DNA products were extracted from samples using the genomic DNA extraction kit (purchased from Beijing Tianwei Ear Technology Co., Ltd.). Extracted DNA products were preserved at −20℃. HPV-DNA was
Table 1 Newly obtained HPV58 sequences
GenBank accession No Gene Source Length/bp Collection year
EU999957 L1 Sichuan 1125 2005
EU999958 L1 Sichuan 1125 2005
EU999959 L1 Jiangsu 1124 2004
EU999960 L1 Shanghai 1180 2006
EU999961 L1 Shanghai 1168 2006
EU999962 L1 Jiangsu 1135 2004
EU999963 L1 Jiangsu 1143 2004
EU999964 E6 Sichuan 450 2005
EU999969 E6 Sichuan 436 2005
EU999970 E6 Jiangsu 435 2004
EU999965 E6 Shanghai 450 2006
EU999966 E6 Shanghai 450 2006
EU999967 E6 Jiangsu 450 2004
EU999968 E6 Jiangsu 450 2004
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1166
amplified by PCR by using HPV-L1 universal primer MY09/MY11 to amplify all types of HPV. Then the positive samples were amplified by PCR using HPV58-E6 specific primers and L1 specific sectional primers to obtain final HPV58-E6 DNA fragments (400—500 bp) and HPV58-L1 DNA sectional fragments (300—500 bp). Both positive control (HeLa cells) and negative control (distilled water) were included in each PCR reaction, which was repeated at least three times. PCR products were analyzed by 2% agarose gel electrophoresis and photographed by the gel image analysis system. All PCR reactions were performed by PCR System 2700 from Applied Biosystems Company. According to a previous reference[17], HPV-L1 universal primers MY09/MY11 are 5′-CGTCCMARRGGAWACT- GATC-3′ (MY09) and 5′-GCMCAGGGWCATAAYAA- TGG-3′ (MY11). We designed HPV58-E6 specific primers and L1 specific sectional primers by ourselves as follows: Primer E6 A: 5′-ACCTCCTCCGTCTGCC- AGTT- 3′; Primer E6 B: 5′-ATAGTGTAAGTACCACA- ACAATTAT-3′; L1-1R(bp6004C): 5′-TGACCACTTAC- GCCAACACC-3′, L1-1F(bp5600): 5′-ATTTTGCGTC- GCAGACGTAA-3′; L1-2R(bp6494C): 5′-TGCAGTAT- TACCGGACCCTT-3′, L1-2F(bp5965): 5′-TAGGTAG- GGGACAGCCATTG-3′; L1-3R(bp6913C): 5′-GCCT- GGGAGGTAACAAATCTAT-3′, L1-3F(bp6424): 5′-TT- AATAGGGCTGGAAAACTTGG-3′; L1-4R(bp7280C): 5′-CAGGAAACTGACAAGGACATAGA-3′, L1-4F- (bp6840): 5′-GACTGGCAATTTGGTTTAACAC-3′. They are synthesized by Invitrogen Biotechnology Limited Company (Shanghai, China).
PCR products were sent to Fudan University for genetic sequencing by the ABI PRISM 310 automated DNA sequencer. A two-way connection for the full- length sequence was performed. Since the L1 gene was too long (1575 bp), it was divided into four sections for sequencing and combined later. Sequences that were newly obtained were compared with those deposited in GenBank.
1.3 Data analysis
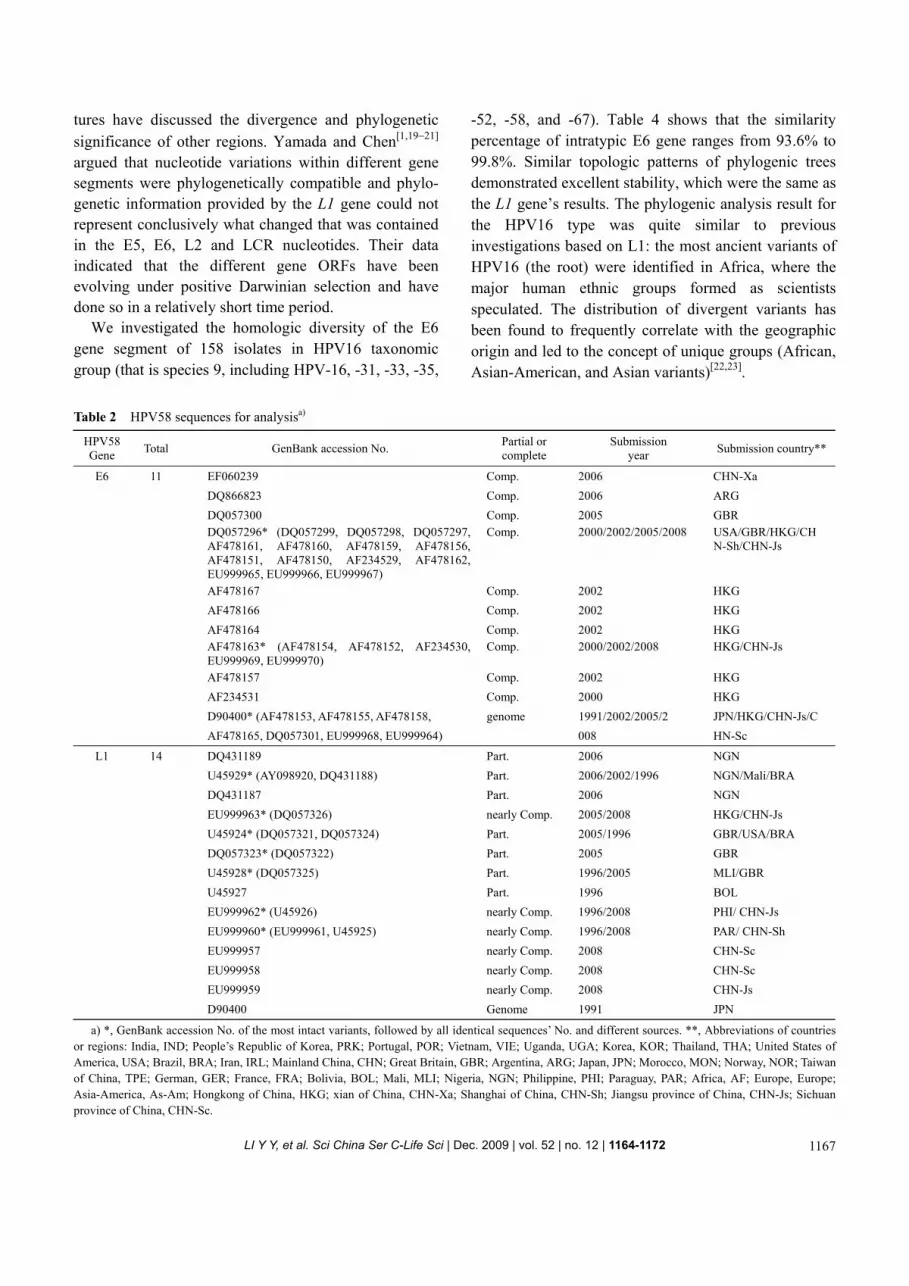
30 isolates of the complete E6 gene sequence (450 bp) and 17 isolates of the partial L1 gene sequence (359—449 bp) were downloaded from GenBank. We combined them with 14 newly obtained sequences to build a data set. We selected the identical sequences and only left the most complete ones of them behind, noted by different
source loca. 11 variants of E6 and 14 variants of L1 remained (Table 2).
158 isolates sequences of the HPV16 phylogenic group (HPV-16, -31, -33, -35, -52, -58, and -67), whose E6 genes are functional and complete (450 bp), or nearly complete (≥385 bp), were download from GenBank. After selection, 93 different variants remained for analysis (Table 3).
Homologic analyses of intratypic HPV nucleotide sequences were performed by DNASTAR software. Confidence intervals between the maximum and the minimum represented the similarity percentage among variants. Gene sequences were aligned using the Clustal W software program. Phylogenetic trees were nstructed by Maximum Parsimony (MP) method. Bootstrap analysis with 100 replicates of heuristic searches was used to test the confidence of the tree and each node. BPV1 (NC 001522 ) and HPV33 (PPH33CG ) were set as the out-group taxon.
The two phylogenetic trees of E6 and L1 were matched with each other with identical geographic loca between branches, and then marked in the map of the world for construction of HPV58’s evolutional path: nodes in the map stood for distinct loca where isolates were sampled, while the arrows between nodes could be inferred to represent the virus’s migratory direction according to the topologic structure of trees. The thickness of arrows from node to node stands for the gradient of branches in phylogenetic trees and succession of migration events among loca. In the end, we integrated HPV58’s migration path into the statistic map of cervical cancer incidence all over the world.
2 Results and discussion
2.1 Typing potential of the E6 gene
The HPV genome is a small, circular, double stranded DNA, which consists of three genetic regions: early transcription regions encoding the E1—E8 proteins, late transcription regions encoding the L1 and L2 proteins, and a non-coding region called the upper region of regulation (URR) or long control region (LCR)[18]. The L1 region, which is considered relatively conservative, is a traditional standard for the virus’s typing. An HPV type is defined as a separate taxon when the nucleotide sequence of its L1 gene differs from that of any other HPV type by at least 10%. Although L1 is traditionally used for typing and analysis, recently published litera-
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1167
tures have discussed the divergence and phylogenetic significance of other regions. Yamada and Chen[1,19−21]
argued that nucleotide variations within different gene segments were phylogenetically compatible and phylo- genetic information provided by the L1 gene could not represent conclusively what changed that was contained in the E5, E6, L2 and LCR nucleotides. Their data indicated that the different gene ORFs have been evolving under positive Darwinian selection and have done so in a relatively short time period.
We investigated the homologic diversity of the E6 gene segment of 158 isolates in HPV16 taxonomic group (that is species 9, including HPV-16, -31, -33, -35,
-52, -58, and -67). Table 4 shows that the similarity percentage of intratypic E6 gene ranges from 93.6% to 99.8%. Similar topologic patterns of phylogenic trees demonstrated excellent stability, which were the same as the L1 gene’s results. The phylogenic analysis result for the HPV16 type was quite similar to previous investigations based on L1: the most ancient variants of HPV16 (the root) were identified in Africa, where the major human ethnic groups formed as scientists speculated. The distribution of divergent variants has been found to frequently correlate with the geographic origin and led to the concept of unique groups (African, Asian-American, and Asian variants)[22,23].
a) *, GenBank accession No. of the most intact variants, followed by all identical sequences’ No. and different sources. **, Abbreviations of countries or regions: India, IND; People’s Republic of Korea, PRK; Portugal, POR; Vietnam, VIE; Uganda, UGA; Korea, KOR; Thailand, THA; United States of America, USA; Brazil, BRA; Iran, IRL; Mainland China, CHN; Great Britain, GBR; Argentina, ARG; Japan, JPN; Morocco, MON; Norway, NOR; Taiwan of China, TPE; German, GER; France, FRA; Bolivia, BOL; Mali, MLI; Nigeria, NGN; Philippine, PHI; Paraguay, PAR; Africa, AF; Europe, Europe; Asia-America, As-Am; Hongkong of China, HKG; xian of China, CHN-Xa; Shanghai of China, CHN-Sh; Jiangsu province of China, CHN-Js; Sichuan province of China, CHN-Sc.
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1168
2.2 Characteristics of HPV58 variants in Mainland China and its phylogenic relationship
We have newly obtained five full-length (450 bp) HPV58-E6, two nearly full-length (435 and 436 bp) E6 sequences and seven nearly full-length (1124—1480 bp) HPV58-L1 sequences from cervical cancer tissue samples in Mainland China (from Sichuan, Jiangsu and Shanghai). Gene homology analysis shows that the newly obtained nucleotide sequences have high similarity with the ones from GenBank: E6: 97.3%—
99.8%; L1: 97.6%—99.8%, which are confirmed to be the same type based on the law of typing[24−26] (Table 4). BLASTed with GenBank’s standard strains, both identical and new variants were confirmed. In E6 sequences: 3 isolates (EU999965, EU999966, EU999- 967) coming from Shanghai and Jiangsu were identical with isolate DQ057296 (from Hong Kong, the United States, and Germany); 2 isolates (EU999968, EU999964) from Jiangsu and Sichuan were confirmed to be identical with D90400 (from Japan and Hong Kong), and 2 isolates (EU999969, EU999970) from Sichuan and Jiangsu were identical with AF478163 (from Hong
Kong). In L1 sequences: 2 isolates (EU999960, EU999961) from Shanghai were identical with U45925 (from Paraguay), one isolate of Jiangsu (EU999962) was identical with U45926 (from Philippines), isolate EU999963 coming from Jiangsu was identical with DQ057326 (from Hong Kong), while the other 3 series (EU999957, EU999958, EU999959) from Sichuan and Jiangsu were new variants (Table 2). Finally we integrated the Chinese data into the global data system and conducted two phylogenic trees based on E6 and L1, respectively (Figure 1, marking Chinese data with red font).
The phenomena from the two phylogenetic trees are as follows: Homologous strains of E6 (AF478163, D90400) appeared in Japan, Hong Kong, Jiangsu, and Sichuan of China. Homologous strains of E6 (AF478164, DQ057296) emerged in Hong Kong, Jiangsu, and Shanghai. Homologous strains of L1 (EU999963, D90400, EU999958) emerged in Hong Kong, Jiangsu, Sichuan, and Japan. Another set of homologous strains (EU999959, EU999960, EU999957, EU999962) appea- red in Jiangsu, Shanghai, Sichuan, Paraguay and Philippines. To sum up, the newly obtained sequences in
Table 3 E6 sequences in HPV16 phylogenic groupa)
Type Sum of variants Source* Year first
submission Country first submission* GenBank accession No.
HPV67 1 JPN 1998 JPN D21208 a) *, Abbreviations of countries or regions are the same with Table2.
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1169
Mainland China are more homologous with the strains from Hong Kong, Xi’an, Japan, Philippines, and other areas of Southeast Asia, and they are prone to gather in the lower part of the tree. This suggests us that Mainland China and Southeast Asia may format the unique “transit points” of HPV58 strain dissemination after receiving variants from the “root”. Mainland China and Southeast Asia may play an important role in HPV58’s distribution and migration all over the world.
2.3 Phylogeographic analysis of HPV58 and its significance
Results from Clifford GM’s Meta analysis[5] show that HPV58 is more frequently identified in invasive cervical cancer (6%) than HPV31, HPV33, and HPV45 in Asia
and South Africa, much higher than that in Europe, North America, Australia, and so on. In some cases, HPV58 has become a dominant type even exceeding HPV16 and HPV18. Xi et al.[27] pointed out that, in addition to HPV16, HPV58 was the most frequently detected HPV type associated with cervical precancer ous lesions in women of Western Africa. Wu et al.[8], Chan et al.[6], Lai et al.[12], Xin et al.[28], and Cho et al.[29] have also reported unusually high prevalence of HPV58 in the populations living in Mainland China, China Hongkong, China Taiwan, Japan, and Korea. In these areas, HPV58 was regarded as the second most common HPV type in cervical cancer after HPV16 and its prevalence was associated statistically with the severity of neoplasia, which has aroused much attention all over the world.
Table 4 Homologic analysis of intratypic HPV nucleotide sequences
Type and Gene Sum of isolates Confidence interval of similarity percentage (%)
HPV16-E6 55 96.2—9.8
HPV31-E6 9 98.4—99.8
HPV33-E6 3 93.6—99.3
HPV35-E6 3 99.3—99.6
HPV52-E6 10 97.8—99.6
HPV58-E6 11 97.3—99.8
HPV58-L1 14 97.6—99.8
HPV67 1 −
Figure 1 Two phylogenic trees of HPV58-E6, -L1. Left is the E6 tree, right is the L1 tree. The data set for analysis includes newly obtained sequences and GenBank sequences (the identical sequences merged, noting different source loca). HPV33 (PPH33CG) was set as an out-group taxon. Phylogenetic trees were constructed by MP method. Abbreviations of countries or regions are the same as with Table 2.
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1170
How has such unique distribution of HPV58 formed? Is it related to migration and evolution of HPV variants, or only linked to the susceptibility of the population and screening discrimination in different areas? Genetic phylogeography may provide a good method for us, which can not only track the viral migration and ancient spread but also provide a powerful epidemiological tool for revealing the association of HPV 58 with cervical cancer. Previously, phylogeography has been used for analysis of HPV16 by scientists and has given us inspiration and clues. Variance of HPV has been recognized as relatively limited for a long time, because diversity of the nucleotide sequence in L1 ranged within 2%. Therefore, the identification of molecular HPV variants is considered a powerful epidemiological and phylogenic tool to reveal the ancient spreading route of HPVs, while its trace through the world has not yet been completely lost[30]. The variants specific for some regions formed at the same time when major human ethnic groups formed. Consequently, evolution of HPV is reflective of patterns of migration and evolution of humankind worldwide. However, there are only a few previous studies about phylogeography of HPV58. In 2005, Calleja-Macias IE[4] found that no clear similari- ties between phylogenetic topologies of HPV58 and that of HPV16 existed. The tree of HPV58 based on 21 variants did not reflect human evolution and spread to the extent illustrated by HPV16. Two other studies based on partial L1 sequences from seven[15]and eight[16] genomic variants of HPV-58 showed the same results.
In the current study, we constructed phylogenic trees of E6 and L1 separately and matched them through geographic mapping between variants (branches). From the E6 tree in Figure 1, we can see that the root of the tree is the isolate AF234531 coming from Hongkong. On lower phylogenic levels, the trunk of the tree is divided into two main branches, one of which is located in Hong Kong and Europe, while the other in Southeast Asia and Latin America. With more global coverage of geographic information, the L1 phylogenetic tree showed a global trend: the variant of the root of the tree (DQ431189) was identified in Western Africa; all of the variants from Africa(U45929, U45927, DQ431187, U45928)are completely confined to the trunk of the tree. When the virus was introduced to Southeast Asia (EU999962, Jiangsu and Philippines), it formed a major branch. The lower topologic pattern of phylogeny is
similar to the E6 tree. Variants of tip branches build up phylogenetic clusters for European, Latin and Southeast Asian variants. To analyze these results, we tracked HPV58’s evolution path by marking migratory events in the map of the world based on the two phylogenic trees (Figures 2A and B)) and superimposing them on Figure 2C.
With the careful analysis of Figure 2C, we have drawn some conclusions as follows: the E6 tree which is
Figure 2 Phylogeographic analysis of HPV58. A, HPV58-E6’s migratory path reconstructed; B, HPV58-L1’s migratory path recons- tructed; C, A + B. The global maps of cervical cancer incidence statistics were introduced as the background of HPV58’s migration path. Thickness of arrows from node to node stands for the gradient of branches in phylogenic trees. Abbreviations of countries or regions are the same with Table 2.
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1171
lacking African data misplaces Southeast Asia (Hong Kong) as the “phylogenic root”, while the L1 tree demonstrates the relatively reliable and actual origin of ancient HPV58. The two trees complementally reinforced each other and matched very well. HPV 58 variants analyzed in this research were isolated within the last two decades and may reveal the specific patterns of HPV58 evolution and spread over a long period of time. Western Africa is probably the original source of ancient HPV58, while Southeast Asia appears to be the next “relay center” or another new weightily “cradleland” of variants, demonstrating multi-directional dispersal potential influencing other regions in Southeast Asia, Europe, North America, Latin America, etc. Therefore, Africa and South Asia are the most important reservoirs of the virus’s variants, which precisely meet the epidemiologic result of high HPV58’s positive rate in these areas. It is inferred that the unusually high prevalence is probably largely related to preoccupant existence (or introduction) of HPV58 variants in Africa and South Asia. But another possible explanation, that the ancient variants (closer to the “root”) are more virulent, cannot be excluded.
Neither Europe nor Latin America turned into a “relay center” for HPV58 like Hongkong, although both of them have encountered the attack of variants several times, some of which even come from ancient and virulent isolates. There are two possible explanations. First, variants have changed. All variants introduced into these areas have encountered some mutations in order to adapt to the environment’s change such as change of specific race, weather, and geographic environment. Those mutations could weaken the variants’ virulence. This explanation is equivalent to DeFilippis VR’s[31]
argument in 2002. Second, susceptibility of population differs in these areas. The susceptibility of populations in Europe and Latin America is possibly lower than that
in Africa and South Asia. In order to investigate the potential associations
between HPV58 migration events and global spread of cervical cancer, we intentionally designated the WHO statistic chart of cervical cancer’s incidence as the background of HPV58’s migratory path (Figures 2A, B and C). It showed us that Africa suffers most for cervical cancer all over the world; India, Mongolia, Southeast Asian island and some regions of Latin America are also in the high-incidence trend; while Europe and North America have much lower incidence. Such special distribution of cancer coincides surprisingly with the successive migratory movement of HPV58 (“Africa→ South Asia→Europe, North America”).
In summary, HPV58 may have an important part in the prevalence of cervical cancer in distinct regions. As an ancient HPV type derived from Africa , HPV58 is also a main type associated with cervical cancer just like HPV16 and HPV18. At present, all of the existing vaccines for cervical cancer that have been developed for clinical application are against HPV16 and HPV18[32,33]. Therefore, our study provides a new concept for exploiting or developing a vaccine for some specific areas (such as West Africa and Southeast Asia). It is worth noting that the virulence of HPV58 variants decreased gradually as the frequency and order of migration increased, but this is not the case in South Asia. We would like to thank Larissa Wenren (Harvard University) and Joan Kirtland (University of Virginia) for their comments and suggestions. Specimens were offered by People's Hospital of Guangyuan City in Sichuan Province, First People’s Hospital of Huaian City in Jiangsu Province, Public Benefit Hospital of Shanghai City, and Central Hospital of Huizhou City in Guangdong Province
1 Lombard I, Vincent-Salomon A, Validier P, et al. Human papilloma-
virus genotype as a major determinant of the course of cervical. J Clin Oncol, 1998, 16: 2613―2619
2 Mclachlin C M. Human papillomavirus in cervical neoplasia: Role, risk factors and implications. Clin Lab Med, 2000, 20: 257―270
3 Franco E L, Duarte-Franco E, Ferenczy A. Cervical cancer: Epidemiology, prevention and the role of human papillomavirus infection. CMAJ, 2001,164: 1017―1025
4 Calleja-Macias I E, Villa L L, Prado J C, et al. Worldwide genomic diversity of the high-risk human papillomavirus types 31, 35, 52, and 58, four close relatives of human papillomavirus type 16. J Virol, 2005, 79: 13630―13640
5 Clifford G M, Smith J S, Plummer M, et al. Human papillomavirus types in invasive cervical cancer worldwide: A meta-analysis. Br J Cancer, 2003, 88: 63―73
6 Chan P K, Lam C W, Cheung TH, et al. Association of human papillomavirus type 58 variant with the risk of cervical cancer. J Natl Cancer Inst, 2002, 94: 1249―1301
7 Lin Q Q ,Yu S Z,Qu W, et al. Human papillomavirus types 52 and 58. Int J Cancer, 1998, 75: 484―485
8 Wu Y, Chen Y, Li L, et al. Associations of high-risk HPV types and viral load with cervical cancer in China. J Clin Virol, 2006, 35: 264―272
9 Huang S, Afonina I, Miller B A, et al. Human papillomavirus types 52
LI Y Y, et al. Sci China Ser C-Life Sci | Dec. 2009 | vol. 52 | no. 12 | 1164-1172 1172
and 58 are prevalent in cervical cancer from Chinese women. Int J Cancer,1997, 70: 408―411
10 Liu J H, Huang X, Liao G W, et al. The comparative study of HPV infection in cervical cancer patients and other risk factors between China and Australia. Chin J Med, 2003, 83: 748―752
11 Lai H C, Sun C A, Yu M H, et al. Favorable clinical outcome of cervical cancers infected with human papilloma virus type 58 and related types. Int J Cancer, 1999, 84: 553―557
12 Lai C H, Huang H J, Hsueh S, et al. Human papillomavirus genotype in cervical cancer: A population-based study. Int J Cancer, 2007,120: 1999―2006
13 Chan P K, Li W H, Chan M Y, et al. High prevalence of human papillomavirus type 58 in Chinese women with cervical cancer and precancerous lesions. J Med Virol, 1999, 59: 232―238
14 Wallace R G, Hodac H, Lathrop R H, et al. Lathrop, et al. A statistical phylogeography of influenza A H5N1. Proc Natl Acad Sci USA, 2007, 104: 4473―4478
15 Stewart A C, Eriksson A M, Manos M M, et al. Intratype variation in 12 human papillomavirus types: A worldwide perspective. J Virol, 1996, 70: 3127―3136
16 Cerqueira D M, Camara G N, da Cruz M R, et al. Variants of human papillomavirus types 53, 58 and 66 identified in Central Brazil. Virus Genes, 2003, 26: 83―87
17 Qu W, Jiang G, Cruz Y, et al. PCR detection of human papillomavirus: Comparison between MY09/MY11 and GP5+/GP6+ primer systems. J Clin Microbiol, 1997, 35: 1304―1310
18 Cheah P L, Looi L M. Biology and pathological associations of the human papillomaviruses: A review. Malays J Pathol, 1998, 20: 1―10
19 Sichero L, Villa L L. Epidemiological and functional implications of molecular variants of human papillomavirus. Braz J Med Biol Res, 2006, 39: 707―717
20 Yamada T, Wheeler C M, Halpern A L, et al. Human papillomavirus type 16 variant lineages in United States populations characterized by nucleotide sequence analysis of the E6, L2, and L1 coding segments. J Virol, 1995, 69: 7743―7795
21 Chen Z, Terai M, Fu L, et al. Diversifying selection in human papillomavirus type 16 lineages based on complete genome analyses. J Virol, 2005, 79: 7014―7036
22 Ho L, Chan S Y, Burk R D, et al. The genetic drift of human papillomavirus type 16 is a means of reconstructing prehistoric viral spread and the movement of ancient human populations. J Virol, 1993,
67: 6413―6435 23 Ho L, Chan S Y, Chow V, et al. Sequence variants of human
papillomavirus type 16 in clinical samples permit verification and extension of epidemiological studies and construction of a phylogenetic tree. J Clin Microbiol, 1991, 29: 1765―1836
24 Kirii Y, Iwamoto S, Matsukura T. Human papillomavirus type 58 DNA sequence. Virology, 1991,185: 424―430
25 Chan P K, Chang A R, Cheung J L, et al. Determinants of cervical human papillomavirus infection: Differences between high- and low-oncogenic risk types. J Infect Dis, 2002, 185: 28―35
26 Chan P K, Chang A R, Tam W H, et al. Prevalence and genotype distribution of cervical human papillomavirus infection: Comparison between pregnant women and non-pregnant controls. J Med Virol, 2002, 67: 583—590
27 Xi L F, Touré P, Critchlow C W, et al. Prevalence of specific types of human papillomavirus and cervical squamous intraepithelial lesions in consecutive, previously unscreened, West-African women over 35 years of age. Int J Cancer, 2003, 103: 803―811
28 Xin C Y, Matsumoto K, Yoshikawa H, et al. Analysis of E6 variants of human papillomavirus type 33, 52 and 58 in Japanese women with cervical intraepithelial neoplasia/cervical cancer in relation to their oncogenic potential. Cancer Lett, 2001, 170: 19―24
29 Cho N H, An H J, Jeong J K, et al. Genotyping of 22 human papillomavirus types by DNA chip in Korean women: comparison with cytologic diagnosis. Am J Obstet Gynecol, 2003, 188: 56―62
30 Bernard H U, Calleja-Macias I E, Dunn S T. Genome variation of human papillomavirus types: Phylogenetic and medical implications. Int J Cancer, 2006, 118: 1071―1076
31 DeFilippis V R, Ayala F J, Villarreal L P. Evidence of diversifying selection in human papillomavirus type 16 E6 but not E7 oncogenes. J Mol Evol, 2002, 55: 491―499
32 Villa L L, Costa R L, Petta C A , et al. Prophylactic quadrivalent human papillomavirus (types 6, 11 16 and 18) L1 virus-like particle vaccine in young women a randomized double-blind placebo- controlled multicentre phase efficacy trial. Lancet Oncol, 2005, 6: Ⅱ
271―278 33 Harper D M, Franco E L, Wheeler C, et al. Efficacy of a bivalent L1
virus-like particle vaccine in prevention of infection with human papillomavirus types 16 and 18 in young women: A randomized controlled trial. Lancet, 2004, 364: 1757―1765