2007 The Paleontological Society. All rights reserved. 0094-8373/07/3302-0001/$1.00 Paleobiology, 33(2), 2007, pp. 165–181 MATTERS OF THE RECORD The problem with the Paleozoic Shanan E. Peters Shanan E. Peters. Department of Geological Sciences and Museum of Paleontology, University of Michigan, Ann Arbor, Michigan 48109. E-mail: [email protected]Accepted: 13 December 2006 Unfossiliferous marine sedimentary rocks of Phanerozoic age are known to all field-oriented paleontologists. These troublesome units are of- ten encountered in the field, perhaps cursed roundly for a moment or two, and usually shrugged off in pursuit of the next fossiliferous interval. Paleontologists tend not to discuss bar- ren units, and they rarely publish on the absence of a fauna from what appears to be unaltered marine rock. But aren’t barren marine sedi- ments revealing something important about their paleoenvironment and possibly about the paleoenvironments of conformably adjacent fos- sil-bearing units? Shouldn’t paleontologists be just as interested in knowing the locations and ages of unfossiliferous sediments as they are fossiliferous strata? In this essay, I discuss the possible environ- mental significance of barren marine sedi- ment and present preliminary data to suggest that its frequency may have varied consider- ably during the Phanerozoic. I also present ev- idence to suggest that this variation is associ- ated with shifts in the prevailing location of preserved marine sediment accumulation from expansive epicontinental seas to com- paratively narrow continental shelves. Finally, I propose that such shifts in the physiographic contexts of marine shelf habitats and their pre- served sediments are associated with ocean- ographic effects that have greatly influenced environmental and biological evolution, the character of the fossil record, and our percep- tion of marine biodiversity and macroevolu- tion at many different temporal and spatial scales. Nobody Home: Causes In the short time that I knew Jack Sepkoski, he told me at least twice about all of his fa- vorite Cambrian formations. He was particu- larly fond of puzzling over those unaltered marine sedimentary rocks that contained no, or exceedingly few, body fossils. The absence of skeletal material might, he reasoned, be the result of taphonomic or diagenetic loss. After all, it is widely appreciated that dissolution can substantially modify the diversity and compo- sition of fossil assemblages (e.g., Cherns and Wright 2000). The wholesale removal of skel- etal material was not, however, a satisfying ex- planation for many unfossiliferous units be- cause, even in the most extreme taphonomic and diagenetic settings, skeletonized animals are generally expected to leave at least some remnants behind. Although Sepkoski would never completely dismiss the possibility that dissolution was important, and for good rea- son (e.g., Reaves 1986), he would always con- clude that ‘‘no, it’s much more likely that there’s just nobody home.’’ Sepkoski’s quip about ‘‘nobody home’’ probably rings true for the majority of barren marine sediments. Although pre-compaction carbonate dissolution can remove all traces of calcareous skeletal material (e.g., Reaves 1986; Schovsbo 2001), lithology, sedimentary struc- tures, stratigraphic context, and occasional characteristic body fossils (Byers 1977; Gaines and Droser 2003) or ichnofossils (Byers 1977; Savrda and Bottjer 1986; Fo ¨llmi and Grimm 1990) often betray the fact that unfossiliferous sediments accumulated in environments that were inhospitable to most metazoans. What environmental factors might prevent the suc- cessful recruitment of skeletonized benthic animals for prolonged periods of time and therefore promote the accumulation of barren marine sediments?
Transcript
! 2007 The Paleontological Society. All rights reserved. 0094-8373/07/3302-0001/$1.00
Paleobiology, 33(2), 2007, pp. 165–181
MATTERS OF THE RECORD
The problem with the Paleozoic
Shanan E. Peters
Shanan E. Peters. Department of Geological Sciences and Museum of Paleontology, University ofMichigan, Ann Arbor, Michigan 48109. E-mail: [email protected]
Accepted: 13 December 2006
Unfossiliferous marine sedimentary rocks ofPhanerozoic age are known to all field-orientedpaleontologists. These troublesome units are of-ten encountered in the field, perhaps cursedroundly for a moment or two, and usuallyshrugged off in pursuit of the next fossiliferousinterval. Paleontologists tend not to discuss bar-ren units, and they rarely publish on the absenceof a fauna from what appears to be unalteredmarine rock. But aren’t barren marine sedi-ments revealing something important abouttheir paleoenvironment and possibly about thepaleoenvironments of conformably adjacent fos-sil-bearing units? Shouldn’t paleontologists bejust as interested in knowing the locations andages of unfossiliferous sediments as they arefossiliferous strata?
In this essay, I discuss the possible environ-mental significance of barren marine sedi-ment and present preliminary data to suggestthat its frequency may have varied consider-ably during the Phanerozoic. I also present ev-idence to suggest that this variation is associ-ated with shifts in the prevailing location ofpreserved marine sediment accumulationfrom expansive epicontinental seas to com-paratively narrow continental shelves. Finally,I propose that such shifts in the physiographiccontexts of marine shelf habitats and their pre-served sediments are associated with ocean-ographic effects that have greatly influencedenvironmental and biological evolution, thecharacter of the fossil record, and our percep-tion of marine biodiversity and macroevolu-tion at many different temporal and spatialscales.
Nobody Home: CausesIn the short time that I knew Jack Sepkoski,
he told me at least twice about all of his fa-
vorite Cambrian formations. He was particu-larly fond of puzzling over those unalteredmarine sedimentary rocks that contained no,or exceedingly few, body fossils. The absenceof skeletal material might, he reasoned, be theresult of taphonomic or diagenetic loss. Afterall, it is widely appreciated that dissolution cansubstantially modify the diversity and compo-sition of fossil assemblages (e.g., Cherns andWright 2000). The wholesale removal of skel-etal material was not, however, a satisfying ex-planation for many unfossiliferous units be-cause, even in the most extreme taphonomicand diagenetic settings, skeletonized animalsare generally expected to leave at least someremnants behind. Although Sepkoski wouldnever completely dismiss the possibility thatdissolution was important, and for good rea-son (e.g., Reaves 1986), he would always con-clude that ‘‘no, it’s much more likely thatthere’s just nobody home.’’
Sepkoski’s quip about ‘‘nobody home’’probably rings true for the majority of barrenmarine sediments. Although pre-compactioncarbonate dissolution can remove all traces ofcalcareous skeletal material (e.g., Reaves 1986;Schovsbo 2001), lithology, sedimentary struc-tures, stratigraphic context, and occasionalcharacteristic body fossils (Byers 1977; Gainesand Droser 2003) or ichnofossils (Byers 1977;Savrda and Bottjer 1986; Follmi and Grimm1990) often betray the fact that unfossiliferoussediments accumulated in environments thatwere inhospitable to most metazoans. Whatenvironmental factors might prevent the suc-cessful recruitment of skeletonized benthicanimals for prolonged periods of time andtherefore promote the accumulation of barrenmarine sediments?
The most geographically widespread envi-ronmental factor is probably oxygen depletion(Rhoads and Morse 1971; Byers 1977; Levin2003). Many benthic marine animals haveunique physiological or behavioral adapta-tions to cope with low oxygen concentrations(e.g., Diaz and Rosenberg 1995; Levin 2003),but intense or prolonged hypoxia can none-theless result in the complete extermination ofbenthic metazoans and the spread of persis-tent ‘‘dead zones’’ on the ocean floor (e.g.,Nordberg et al. 2001; Rabalais et al. 2002; Hel-ly and Levin 2004). Because hypoxia is pro-moted by biologically mediated oxidation oforganic matter in the water column, it is mostprevalent in bottom waters located below thewind-mixed surface layer in regions of highproductivity (Levin 2003) and in restricted ba-sins with poor circulation and stratified watercolumns (Rhoads and Morse 1971; Byers 1977;Kamykowski and Zentara 1990; Tyson andPearson 1991a; Wignall 1994).
Although humans are often implicated inmany cases of marine hypoxia (Levin 2003),there are presently over 106 km2 of shelves thatare naturally hypoxic owing to incursions ofpermanent oceanic thermoclines and their as-sociated oxygen minimum zones (OMZs) aswell as to precipitation and runoff-induced sa-linity stratification (Helly and Levin 2004).Thus, despite the important role that humansplay in promoting hypoxia (Diaz and Rosen-berg 1995; Diaz 2001), large areas would ex-perience episodic hypoxia even in the absenceof humans (Diaz and Rosenberg 1995; Diaz2001; Rabalais et al. 2002; Sorokin 2002; Levin2003; Helly and Levin 2004; Osterman et al.2005).
Low dissolved oxygen concentration isprobably the most geographically widespreadfactor that can result in the accumulation ofsignificant quantities of barren marine sedi-ment, but unusual salinity can also excludemetazoans for extended periods of time. Un-like hypoxia, hypersalinity typically occurs inwell-mixed, shallow-water settings, such asrimmed carbonate platforms and arid margin-al marine environments. Brackish water with-in estuaries and other humid, marginal ma-rine embayments may also contain a relativelydepauperate benthic fauna, but because many
taxa are uniquely adapted to variably brackishconditions (e.g., Fursich 1993), brackish wateralone is usually insufficient to exclude benthicorganisms for long periods of time.
Water-mass properties, such as oxygen con-centration and, to a lesser degree, salinity, areimportant factors that control the abundanceand diversity of benthic animals, but if sedi-mentation rates are high enough to dilute skel-etal input or if sedimentary processes them-selves preclude successful benthic recruit-ment, then unfossiliferous sediment may alsoaccumulate under fully normal marine con-ditions. Depositional settings with the neces-sary subsidence and sedimentation rates toexclude benthic organisms typically are foundonly in regions of active tectonic faulting, suchas the pull-apart basins in southern California(e.g., Kidwell 1988), and in pro-deltaic settingsin foreland basins.
All three proximal causes of barren marinesediment in the rock record, namely hypoxia,salinity, and sedimentation, are directly con-trolled by circulation patterns on and adjacentto shelves, precipitation patterns over the shelfand adjacent landmasses, and temperature.These parameters, in turn, are controlled byclimate and shelf physiography (Fig. 1). Be-cause hypoxia can be promoted by many dif-ferent factors, it is likely to be a prevalent, butby no means exclusive, cause of barren marinesediment in the geologic record. It is of coursealso likely that several factors might combineso as to effectively exclude benthic organismseven when any one factor by itself would beinsufficient to prevent the successful recruit-ment of skeletonized benthos.
Nobody Home: Temporal Patterns
‘‘Paleozoic black shales indicate by their con-tained faunas and lithologic associations the prev-alence of somewhat abnormal, toxic bottom condi-tions that either impoverished or completely pre-vented all bottom life . . . over vast areas andthroughout long intervals of time’’ Ruedemann(1935: p. 90).
Ruedemann’s (1935) observations character-ize a few familiar manifestations of the oxy-gen-deficient bottom-water conditions thatcommonly result in the accumulation of thick,
FIGURE 1. Causes of barren marine sediment (excluding dissolution). Arrows indicate direction of forcing, but notmagnitude or nature of interaction. Shelf physiography subsumes tectonic context of shelves, bathymetric profiles,geometry, geographic position, and characteristics of adjacent oceanic water masses and currents. Climate includesboth global and local effects. Circulation includes exchange of water between the shelf and open ocean as well aswind-driven mixing. Oval boxes list some of the general signal types that may be unique to each proximal causeof barren sediment. Strat., stratification of the water column; O2 sol., oxygen solubility.
organic-rich, unfossiliferous sedimentary suc-cessions (Rhoads and Morse 1971; Byers 1977).Two aspects of Ruedemann’s comments areparticularly noteworthy. First, he specificallyhighlights the Paleozoic as a time that is char-acterized by such deposits. Second, he notesthat ‘‘toxic bottom conditions’’ are commonlywidespread and geologically long lived. Al-though Ruedemann’s perceptions must havebeen colored by his work on graptolites fromthe well-known Paleozoic mudstones of theAppalachian foreland basin, his commentarymakes a testable prediction regarding the fre-quency of unfossiliferous marine sedimentover geologic time.
To indirectly evaluate the possibility that‘‘toxic bottom conditions,’’ as evidenced byunfossiliferous marine sediment, may havedecreased in frequency during the Phanero-zoic, I searched the Georef bibliographic indexfirst for the term ‘‘unfossiliferous’’ and thenfor the term ‘‘fossiliferous’’ occurring any-where in a citation (searches conducted in
March 2006). I then read the abstracts of therecovered references (extending only back to1957) to determine whether the referencedsediments were marine and to determine theirgeologic age. If age was given to period andthe unit was determined to be marine in ori-gin (either explicitly or by inference), one unitwas scored for all of the relevant time periodslisted in the reference.
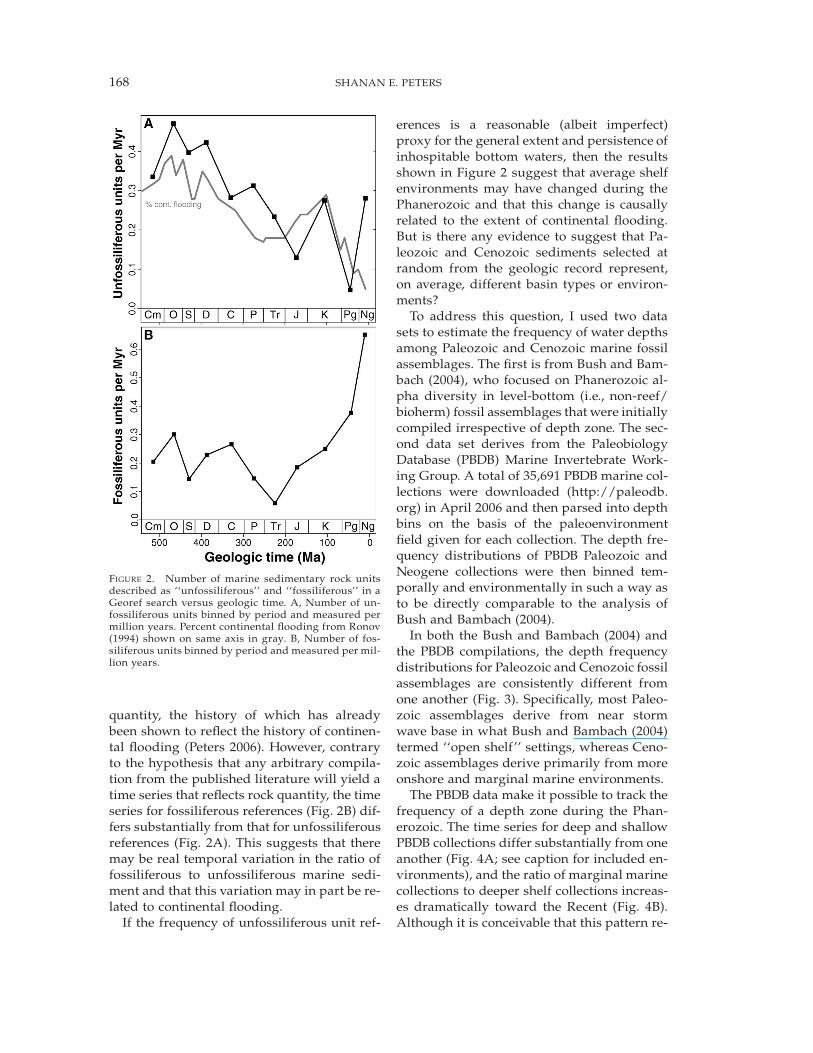
According to the Georef search, there is adecline in the frequency of unfossiliferousunits from a Phanerozoic high in the Ordo-vician to a low in the Paleogene (Fig. 2A). Thisdecline is broadly similar to Phanerozoic sealevel estimates (Miller et al. 2005) and the his-tory of continental flooding (Fig. 2A, grayline) (Ronov 1994). Although the time series ofunfossiliferous sediment (Fig. 2A) suggeststhat there may have been significant temporalchanges in the prevalence of environmentalconditions favoring the accumulation of bar-ren marine sediment, it is also possible thatthe pattern is controlled by preserved rock
FIGURE 2. Number of marine sedimentary rock unitsdescribed as ‘‘unfossiliferous’’ and ‘‘fossiliferous’’ in aGeoref search versus geologic time. A, Number of un-fossiliferous units binned by period and measured permillion years. Percent continental flooding from Ronov(1994) shown on same axis in gray. B, Number of fos-siliferous units binned by period and measured per mil-lion years.
quantity, the history of which has alreadybeen shown to reflect the history of continen-tal flooding (Peters 2006). However, contraryto the hypothesis that any arbitrary compila-tion from the published literature will yield atime series that reflects rock quantity, the timeseries for fossiliferous references (Fig. 2B) dif-fers substantially from that for unfossiliferousreferences (Fig. 2A). This suggests that theremay be real temporal variation in the ratio offossiliferous to unfossiliferous marine sedi-ment and that this variation may in part be re-lated to continental flooding.
If the frequency of unfossiliferous unit ref-
erences is a reasonable (albeit imperfect)proxy for the general extent and persistence ofinhospitable bottom waters, then the resultsshown in Figure 2 suggest that average shelfenvironments may have changed during thePhanerozoic and that this change is causallyrelated to the extent of continental flooding.But is there any evidence to suggest that Pa-leozoic and Cenozoic sediments selected atrandom from the geologic record represent,on average, different basin types or environ-ments?
To address this question, I used two datasets to estimate the frequency of water depthsamong Paleozoic and Cenozoic marine fossilassemblages. The first is from Bush and Bam-bach (2004), who focused on Phanerozoic al-pha diversity in level-bottom (i.e., non-reef/bioherm) fossil assemblages that were initiallycompiled irrespective of depth zone. The sec-ond data set derives from the PaleobiologyDatabase (PBDB) Marine Invertebrate Work-ing Group. A total of 35,691 PBDB marine col-lections were downloaded (http://paleodb.org) in April 2006 and then parsed into depthbins on the basis of the paleoenvironmentfield given for each collection. The depth fre-quency distributions of PBDB Paleozoic andNeogene collections were then binned tem-porally and environmentally in such a way asto be directly comparable to the analysis ofBush and Bambach (2004).
In both the Bush and Bambach (2004) andthe PBDB compilations, the depth frequencydistributions for Paleozoic and Cenozoic fossilassemblages are consistently different fromone another (Fig. 3). Specifically, most Paleo-zoic assemblages derive from near stormwave base in what Bush and Bambach (2004)termed ‘‘open shelf’’ settings, whereas Ceno-zoic assemblages derive primarily from moreonshore and marginal marine environments.
The PBDB data make it possible to track thefrequency of a depth zone during the Phan-erozoic. The time series for deep and shallowPBDB collections differ substantially from oneanother (Fig. 4A; see caption for included en-vironments), and the ratio of marginal marinecollections to deeper shelf collections increas-es dramatically toward the Recent (Fig. 4B).Although it is conceivable that this pattern re-
FIGURE 3. Total number of Paleozoic (Pz, Ordovician–Devonian, solid line) and Neogene (Ng, dashed line) fos-sil assemblages sampled from different depth zones. A,Assemblage data from Bush and Bambach (2004). Depthzones given by Bush and Bambach (2004) are (e1) shore-lines, (e2) nearshore shelf, (e3) open shelf, (e4) distalopen shelf, (e5) outer shelf margin. Estimated positionof average storm wave base (swb) is shown by dashedline. B, Percent collections in the PBDB Marine Inverte-brate Working Group for the Pz (4742 total collections)and Ng (1198 total collections). Environmental bin e1corresponds to PBDB marginal marine, coastal, andforeshore collections, e2 corresponds to PBDB shallowsubtidal and shoreface collections, e3 corresponds toPBDB deep subtidal and lower shoreface collections, e4corresponds to PBDB offshore collections, and e5 cor-responds to PBDB slope and basinal collections.
FIGURE 4. Number of Paleobiology Database (PBDB)collections from deep water and marginal marine en-vironments versus geologic time. A, Absolute time se-ries. Solid line labeled ‘‘deep’’ includes collections iden-tified as deep subtidal, transition zone/lower shoreface,offshore, basinal, slope, and submarine fan. Dashed linelabeled ‘‘marginal’’ includes collections designated ascoastal, marginal marine, estuary/bay, paralic, lagoon-al, peritidal, and foreshore. Compare the deep curve tothe curve for continental flooding (Fig. 2A) and comparethe marginal curve to the number of fossiliferous Georefreferences (Fig. 2B). B, Ratio of marginal to deep PDBDcollections over time.
flects systematic environmental error in thePBDB, the time series for the number of deepshelf collections mirrors global sea level (Mil-ler et al. 2005) and the number of unfossilif-erous unit references (Fig. 2A), and the timeseries for the number of marginal marine col-lections resembles the number of fossiliferousunit references (Fig. 1B) and the history of ma-rine genus richness (Sepkoski 2002; curve notshown).
What might account for the observed dif-
ferences in the depth frequency distributionsof Cenozoic and Paleozoic fossil assemblages(Fig. 3) as well as for different temporal pat-terns in the frequency of marginal and deepershelf collections (Fig. 4)? I argue here that themost likely explanation is a shift in the aver-age location of deposition of preserved marinesediment from broad epicontinental seas inthe Paleozoic to narrow continental margins
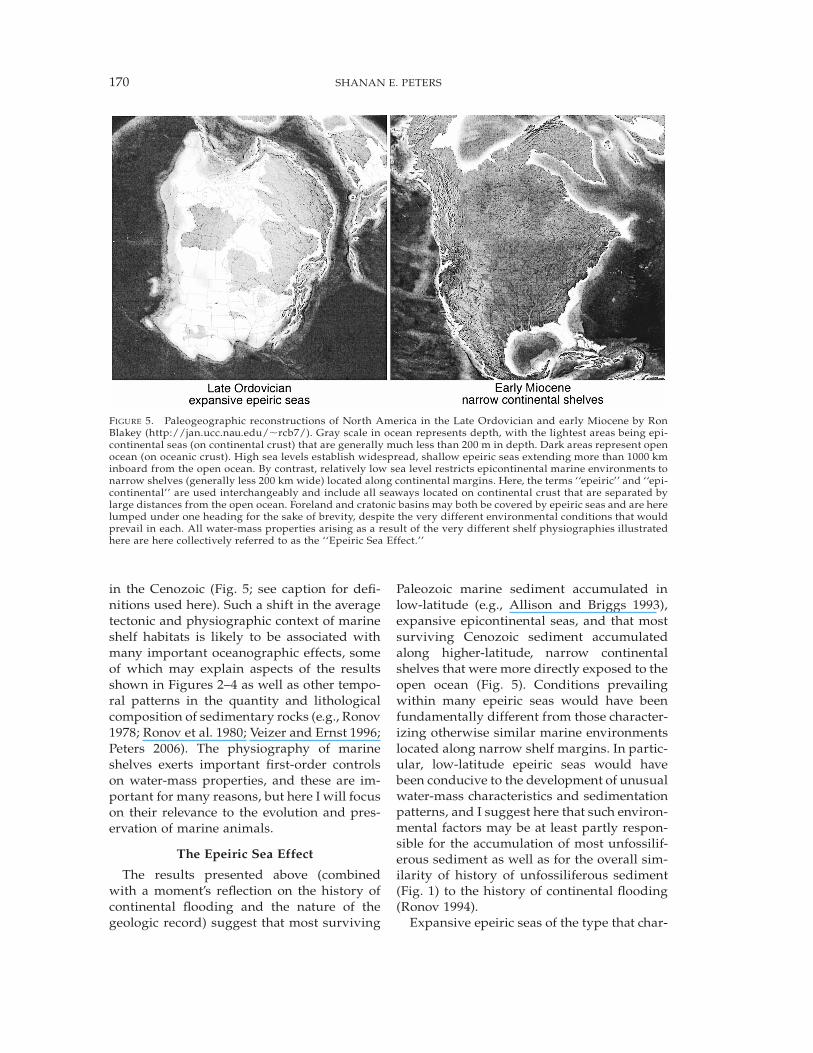
FIGURE 5. Paleogeographic reconstructions of North America in the Late Ordovician and early Miocene by RonBlakey (http://jan.ucc.nau.edu/!rcb7/). Gray scale in ocean represents depth, with the lightest areas being epi-continental seas (on continental crust) that are generally much less than 200 m in depth. Dark areas represent openocean (on oceanic crust). High sea levels establish widespread, shallow epeiric seas extending more than 1000 kminboard from the open ocean. By contrast, relatively low sea level restricts epicontinental marine environments tonarrow shelves (generally less 200 km wide) located along continental margins. Here, the terms ‘‘epeiric’’ and ‘‘epi-continental’’ are used interchangeably and include all seaways located on continental crust that are separated bylarge distances from the open ocean. Foreland and cratonic basins may both be covered by epeiric seas and are herelumped under one heading for the sake of brevity, despite the very different environmental conditions that wouldprevail in each. All water-mass properties arising as a result of the very different shelf physiographies illustratedhere are here collectively referred to as the ‘‘Epeiric Sea Effect.’’
in the Cenozoic (Fig. 5; see caption for defi-nitions used here). Such a shift in the averagetectonic and physiographic context of marineshelf habitats is likely to be associated withmany important oceanographic effects, someof which may explain aspects of the resultsshown in Figures 2–4 as well as other tempo-ral patterns in the quantity and lithologicalcomposition of sedimentary rocks (e.g., Ronov1978; Ronov et al. 1980; Veizer and Ernst 1996;Peters 2006). The physiography of marineshelves exerts important first-order controlson water-mass properties, and these are im-portant for many reasons, but here I will focuson their relevance to the evolution and pres-ervation of marine animals.
The Epeiric Sea Effect
The results presented above (combinedwith a moment’s reflection on the history ofcontinental flooding and the nature of thegeologic record) suggest that most surviving
Paleozoic marine sediment accumulated inlow-latitude (e.g., Allison and Briggs 1993),expansive epicontinental seas, and that mostsurviving Cenozoic sediment accumulatedalong higher-latitude, narrow continentalshelves that were more directly exposed to theopen ocean (Fig. 5). Conditions prevailingwithin many epeiric seas would have beenfundamentally different from those character-izing otherwise similar marine environmentslocated along narrow shelf margins. In partic-ular, low-latitude epeiric seas would havebeen conducive to the development of unusualwater-mass characteristics and sedimentationpatterns, and I suggest here that such environ-mental factors may be at least partly respon-sible for the accumulation of most unfossilif-erous sediment as well as for the overall sim-ilarity of history of unfossiliferous sediment(Fig. 1) to the history of continental flooding(Ronov 1994).
acterized much of the Paleozoic and the Juras-sic–Cretaceous have no close modern ana-logues, and few oceanographic studies havesystematically explored the expected water-mass properties of epeiric seas. Nevertheless,several authors have pointed out the oceano-graphic differences between narrow continen-tal shelves and epicontinental seaways, andsome of the most consistent and importantdifferences involve the expected frequencyand extent of bottom-water hypoxia. For ex-ample, Tyson and Pearson (1991a) summarizethe causes of hypoxia on modern shelves, ar-gue that most epeiric seas must have been sub-ject to episodic hypoxia, and offer an actualis-tic model for its development in epeiric seas.Allison and Wright (2005) and Algeo et al.(2007) emphasize the non-actualistic aspectsof epeiric seas and offer what the latter studycalls a ‘‘superesturarine model’’ for the devel-opment of widespread salinity anomalies andconcomitant hypoxia in thin bottom-waterlayers. Some of the basic features of thesemodels that are most relevant to paleobiology,along with a few modifications and additions,are briefly outlined below.
Hypoxia in Epeiric Seas. The frequency andintensity of hypoxia in any epeiric sea wouldhave been controlled by boundary conditionsspecific to the physiography of the seaway andadjacent water masses as well as to local cli-mate, just as it is on modern shelves (Algeo etal. 2007). However, two factors are consistent-ly important on long time scales: (1) sea levelrises that establish widespread seaways withwater depths capable of supporting pycno-clines on at least a temporary basis, and (2) cy-clicity on annual and Milankovitch time-bands in temperature, productivity, freshwa-ter input, and storm-driven mixing of the wa-ter column. Tidal mixing may also serve as animportant deterrent of stratification on manycontinental shelves (Algeo et al. 2007), but tid-al amplitudes were probably negligible inmost large epeiric seas (e.g., Allison andWright 2005; Wells et al. 2005).
The combination of, and interactions be-tween, sea level changes and climate likely re-sulted in at least seasonal development of apycnocline and concomitant hypoxia in thinbottom-water layers over vast areas in many
epeiric seas, particularly those located at lowlatitudes. Because wind-driven mixing is crit-ical to preventing stratification of the watercolumn and therefore the development of hyp-oxia in shallow ("200 m) water settings, theneven if Milankovitch forcing resulted in nosubstantial changes in sea level, its influenceon storm frequency or intensity could have anoceanographic effect in offshore settings thatis equivalent to sea level change. Seasonalityand climatic variations would have been par-ticularly effective promoters of hypoxia inepeiric seas located at low latitudes, such asthose that dominated during much of the Pa-leozoic.
Hypoxia would tend to develop initially inthe deepest basins, some of which could po-tentially have permanent thermoclines andtherefore permanently anoxic bottom waters,and then spread outward across the shelf inresponse to sea level rise or changes in mixingparameters. This is comparable to what Wig-nall (1994) called the ‘‘expanding puddlemodel’’ because of the potentially sharp, wa-terline-like oxycline that would intersect theseafloor, possibly filling subtle seafloor topog-raphy and contributing to strong faunal patch-iness both regionally and locally (e.g., Brett1999).
It is obvious, but important to point out,that oxygen concentration varies continuouslyfrom anoxic to fully oxic, and that hypoxia inepeiric seas must also have varied in intensity,frequency, duration, and distribution depend-ing upon a large number of boundary condi-tions, including circulation patterns, waterdepths, nutrient supplies, storm intensities,freshwater inputs, mean annual temperature(e.g., Keeling and Garcia 2002), and even theconcentration of oxygen in the atmosphere(e.g., Berner et al. 2003). Barren, laminated‘‘black shales’’ represent but one end-memberin a continuum of bottom-water oxygen con-ditions, and the absence of a stereotypic ‘‘an-oxic’’ facies cannot be interpreted as evidencefor the absence of episodic hypoxia. This isparticularly true because all sedimentological,geochemical, and biological signatures of briefbouts of hypoxia could be overprinted duringintervals of normal oxygen concentration.
hypoxia, modulated by longer-wavelength cy-clicity in water depth or storm intensity, pro-ductivity, and temperature could have result-ed in the widespread mortality of benthic ma-rine animals on an annual or longer-term ba-sis over vast areas in many epeiric seas. Therecovery of benthic communities from thesehypoxic episodes would have been difficultand much slower than their initial extermi-nation (e.g., Stachowitsch 1991). Moreover, inepeiric seas that experienced frequent hypox-ia, only opportunistic, rapidly dispersing taxawith short life cycles would have been able tocolonize the seafloor during temporary im-provements, possibly resulting in the preser-vation of single, nearly monospecific, settle-ment cohorts within otherwise mostly barrensediment. Slowly growing, longer-lived taxawould generally have been excluded fromsuch environments altogether. More persis-tently hypoxic environments would have pro-moted specialization and morphological/physiological adaptation, and this has beenobserved in the fossil record (e.g., Savrda andBottjer 1987; Gaines and Droser 2003; Hen-derson 2004).
Rhodes and Morse (1977), Tyson and Pear-son (1991a), Wignall (1994), Levin (2003), andothers provide comprehensive summaries ofthe many dramatic effects that hypoxia canhave on the behavior, diversity, composition,and abundance of benthic organisms, and I re-fer the reader to these excellent sources formore information. It is worth noting, however,that most of these references focus on the bi-otic characteristics of persistently hypoxic oreven anoxic environments, such as those lo-cated beneath permanent OMZs and recordedby black shales. However, because hypoxia islikely to have been transient over wide areasin many epeiric seas, the presence of rather di-verse and abundant stenotopic benthic organ-isms does not necessarily preclude the possi-bility that a marine community was influ-enced by episodic hypoxia. Brief episodesbrought about by unusually low storm fre-quency/intensity (low mixing), peaks in pro-ductivity (high oxygen consumption), orfreshwater pulses (enhanced stratification)could have resulted in the wholesale or selec-tive extermination of benthic animals over
vast areas of seafloor. Thus, even in sedimentsthat yield abundant and well-preserved ma-rine animals, hypoxia may have resulted inthe episodic extermination and reestablish-ment of entire benthic communities (e.g.,Boesch and Rabalais 1991).
A Possible Example of Hypoxia in an Epeiric SeaSetting. The richly fossiliferous CincinnatianSeries of the Cincinnati Arch region accumu-lated in an epeiric sea that is typically thoughtof as being ‘‘normal marine’’ in character.However, much of this thick succession ofmudstone, siltstone, and carbonate accumu-lated in a terrigenous sediment-catchingepeiric sea separated by hundreds of kilome-ters from the open ocean. Episodic hypoxiamay have been common in this depositionalsetting, and I suggest that some features of thestratigraphy and taphonomy of the type Cin-cinnatian Series may have been influenced bycyclical variation in the frequency and inten-sity of widespread hypoxia, particularly inoffshore sediments that contain well-pre-served fossil assemblages.
One of the best-known examples of thespectacular preservation that can characterizeoffshore and deep subtidal deposits withinthe type Cincinnatian are articulated speci-mens of Flexicalymene. This medium-sized (upto about 7 cm) calymenid trilobite is oftenfound in dense aggregations within blue-graymudstones (‘‘butter shales’’; Hunda et al.2006), some of which can be traced for tens ofkilometers (Brett et al. 2007). Many of the in-dividuals within these aggregations are whol-ly articulated (including in-place hypostomes)and display numerous postures, including en-rollment (Fig. 6). One common interpretationis that these assemblages formed during thedeposition of distal tempestites that rapidlysmothered trilobites in life and escape posi-tions (e.g., Hughes and Cooper 1999). How-ever, articulated Flexicalymene individuals areoften distributed throughout 40–100 cm thickmudstone intervals that contain subtle centi-meter-scale and finer bedding features, andthere appears to be no consistent associationbetween articulated trilobites and storm-gen-erated sedimentary features (Schumacher andShrake 1997; Hunda et al. 2006).
FIGURE 6. Flexicalymene aggregation of four articulated individuals preserved in pyrite-bearing gray mudstonefrom the type Cincinnatian Series in Ohio. It is here hypothesized that episodic bouts of hypoxia resulted in thedeath of trilobites and other benthic animals in many epicontinental settings. Subsequent burial by modest quan-tities of sediment preserved the dead animals in fantastic death poses (shown here), modified death poses, or asscattered carcasses in various states of decay and disarticulation. See text for further explanation and for discussionof similar hypoxia-related enrollment among modern horseshoe crabs (Fisher 1977). Individual specimen at lowerleft side of photo has been removed from the matrix and turned upside down in order to reveal the enrolled spec-imen located directly beneath the partially prone individual. Specimen and photo courtesy of Kenneth D. Karns.
could be consistently linked to centimeter-scale graded silt and mud beds, it seems un-likely that an entire community of trilobiteswith lengths of 5 cm or more could be ren-dered helpless by burial in what must havebeen rather modest amounts of suspendedfine-grained sediment (generally much lessthan 10 cm, even after correction for compac-tion). Moreover, fully articulated specimens ofIsotelus, an asaphid trilobite reaching over 30cm in length, also occur in enrolled and flatpostures within similar muddy intervals(Schumacher and Shrake 1997). It is unlikelythat these giants were overly sensitive to buri-al by the modest quantities of sediment thatare often suggested by detailed microstrati-graphic studies (Schumacher and Shrake 1997;Hunda et al. 2006), particularly because sometrilobites, including calymenids and asaphids,are known to be capable burrowers (e.g., Os-good 1970; Cherns et al. 2006).
In many cases of trilobite Konservat-Lager-statte, such as the Cincinnatian trilobite beds,enrolled specimens are common. For example,Hunda et al. (2006) reported an enrollmentfrequency of 42% among Flexicalymene in a sin-gle mudstone interval (56% if partial enroll-ment is included). Although enrollment is of-
ten interpreted as a defensive posture as-sumed in response to the physical burial pro-cess, Fisher (1977, personal communication2006) has shown that small horseshoe crabscan easily escape when buried by at least sev-eral body lengths of fine sediment. When sub-ject to hypoxia, however, these same horse-shoe crabs will enroll in a fashion that is sim-ilar to that of trilobites and remain so evenwhen shallowly buried. If hypoxia persists,the animals will die in an enrolled posture onthe surface or beneath a thin layer of sediment.Partly because of the experiments conductedby Fisher (1977), enrollment among some tri-lobites has been interpreted as a behavioral re-sponse to hypoxia by Fisher and subsequentlyby other authors (e.g., Speyer 1990). This hy-pothesis is supported by studies that haveidentified beds of enrolled trilobites as beingparticularly prevalent in pyritic mudstone fa-cies indicative of hypoxia (Speyer and Brett1986).
In the hypoxia model for trilobite Lager-statte (and other deposits that contain articu-lated multi-element skeletons buried beneathrelatively thin mud drapes), all animals thatsuccumbed to hypoxia would be lying undis-turbed (and not disturbing one another) in
death postures on the seafloor. The entire as-semblage could then be buried by one or morestorm-induced mud transport events, or itcould be left exposed, only to undergo even-tual decay, disarticulation, and dispersal byscavengers during the next normal oxygenphase or by currents during the next largestorm. Cyclic alternations between favorableand hypoxic conditions would result in the ac-cumulation of significant quantities of sedi-ment with comparable fine-scale sedimento-logic and taphonomic signatures. Under thismodel, there need be no consistent relation-ship between storm-induced sedimentarystructures and articulated or incipiently de-caying benthic organisms, but any wide-spread hypoxia-related death assemblage canbe preserved only if it is buried, and rapidburial beneath a silt/mud tempestite, trans-ported in suspension over wide areas alongsubmarine nepheloid layers (e.g., Kohrs et al.2007), is a very probable scenario.
Paleozoic Lagerstatte of all types should bereevaluated for ichnological, sedimentologi-cal, and taphonomic evidence of hypoxia asthe primary killing mechanism and later buri-al beneath thin tempestites as the entombingmechanism. Distinguishing between the ob-rution and hypoxia models may seem like asubtle distinction in taphonomic mode, but itis of great importance in understanding thewater-mass properties in which the preservedtaxa lived and evolved and for determiningthe potential influence and scope of the Epeir-ic Sea Effect in structuring benthic marinecommunities.
Stratigraphic Consequences of Hypoxia. Giv-en the possibility of widespread hypoxia inthe Cincinnati Arch region, it is interesting tohypothesize that meter-scale cyclicity in fau-nal composition within the Cincinnatian (e.g.,Holland et al. 2000; Miller et al. 2001; Webber2002), and other similar units with cyclicitymanifested by the abundance and composi-tion of skeletal material (e.g., Brett et al. 2006),might be influenced by the effects of cyclichypoxia as well as changes in bottom-currentenergy and the frequency of sediment trans-port events.
During times of more frequent or intensehypoxia, poorly fossiliferous sediments con-
taining only those taxa that were able to cope,or that had short generation times permittingthem to take advantage of brief intervals ofnormal oxygen concentration, would accu-mulate. Less frequent or less intense hypoxiawould, by contrast, permit the establishmentof communities with lower hypoxia tolerancesand would typically support a higher densityof individuals. In this situation, shelly lags orskeletal grainstones and packstones with agenerally different taxonomic compositionfrom interbedded, less fossiliferous sedimentswould accumulate (see Tsujita et al. 2006 foran example of this phenomenon in the Devo-nian Arkona Shale). The thicknesses of eachfacies (sparingly fossiliferous fine-grainedsediment versus higher carbonate content andmore shell beds) would be largely indicativeof the average amount of time spent in eachstate (frequent/intense hypoxia versus moreoxygenated conditions), and it would be pos-sible to have oscillations between end-mem-bers on the shortest time and thickness scales.
Interestingly, many of the same factors thatwould contribute to cyclicity in hypoxia, suchas Milankovitch forcing of climate (see above),would also influence wave energy at the sed-iment-water interface and it is therefore likelythat hypoxia would be acting in conjunctionwith bottom current energy to structure sed-imentologic and taxonomic signatures. Dur-ing periods of weaker storms, hypoxia wouldgenerally be more widespread and intenseowing to weaker mixing of the water column,and this would promote the accumulation ofless fossiliferous sedimentary intervals con-sisting of relatively fine-grained sediment. Pe-riods of shallower water or stronger stormswould be characterized by less frequent bot-tom-water hypoxia and would produce morefossiliferous sedimentary intervals, more win-nowed and amalgamated storm beds, andcoarser grain sizes. Disentangling the relativecontributions of hypoxia and wave energy atthe sediment-water interface may prove diffi-cult, but it should be possible with detailedmicrostratigraphic taphonomic, ichnologic,and paleoecological studies because hypoxiamakes many testable predictions at thesemuch finer stratigraphic scales (e.g., see Wig-nall 1994 and references therein).
Salinity in Epeiric Seas. Hypoxia is not theonly environmental stress that would havebeen widespread in many epeiric sea settings.Carbonate platforms in the Paleozoic, such asthe expansive Great American CarbonateBank (Cambro-Ordovician) and the continent-scale carbonate platforms and ramps of the Si-lurian, were maintained at or near sea level formillions of years (e.g., Ginsburg 1982). Thewater masses on these shallow platforms andramps were probably often restricted fromopen-ocean circulation by virtue of marginalbuildups or their sheer geographic scale. Insuch settings, high water temperatures, ele-vated salinity, and rapid salinity changes ow-ing to alternating high evaporation and rain-fall probably prevented all but algal and mi-crobial life from becoming established. It isprobably for this reason that many platformcarbonate successions contain barren, cryptal-gal laminated, shallow subtidal to peritidalcarbonates that are both devoid of benthic an-imal remains and similar to those found in sa-linity-stressed modern tropical carbonatebuildups (e.g., Fagerstrom and Weidlich 2005).
Although shallow carbonate platforms inthe Paleozoic were certainly capable of sup-porting diverse benthic communities, it is alsotrue that extensive carbonate platforms werefrequently subjected to anomalously high orfluctuating salinities. As with hypoxia, thesetwo end-members bracket a continuum in thefrequency and magnitude of episodicallystressful salinity conditions. At any particularpoint in space, normal and stressful condi-tions are likely to have alternated in time suchthat barren intervals come to be interbeddedwith fossiliferous strata. Changes in the ge-ometry of carbonate environments from plat-forms to more openly circulating ramps arealso likely to have resulted in important wa-ter-mass changes, and such changes have beenlinked to the abundance of crinoids in the fos-sil record (Kammer and Ausich 2006).
High salinity may have prevailed on manyexpansive carbonate platforms during the Pa-leozoic, but low salinity may have been aneven more pernicious problem in many epeiricseas. Allison and Wright (2005) argue for thedevelopment of extensive freshwater lenses inepeiric seas and describe the environmental
changes they promote. In particular, theypoint out that salinity stratification, in com-bination with weak tidal mixing, can result inshallow-water hypoxia. The ponding of brack-ish water within epeiric seas is therefore likelyto have imposed salinity stresses that both di-rectly and indirectly influenced the abun-dance and diversity of marine animals.
Sedimentation in Epeiric Seas. During timesof high continental flooding and expansiveepicontinental seas, most of the world’s sili-ciclastic sediment load would have been trans-ported by rivers onto shallow shelves span-ning hundreds of kilometers. Some of thissediment would have been trapped in rapidlysubsiding foreland basins, but much of itwould have been dispersed by storm waves(which form effective base level) over wide ar-eas. Suspended sediment, transported direct-ly from rivers or resuspended during storms,would contribute not only to productivity inthe water column and therefore to bottom-wa-ter hypoxia, but also to the near simultaneoussmothering of wide areas as storm-transport-ed sediment encountered and then spreadalong pycnoclines before finally settling.
As for hypoxia and salinity, it is likely thatexpansive epicontinental seas promoted dif-ferent sediment transport dynamics than oth-erwise similar narrow continental shelves,and that such dynamics are relevant to the dis-tribution and abundance of benthic organ-isms. The comparative sedimentology of nar-row shelves versus widespread epicontinentalseas needs to be explored more thoroughly be-fore evolutionary hypotheses can be adequate-ly formulated and then tested in the field.
The Cenozoic Contrast. In contrast to theepeiric sea-dominated Paleozoic, most Ceno-zoic marine sediments, and other Phanerozoictime intervals characterized by low levels ofcontinental flooding, represent ‘‘bathtub rings’’that were deposited on narrow continentalmargins with a comparatively small area ofdeeper shelf. Most of these narrow shelf en-vironments would have been more freely cir-culating with the open ocean and less prone(but by no means immune) to episodic strati-fication and concomitant hypoxia. Estuarineand other restricted marine environments arecertainly present in many Cenozoic sequenc-
es, and numerous sedimentary basins duringsea level lowstands could have been prone tounusual water-mass characteristics and strongstratification, but, on average, shelf environ-ments of the type that characterize the Ceno-zoic are quite unlike those that dominatedduring the Paleozoic. The oceanographic con-sequences of such critical changes in marineshelf physiography cannot go unrecognizedby ecological and evolutionary studies of thefossil record.
The Epeiric Sea Effect: MacroevolutionaryConsequences
The macroevolutionary consequences oftemporal shifts in the average character of ma-rine shelf habitats and their preserved sedi-ments (the Epeiric Sea Effect) are twofold.First, because these shifts reflect, at least inpart, real changes in the relative proportionsof fundamentally different types of marinehabitats, at least one important aspect of theenvironmental mosaic upon which marine an-imal evolution occurred must have changed.Second, because at any point in time benthicanimals would have been influenced by theEpeiric Sea Effect, temporal patterns derivedfrom the aggregate fossil record may not beevolutionary or ecological trends per se, butmay instead reflect differential environmentalsampling (i.e., empirical temporal patterns re-flect spatial patterns). It is also possible thatreal evolutionary or ecological patterns mightresult only from shifting relative proportionsof habitats in an otherwise constant environ-mental mosaic (see Rhoads and Morse 1977:Fig. 5 for an illustration of the basic idea).Here, I briefly discuss these issues and suggestapproaches to overcoming some of the poten-tial empirical problems imposed by the Epeir-ic Sea Effect.
Macroevolutionary Consequences. The poten-tial macroevolutionary consequences of theexpansion and contraction of epeiric seas andtheir unique shallow marine environmentshave been discussed for some 60 years (e.g.,Newell 1949, 1952; Valentine and Moores 1970;Johnson 1974; Schopf 1974; Simberloff 1974).Most of these discussions have focused onspecies-area effects and the possibility thatmass extinctions may have been induced by
the elimination of shallow marine environ-ments and associated ‘‘perched’’ endemic fau-nas. There is now good quantitative evidenceto suggest that such factors have indeed ex-ercised a first-order control on global biodi-versity and on rates of extinction and origi-nation during the Phanerozoic (Peters 2005),and that similar processes can affect the com-position and evolution of marine faunas onmillion-year time scales within individual de-positional basins (e.g., Bowersox 2005). How-ever, few studies have addressed the potentiallarge-scale macroevolutionary significance ofthe Epeiric Sea Effect in terms of changes inthe character of marine shelf habitats in gen-eral.
We have much to learn about the specific en-vironmental conditions that would have pre-vailed in most epeiric seas, but there is abun-dant evidence to suggest that hypoxia wouldhave been particularly widespread and fre-quent (Tyson and Pearson 1991a; Allison andWright 2005; Algeo et al. 2007). Because hyp-oxia probably occurred on short, even season-al, time scales in many epeiric seas, living ator above the sediment-water interface couldhave conferred a substantial survival advan-tage over being infaunal. In fact, at the onsetof hypoxia, modern infaunal stenoxic benthicorganisms extend from the sediment andeventually remove themselves completely inorder to reach upward into the water column,and mobile taxa will aggregate on topograph-ic highs or climb on top of one another to es-cape the benthic boundary layer (Tyson andPearson 1991a). The dominance of epibenthictaxa, shallow bioturbation, and vertical water-column tiering (Ausich and Bottjer 1982; Bott-jer and Ausich 1986) in many Paleozoic com-munities may therefore partially reflect thefact that elevation above the sediment-waterinterface is a very effective way for benthic an-imals to minimize the impacts of bottom-wa-ter hypoxia.
In support of this hypothesis, Brett (1984)and Frest et al. (1999) have noted that long-stemmed Devonian crinoids are particularlyabundant in dysoxic facies lacking surfacedwellers like brachiopods, bryozoans, andother shorter-stemmed crinoids, and thatlong-stemmed crinoids are not so abundant in
more oxygenated and crowded benthic com-munities where the traditional view of nichepartitioning would predict strong selectivepressure for vertical tiering. Because swim-ming predators often take advantage of the ef-fects that hypoxia has on benthic prey (e.g.,Pihl et al. 1992; Rabalais et al. 2002), an epi-faunal, vertically positioned life habit wouldhave been a particularly effective strategy todeal with frequent hypoxia before the evolu-tion of abundant durophagous predators(Signor and Brett 1984; Baumiller and Gahn2004).
Although widespread hypoxia in epeiricseas may have been stressful for many benthicanimals, it may also have afforded an impor-tant opportunity in the form of an effective, ifepisodically costly, means of escaping preda-tors. Benthic marine invertebrates that wereable to cope with hypoxia could have activelysought out these environments to aggregate induring vulnerable stages of their life history(e.g., reproduction and molting), to exploit lit-tle-used resource pools, or to escape predatorswith higher oxygen demands. ‘‘Oxygen refu-gia’’ have been demonstrated to reduce ratesof predation in some freshwater fish (e.g.,Chapman et al. 1995) and it is likely that ma-rine invertebrates use similar avoidance strat-egies, either actively or opportunistically(Levin 2003). Precarious tradeoffs associatedwith life near the oxycline would contribute tothe frequent mortality (and subsequent spec-tacular preservation) of densely aggregated,fully articulated individuals of trilobites (e.g.,Gaines and Droser 2003) and other taxa, but itis possible that such episodic risks would beoffset by the persistent advantage of having arefugium from predators with higher oxygendemands.
Episodic hypoxia would also tend to favorrapidly developing taxa that are able to takeadvantage of boom-bust oxygen cycles. Unlesspossessing unique physiological adaptations,slowly developing, high-biomass taxa wouldbe at a disadvantage in hypoxia-prone epeiricseas. Stenotopic taxa may therefore tend to berestricted to, and evolve in, regions that wererarely, if ever, subject to water column strati-fication and hypoxia. In fact, the tendency forhigher taxa to originate in nearshore settings
(e.g., Jablonski et al. 1983; Jablonski and Bottjer1991) has been specifically related to sea levelhighstands and the prevalence of stressfulhypoxia in offshore epeiric sea settings (Jacobsand Lindberg 1998). It is therefore possiblethat hypoxia in widespread epeiric seas hasaffected environmental patterns of clade orig-ination and diversification, thereby influenc-ing the familiar onshore-offshore patterns offaunal distribution (e.g., Sepkoski and Miller1985).
Biases. Given the potential of the EpeiricSea Effect to substantially impact the restruc-ture benthic marine communities, there is adanger in any paleontological study of con-founding spatial and environmental patternswith temporal evolutionary phenomena. Fullyaccounting for the Epeiric Sea Effect is, how-ever, a challenging task because the environ-mental stresses related to water-mass prop-erties can be difficult to identify without ap-pealing to the fossil assemblages themselves,a strategy that would lead to circular reason-ing in any study of fossil biodiversity.
The difficulty of identifying and fully con-trolling for the Epeiric Sea Effect, combinedwith the lack of clearly defined biological ex-pectations, makes most studies of Phanero-zoic marine biodiversity problematical to in-terpret in terms of their evolutionary signifi-cance (e.g., Bambach 1977; Sepkoski 1988; Al-roy et al. 2001; Powell and Kowalewski 2002;Fraiser and Bottjer 2004; Peters 2004; Bush andBambach 2004; and many other similar stud-ies). Comparisons between Paleozoic and Ce-nozoic fossil assemblages (e.g., Bush andBambach 2004) are particularly prone to biasin the direction of increasing diversity simplybecause of the much greater prevalence ofsamples narrow continental shelves than fromepeiric seas, but shorter-term studies (e.g., Pe-ters 2004), and even studies that focus on justone stratigraphic section, are also subject toprofound bias due to shifting water-mass ef-fects.
One potentially powerful approach to con-trolling for the Epeiric Sea Effect would be tolimit all comparisons to fossil assemblagesthat derive from similar oceanographic set-tings. This could be accomplished by consult-ing paleogeographic maps and analyzing
samples that come from continental marginsfacing the open ocean separately from thosethat derive from interior epicontinental seas.Perhaps more effectively, paleogeographic re-constructions could be used to identify re-gions that are expected, on purely physicalgrounds, to have comparable oceanographicparameters and then appropriate paleonto-logical samples could be sought out in thoseregions. Although an effective first step, thisapproach is not failsafe because many factorscan cause strong Epeiric Sea Effects besidesgeographic location (see discussion above),and it would still be necessary to control forabsolute water depth, taphonomy, diagenesis,and other sample-specific factors that can con-found biodiversity patterns.
Summary
The problem with the Paleozoic is that, onaverage, Paleozoic marine sediments derivefrom expansive epicontinental seas that weremarkedly environmentally different fromthose found on narrow, open-ocean-facingcontinental margins. Episodic hypoxia in sil-iciclastic environments, salinity variations oncarbonate platforms, freshwater lenses withinshallow and expansive epeiric seas, and otherwater-mass properties may have structuredmany benthic animal communities, includingthose that are typically considered diverseand ‘‘normal marine.’’ This is not to suggestthat all epeiric sea environments were inhos-pitable or harsh, but rather that taxa living inepeiric seas would have been subject to a dif-ferent suite of environmental conditions thanthose living in otherwise similar shelf settingslocated along narrow continental margins.The general influences that basin configura-tions, circulation patterns, and climate haveon biologically essential water-mass proper-ties and sedimentation patterns are collective-ly referred to as the ‘‘Epeiric Sea Effect.’’
Paleobiological studies that control only forwater depth, lithology, taphonomy, diagene-sis, and other similar sample-specific factorsdo not address the Epeiric Sea Effect and aretherefore subject to substantial biases that canonly be overcome by additional paleo-ocean-ographic control. However, the Epeiric Sea Ef-fect does not just impose an important envi-
ronmental sampling bias that has largely beenignored by paleobiologists interested in large-scale macroevolutionary patterns. Instead,changes in the degree of continental floodinghave forced real shifts in the average characterof shelf habitats over time, and these shifts arelikely to have had many important evolution-ary, taphonomic, and stratigraphic conse-quences. This has long been recognized byfield-oriented paleontologists, but it is nownecessary to begin incorporating these in-sights into larger-scale macroevolutionarystudies.
Fully accounting for the environmental ef-fects of expanding and contracting epeiricseas and testing the evolutionary and ecolog-ical consequences of sea level changes and theresultant formation and destruction of uniqueepeiric sea habitats requires (1) quantifyingthe shifting geographic, tectonic, and environ-mental contexts of preserved marine sedi-ments; (2) documenting the distribution offossil organisms within the environmentalmosaic of the sedimentary rock record; and (3)developing rigorous field tests to estimate pa-leo-water-mass properties and to effectivelydisentangle the relative contributions of hyp-oxia, salinity, and sedimentation in control-ling the composition, abundance, and evolu-tion of marine life.
Acknowledgments
I thank C. Badgley, M. Foote, B. Gaines, andB. Wilkinson for feedback on early drafts ofthis manuscript. C. E. Brett and S. Hollandprovided insightful reviews that greatly im-proved this manuscript. T. Baumiller served asmanaging editor. Acknowledgment is made tothe donors of the American Chemical SocietyPetroleum Research Fund for partial supportof this research and to the Michigan Society ofFellows. This work also partially supported byNational Science Foundation EAR 0544941.This is Paleobiology Database Publication 51.
Literature Cited
Algeo, T. J., P. H. Heckel, J. B. Maynard, R. C. Blakey, and H.Rowe. 2007. Modern and ancient epicontinental seas and thesuperestuarine circulation model of marine anoxia. In C.Holmden and B. Pratt, eds. Geology of epeiric seas. Geolog-ical Association of Canada (in press).
179THE EPEIRIC SEA EFFECT
Allison, P. A., and D. E. G. Briggs. 1993. Exceptional fossil rec-ord: distribution of soft-tissue preservation through the Phan-erozoic. Geology 21:527–530.
Allison, P. A., and V. P. Wright. 2005. Switching off the carbonatefactory: a-tidality, stratification and brackish wedges in epeir-ic seas. Sedimentary Geology 179:175–184.
Alroy, J., C. R. Marshall, R. K. Bambach, K. Bezusko, M. Foote,F. T. Fursich, T. A. Hansen, S. M. Holland, L. C. Ivany, D. Ja-blonski, D. K. Jacobs, D. C. Jones, M. A. Kosnik, S. Lidgard, S.Low, A. I. Miller, P. M. Novack-Gottshall, T. D. Olszewski, M.E. Patzkowsky, D. M Raup, K. Roy, J. J. Sepkoski Jr., M. G. Som-mers, P. J. Wagner, and A. Webber. 2001. Effects of samplingstandardization on estimates of Phanerozoic marine diversi-fication. Proceedings of the National Academy of SciencesUSA 98:6261–6266.
Ausich, W. I., and D. J. Bottjer. 1982. Tiering in suspension-feed-ing communities on soft substrata throughout the Phanero-zoic. Science 216:173–174.
Bambach, R. K. 1977. Species richness in marine benthic habitatsthrough the Phanerozoic. Paleobiology 3:152–167.
Baumiller, T. K., and F. J. Gahn. 2004. Testing predator-drivenevolution with Paleozoic crinoid arm regeneration. Science305:1453–1455.
Berner, R. A., D. J. Beerling, R. Dudley, J. M. Robinson, and R.A. Wildman Jr. 2003. Phanerozoic atmospheric oxygen. An-nual Review of Earth and Planetary Sciences 31:105–134.
Boesch, D. F., and N. N. Rabalais. 1991. Effects of hypoxia oncontinental shelf benthos: comparisons between the New YorkBight and the Northern Gulf of Mexico. Pp. 27–34 in Tysonand Pearson 1991b.
Bottjer, D. J., and W. I. Ausich. 1986. Phanerozoic developmentof tiering in soft substrata suspension-feeding communities.Paleobiology 12:400–420.
Bowersox, J. R. 2005. Reassessment of extinction patterns of Pli-ocene mollusks from California and environmental forcing ofextinction in the San Joaquin Basin. Palaeogeography, Palaeo-climatology, Palaeoecology 221:55–82.
Brett, C. E. 1984. Autecology of Silurian pelmatozoan echino-derms. Special Papers in Palaeontology 32:87–120.
———. 1999. Wenlockian fossil communities in New York Stateand adjacent areas paleontology and paleoecology. Pp. 592–637 in A. J. Boucot and J. D. Lawson, eds. Paleocommunities:a case study from the Silurian and Lower Devonian (Worldand Regional Geology 11). Cambridge University Press, NewYork.
Brett, C. E., P. A. Allison, C. J. Tsujita, D. Soldani, and H. A. Mof-fat. 2006. Sedimentology, taphonomy, and paleoecology ofmeter-scale cycles from the Upper Ordovician of Ontario. Pa-laios 21:530–547.
Brett, C. E., R. H. Kohrs, and B. Kirchner. 2007. Paleontologicalevent beds from the Upper Ordovician of Ohio and northernKentucky and the limits of high-resolution stratigraphy. In C.Holmden and B. Pratt, eds. Geology of epeiric seas. Geolog-ical Association of Canada (in press).
Bush, A. M., and R. K. Bambach. 2004. Did alpha diversity in-crease during the Phanerozoic? Lifting the veils of taphonom-ic, latitudinal, and environmental biases. Journal of Geology112:625–642.
Byers, C. W. 1977. Biofacies patterns in euxinic basins: a generalmodel. In H. E. Cook and P. Enos, eds. Deep-water carbonateenvironments: Society of Economic Paleontologists and Min-eralogists Special Publication 25:5–17.
Chapman, L. J., L. S. Kaufman, C. A. Chapman, and F. E.McKenzie. 1995. Hypoxia tolerance in twelve species of EastAfrican cichlids: potential for low oxygen refugia in Lake Vic-toria. Conservation Biology 9:1274–1287.
Cherns, L., and V. P. Wright. 2000. Missing molluscs as evidenceof large-scale, early skeletal aragonite dissolution in a Siluriansea. Geology 28:791–794.
Cherns, L., J. R. Wheeley, and L. Karis. 2006. Tunneling trilo-bites: habitual infaunalism in an Ordovician carbonate floor.Geology 34:657–660.
Diaz, R. J. 2001. Overview of hypoxia around the world. Envi-ronmental Quality 30:275–281.
Diaz, R. J., and R. Rosenberg. 1995. Marine benthic hypoxia: areview of its ecological effects and the behavioural responsesof benthic macrofauna. Oceanography and Marine BiologyAnnual Review 33:245–303.
Fagerstrom, J. A., and O. Weidlich. 2005. Biologic response toenvironmental stress in tropical reefs: lessons from modernPolynesian coralgal atolls and Middle Permian sponge andShamovella-microbe reefs (Capitan Limestone USA). Facies 51:501–515.
Fisher, D. C. 1977. Mechanism and significance of enrollment inxiphosurans (Chelicerata, Merostomes). Geological Society ofAmerica Abstracts with Programs 9:264–265.
Follmi, K. B., and K. A. Grimm. 1990. Doomed pioneers: gravityflow deposition and bioturbation in marine oxygen-deficientenvironments. Geology 18:1069–1072.
Fraiser, M. L., and D. J. Bottjer. 2004. The non-actualistic EarlyTriassic gastropod fauna: a case study of the Lower TriassicSinbad Limestone Member. Palaios 19:259–275.
Frest, T. J., C. E. Brett, and B. J. Witzke. 1999. Caradocian-Ged-innian echinoderm associations of central and eastern NorthAmerica. World and Regional Geology 11:638–783.
Fursich, F. T. 1993. Palaeoecology and evolution of Mesozoic sa-linity-controlled benthic macroinvertebrate associations. Le-thaia 26:327–346.
Gaines, R. R., and M. L. Droser. 2003. Paleoecology of the fa-miliar trilobite Elrathia kingii: an early exaerobic zone inhab-itant. Geology 31:941–944.
Ginsburg, R. N. 1982. Actualistic depositional models for theGreat American Bank (Cambro-Ordovician). InternationalCongress on Sedimentology 11:114.
Helly, J. J., and L. A. Levin. 2004. Global distribution of naturallyoccurring marine hypoxia continental margins. Deep-Sea Re-search Part I 51:1159–1168.
Henderson, R. A. 2004. A mid-Cretaceous association of shellbeds and organic-rich shale: bivalve exploitation of a nutrient-rich, anoxic sea-floor environment. Palaios 19:156–169.
Holland, S. M., A. I. Miller, and D. L. Meyer. 2000. High-reso-lution correlation in apparently monotonous rocks: Upper Or-dovician Kope Formation. Palaios 15:73–80.
Hughes, N. C., and D. L. Cooper. 1999. Paleobiologic and taph-onomic aspects of the ‘‘granulosa’’ trilobite cluster, Kope For-mation (Upper Ordovician, Cincinnati region). Journal of Pa-leontology 73:306–319.
Hunda, B. R., N. C. Hughes, and K. W. Flessa. 2006. Trilobitetaphonomy and temporal resolution in the Mt. Orab ShaleBed (Upper Ordovician, Ohio, U.S.A.). Palaios 21:26–45.
Jablonski, D., and D. J. Bottjer. 1991. Environmental patterns inthe origins of higher taxa: the post-Paleozoic fossil record.Science 252:1831–1833.
Jablonski, D., J. J. Sepkoski Jr., D. J. Bottjer, and P. M. Sheehan.1983. Onshore-offshore patterns in the evolution of Phaner-ozoic shelf communities. Science 222:1123–1125.
Jacobs, D. K., and D. R. Lindberg. 1998. Oxygen and evolution-ary patterns in the sea: onshore/offshore trends and recentrecruitment of deep-sea faunas. Proceedings of the NationalAcademy of Sciences USA 95:9396–9401.
Johnson, J. G. 1974. Extinction of perched faunas. Geology 2:479–482.
Kammer, T. W., and W. I. Ausich. 2006. The ‘‘age of crinoids’’: aMississippian biodiversity spike coincident with widespreadcarbonate ramps. Palaios 21:238–248.
Kamykowski, D., and S.-J. Zentara. 1990. Hypoxia in the worldocean as recorded in the historical data set. Deep-Sea Re-search 37:1861–1874.
Keeling, R. F., and H. E. Garcia. 2002. The change in oceanic O2
inventory associated with recent global warming. Proceed-ings of the National Academy of Sciences USA 99:7848–7853.
Kidwell, S. M. 1988. Taphonomic comparison of passive and ac-tive continental margins: Neogene shell beds of the Atlanticcoastal plain and northern Gulf of California. Palaeogeogra-phy, Palaeoclimatology, Palaeoecology 63:201–223.
Kohrs, R., C. E. Brett, and N. O’Brien. 2007. Sedimentology ofUpper Ordovician mudstones from the Cincinnati Arch re-gion, Ohio/Kentucky: toward a general model of mud eventdeposition. In P. I. McLaughlin, C. E. Brett, S. L. Taha Mc-Laughlin, and J. Bazeley, eds. Stratigraphic renaissance in theCincinnati Arch: implications for Upper Ordovician paleon-tology and paleoecology. Cincinnati Museum Center SpecialPublication 2 (in press).
Levin, L. A. 2003. Oxygen minimum zone benthos: adaptationand community responses to hypoxia. Oceanography andMarine Biology 41:1–45.
Miller, A. I., S. M. Holland, D. L. Meyer, and B. F. Dattilo. 2001.The use of faunal gradient analysis for intraregional correla-tion and assessment of changes in sea-floor topography in thetype Cincinnatian. Journal of Geology 109:603–613.
Miller, K. J., M. A. Kominz, J. V. Browning, J. D. Wright, G. S.Mountain, M. E. Katz, P. J. Sugarman, B. S. Cramer, N. Chris-tie-Blick, and S. F. Pekar. 2005. The Phanerozoic record ofglobal sea-level change. Nature 310:1293–1298.
Newell, N. D. 1949. Periodicity in invertebrate evolution. Geo-logical Society of America Bulletin 60:1911–1912.
———. 1952. Periodicity in invertebrate paleontology. Journal ofPaleontology 26:371–385.
Nordberg, K., H. L. Filipsson, M. Gustafsson, R. Harland, andP. Ross. 2001. Climate, hydrographic variations and marinebenthic hypoxia in Koljo Fjord, Sweden. Journal of Sea Re-search 46:187–200.
Osgood, R. G. 1970. Trace fossils of the Cincinnati area. Palaeon-tographica Americana 6:281–444.
Osterman, L. E., R. Z. Poore, P. W. Swarzenski, and R. E. Turner.2005. Reconstructing a 180 yr record of natural and anthro-pogenic induced low-oxygen conditions from Louisiana con-tinental shelf sediments. Geology 33:329–332.
Peters, S. E. 2004. Evenness of Cambrian-Ordovician benthicmarine communities in North America. Paleobiology 30:325–346.
———. 2005. Geologic constraints on the macroevolutionaryhistory of marine animals. Proceedings of the National Acad-emy of Sciences USA 102:12326–12331.
———. 2006. Macrostratigraphy of North America. Journal ofGeology 114:391–412.
Pihl, L., S. P. Baden, R. J. Diaz, and L. C. Shaffner. 1992. Hyp-oxia-induced structural changes in the diet of bottom-feedingfish and Crustacea. Marine Biology 112:349–361.
Powell, M. G., and M. Kowalewski. 2002. Increase in evennessand sampled alpha diversity through the Phanerozoic: com-parison of early Paleozoic and Cenozoic marine fossil assem-blages. Geology 30:331–334.
Rabalais, N. N., E. Turner, and W. J. Wiseman Jr. 2002. Gulf ofMexico hypoxia, a.k.a. ‘‘the dead zone.’’ Annual Review ofEcology and Systematics 33:235–363.
Reaves, C. M. 1986. Organic matter metabolizability and calci-um carbonate dissolution in nearshore marine muds. Journalof Sedimentary Petrology 56:486–494.
Rhoads, D. C., and J. W. Morse. 1971. Evolutionary and ecologicsignificance of oxygen-deficient basins. Lethaia 4:413–428.
Ronov, A. B. 1978. The Earth’s sedimentary shell. InternationalGeology Review 24:1313–1363.
———. 1994. Phanerozoic transgressions and regressions on thecontinents: a quantitative approach based on areas flooded bythe sea and areas of marine and continental deposition.American Journal of Science 294:777–801.
Ronov, A. B., V. E. Khain, A. N. Balukhovsky, and K. B. Sesla-vinsky. 1980. Quantitative analysis of Phanerozoic sedimen-tation. Sedimentary Geology 25:311–325.
Ruedemann, R. 1935. Ecology of black mud shales of easternNew York. Journal of Paleontology 9:79–91.
Savrda, C. E., and D. J. Bottjer. 1986. Trace-fossil model for re-construction of paleo-oxygenation in bottom waters. Geology14:306–309.
———. 1987. The exaerobic zone, a new oxygen-deficient marinebiofacies. Nature 327:54–56.
Schopf, T. J. M. 1974. Permo-Triassic extinctions: relation to sea-floor spreading. Journal of Geology 82:129–143.
Schovsbo, N. H. 2001. Why barren intervals? A taphonomic casestudy of the Alum Shale and its faunas. Lethaia 34:271–285.
Schumacher, G. A., and D. L. Shrake. 1997. Paleoecology andcomparative taphonomy of an Isotelus (Trilobita) fossil lag-erstatten from the Waynesville Formation (Upper Ordovician,Cincinnatian Series) of southwestern Ohio. Pp. 131–161 in C.E. Brett and G. C. Baird, eds. Paleontological events: strati-graphic, ecological, and evolutionary implications. ColumbiaUniversity Press, New York.
Sepkoski, J. J., Jr. 1988. Alpha, beta, or gamma: where does allthe diversity go? Paleobiology 14:221–234.
———. 2002. A compendium of fossil marine animal genera.Bulletins of American Paleontology 363:560.
Sepkoski, J. J., Jr., and A. I. Miller. 1985. Evolutionary faunas andthe distribution of Paleozoic benthic communities in spaceand time. Pp. 153–190 in J. W. Valentine, ed. Phanerozoic di-versity patterns: profiles in macroevolution. Princeton Uni-versity Press, Princeton, N.J.
Signor, P. W., III, and C. E. Brett. 1984. The mid-Paleozoic pre-cursor to the Mesozoic marine revolution. Paleobiology 10:229–245.
Simberloff, D. S. 1974. Permo-Triassic extinctions: effects of areaon biotic equilibrium. Journal of Geology 82:267–274.
Sorokin, Y. I. 2002. The Black Sea: ecology and oceanography.Backhuys, Leiden.
Speyer, S. E. 1990. Enrollment in trilobites. Pp. 450–455 in A. J.Boucot, ed. Evolutionary paleobiology of behavior and coevo-lution. Elsevier, Amsterdam.
Speyer, S. E., and C. E. Brett. 1986. Trilobite taphonomy andMiddle Devonian taphofacies. Palaios 1:312–327.
Stachowitsch, M. 1991. Anoxia in the Northern Adriatic Sea:rapid death, slow recovery. Pp. 119–130 in Tyson and Pearson1991b.
Tsujita, C. J., C. E. Brett, M. Topor, and J. Topor. 2006. Evidenceof high-frequency storm disturbance in the Middle DevonianArkona Shale, southwestern Ontario. Journal of Taphonomy4:49–68.
Tyson, R. V., and T. H. Pearson. 1991a. Modern and ancient con-tinental shelf anoxia: an overview. Pp. 1–26 in Tyson and Pear-son 1991b.
Tyson, R. V., and T. H. Pearson, eds. 1991b. Modern and ancientcontinental shelf anoxia. Geological Society of London SpecialPublication 58.
Valentine, J. W., and E. M. Moores. 1970. Plate-tectonic regula-tion of faunal diversity and sea level: a model. Nature 228:657–659.
Veizer, J., and R. E. Ernst. 1996. Temporal patterns of sedimen-tation: Phanerozoic of North America. Geochemistry Inter-national 33:64–76.
Webber, A. J. 2002. High-resolution faunal gradient analysis andan assessment of the causes of meter-scale cyclicity in the typeCincinnatian Series (Upper Ordovician). Palaios 17:545–555.
Wells, M. R., P. A. Allison, G. J. Hampson, M. D. Piggott, and C.C. Pain. 2005. Modeling ancient tides: the Upper Carbonifer-ous epi-continental seaway of Northwest Europe. Sedimen-tology 52:715–735.
Wignall, P. B. 1994. Black shales. Oxford Monographs on Geol-ogy and Geophysics No. 30. Clarendon, Oxford.