The Radiolarian biotic response during OAE2. A high-resolution studyacross the Bonarelli level at Bottaccione (Gubbio, Italy)
La reponse biotique des Radiolaires durant le OAE2. Une etude ahaute-resolution a travers le niveau Bonarelli a
Bottaccione (Gubbio, Italie)
Benjamin Musavu-Moussavou a,b,∗, Taniel Danelian b, Francois Baudin c,Rodolfo Coccioni d, Francois Frohlich e
a Departement de geologie et Ureste, universite des sciences et techniques de Masuku, BP 943, Franceville, Gabonb Universite Pierre-et-Marie-Curie (Paris 6), CNRS-UMR 5143, case 104, 4, place Jussieu, 75252 Paris cedex 05, Francec Universite Pierre-et-Marie-Curie (Paris 6), CNRS-UMR 7072, case 117, 4, place Jussieu 75252 Paris cedex 05, France
d Istituto di Geologia e Centro di Geobiologia dell’Universita degli Studi “Carlo Bo”, Campus Scientifico, Localita Crocicchia, 61029 Urbino, Italye Museum national d’histoire naturelle, 43, rue Buffon, 75005 Paris, France
bstract
In order to improve our understanding of the radiolarian biotic response and the palaeoceanographic changes associated with the Late Cenomanianceanic anoxic event (OAE2), a high-resolution radiolarian study was carried out across the Bonarelli level at its typical locality (Bottaccioneection, central Italy). Our results confirm that a drastic radiolarian faunal change took place during the OAE2. The radiolarian turnover took placeithin the median part of the Bonarelli level, associated with an interval of high organic matter preservation, but relatively low values of silica.herefore, this part constitutes a critical period in the evolutionary history of Radiolaria. A synthesis of all data available in the Umbria-Marcheasin suggests that the lower part of the Bonarelli level (and underlying limestones) records mainly radiolarian extinctions (ca. 41%), while itspper part (and overlying limestones) records mainly first occurrences (ca. 35%). Our results improve understanding of the paleoecology of theamily Saturnalidae, which would seem to prefer oligotrophic environments.
2007 Elsevier Masson SAS. All rights reserved.
esume
Dans l’optique d’ameliorer notre comprehension de la reponse biotique des radiolaires et des changements paleoceanographiques associes avec’evenement anoxique du Cenomanien terminal (OAE2), une etude a haute-resolution a ete effectuee a travers le niveau Bonarelli de la coupe deottaccione (Italie centrale). Les donnees de cette etude ont ete completees par celles issues de la bibliographie. Nos resultats confirment qu’unhangement drastique des faunes de radiolaires a eu lieu au cours de l’OAE2. Ce renouvellement des radiolaires a eu lieu au sein de la partieediane du niveau Bonarelli, qui est associee avec un intervalle de preservation d’une quantite importante de matiere organique, mais des quantites
elativement faibles de silice. Par consequent, cet intervalle constitue une periode critique dans l’histoire evolutive des radiolaires. Juste avant etment des extinctions, estimees a 41 %. En revanche, un large nombre deieme moitie de ce niveau critique et juste apres celui-la. L’enregistrementpeu mieux la paleoecologie de la famille Saturnalidae, qui semblerait avoir
u sein de la premiere moitie du niveau Bonarelli, on note essentielleremieres occurrences, estimees a 35 %, sont enregistrees dans la deuxes radiolaires dans la coupe de Bottaccione permet de comprendre unne preference pour des environnements oligotrophiques.
2007 Elsevier Masson SAS. All rights reserved.
eywords: Radiolaria; Oceanic anoxic events; OAE2; Umbria-Marche basin; Italy; C
Fig. 1. Geological sketch map of the Gubbio area (central Italy) with locationof the Bottaccione section.Fm
TSpc1
tsBbl
goa(abPp
54 B. Musavu-Moussavou et al. / Revue
. Introduction
The mid Cretaceous (Barremian–Turonian) represents aemarkable greenhouse period in earth history. Sediments richn organic carbon accumulated during distinct intervals, knowns oceanic anoxic events (OAEs), thought to represent relativelyhort intervals of major disturbance in the marine environmentSchlanger and Jenkyns, 1976; Jenkyns, 1980). Amongst thesevents, the Late Cenomanian OAE2 has a global distribution anduration estimated between 400 and 600 ka (Morel, 1998; Caront al., 1999; Scopelliti et al., 2004). OAE2 had a profound impactn the evolution and productivity of the marine biosphere (Kuhntt al., 1986; Hallam and Wignall, 1997; Harries and Little, 1999;eckie et al., 2002).
During OAE2, Radiolaria appear to have responded tohe environmental perturbation by significant changes in theirbundance and assemblage composition (Marcucci Passerinit al., 1991; Erbacher, 1994; O’Dogherty, 1994; Gallicchio etl., 1996; Erbacher and Thurow, 1997; Premoli Silva et al.,999; O’Dogherty and Guex, 2002; Musavu-Moussavou andanelian, 2006). Based on data provided by Erbacher, 1994;’Dogherty, 1994; Erbacher and Thurow, 1997; O’Dogherty
nd Guex (2002), it was respectively estimated that approxi-ately 60% of Cenomanian species went extinct during OAE2,hile ca. 50% of Turonian species first occurred during or just
fter this oceanic event. Both studies were based on data frommbria-Marche basin of Italy.However, major discrepancies exist in the type and quantity
f data used by the above authors: (1) the total number of speciesnvestigated for the late Cenomanian–early Turonian interval isuite different in each study (i.e., 70 species were consideredy Erbacher, 1994 and 152 by O’Dogherty, 1994), (2) taxo-omic concepts are quite different for a great number of speciesetween the two authors, (3) calculation methods are differentn the two works for percent extinctions and originations, (4)’Dogherty did not consider any radiolarian data from within
he Bonarelli level (BL). On the other hand, although, Erbacher1994) studied samples from within the critical interval (BL), theumber of species one finds since his work has increased sub-tantially, or the stratigraphic range of previously found speciess now modified.
During the present work, we deployed a high-resolution radi-larian study across the BL and surrounding limestones with thebjective to obtain an improved insight on the pattern and inten-ity of the radiolarian faunal change across the OAE2 in thembria-Marche basin (UMB).
. Stratigraphic framework
The Cretaceous part of the Umbria-Marche pelagic sequenceccumulated in a basin of the Apulian block formed since theower Jurassic. The BL and surrounding limestones are wellxposed in the classical reference section of the Bottaccione
orge (Gubbio area, Fig. 1) located a few kilometres north-ast of the town of Gubbio, along the road S.S. 298. The BLepresents a regional stratigraphic marker horizon that occursowards the top of the uppermost Cenomanian to lowermost
wpps
ig. 1. Esquisse de la carte geologique de la region de Gubbio (Italie centrale)ontrant la localisation de la section de Bottaccione.
uronian Scaglia Bianca limestone formation. The facies of thecaglia Bianca limestones corresponds to lithified nannofossil-lanktonic foraminiferal ooze, which accumulated above thealcite compensation depth, at an estimated water-depth of500–2500 m (Arthur and Premoli Silva, 1982; Kuhnt, 1990).
The interval studied here includes the BL and nearly 12 m ofhe underlying and 1 m of the overlying Scaglia Bianca lime-tones (Fig. 2). The rather monotonous sequence of the Scagliaianca limestones underlying the BL is marked at Bottaccioney the intercalation of black marls and shales, as well as severalayers of nodules and lenses of dark-grey to black chert.
In the studied section, the BL consists of carbonate-free olive-reen to black mudstones and Corg-rich finely laminated shales,ften rich in fish remains, pyrite nodules and/or radiolaria. Shalesnd mudstones alternate with radiolarian-rich siliceous layersArthur and Premoli Silva, 1982; Coccioni et al., 1991; Coccionind Luciani, 2004). Based on lithological criteria, the BL cane subdivided into three parts throughout the UMB (Arthur andremoli Silva, 1982; Coccioni et al., 1991). Its lower and upperarts consist, in general, of radiolarian-rich siliceous layers,
hereas its median part is dominated by black mudstones. Morearticularly, the 82-cm-thick BL of the Bottaccione section dis-lays a 32-cm-thick lower siliceous part, a 16-cm-thick medianhaly part and a 34-cm-thick siliceous upper part.
B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287 255
Fig. 2. Lithostratigraphy of the studied interval of the Bottaccione section plotted against (A), radiolarian % abundance estimated on thin section, according to thediagrams of visual percentage estimation suggested by Baccelle and Bosellini (1965) (cross (x) corresponds to samples for which no thin section was produced);(B), radiolarian species diversity and (C), radiolarian preservation according to the scale suggested by Kiessling (1996) (BL, Bonarelli level).Fig. 2. Lithostratigraphie de l’intervalle etudie de la coupe de Bottaccione, correlee avec (A), l’abondance des radiolaires en lame-mince, exprimee en pourcentageselon la charte de Baccelle et Bosellini, (1965) (les croix (x) correspondent a des echantillons pour lesquels des lames minces n’ont pas ete realisees); (B), la diversitespecifique des radiolaires et (C), leur preservation selon l’echelle proposee par Kiessling (1996) (BL, Bonarelli level).
2 de mi
3
stslBasparemdai
wwKB(wfoai
ap
hR1
csf
FpFs
iT
4
4
iomatdBtpa(
mttqaKocs
tttca(
56 B. Musavu-Moussavou et al. / Revue
. Material and methods
Study of radiolaria was carried out based on a total of 120amples collected from the BL and surrounding limestones ofhe Bottaccione section (Fig. 2). The sampled lithologies con-ist of limestone and chert nodules and lenses from the carbonateevels and radiolarian cherts and black shales from within theL. Samples were collected every 2 cm from within the BLnd with a lower resolution (between 2 and 30 cm) from theurrounding limestones. Radiolarians were extracted followingrocessing with 10% hydrochloric acid (for limestones samples)nd 4% hydrofluoric acid (for siliceous samples), according tooutine laboratory procedures (Dumitrica, 1970; Baumgartnert al., 1981; De Wever, 1982). Radiolarian abundance was esti-ated based on thin section observations, according to the
iagrams of visual percentage estimation suggested by Baccellend Bosellini (1965). Preservation was evaluated based on thendex scale suggested by Kiessling (1996).
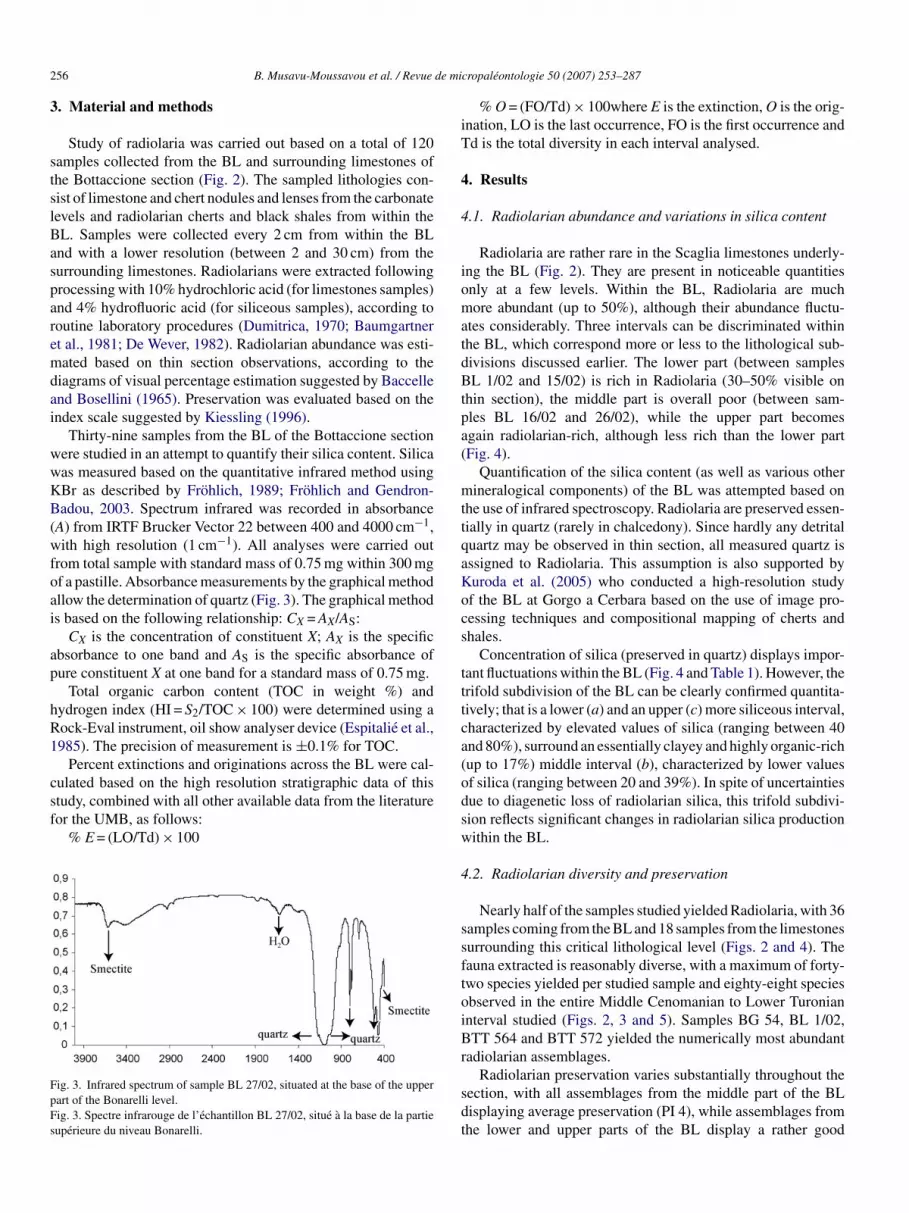
Thirty-nine samples from the BL of the Bottaccione sectionere studied in an attempt to quantify their silica content. Silicaas measured based on the quantitative infrared method usingBr as described by Frohlich, 1989; Frohlich and Gendron-adou, 2003. Spectrum infrared was recorded in absorbance
A) from IRTF Brucker Vector 22 between 400 and 4000 cm−1,ith high resolution (1 cm−1). All analyses were carried out
rom total sample with standard mass of 0.75 mg within 300 mgf a pastille. Absorbance measurements by the graphical methodllow the determination of quartz (Fig. 3). The graphical methods based on the following relationship: CX = AX/AS:
CX is the concentration of constituent X; AX is the specificbsorbance to one band and AS is the specific absorbance ofure constituent X at one band for a standard mass of 0.75 mg.
Total organic carbon content (TOC in weight %) andydrogen index (HI = S2/TOC × 100) were determined using aock-Eval instrument, oil show analyser device (Espitalie et al.,985). The precision of measurement is ±0.1% for TOC.
Percent extinctions and originations across the BL were cal-
ulated based on the high resolution stratigraphic data of thistudy, combined with all other available data from the literatureor the UMB, as follows:
% E = (LO/Td) × 100
ig. 3. Infrared spectrum of sample BL 27/02, situated at the base of the upperart of the Bonarelli level.ig. 3. Spectre infrarouge de l’echantillon BL 27/02, situe a la base de la partieuperieure du niveau Bonarelli.
odsw
4
ssftoiBr
sdt
cropaleontologie 50 (2007) 253–287
% O = (FO/Td) × 100where E is the extinction, O is the orig-nation, LO is the last occurrence, FO is the first occurrence andd is the total diversity in each interval analysed.
. Results
.1. Radiolarian abundance and variations in silica content
Radiolaria are rather rare in the Scaglia limestones underly-ng the BL (Fig. 2). They are present in noticeable quantitiesnly at a few levels. Within the BL, Radiolaria are muchore abundant (up to 50%), although their abundance fluctu-
tes considerably. Three intervals can be discriminated withinhe BL, which correspond more or less to the lithological sub-ivisions discussed earlier. The lower part (between samplesL 1/02 and 15/02) is rich in Radiolaria (30–50% visible on
hin section), the middle part is overall poor (between sam-les BL 16/02 and 26/02), while the upper part becomesgain radiolarian-rich, although less rich than the lower partFig. 4).
Quantification of the silica content (as well as various otherineralogical components) of the BL was attempted based on
he use of infrared spectroscopy. Radiolaria are preserved essen-ially in quartz (rarely in chalcedony). Since hardly any detritaluartz may be observed in thin section, all measured quartz isssigned to Radiolaria. This assumption is also supported byuroda et al. (2005) who conducted a high-resolution studyf the BL at Gorgo a Cerbara based on the use of image pro-essing techniques and compositional mapping of cherts andhales.
Concentration of silica (preserved in quartz) displays impor-ant fluctuations within the BL (Fig. 4 and Table 1). However, therifold subdivision of the BL can be clearly confirmed quantita-ively; that is a lower (a) and an upper (c) more siliceous interval,haracterized by elevated values of silica (ranging between 40nd 80%), surround an essentially clayey and highly organic-richup to 17%) middle interval (b), characterized by lower valuesf silica (ranging between 20 and 39%). In spite of uncertaintiesue to diagenetic loss of radiolarian silica, this trifold subdivi-ion reflects significant changes in radiolarian silica productionithin the BL.
.2. Radiolarian diversity and preservation
Nearly half of the samples studied yielded Radiolaria, with 36amples coming from the BL and 18 samples from the limestonesurrounding this critical lithological level (Figs. 2 and 4). Theauna extracted is reasonably diverse, with a maximum of forty-wo species yielded per studied sample and eighty-eight speciesbserved in the entire Middle Cenomanian to Lower Turoniannterval studied (Figs. 2, 3 and 5). Samples BG 54, BL 1/02,TT 564 and BTT 572 yielded the numerically most abundant
adiolarian assemblages.
Radiolarian preservation varies substantially throughout the
ection, with all assemblages from the middle part of the BLisplaying average preservation (PI 4), while assemblages fromhe lower and upper parts of the BL display a rather good
B.M
usavu-Moussavou
etal./Revue
dem
icropaleontologie50
(2007)253–287
257
Fig. 4. Lithostratigraphic detail of the Bonarelli level at Bottaccione, plotted against (A), radiolarian % abundance estimated on thin section, according to the diagrams of visual percentage estimation suggestedby Baccelle and Bosellini (1965) (cross (x) corresponds to samples for which no thin section was produced) ; (B), radiolarian species diversity and (C), radiolarian preservation according to the scale suggested byKiessling (1996) ; (D), TOC values (according to Scopelliti et al., 2006) ; (E), HI (mgHC/gTOC) (data after Scopelliti et al., 2006) ; (F), abundance of silica measured as quartz by infrared spectroscopy ; (G), thecurve of �13C after Tsikos et al. (2004) and (H), radiolarian events based on this study.Fig. 4. Description lithologique du niveau Bonarelli de la coupe de Bottaccione correlee avec (A), l’abondance des radiolaires exprimee en pourcentage selon la charte de Baccelle et Bosellini (1965) (les croix (x)correspondent a des echantillons pour lesquels des lames minces n’ont pas ete realisees) ; (B), la diversite specifique des radiolaires ; (C), leur preservation selon l’echelle proposee par Kiessling (1996) ; (D), lesvaleurs de carbone organique total (TOC) ; (E), de l’IH (mgHC/gCOT) (d’apres Scopelliti et al., 2006) ; (F), l’abondance de silice (en forme de quartz) ; (G), la courbe du �13C selon Tsikos et al. (2004) et (H), lesquelques bioevenements des radiolaires reconnus sur la base de cette etude.
258 B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287
F ion.F e Bot
ptfvp(
4
pUd
taP1M
Boa
Fr*Fc(
ig. 5. Occurrence of Radiolaria in the studied samples of the Bottaccione sectig. 5. Occurrence des radiolaires au sein des echantillons etudies de la coupe d
reservation (PI 2). All assemblages from limestones underlyinghe BL display poor preservation (PI 8), whereas assemblagesrom limestones overlying the BL display a rather good preser-ation. No correlation appears to exist between radiolarianreservation, radiolarian diversity and radiolarian abundancesee Figs. 2 and 4).
.3. Radiolarian bioevents and patterns of faunal changes
Detailed occurrence data as resulted from the study of sam-les from Bottaccione section are presented in Fig. 5. BecauseMB is currently the palaeogeographic realm with the bestocumented radiolarian record across the BL, Fig. 6 combines
laae
ig. 6. Radiolarian distribution across the BL in UMB. This figure synthesises dataecord across the BL in UMB: (a) Marcucci Passerini et al. (1991), (b) Erbacher (1, species identified under a different specific name by the cited authors.ig.6. Distribution des radiolaires a travers le niveau Bonarelli (NB) dans le bassin deette etude (e) avec toutes les autres disponibles sur les radiolaires a travers le NB de B1994) et (d) Salvini et Marcucci Passerini (1998). *, espece determinee sous un autr
taccione.
he data issued from the present high-resolution study withll those provided in previous studies from UMB (Marcucciasserini et al., 1991; Erbacher, 1994; Erbacher and Thurow,997; O’Dogherty, 1994; Gallicchio et al., 1996; Salvini andarcucci Passerini, 1998).The radiolarian turnover that occurs in the middle part of the
L allows the distinction of a “lower fauna” from an “upper”ne, in agreement with Marcucci Passerini et al., 1991; Salvinind Marcucci Passerini (1998). Fifty-four radiolarian species
ast occur during the lower–middle part of the BL (lower fauna)nd underlying limestones. They represent 40.6% of extinctionsmongst the Upper Cenomanian radiolarian fauna. Extinctionvents were initiated since the Middle Cenomanian only to
obtained during this study (e) and all those data available on the radiolarian994), (c) O’Dogherty (1994) and (d) Salvini and Marcucci Passerini (1998).
l’Ombrie-Marches (BOM). Cette figure constitue une synthese des donnees deOM : (a) Marcucci Passerini et al. (1991), (b) Erbacher (1994), (c) O’Doghertye nom par les auteurs correspondants.
B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287 259
260 B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287
Fig. 6. (Continued ).
B. Musavu-Moussavou et al. / Revue de mi
Table 1Quantitative results of infrared spectroscopy on 39 samples from the BL of theBottaccione sectionTableau 1Resultats d’analyses en spectroscopie infrarouge quantitative par la methode dupastillage dans le KBr de 39 echantillons du NB de la section de Bottacione
ttiabsratalso corresponds to the lowest values in radiolarian abundancethroughout the BL, indicating a radiolarian productivity crush in
ttend their climax within the middle part of the BL. This latternterval represents a turning point in radiolarian evolution, afterhich mainly first occurrences are recognised in a progressiveay. Forty-two species first occur in the middle–upper part of
he BL (upper fauna) and the overlying lower Turonian lime-tones. They represent 34.7% of originations within the latestenomanian radiolarian fauna.
Within the BL, assemblages yielded during this study areominated by Nassellaria, and more particularly by speciesrchaeodityomitra sliteri and Stichomitra communis. Membersf the family Saturnalidae, although abundant and diverse in thenderlying limestones, disappear temporarily at the base of the
L, only to become again abundant and diverse at the base of
he overlying limestones.tr
cropaleontologie 50 (2007) 253–287 261
. Discussion
The BL highlights the special paleoceanographic signifi-ance of the OAE2 in the UMB. Based on sedimentological,icropalaeontological and geochemical data, the BL is
egarded as reflecting an interval of eutrophic, anoxic to euxiniconditions in the water column of the UMB, and contrastsith the oligotrophic conditions of the surrounding limestones
Jenkyns, 1980; Arthur and Premoli Silva, 1982; Coccioni et al.,991, 1992; Tsikos et al., 2004; Scopelliti et al., 2004, 2006).
Although radiolarian occurrence is greatly influenced byreservation, the absence or relative paucity of the family Satur-alidae cannot be solely explained by a differential preservation,ecause the ring present in all members of this family has aigh preservation potential, often found as broken fragments.herefore, their absence (or rarity) and contrasting abundancend diversity in the surrounding limestones is here interpreteds bearing an original palaeoecological significance. It pointso a preference for members of this family for oligotrophiconditions, as those that prevailed during the deposition ofhe Scaglia limestones (Coccioni and Luciani, 2004). On thether hand, the relative abundance of Archaeodictyomitra sli-eri and Stichomitra communis in the assemblages recoveredrom the BL, may reflect the preference of these species forutrophic environments, although their abundance is probablylso influenced by the better preservational potential of theirtout tests.
Within the BL, we can clearly recognize a lower and an upperssemblage (lower and upper fauna, respectively), which corre-pond to those previously recognized by Marcucci Passerini etl., 1991; Salvini and Marcucci Passerini, 1998. The significantadiolarian turnover that is crystallised in the middle shaly part ofhe BL (separating step-by-step extinctions below, from an uppernterval of progressive enrichment in new species) correlatesith the highest values of preserved organic matter, presencef abundant pyrite crystals and the lowest values of radiolarianbundance within the critical interval. All these suggest that theurning point in the pattern of change in radiolarian diversity tooklace during an interval of intensified anoxic conditions in theater column associated with a crush in radiolarian productivity.
. Conclusions
The high resolution analysis of the radiolarian record acrosshe BL in its typical locality confirms and further specifies theiming and pattern of radiolarian turnover which took place dur-ng the OAE2 in UMB, as suggested previously by Erbachernd Thurow (1998). The lower part of the BL is characterizedy step-by-step extinctions, which took place up to a critical levelituated at the center of the middle shaly interval. This level rep-esents an interval of intensified anoxic conditions in the UMB,s these are reflected by the abundance of preserved organic mat-er and pyrite crystals in the middle shale interval of the BL. It
he UMB. It crystallises a critical turning point in the record ofadiolarian diversity in this pelagic basin of Tethys, after which
2 de mi
aaulec
Swooarf
7
p
p
t1rT
P
O
tQ
6a
P
olUvu
F
a
2
t1C
F
e
P
c
P
a
uSln
AaeC
62 B. Musavu-Moussavou et al. / Revue
large number of new species occurred progressively. Newnd revised taxonomic and stratigraphic data allow improvednderstanding of the level of extinctions (nearly 41% of theate Cenomanian fauna) and originations (nearly 35% of the lat-st Cenomanian–Turonian fauna) that took place in radiolarianommunities, which were overestimated in previous studies.
Two Nassellarian species (Archaeodityomitra sliteri andtichomitra communis) dominate the radiolarian assemblagesithin the BL, which may be indicative of a specific preferencef these species to eutrophic conditions. The absence (or rarity)f Saturnalidae within the BL contrasts with their abundancend diversity in the underlying and overlying limestones and isegarded here as an ecological preference of members of thisamily for oligotrophic environments.
. Systematic palaeontology
Order ENTACTINARIA Kozur and Mostler, 1982Family QUINQUECAPSULARIIDAE Dumitrica, 1975Genus Protoxiphotractus Pessagno, 1973Type species: Protoxiphotractus perplexus Pessagno, 1973
. S403, Pl. 18, Fig. 1.Occurrence: In the UMB, this species is known to occur in
he Lower Turonian limestones overlying the BL (O’Dogherty,994 and this study, sample BTT 566B). However, it is alsoeported from Middle–Upper Cenomanian strata of northernurkey (Bragina, 2004).
Material: Two specimens.
Genus Quinquecapsularia Pessagno, 1971bType species: Quinquecapsularia spinosa Pessagno, 1971b
Quinquecapsularia parvipora (Squinabol, 1903b)Plate 1, Fig. 21903b. Acanthosphaera parvipora n. sp. – Squinabol, p. 115,
l. 8, Fig. 5.1994. Quinquecapsularia parvipora (Squinabol) –
’Dogherty, p. 269, Pl. 47, Figs. 25–28.Occurrence: This species is reported from the Middle Albian
o Lower Turonian of the UMB (O’Dogherty, 1994). In this study. parvipora was found only in the limestones overlying the BL.Material: Four specimens.
Family SATURNALIDAE Deflandre, 1953Subfamily SATURNALINAE Deflandre, 1953
Genus Acanthocircus Squinabol, 1903bType species: Acanthocircus irregularis Squinabol, 1903b
Acanthocircus bestiarius O’Dogherty, 1994
cropaleontologie 50 (2007) 253–287
Plate 1, Fig. 31973. Spongosaturnalis (?) sp. – Foreman, p. 251, Pl. 14, Figs.
–8; non Figs. 4 and 5 (=A. venetus), 9 (=A. dendroacanthos)nd Pl. 15, Figs. 2 and 3.
Occurrence: During this study, A. bestiarius was found innly one sample (BL 572) coming from the limestones over-ying the BL. However, this species is reported to occur in theMB throughout Upper Cenomanian to Lower Turonian inter-al (U.A. 18–21 of O’Dogherty, 1994), including the limestonesnderlying the BL.
Material: Two specimens.
Acanthocircus euganeus (Squinabol, 1914)Plate 1, Fig. 41914. Saturnalis euganeus n. f. – Squinabol, p. 300, Pl. 24(5),
bol – Foreman, Pl. 15, Fig. 5.1994. Acanthocircus euganeus (Squinabol) – O’Dogherty, p.
57, Pl. 45, Figs. 14–16 (and entire synonymy).Occurrence: In the UMB, this species is known to occur in
he Lower Turonian limestones overlying the BL (O’Dogherty,994 and this study, sample BTT 572). It is also reported fromampanian sediments of DSDP Leg 20 (Foreman, 1973).
Material: Three specimens.
Acanthocircus hueyi (Pessagno, 1976)Plate 1, Figs. 5–91976. Spongosaturninus hueyi n. sp. – Pessagno, p. 39, Pl. 12,
Description: Form bearing a subrectangular ring which pos-esses at least six small peripheral spines and displays a slightlyeveloped internal blade.
Material: One specimen.
Acanthocircus horridus Squinabol, 1903bPlate 1, Figs. 11 and 121903b. Acanthocircus horridus n. sp. – Squinabol, p. 125,
l. 9, Fig. 3.1994. Acanthocircus horridus Squinabol – O’Dogherty,
. 253, Pl. 44, Figs. 1–6 (and entire synonymy).2003. Mesosaturnalis horridus (Squinabol) – Yurtsever et al.,
ext-Fig. 6H.2004. Acanthocircus horridus Squinabol – Bragina, p. S445,
l. 27, Figs. 4 and 9.Occurrence: In the UMB, A. horridus is reported from
iddle Albian to Upper Cenomanian levels (U.A. 12–18 of’Dogherty, 1994). During this study, this species was found toccur in limestones overlying the BL (Lower Turonian), extend-ng thus its currently known age range. The species is alsoeported from Middle–Upper Cenomanian levels of northernurkey (Yurtsever et al., 2003; Bragina, 2004).
Material: Three specimens.
Acanthocircus irregularis Squinabol, 1903bPlate 1, Fig. 131903b. Acanthocircus irregularis n. sp. – Squinabol, p. 125,
l. 9, Fig. 6.1994. Acanthocircus sp. EJ1 – Erbacher, p. 90, Pl. 17, Figs. 4
. 264, Pl. 46, Figs. 17–25 (and entire synonymy).Occurrence: This species is known from Middle Albian to
pper Cenomanian levels of the UMB (U.A 11–18, O’Dogherty,994). During this study, the species was found only in oneample (BG 54) dated as Middle Cenomanian.
Material: One specimen.
Acanthocircus tympanum O’Dogherty, 1994Plate 1, Figs. 14 and 151986. Acanthocircus sp. nov. – Kuhnt et al., Pl. 8, Fig. M.1994. Mesosaturnalis sp. EJ2 – Erbacher, p. 102, Pl. 19, Figs.
and 7.1994. Acanthocircus tympanum nov. sp. – O’Dogherty,
. 259, Pl. 45, Figs. 17–24.1999. Acanthocircus tympanum O’Dogherty – Khan et al.,
t al., Text-Fig. 6I.2004. Acanthocircus tympanum O’Dogherty – Bragina,
. S447, Pl. 34, Fig. 15. P
cropaleontologie 50 (2007) 253–287
Occurrence: In the UMB, this species is common in theower Turonian limestones overlying the BL (O’Dogherty, 1994nd this study). However, it is also reported from the top of theL of the southern Alps (Salvini and Marcucci Passerini, 1998).therwise, it is also reported from Lower Turonian levels oforocco (Kuhnt et al., 1986), Turkey (Yurtsever et al., 2003)
nd the Crimean Mountains (Bragina, 2004).Material: Three specimens.
Acanthocircus venetus (Squinabol, 1914)Plate 1, Figs. 16 and 171914. Saturnalis venetus n. f. – Squinabol, p. 269, 299,
l. 20.1, Fig. 2; Pl. 24.5, Fig. 1.1973. Spongosaturnalis (?) sp. – Foreman, p. 261, Pl. 14, Figs.
and 5; non Figs. 6, 7, 8 (=A. bestiarius), 9 (=A. dendroacanthos)nd Pl. 15, Fig. 2 and 3.
1994. Mesosaturnalis praeclarus (Foreman) – Erbacher,. 102, Pl. 19, Fig. 5.
2004. Vitorfus brustolensis (Squinabol) – Bragina, p. S447,l. 28, Figs. 6, 8; Pl. 34, Fig. 13.
Occurrence: In the UMB, this species is known from Upperlbian to Lower Turonian levels (Erbacher, 1994; O’Dogherty,994). During this study, V. brustolensis was found only inne sample (BG 54, Middle Cenomanian). It is also reportedrom Middle Cenomanian strata of northern Turkey and fromhe Lower Turonian of the Crimean Mountains (Bragina,004).
Material: One specimen.
Vitorfus minimus (Squinabol, 1914)Plate 1, Fig. 201914. Saturnalis minimus n. f. – Squinabol, p. 287, Pl. 23[4],
igs. 6 and 6a.1944. Saturnalis lateralis n. sp. – Campbell and Clark, p. 6,
l. 1, Figs. 7, 10, 11, 13, 15.1994. Vitorfus brustolensis (Squinabol) – Marcucci et al.,
l. 1, Fig. 11.1994. Vitorfus minimus (Squinabol) – O’Dogherty, p. 266,
l. 47, Figs. 4–7.1999. Vitorfus minimus (Squinabol) – Khan et al., Pl. 2,
ig. j.Occurrence: During this study, this species was found only
n the Lower Turonian limestones overlying the BL (sampleTT 567B). However, in the UMB, V. minimus is known frompper Albian to Lower Turonian levels (Marcucci et al., 1994;’Dogherty, 1994). It is also reported from Cenomanian to Tur-nian strata of Spain (Khan et al., 1999).
Material: One specimen.
Vitorfus morini Empson-Morin, 1981Plate 1, Fig. 211981. Vitorfus morini n. sp. – Empson-Morin, p. 261, Pl. 4,
igs. 7a–8d.1994. Vitorfus morini Empson-Morin – Marcucci et al., Pl.
, Fig. 10.1994. Vitorfus morini Empson-Morin – O’Dogherty, p. 267,
l. 47, Figs. 12–15 (and entire synonymy).1998. Vitorfus morini Empson-Morin – Salvini and Marcucci
ig. 6K.2004. Vitorfus morini Empson-Morin – Bragina, p. S449,
l. 34, Fig. 8.2004. Vitorfus morini Empson-Morin – Scopelliti et al., Text-
ig. 8(14).Occurrence: Lower Turonian limestones overlying the BL
f UMB (O’Dogherty, 1994 and this study). V. morini is notound so far within the BL of the UMB, but it is reported byalvini and Marcucci Passerini (1998) in the BL of the South-rn Alps. Outside Italy, this species is also known to occur in
p
a
cropaleontologie 50 (2007) 253–287 265
ower Turonian levels of Turkey (Yurtsever et al., 2003) andn Lower Turonian strata of the Crimean Mountains (Bragina,004).
Archaeodictyomitra crassispina (Squinabol, 1903b)Plate 2, Fig. 11903b. Diplostrobus crassispina n. sp. – Squinabol, p. 140,
l. 8, Fig. 37.1986. Mita gracilis (Squinabol) – Kuhnt et al., Pl. 7, Fig. O.1994. Dictyomitra crassispina (Squinabol) – O’Dogherty,
. 79, Pl. 4, Figs. 2–7.2001. Archaeodictyomitra crassispina (Squinabol) – De
ever et al., Text-Fig. 173.4.Occurrence: In the UMB, this species is reported in
pper Cenomanian strata underlying the BL (U.A. 17–18 of’Dogherty, 1994). During the present study, A. crassispinaas found in the lowermost part of the BL. The species is also
eported from Upper Cenomanian strata of Morocco (Kuhnt etl., 1986).
Material: Two specimens.
Archaeodictyomitra sp. cf. A. crebrisulcata (Squinabol,904)
Plate 2, Fig. 2cf. 1904. Dictyomitra crebrisulcata n. sp. – Squinabol, p. 231,
l. 10, Fig. 1.cf. 1994. Dictyomitra crebrisulcata Squinabol – O’Dogherty,
. 75, Pl. 2, Figs. 12–17.Occurrence: This species is reported from Upper Cenoma-
ian to Lower Turonian of the UMB (U.A. 17–20 of O’Dogherty,994). During this study, few poorly preserved specimens werenly found within the BL.
Material: Four specimens.
Archaeodictyomitra montisserei (Squinabol, 1903b)Plate 2, Fig. 31903b. Stichophormis Montis Serei n. sp. – Squinabol, p. 137,
l. 8, Fig. 38.cf. 1977b. Zifondium lassensis n. sp. – Pessagno, p. 47, Pl. 7,
Occurrence: In the UMB, this species is reported through-ut the Middle Albian to Lower Turonian (O’Dogherty, 1994nd this study). It is also reported from Cenomanian strata oforthern Turkey (Bragina, 2004).
Occurrence: In the UMB, this species is reported fromower to Upper Cenomanian strata underlying the BL (U.A.6–18 of O’Dogherty, 1994). During the present study, A. tur-itum was found at the base of the BL.
de mi
P
p
p
O(
n
p
P
M
onTat
P
P
P
P
F
O
F
a
M
pF
M
U
tMnoMw
F
F
p
F
b
P
F
inkif(
F
P
P
b
P
nF
a
B. Musavu-Moussavou et al. / Revue
Material: One specimen.
Archaeodictyomitra simplex Pessagno, 1977bPlate 2, Fig. 51977b. Archaeodictyomitra simplex n. sp. – Pessagno, p. 43,
’Dogherty, p. 77, Pl. 3, Figs. 1–4, 6–10, 16, 17, 24only).
?1994. Archaeodictyomitra sp. cf. A. sliteri Pessagno – Pig-otti, Pl. 1, Fig. 17.
2004. Archaeodictyomitra simplex Pessagno – Bragina,. S370, Pl. 7, Figs. 12 and 14.
2004. Dictyomitra montisserei Pessagno – Bragina, p. S370,l. 8, Figs. 2 and 3.
2006. Archaeodictyomitra simplex Pessagno – Musavu-oussavou and Danelian, p. 149, Pl. 1, Figs. 1–3.Occurrence: Albian of California (Pessagno, 1977b), Albian
f Japan (Tumanda, 1989), Middle–Upper Cenomanian oforthern Turkey (Bragina, 2004), Upper Cenomanian-Loweruronian of Demerara Rise, ODP Leg 207 (Musavu-Moussavound Danelian, 2006). During this study, A. simplex was found inhe lower part of the BL and underlying limestones.
Material: Approximately 60 specimens.
Archaeodictyomitra sliteri Pessagno, 1977bPlate 2, Figs. 6–81977b. Archaeodictyomitra sliteri n. sp. – Pessagno, p. 44,
l. 6, Figs. 3, 4, 22, 23, 27.1989. Archaeodictyomitra sliteri Pessagno – Tumanda, p. 36,
l. 7, Fig. 2.1991. Archaeodictyomitra sliteri Pessagno – Marcucci
asserini et al., Text-Figs. 3n-o.1994. Archaeodictyomitra sliteri Pessagno – Erbacher, p. 93,
l. 20, Fig. 3.1994. Archaeodictyomitra sp. – Munasri and Bambang Text-
. S372, Pl. 7, Figs. 6, 10, 11; Pl. 8, Figs. 8–10, 12; Pl. 34,ig. 6.
2006. Archaeodictyomitra sliteri Pessagno – Musavu-oussavou and Danelian, p. 149, Pl. 1, Fig. 11.Occurrence: Cenomanian of California (Pessagno, 1977b);
pper Albian to Lower Turonian of Italy and Upper Albian
o(U
cropaleontologie 50 (2007) 253–287 267
o Cenomanian of the northern Atlantic (Erbacher, 1994);iddle–Upper Cenomanian of northern Turkey; Lower Turo-
ian of Crimean Mountains (Bragina, 2004); Upper Cenomanianf the central Atlantic (Demerara Rise, ODP Leg 207, Musavu-oussavou and Danelian, 2006). During this study, A. sliterias found throughout the BL.Material: More than 150 specimens.
Genus Dictyomitra Zittel, 1876Type species: Dictyomitra multicostata Zittel, 1876
Dictyomitra formosa Squinabol, 1904Plate 2, Figs. 9–111904. Dictyomitra formosa n. sp. – Squinabol, p. 232, Pl. 10,
ig. 4.1973. Dictyomitra torquata Foreman – Foreman, Pl. 15,
l. 4, Figs. 8–12 (and entire synonymy).1994. Dictyomitra formosa Squinabol – Pignotti, Pl. 2,
igs. 5, 16.Occurrence: In this study, species D. formosa was only found
n lower–middle part of the BL and the underlying Cenoma-ian limestones. However, in other studies from the UMB it isnown to occur throughout the Late Albian to Early Turoniannterval (O’Dogherty, 1994; Pignotti, 1994). It is also reportedrom Lower Coniacian to Lower Campanian strata of CaliforniaPessagno, 1976).
Material: 35 specimens.
Dictyomitra multicostata Zittel, 1876Plate 2, Figs. 12 and 131876. Dictyomitra multicostata n. sp. – Zittel, p. 81, Pl. 2,
igs. 2–4.1976. Dictyomitra multicostata Zittel – Pessagno, p. 52,
l. 14, Figs. 4–9.1994. Dictyomitra multicostata Zittel – O’Dogherty, p. 82,
l. 4, Figs. 17–19 (and entire synonymy).1994. Dictyomitra multicostata Zittel – Munasri and Bam-
ang, Text-Figs. 7.3–7.5.1998. Dictyomitra multicostata Zittel – Erbacher, p. 369,
l. 1, Fig. 5.1998. Dictyomitra ex. gr. multicostata Zittel – Vish-
evskaya and De Wever, Pl. 2, Figs. 22–25; Pl. 3,ig. 17–19.
2006. Dictyomitra multicostata Zittel – Musavu-Moussavound Danelian, p. 151, Pl. 1, Fig. 6.
Occurrence: In the UMB, this species is known to firstccur in the Lower Turonian limestones overlying the BLO’Dogherty, 1994 and this study). It is also reported frompper Turonian to Santonian levels of the equatorial Atlantic
2 de mi
(f(
P
a
Dm
F
F
P
F
F
P
a
P
F
P
f1cM(A
P
F
3
rOw
F
F
P
F
F
F
a
P
1
P
P
T
MOfaI(P2
68 B. Musavu-Moussavou et al. / Revue
Erbacher, 1998; Musavu-Moussavou and Danelian, 2006) androm Middle Campanian to Mastrichtian strata of CaliforniaPessagno, 1976).
Material: Five specimens.
Dictyomitra sp. cf. D. densicostata Pessagno, 1976Plate 2, Figs. 14 and 15cf. 1976. Dictyomitra densicostata n. sp. – Pessagno, p. 51,
l. 14, Figs. 10–14.cf. 1998. Dictyomitra densicostata Pessagno – Vishnevskaya
nd De Wever, Pl. 3, Fig. 20.Remarks: These specimens are doubtfully assigned to
. densicostata because of their reduced height (200 �m inaximum).Material: Three specimens.
Genus Mita Pessagno, 1977bType species: Mita magnifica Pessagno, 1977b
Mita gracilis (Squinabol, 1903b)Plate 2, Figs. 16 and 171903b. Sethoconus gracilis n. sp. – Squinabol, p. 131, Pl. 10,
ig. 13.1982. Mita gracilis (Squinabol) – Taketani, p. 60, Pl. 5,
igs. 2a and 2b; Pl. 12, Fig. 3.1982. Mita regina (Campbell and Clark) – Taketani, p. 60,
l. 5, Figs. 3a and 3b.1988. Mita gracilis (Squinabol) – Thurow, p. 402, Pl. 3,
ig. 2.1994. Mita gracilis (Squinabol) – Erbacher, p. 102, Pl. 9,
igs. 10 and 11.1994. Dictyomitra gracilis (Squinabol) – O’Dogherty, p. 73,
l. 1, Figs. 12–25.1994. Mita gracilis (Squinabol) - Urquhart, Text-Figs. 5.8
nd 5.14.1996. Dictyomitra gracilis (Squinabol) – Gallicchio et al.,
l. 1, Fig. 4.1998. Mita gracilis (Squinabol) – Erbacher, p. 370, Pl. 1,
ig. 13.non 2004. Thanarla gracilis (Squinabol) – Bragina, p. S369,
l. 7, Fig. 9.Occurrence: In the UMB, this species was known previously
rom Lower Albian to Middle Cenomanian strata (Erbacher,994; O’Dogherty, 1994). During this study, we found M. gra-ilis at the base of the BL. The species is also reported fromiddle to Upper Cenomanian levels of the equatorial Atlantic
Erbacher, 1998) and from lower Albian levels of the northerntlantic (Erbacher and Thurow, 1998).Material: 13 specimens.
Genus Thanarla Pessagno, 1977bType species: Phormocyrtis veneta Squinabol, 1903b
Thanarla brouweri (Tan, 1927)Plate 2, Fig. 18 F
cropaleontologie 50 (2007) 253–287
1927. Eucyrtidium Brouweri spec. nov. typ. – Tan, p. 58,l. 11, Figs. 89a and 89b.
1994. Thanarla brouweri (Tan) – O’Dogherty, p. 86, Pl. 5,igs. 1–12 (and entire synonymy).
2000. Thanarla conica (Aliev) – Mekik Pl. 5, Fig. 28.2003. Thanarla brouweri (Tan) – Ziabrev et al. Figs.
–43.Occurrence: In UMB, this species is reported from Bar-
emian (and older) to middle Albian strata (U.A. 1–11 of’Dogherty, 1994). It is the first time that it is reported fromithin the BL.Material: One specimen.
Thanarla pulchra (Squinabol, 1904)Plate 2, Figs. 19 and 201904. Sethamphora pulchra n. sp. – Squinabol, p. 213, Pl. 5,
ig. 2.7.1976. Lithocampe (?) elegantissima Cita – Pessagno, p. 55,
l. 3, Fig. 6.1982. Thanarla pulchra (Squinabol) – Taketani, p. 59, Pl. 11,
ig. 19.1982. Thanarla elegantissima (Cita) – Taketani, p. 59, Pl. 11,
igs. 17 and 18.1989. Thanarla pulchra (Squinabol) – Tumanda, p. 40, Pl. 3,
ig. 17.1991. Thanarla elegantissima (Cita) – Marcucci Passerini et
l., Text-Figs. 3a and 3b.1994. Thanarla pulchra (Squinabol) – O’Dogherty, p. 91,
l. 5, Figs. 28–33 (and entire synonymy).1995. Thanarla elegantissima (Cita) – Bak, p. 21, Fig-text.
0k, l.1998. Thanarla pulchra (Squinabol) – Salvini and Marcucci
asserini, Text-Fig. 6u.1998. Thanarla elegantissima (Cita) – Salvini and Marcucci
asserini, Text-Fig. 6t.1999. Thanarla pulchra (Squinabol) – Bak, Pl. 1, Fig. 9.2002. Thanarla pulchra (Squinabol) – Suzuki et al., p. 57,
ext-Figs. 2 and 3.Occurrence: In the UMB, this species is reported from
iddle Albian to Upper Cenomanian strata (U.A. 11–18 of’Dogherty, 1994; this study). However, it is also reported
rom within the BL by Marcucci Passerini et al., 1991; Salvinind Marcucci Passerini, 1998, as was the case of this study.t is also reported from the Lower Cenomanian of CaliforniaPessagno, 1976), the Middle Albian to Upper Cenomanian ofoland (Bak, 1999) and the Albian of Colombia (Suzuki et al.,002).
Material: 23 specimens.
Thanarla veneta (Squinabol, 1903b)Plate 2, Figs. 21 and 221903b. Phormocyrtis veneta n. sp. – Squinabol, p. 134, Pl. 9;
2004. Thanarla veneta (Squinabol) – Scopelliti et al.,ig. 8(13).
2004. Thanarla veneta (Squinabol) – Bragina, p. S369, Pl. 7,igs. 7 and 8; Pl. 33 Fig. 11.
Occurrence: In the UMB, this species is known from Middlelbian to Lower Turonian levels (Erbacher, 1994; O’Dogherty,994). More precisely, during this study, T. veneta was found toccur last in the upper part of the BL, while Erbacher (1994)ound it also above the critical interval. This species is alsoeported from the Upper Cenomanian of northern Turkey, theower Turonian of the Crimean Mountains (Bragina, 2004) and
rom Santonian strata of Russia (Vishnevskaya and De Wever,998).
Material: 15 specimens.
Family CANNOBOTRYIDAE Haeckel, 1881Genus Rhopalosyringium Campbell and Clark, 1944Type species: Rhopalosyringium magnificum Campbell and
lark, 1944
Rhopalosyringium elegans (Squinabol, 1903b)Plate 3, Fig. 11903b. Lychnocanium elegans n. sp. – Squinabol, p. 130,
l. 8, Fig. 34.1994. Rhopalosyringium elegans (Squinabol) – O’Dogherty,
. 166, Pl. 23, Figs. 1–6.2003. Rhopalosyringium elegans (Squinabol) – Yurtsever et
l., Text-Fig. 7.P.Occurrence: In the UMB, R. elegans is known from Cenoma-
ian limestones underlying the BL (U.A. 17–19 of O’Dogherty,994). During this study, the species was found in only oneample (BL 1/02) coming from the base of the BL. It is alsoeported from Cenomanian strata of Turkey (Yurtsever et al.,003).
Material: Three specimens.
Rhopalosyringium majuroensis Schaaf, 1981Plate 3, Fig. 21981. Rhopalosyringium majuroensis n. sp. – Schaaf, p. 437,
l. 6, Figs. 2 and 3; Pl. 23, Fig. 5.
cropaleontologie 50 (2007) 253–287 269
1994. Rhopalosyringium mosquense (Smirnova and Aliev) -’Dogherty, p. 165, Pl. 22, Figs. 1–6.1998. Rhopalosyringium majuroensis Schaaf – Salvini and
oussavou and Danelian, p. 154, Pl. 2, Fig. 1.Occurrence: During this study, this species was found in
nly one sample (BL 1/02) coming from the base of the BL.ut in the UMB, R. majuroensis (=R. mosquense) is known
hroughout the Albian to uppermost Cenomanian (U.A. 10–19f O’Dogherty, 1994; Salvini and Marcucci Passerini, 1998). It islso known from Middle–Upper Cenomanian levels of northernurkey (Bragina, 2004) and Upper Cenomanian sediments of
he equatorial Atlantic (ODP Leg 207, Musavu-Moussavou andanelian, 2006).Material: Two specimens.
. 168, Pl. 23, Figs. 12–16.1998. Rhopalosyringium scissum O’Dogherty – Salvini and
arcucci Passerini, Fig. 10l.Occurrence: In the UMB, this species is known to occur in
he Lower Turonian limestones overlying the BL (O’Dogherty,994; this study, sample BTT 566B). However, it is also reportedrom the top of the BL of Southern Alps (Salvini and Marcucciasserini, 1998).
Material: Two specimens.
Family CARPOCANIIDAE Haeckel, 1881 emend. Riedel,967
Diacanthocapsa ovoidea Dumitrica, 1970Plate 3, Fig. 41970. Diacanthocapsa ovoidea n. sp. – Dumitrica, p. 63,
l. 5, Figs. 25a and 25b; Pl. 6, Figs. 26–29b.1982. Diacanthocapsa brevithorax Dumitrica – Taketani,
. 68, Pl. 8, Figs. 1a and 1b.1994. Diacanthocapsa ovoidea Dumitrica – O’Dogherty,
. 220, Pl. 37, Figs. 1–6 (and entire synonymy).1998. Diacanthocapsa ovoidea Dumitrica – Salvini and Mar-
ucci Passerini, Text-Fig. 9o.Occurrence: In the UMB, D. ovoidea is reported from the
iddle Cenomanian to Lower Turonian interval (O’Dogherty,994; Salvini and Marcucci Passerini, 1998). During this study,t was found in only one sample (BL 29/02) coming from the BL.
Material: Three specimens.
Genus Guttacapsa O’Dogherty, 1994Type species: Halicapsa gutta Squinabol, 1903b
270 B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287
Occurrence: In the UMB, this species is known to occur inhe Middle–Upper Cenomanian limestones below the BL (U.A.
de mi
11pfA
F
P
F
t1fts
1
a
F
F
P
T
T
F
iofaLS
F
F
P
(iB
P
p
c
U1P
P
p
P
M
bsrpDin
B. Musavu-Moussavou et al. / Revue
7–19 of O’Dogherty, 1994; Salvini and Marcucci Passerini,998). During this study, G. biacuta was found in only one sam-le (BL 1/02) coming from the base of the BL. It is also reportedrom Middle–Upper Cenomanian sediments of the equatorialtlantic (Erbacher, 1998).Material: Two specimens.
Type species: Guttacapsa gutta (Squinabol, 1903b)Plate 3, Fig. 61903b. Cenellipsis biacutus n. sp. – Squinabol, p. 116, Pl. 8,
ig. 24.1994. Guttacapsa gutta (Squinabol) – O’Dogherty, p. 227,
l. 38, Figs. 1–6.1998. Guttacapsa gutta (Squinabol) – Erbacher, p. 369, Pl. 1,
ig. 8.Occurrence: In the UMB, this species is known to occur in
he Middle–Upper Cenomanian limestones below the BL (U.A.7–19 of O’Dogherty, 1994). During this study, G. gutta wasound in only one sample (BL 1/02) coming from the base ofhe BL. It is also reported from Upper Turonian to Santonianediments of the equatorial Atlantic (Erbacher, 1998).
Eostichomitra bonum (Kozlova) in Kozlova and Gorbovetz,966
Plate 4, Figs. 1 and 21986. Xitus sp. – Kuhnt et al., Pl. 7, Fig. y.1994. Xitus (?) sp. EJ2 – Erbacher, p. 118, Pl. 19, Figs. 13
nd 14.1994. Xitus (?) bonarelli n. sp. – Marcucci et al., p. 30, Pl. 2,
ig. 3.1994. Stichomitra sp. – Munasri and Bambang, Text-
ig. 7.11.1994. Eostichomitra bonum (Kozlova) – O’Dogherty, p. 151,
l. 18, Figs. 16–24 (and entire synonymy).1998. Xitus (?) bonarelli – Salvini and Marcucci Passerini,
ext-Fig. 10u.2003. Eostichomitra bonum (Kozlova) – Yurtsever et al.,
ext-Fig. 7L.2004. Eostichomitra bonum (Kozlova) – Scopelliti et al.,
ig. 8.8.Occurrence: In the UMB, this species was known to occur
n the Lower Turonian limestones overlying the BL (U.A. 20–21f O’Dogherty, 1994). However, during this study, it also wasound to occur in the upper part of the BL (sample BL 37/02),s well as in the overlying limestones. It is also reported fromower Turonian levels of Turkey (Yurtsever et al., 2003) andicily (Scopelliti et al., 2004).
Material: 15 specimens.
Genus Phalangites O’Dogherty, 1994Type species: Phalangites calamus O’Dogherty, 1994
cropaleontologie 50 (2007) 253–287 271
Phalangites telum O’Dogherty, 1994Plate 4, Figs. 3 and 41994. Phalangites telum nov. sp. – O’Dogherty, p. 157, Pl. 20,
igs. 16–23.1996. Phalangites telum O’Dogherty – Gallicchio et al., Pl. 1,
ig. 13.1998. Phalangites telum O’Dogherty – Salvini and Marcucci
asserini, Fig. 10h.Occurrence: Middle Albian to Lower Turonian of UMB
O’Dogherty, 1994; Salvini and Marcucci Passerini, 1998). Dur-ng this study, P. telum was only reported from the top of theL.
Material: Nine specimens.
Genus Pseudoeucyrtis Pessagno, 1977aType species: Eucyrtis (?) zhamoidai Foreman, 1973
Pseudoeucyrtis pulchra (Squinabol, 1904)Plate 4, Fig. 51904. Theosyringium pulchrum n. sp. – Squinabol, p. 222,
l. 8, Fig. 7.1994. Pseudoeucyrtis pulchra (Squinabol) – O’Dogherty,
. 184, Pl. 27, Figs. 9–13 (and entire synonymy).1998. Pseudoeucyrtis pulchra (Squinabol) – Salvini and Mar-
ucci Passerini, Text-Fig. 10j.Occurrence: In the UMB, this species is reported from the
pper Cenomanian to Lower Turonian interval (O’Dogherty,994; Salvini and Marcucci Passerini, 1998). During this study,. pulchra was found only in the lower part of the BL.
Material: Two specimens.
Pseudoeucyrtis sp. cf. P. spinosa (Squinabol, 1903b)Plate 4, Fig. 6cf. 1903b. Eusyringium spinosum n. sp. – Squinabol, p. 141,
l. 8, Fig. 42.cf. 1994. Pseudoeucyrtis spinosa (Squinabol) – O’Dogherty,
. 183, Pl. 27, Figs. 1–8 (and entire synonymy).cf. 1994. Eusyringium spinosum Squinabol – Erbacher, p. 99,
l. 16, Figs. 4 and 5.cf. 1998. Pseudoeucyrtis spinosa (Squinabol) – Salvini and
arcucci Passerini, Text-Fig. 10k.Occurrence: In the UMB, this species is known to occur
elow the BL (U.A. 11–19 of O’Dogherty, 1994). The fewpecimens found at the base of the BL (sample BL 1/02)esemble to P. spinosa by the subspherical shape of theostabdominal segment and traces of spines on its surface.oubtful identification is due to poor preservation. The species
s also reported from Middle to Upper Albian sediments of theorthern Atlantic (Erbacher, 1994).
Material: Two specimens.
Genus Stichomitra Cayeux, 1897
Type species: Stichomitra bertrandi Cayeux, 1897
Stichomitra communis Squinabol, 1903bPlate 4, Figs. 7 and 8
272 B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287
2006. Stichomitra communis Squinabol – Musavu-oussavou and Danelian, p. 155, Pl. 32 Figs. 14–16.Occurrence: In the UMB, S. communis is known from
he Middle Aptian to Lower Turonian (O’Dogherty, 1994;alvini and Marcucci Passerini, 1998, this study). It is alsoeported from the Cenomanian to Lowermost Coniacian ofapan (Taketani, 1982), from the Upper Albian to Loweruronian of the equatorial Atlantic (Erbacher, 1998; Musavu-oussavou and Danelian, 2006) and from the Cenomanian
o Turonian of Turkey (Yurtsever et al., 2003; Bragina,004).
Material: Approximately 100 specimens.
Stichomitra stocki (Campbell and Clark, 1944)Plate 4, Figs. 9–121944. Stichocapsa (?) stocki n. sp. – Campbell and Clark,
. 44, Pl. 8, Figs. 31–33.1986. Amphipyndax stocki (Campbell and Clark) – Kuhnt et
l., Pl. 7, Fig. u.1994. Stichomitra stocki (Campbell and Clark) – O’Dogherty,
. 146, Pl. 18, Figs. 10–15.1998. Amphipyndax stocki (Campbell and Clark) – Vish-
evskaya and De Wever, p. 255, Pl. 2, Fig. 21.1998. Stichomitra stocki (Campbell and Clark) – Erbacher,
. 370, Pl. 1, Fig. 14 non Pl. 2, Fig. 1 (=S. mediocris).2004. Amphipyndax stocki (Campbell and Clark) – Bragina,
. S375, Pl. 9, Fig. 11 (only); Pl. 32, Fig. 8.2006. Stichomitra stocki (Campbell and Clark) – Musavu-
oussavou and Danelian, p. 155, Figs. 11–13.Occurrence: This species is recorded from the Upper Ceno-
anian to Lower Turonian of the UMB (O’Dogherty, 1994;his study). It is also reported from the Uppermost Cenoma-
a
A1
cropaleontologie 50 (2007) 253–287 273
ian to lowermost Turonian of Morocco (Kuhnt et al., 1986),rom the Albian to Turonian, Santonian of Russia (Vishnevskayand De Wever, 1998); from the Upper Cenomanian of Demer-ra Rise, ODP leg 207 (Musavu-Moussavou and Danelian,006).
Material: Five specimens.Stichomitra tosaensis Nakaseko and Nishimura in Nakaseko
t al., 1979Plate 4, Fig. 131979. Stichomitra tosaensis n. sp. – Nakaseko and Nishimura,
. 24, Pl. 7, Fig. 11.1994. Stichomitra sp. EJ1 – Erbacher, p. 114, Pl. 15, Fig. 2;
l. 19, Fig. 10.1994. Stichomitra tosaensis Nakaseko and Nishimura –
’Dogherty, p. 146, Pl. 18, Figs. 1–8.Occurrence: In the UMB, this species is known to occur
hroughout the Lower Albian to Lower Turonian intervalErbacher, 1994; O’Dogherty, 1994; this study).
Sciadiocapsa sp. cf. S. monticelloensis (Pessagno, 1969)Plate 3, Fig. 7cf. 1969. Microsciadocapsa monticelloensis n. sp. – Pes-
agno, p. 407, Pl. 32, Figs. 3–9; Pl. 34, Figs. 1 and 2.cf. 1994. Sciadiocapsa monticelloensis (Pessagno) –
’Dogherty, p. 230, Pl. 39, Figs. 9–12 (and entire synonymy).cf. 2004. Sciadiocapsa monticelloensis (Pessagno) – Scopel-
iti et al., Text-Fig. 8(12).Remarks: Doubtful identification is due to incomplete
reservation of the proximal part of the test.Occurrence: In the UMB, this species occurs only in the
ower Turonian limestones overlying the BL (O’Dogherty,994). However, it is also reported from the Uppermost Ceno-anian levels of Sicily (Scopelliti et al., 2004).Material: Two specimens.
Pignotti, Pl. 1, Fig. 12.1998. Pseudodictyomitra pseudomacrocephala (Squinabol)
Salvini and Marcucci Passerini, Text-Fig. 8h.1999. Pseudodictyomitra pseudomacrocephala (Squinabol)
Bak, Pl. 1, Fig. 12.2003. Pseudodictyomitra pseudomacrocephala (Squinabol)
Yurtsever et al., Text-Fig. 7N.2004. Pseudodictyomitra pseudomacrocephala (Squinabol)
Scopelliti et al., Fig. 8(11).2004. Pseudodictyomitra pseudomacrocephala (Squinabol)
Bragina, p. S367, Pl. 7, Fig. 4; Pl. 32, Figs. 9, 14–16.Occurrence: In the UMB, this species is known to occur in
he Lower Albian to Lower Turonian levels, including the BLMarcucci Passerini et al., 1991; O’Dogherty, 1994; this study).t is also reported from the Middle Albian to Upper Cenomanianf Poland (Bak, 1999), from the Middle Cenomanian to Loweruronian of Turkey (Yurtsever et al., 2003; Bragina, 2004) androm the Upper Cenomanian to Lower Turonian of the Crimean
ountains (Bragina, 2004).Material: More than 100 specimens.
Pseudodictyomitra tiara (Holmes, 1900)Plate 3, Figs. 11 and 121900. Dictyomitra tiara sp. nov. – Holmes, p. 701, Pl. 38,
Occurrence: In the UMB, this species is recorded in Ceno-anian strata (i.e., O’Dogherty, 1994; Salvini and Marcucciasserini, 1998; this study). It last occurs within the middle partf the BL (Salvini and Marcucci Passerini, 1998; this study, sam-le BL 16/02). It is also known from the Cenomanian of TurkeyYurtsever et al., 2003; Bragina, 2004).
Material: 56 specimens.
Family SYRINGOCAPSIDAE Foreman, 1973Genus Distylocapsa Squinabol, 1904Type species: Distylocapsa nova Squinabol, 1904
Distylocapsa sp. cf. D. squama O’Dogherty, 1994Plate 3, Fig. 131994. Distylocapsa squama nov. sp. – O’Dogherty, p. 189,
l. 28, Figs. 16–21.1998. Distylocapsa squama O’Dogherty – Salvini and Mar-
ucci Passerini, Text-Fig. 10s.Occurrence: In the UMB, this species was known to occur in
he Lower Turonian limestones overlying the BL (O’Dogherty,994). The single specimen found during this study comesrom the middle–upper part of the BL (sample BL 29/02). Its doubtfully assigned to D. squama because of its relativelyoor preservation. This species is also known to occur at the topf the BL of the Southern Alps (Salvini and Marcucci Passerini,998).
Material: One specimen.
Genus Dorypyle Squinabol, 1904Type species: Dorypyle cretacea Squinabol, 1904
Dorypyle ovoidea (Squinabol, 1904)Plate 3, Fig. 141904. Lithomespilus ovoideus n. sp. – Squinabol, p. 198,
l. 4, Fig. 8.1994. Dorypyle ovoidea (Squinabol) – O’Dogherty, p. 205,
l. 33, Figs. 1–7.Occurrence: In the UMB, D. ovoidea was known to last
ccur in the Upper Cenomanian limestones underlying the BLO’Dogherty, 1994). In this study, the species was found in oneample (BL 1/02) coming from the base of the BL.
Material: Three specimens.
Genus Squinabollum Dumitrica, 1970Type species: Clistophaena fossile Squinabol, 1903b
de mi
F
F
p
c
P
dafCC
P
F
O
TsU
1uHCSDr
F
e
P
P
F
F
P
P
c
P
LPPA
F
P
c
P
act
F
F
F
F
B. Musavu-Moussavou et al. / Revue
Squinabollum fossile (Squinabol, 1903b)Plate 3, Figs. 15 and 161903b. Clistophaena fossilis n. sp. – Squinabol, p. 130, Pl. 10,
l. 12, Figs. 1–4.Occurrence: In the UMB, this species is recorded from Mid-
le Albian to Lower Turonian strata (O’Dogherty, 1994; Salvinind Marcucci Passerini, 1998; this study). It is also reportedrom the Cenomanian of Romania (Dumitrica, 1975), the Upperenomanian of northern Turkey and the Lower Turonian of therimean Mountains (Bragina, 2004).
’Dogherty, p. 243, Pl. 42, Figs. 10–12 (and entire synonymy).Occurrence: In the UMB, this species is known from Lower
uronian limestones overlying the BL (O’Dogherty, 1994; thistudy, sample BTT 565). However, it is also reported from thepper Albian of California (Pessagno, 1977b).Material: Three specimens.
Family XITIDAE Pessagno, 1977bGenus Xitus Pessagno, 1977bType species: Xitus plenus Pessagno, 1977bRemarks: In the UMB, genus Xitus (sensu O’Dogherty,
994) last occurs in the Upper Cenomanian, and more partic-larly within the BL (Erbacher and Thurow, 1997; this study).owever, it is also reported from Lower Turonian strata of therimean Mountains (Bragina, 2004) and from Coniacian toantonian levels of the Russian platform (Vishnevskaya ande Wever, 1998). Therefore, its last occurrence in the UMB is
egarded as a local extinction.
Xitus mclaughlini (Pessagno, 1977b)Plate 4, Figs. 14 and 151977b. Xitus mclaughlini n. sp. – Pessagno, p. 54, Pl. 9,
non 1998. Novixitus mclaughlini Pessagno – Erbacher, p. 370,l. 2, Fig. 6 (=X. spicularius)
1998. Novixitus mclaughlini Pessagno – Salvini and Mar-ucci Passerini, p. 793, Text-Fig. 6–l.
1999. Xitus mclaughlini (Pessagno) – Bak, Pl. 1, Fig. 10.2004. Novixitus weyli Schmidt-Effing – Bragina, p. S366,
l. 6, Figs. 1, 5–8, 10, 11.Occurrence: In the UMB, this species is reported from
ower Albian to Upper Cenomanian levels (i.e., Marcucciasserini et al., 1991; O’Dogherty, 1994; Salvini and Marcucciasserini, 1998; this study). It is also reported from the Lowerlbian to Upper Cenomanian of Poland (Bak, 1999).Material: 48 specimens.
Xitus sp. cf. X. mclaughlini (Pessagno, 1977b)Plate 4, Fig. 16cf. 1977b. Novixitus mclaughlini – Pessagno, p. 54, Pl. 9,
ig. 17.cf. 1994. Xitus mclaughlini (Pessagno) – O’Dogherty, p. 130,
l. 12, Figs. 14–21.cf. 1998. Novixitus mclaughlini Pessagno – Salvini and Mar-
ucci Passerini, p. 793, Fig. 6-l.cf. 2004. Novixitus weyli Schmidt-Effing - Bragina, p. S366,
l. 6, Figs. 1, 5–8, 10, 11.Remarks: The specimen illustrated here is doubtfully
ssigned to X. mclaughlini because of the rather subcylindri-al shape and poorly developed tubercles on the distal part of itsest.
Material: One specimen.
Xitus spicularius (Aliev, 1965)Plate 4, Figs. 17 and 181965. Dictyomitra spicularia Aliev – Aliev, p. 39, Pl. 6,
ig. 9.1994. Xitus spicularius (Aliev) – O’Dogherty, p. 127, Pl. 11,
igs. 17–31.1994. Xitus spicularius (Aliev) – Erbacher, p. 118, Pl. 3,
ig. 4; Pl. 15, Fig. 7.1998. Xitus spicularius (Aliev) – Erbacher, p. 371, Pl. 1,
igs. 1 and 2.
1998. Novixitus mclaughlini Pessagno – Erbacher, p. 370,
l. 2, Fig. 6.1998. Xitus spicularius (Aliev) – Vishnevskaya and De
ever, p. 258, Pl. 1, Figs. 10, 14, 15.
2 de mi
F
LOorAno
otiab
lrib
P
ps
Tp
F
8
P
P
F
c
a
S
P
LB1BLfZe(a2
1
P
T
3
P
P
1
F
T
76 B. Musavu-Moussavou et al. / Revue
2004. Xitus spicularius (Aliev) – Bragina, p. S362, Pl. 5,igs. 1–6; Pl. 33, Figs. 19, 29, 22.
Occurrence: In the UMB, this species is known fromower Aptian to Upper Cenomanian levels (Erbacher, 1994;’Dogherty, 1994). The present study allows specifying its lastccurrence in the lower part of the BL. The species is alsoeported from the Upper Turonian to Santonian of the equatorialtlantic (Erbacher, 1998), the Middle–Upper Cenomanian oforthern Turkey and the Middle Cenomanian to Lower Turonianf the Crimean Mountains (Bragina, 2004).
Material: 12 specimens.
Xitus sp. APlate 4, Figs. 19–21Description: Test elongated conical bearing an inner layer
f small circular pores and an outer layer composed of smallubercles situated along circumferential ridges. The first ridges more prominent and separates the test into a proximalnd a distal part. Tubercles are interconnected by numerousars.
Material: Three specimens.
Xitus sp. BPlate 4, Fig. 22Description: Test elongated cylindrical, bearing an outer
ayer of three prominent ridges composed of circumferentialows of small tubercles. The first ridge is situated at the prox-mal part of the test, while the two others situated towards thease of the perforated distal part.
Material: One specimen
Genus Torculum O’Dogherty, 1994Type species: Theoconus coronatus Squinabol, 1904
Torculum sp. cf. T. coronatum (Squinabol)Plate 4, Fig. 23cf. 1904. Theoconus coronatus n. sp. – Squinabol, p. 220,
l. 8, Fig. 3.cf. 1994. Torculum coronatum (Squinabol) – O’Dogherty,
. 133, Pl. 12, Figs. 27 and 28; Pl. 14, Figs. 1–29 (and entireynonymy).
Remarks: This morphotype is questionably assigned toorculum coronatum because of the poor preservation of itsroximal part.
Occurrence: In the UMB, this species is reported fromower Albian levels (Erbacher, 1994) and from within theL (up to its upper part; Salvini and Marcucci Passerini,998). During this study, it was found at the base of theL and underlying limestones. It is also reported fromower Maastrichtian levels of California (Foreman, 1968),
rom the Late Valanginian to Middle Albian of the Budvaone (Gorican, 1994), from the Upper Cenomanian of north-rn Turkey and Lower Turonian of the Crimean MountainsBragina, 2004) and from the Upper Cenomanian of Demer-ra Rise, ODP Leg 207 (Musavu-Moussavou and Danelian,006).
Material: 11 specimens.
Genus Holocryptocanium Dumitrica, 1970Type species: Holocryptocanium tuberculatum Dumitrica,
970
Holocryptocanium barbui Dumitrica, 1970Plate 3, Figs. 19 and 201970. Holocryptocanium barbui n. sp. – Dumitrica, p. 76,
l. 17, Figs. 105–108b; Pl. 21, Fig. 136.1975. Holocryptocanium barbui Dumitrica – Dumitrica,
ext-Fig. 2.1.1989. Holocryptocanium barbui Dumitrica – Tumanda, p.
7, Pl. 7, Figs. 20 and 21.1993. Holocryptocanium barbui Dumitrica – Bak p. 197,
l. 4, Fig. 1.1994. Holocryptocanium barbui Dumitrica – Gorican, p. 72,
l. 14, Figs. 10, 13, 14.1994. Holocryptocanium barbui Dumitrica – Erbacher, p.
01, Pl. 5, Fig. 8; Pl. 13, Fig. 11.1994. Holocryptocanium barbui Dumitrica – Pignotti, Pl. 2,
igs. 8 and 17.1995. Holocryptocanium barbui Dumitrica – Bak, p. 16,
ext-Figs. 12k-I.
1998. Holocryptocanium barbui Dumitrica – Salvini and
2001. Holocryptocanium barbui Dumitrica – Bak et al. Text-igs. 17H, I.
2004. Holocryptocanium barbui Dumitrica – Bragina,. S376, Pl. 10, Fig. 12; Pl. 31, Figs. 1 and 2b.
2006. Holocryptocanium barbui Dumitrica – Musavu-oussavou and Danelian, 2006, p. 157, Pl. 3, Fig. 2.Occurrence: In the UMB, this species is known to occur
hroughout the Lower Cenomanian to Lower Turonian inter-al (Erbacher, 1994; Salvini and Marcucci Passerini, 1998).n this study, H. barbui was found only in the lower partf the BL. It is also known from the Cenomanian of Ruma-ia (Dumitrica, 1970), from the Aptian to Upper Turonian ofoland (Bak, 1995, 1999), from the Middle Cenomanian toower Turonian of the Crimean Mountains (Bragina, 2004) and
rom the Upper Cenomanian of the equatorial Atlantic (Demer-ra Rise, ODP Leg 207, Musavu-Moussavou and Danelian,006).
Material: 13 specimens.
Holocryptocanium tuberculatum Dumitrica, 1970Plate 3, Figs. 21 and 221970. Holocryptocanium tuberculatum n. sp. – Dumitrica,
. 75, Pl. 16, Figs. 102, 103a–c; Pl. 21, Figs. 138a and 138b.1975. Holocryptocanium tuberculatum Dumitrica –
umitrica, Text-Fig. 2.2.1977b. Holocryptocanium astiensis n. sp. – Pessagno, p. 40,
l. 1, Fig. 5.Occurrence: In the UMB, this species is known from Upper
enomanian levels (Marcucci Passerini et al., 1991; Salvini andarcucci Passerini, 1998). During this study, it was found in
he lower part of the BL and the underlying limestones. It is alsoeported from the Lower Cenomanian of California (Pessagno,977b) and from Albian to Lower Turonian strata of PolandBak, 1999).
1998. Hemicryptocapsa polyhedra Dumitrica – Salvini andarcucci Passerini, 1998, Fig. 9v.Occurrence: In UMB, this species is reported from the
pper part of the BL (Erbacher, 1994; Salvini and Marcucciasserini, 1998) and the overlying limestones (Erbacher, 1994;’Dogherty, 1994). During this study, it was found in the lowerart of the BL (sample BL 10/02) and its overlying limestonessample BTT 572).
Material: Four specimens.
Order SPUMELLARIA Ehrenberg, 1875Family ANGULOBRACCHIIDAE Baumgartner, 1980
mend. De Wever et al., 2001Genus Paronaella Pessagno, 1971aType species: Paronaella solanoensis Pessagno, 1971a
Paronaella solanoensis Pessagno, 1971aPlate 5, Figs. 1 and 21971a. Paronaella solanoensis n. sp. – Pessagno, p. 48, Pl. 10,
igs. 2 and 3.1994. Paronaella solanoensis Pessagno – O’Dogherty,
. 354, Pl. 66, Figs. 19–24 (and entire synonymy).1998. Paronaella solanoensis Pessagno – Salvini and Mar-
l. 39, Fig. 12.Occurrence: In the UMB, this species is known to occur
rom the Uppermost Cenomanian to the Lower TuronianO’Dogherty, 1994; Salvini and Marcucci Passerini, 1998). Theresence of P. solanoensis in a middle Cenomanian sample (BG-8) of this study extends its known age range in the UMB.his species is also reported from the Upper Turonian to Conia-ian of California (Pessagno, 1971a), from the Middle–Upperenomanian of northern Turkey and from the Upper Cenoma-ian to Lower Turonian of the Crimean Mountains (Bragina,004).
Material: Nine specimens.
Family ARCHAEOSPONGOPRUNIDAE Pessagno, 1973Genus Archaeospongoprunum Pessagno, 1973Type species: Archaeospongoprunum venadoensis
1994. Archaeospongoprunum cortinaensis Pessagno –rbacher, p. 94, Pl. 12, Fig. 2.
1998. Archaeospongoprunum cortinaensis Pessagno –alvini and Marcucci Passerini, Text-Fig. 9b.
2004. Archaeospongoprunum cortinaensis Pessagno – Brag-na, p. S407, Pl. 18, Figs. 5, 10, 11, 13, 14.
2006. Archaeospongoprunum cortinaensis Pessagno –usavu-Moussavou and Danelian, p. 159, Pl. 3, Figs. 11–15.Occurrence: In the UMB, A. cortinaensis is reported from
pper Albian to Lower Turonian levels (Erbacher, 1994). Dur-ng this study, it was found to occur only in the limestonesnderlying and overlying the BL. This species is also knownrom the Lower Cenomanian to Lower Coniacian of CaliforniaPessagno, 1973), from the Uppermost Cenomanian equivalentf the BL of Southern Alps (Salvini and Marcucci Passerini,998), from the Middle–Upper Cenomanian of northern TurkeyBragina, 2004) and from the Lower Turonian of the equatorialtlantic (Demerara Rise, ODP Leg 207, Musavu-Moussavou
. S407, Pl. 35, Fig. 13.Occurrence: This species is reported from the Coniacian
f California (Pessagno, 1976), from the Uppermost Ceno-anian equivalent of the BL of Southern Alps (Salvini andarcucci Passerini, 1998) and from the Lower Turonian of therimean Mountains (Bragina, 2004). As far as we know, it is
he first time that it is reported here from the UMB. Duringhis study, it was found to occur only in one sample (BTT72) coming from the limestones overlying the BL (Loweruronian).
1998. Cavaspongia antelopensis Pessagno – Vishnevskayand De Wever, p. 247, Pl. 3, Fig. 9.
2003. Cavaspongia antelopensis Pessagno – Yurtsever et al.,ext-Fig. 6.L.
2004. Cavaspongia antelopensis Pessagno – Bragina,. S421, Pl. 23, Figs. 9 and 11; Pl. 37, Fig. 7.
Occurrence: In the UMB, the species is known to occur inhe upper part of BL (Salvini and Marcucci Passerini, 1998)nd in the overlying Lower Turonian limestones (O’Dogherty,994). During this study, C. antelopensis was found only inamples overlying the BL. However, it is also reported from
iddle–Upper Cenomanian strata of northern Turkey and Loweruronian levels of the Crimean Mountains (Bragina, 2004).
l. 57, Figs. 8–11.2004. Cavaspongia contracta O’Dogherty – Bragina,
. S421, Pl. 40, Fig. 11.Occurrence: In the UMB, this species is known to occur
hroughout the Lower Cenomanian to Lower Turonian intervalO’Dogherty, 1994 and this study). C. contracta is also knownrom the Upper Cenomanian to Lower Turonian of the Crimean
ountains and from the Upper Cenomanian of northern TurkeyBragina, 2004).
Material: Two specimens.
Cavaspongia euganea (Squinabol, 1904)Plate 5, Fig. 101904. Euchitonia euganea n. sp. – Squinabol, p. 204, Pl. 6,
ig. 1.1976. Dumitrica maxwellensis n. sp. – Pessagno, p. 38, Pl. 4,
igs. 10 and 11.1994. Dumitrica maxwellensis Pessagno – Erbacher, p. 98,
l. 18, Fig. 6.1994. Cavaspongia euganea (Squinabol) – O’Dogherty,
. 309, Pl. 56, Figs. 8–14 (and entire synonymy).1998. Dumitrica maxwellensis Pessagno – Salvini and Mar-
l. 24, Figs. 11 and 12; Pl. 40, Figs. 1 and 8.Occurrence: In the UMB, C. euganea is reported from
iddle Albian to Lower Turonian levels (Erbacher, 1994;’Dogherty, 1994). During this study, it was found only in sam-les coming from limestones underlying the BL. The speciess also known from the Middle–Upper Cenomanian of northernurkey and from the Lower Turonian of the Crimean MountainsBragina, 2004).
2003. Cavaspongia sphaerica O’Dogherty – Yurtsever et al.,ext-Fig. 7.N.
Occurrence: This species is reported from the Upper Albiano Lower Turonian interval of UMB (O’Dogherty, 1994). Duringhis study, C. sphaerica was found in only one sample (BTT 572)oming from limestones overlying the BL. It is also reportedrom the Cenomanian to Lower Turonian of Turkey (Yurtsevert al., 2003).
Material: One specimen.
Family CONOCARYOMMIDAE Lipman, 1969 emend. Deever et al., 2001Genus Praeconocaryomma Pessagno, 1976Type species: Praeconocaryomma universa Pessagno, 1976
Praeconocaryomma lipmanae Pessagno, 1976Plate 5, Figs. 12 and 13?1976. Preconocaryomma lipmanae n. sp. – Pessagno, p. 41,
l. 4, Figs. 12 and 13.1998. Preconocaryomma lipmanae Pessagno – Salvini and
arcucci Passerini, Text-Fig. 8c.Remarks: The specimen illustrated on Fig. 22 is question-
bly assigned to P. lipmanae because of its outline, whichisplays mamae without spines on the top.
Occurrence: In the UMB, this species is known to occurithin the BL (Salvini and Marcucci Passerini, 1998). Dur-
ng this study, P. lipmanae was found in the Lower Turonianimestones overlying the BL, extending thus its known age inhis basin. It is also reported from Upper Cenomanian to Loweruronian levels of California (Pessagno, 1976).
. 340, Pl. 63, Figs. 13–21 (and entire synonymy).1998. Dactyliosphaera lepta (Foreman) – Erbacher, p. 369,
l. 1, Fig. 28.2006. Dactyliosphaera lepta (Foreman) – Musavu-
oussavou and Danelian, p. 160, Pl. 4, Figs. 6 and.
Occurrence: This species is reported from the Late Albiano Late Cenomanian interval of the UMB (O’Dogherty, 1994
F
D
cropaleontologie 50 (2007) 253–287
nd this study). It is also known from the Albian to Cenomanianf the southern Atlantic (Foreman, 1978), from the Late Albiano Late Cenomanian of the northern Atlantic (Erbacher, 1994)nd from the Middle to Upper Cenomanian of the equatorialtlantic (Erbacher, 1998; Musavu-Moussavou and Danelian,006).
Material: Eight specimens.
Dactyliosphaera silviae Squinabol, 1904Plate 5, Fig. 151903b. Gen. and sp. nov. – Squinabol, p. 121, Pl. 9,
ig. 21.1904. Dactyliosphaera silviae n. sp. – Squinabol, p. 196,
l. 4, Fig. 3.1975. Dactyliosphaera silviae Squinabol – Dumitrica, Text-
ig. 2.14.1988. Pseudoaulophacus (?) sp. D – Thurow p. 404, Pl. 5,
l. 25, Figs. 5–8.2006. Dactyliosphaera silviae Squinabol – Musavu-
oussavou and Danelian, p. 160, Pl. 4, Figs. 1–3.Occurrence: In the UMB, this species is known from the
pper Cenomanian limestones underlying the BL (O’Dogherty,994) and from the lower part of the critical interval (Salvini andarcucci Passerini, 1998; this study). D. silviae is also reported
rom the Cenomanian of Roumania (Dumitrica, 1975), fromhe Middle–Upper Cenomanian of northern Turkey (Bragina,004) and from the Upper Cenomanian of the equatorialtlantic (Demerara Rise, ODP Leg 207, Musavu-Moussavou
nd Danelian, 2006).Material: Three specimens.
Family HAGIASTRIDAE Riedel, 1971Genus Crucella Pessagno, 1971aType species Crucella messinae Pessagno, 1971a
Crucella irwini Pessagno, 1971aPlate 5, Fig. 161971a. Crucella irwini n. sp. – Pessagno, p. 55, Pl. 9,
igs. 4–6.1994. Crucella irwini Pessagno – O’Dogherty, p. 369, Pl. 71,
igs. 7–14 (and entire synonymy).1994. Crucella irwini Pessagno – Urquhart, Text-Fig. 5.3.1998. Crucella irwini Pessagno – Vishnevskaya and De
ever, p. 250, Pl. 3, Figs. 11 and 12.
2004. Crucella irwini Pessagno – Bragina, p. S417, Pl. 23,
igs. 4, 5, 7; Pl. 39, Figs 3 and 8.2006. Crucella irwini Pessagno – Musavu-Moussavou and
anelian, p. 160, Pl. 4, Figs. 17 and 18?
de mi
tOof1ptTfM
F
F
P
P
F
F
F
a
oSk1TCmD
F
F
a
F
M
P
P
P
F
P
F
F
F
ml1PotC
e
P
U1
F
F
P
P
P
B. Musavu-Moussavou et al. / Revue
Occurrence: In the UMB, this species is known to occurhroughout the Cenomanian to Lower Turonian (U.A. 16–21 of’Dogherty, 1994). During this study, C. irwini was found innly one Middle Cenomanian sample (BG 54). It is also knownrom the Turonian to Lower Coniacian of California (Pessagno,971a), from the Turonian to Lower Santonian of the Russianlatform (Vishnevskaya and De Wever, 1998), from the Middleo Upper Cenomanian of northern Turkey and from the Loweruronian of the Crimean Mountains (Bragina, 2004) and finallyrom the Upper Cenomanian of the equatorial Atlantic (Musavu-
oussavou and Danelian, 2006).Material: One specimen.
Crucella messinae Pessagno, 1971aPlate 5, Fig. 171971a. Crucella messinae n. sp. – Pessagno, p. 56, Pl. 6,
igs. 1–3.1994. Crucella messinae Pessagno – Erbacher, p. 96, Pl. 2,
ig. 10; Pl. 12, Fig. 3; Pl. 10, Fig. 15; Pl. 16, Fig. 12.1994. Crucella messinae Pessagno – O’Dogherty, p. 368,
l. 70, Figs. 21–24; Pl. 71, Figs. 1–6 (and entire synonymy).1998. Crucella messinae Pessagno – Salvini and Marcucci
nd Danelian, p. 160, Pl. 4, Figs. 20 and 21.Occurrence: In the UMB, this species is reported through-
ut the Middle Albian to Lower Turonian (O’Dogherty, 1994;alvini and Marcucci Passerini, 1998; this study). It is alsonown from the Lower Cenomanian of California (Pessagno,971a), from the Middle–Upper Cenomanian of northernurkey, from the Upper Cenomanian–Lower Turonian of therimean Mountains (Bragina, 2004) and from the Upper Ceno-anian of the equatorial Atlantic (Musavu-Moussavou andanelian, 2006).Material: 45 specimens.
Crucella cachensis Pessagno, 1971aPlate 5, Figs. 18 and 191971a. Crucella cachensis n. sp. – Pessagno, p. 53, Pl. 9,
igs. 1–3.1976. Crucella cachensis Pessagno – Pessagno, p. 31, Pl. 3,
igs. 14 and 15.1991. Crucella cachensis Pessagno – Marcucci Passerini et
l., Text-Figs. 4j–l.1994. Crucella cachensis Pessagno – Erbacher, p. 95, Pl. 18,
ig. 4.
1994. Crucella cachensis tolfaensis Marcucci and Gardin –
arcucci Passerini et al., Pl. 1, Fig. 1.1994. Crucella cachensis Pessagno – O’Dogherty, p. 370,
igs. 7D and 7E.2004. Crucella cachensis Pessagno – Scopelliti et al.,
ig. 8(6).2004. Crucella cachensis Pessagno – Bragina, p. S445, Pl. 22,
ig. 11; Pl. 38, Figs. 8, 10, 11; Pl. 39, Figs. 5, 6, 9.Occurrence: In the UMB, this species is known from the
iddle part of the BL (Uppermost Cenomanian) and the over-ying Lower Turonian limestones (Marcucci Passerini et al.,991; Erbacher, 1994; O’Dogherty, 1994; Salvini and Marcucciasserini, 1998; this study). It is also known from the Turonianf California (Pessagno, 1971a), from the Upper Cenomaniano Lower Turonian of the Crimean Mountains and the Upperenomanian of northern Turkey (Bragina, 2004).
Material: 12 specimens.
Family PARVIVACCIDAE Pessagno, Six and Yang, 1989mend. De Wever et al., 2001
Genus Acaeniotyle Foreman, 1973Type species: Xiphosphaera umbilicata Rust, 1898
Acaeniotyle rebellis O’Dogherty, 1994Plate 6, Figs. 1 and 21994. Acaeniotyle rebellis nov. sp. – O’Dogherty, p. 287,
l. 51, Figs. 5–10.Occurrence: This species is known to occur throughout the
pper Albian to Lower Turonian of the UMB (O’Dogherty,994; this study).
Material: Eight specimens.
Acaeniotyle sp. cf. A. umbilicata (Rust, 1898)Plate 6, Fig. 3cf. 1898. Xiphospaera umbilicata n. sp. – Rust, p. 7, Pl. 1,
ig. 9.cf. 1981. Acaeniotyle umbilicata (Rust) – Schaaf, p. 431, p. 6,
ig. 11; Pl. 15, Figs. 3a and 3b.cf. 1994. Acaeniotyle umbilicata (Rust) – O’Dogherty, p. 289,
l. 51, Figs. 19 and 20.cf. 1994. Acaeniotyle umbilicata (Rust) – Erbacher, p. 89,
l. 1, Fig. 2.cf. 2004. Acaeniotyle umbilicata (Rust) – Bragina, p. S401,
l. 17, Figs. 3 and 7; Pl. 41, Fig. 8.Occurrence: In the UMB, this species is reported through-
ut the Upper Barremian to Lower Albian (Erbacher, 1994;’Dogherty, 1994). The probable presence of A. umbilicata in a
ample coming from the base of the BL may extend its known ageange in the UMB to the uppermost Cenomanian. The species
282 B. Musavu-Moussavou et al. / Revue de micropaleontologie 50 (2007) 253–287
s also known from the Middle to Upper Aptian of the northerntlantic (Erbacher, 1994), from the Middle–Upper Cenomanianf northern Turkey and from the Lower Turonian of the Crimeanountains (Bragina, 2004).Material: Four specimens.
l. 39, Figs. 7 and 10.Occurrence: In the UMB, this species is known to
ccur throughout the Middle Cenomanian to Lower TuronianO’Dogherty, 1994; Salvini and Marcucci Passerini, 1998). Inhis study, the species H. sexangulum was found in only oneample (BTT 572) coming from limestones overlying the BL. Its also known from the Lower Cenomanian to Middle Turonianf California (Pessagno, 1971a), from the Upper Cenomaniano Lower Turonian of the Crimean Mountains and from the
iddle–Upper Cenomanian of northern Turkey (Bragina, 2004).Material: 15 specimens.
Genus Pessagnobrachia Kozur and Mostler, 1978Type species: Patulibracchium testlaensis Pessagno, 1971a
Pessagnobrachia fabianii (Squinabol, 1914)Plate 6, Figs. 6 and 71914. Rhopalastrum Fabianii n. f. – Squinabol, p. 274,
l. 21[2], Fig. 4.1994. Pessagnobrachia fabianii (Squinabol) – O’Dogherty,
igs. 7A and B.2004. Alievium superbum (Squinabol) – Bragina, p. S427,
l. 36, Fig. 1.Occurrence: In the UMB, A. superbum is reported from
he middle part of the BL and the overlying Lower Turonianimestones (Marcucci Passerini et al., 1991; O’Dogherty, 1994;alvini and Marcucci Passerini, 1998). The species is also knownrom the Lower Turonian to Lower Coniacian of CaliforniaPessagno, 1976), from the Lower Turonian of Turkey (Yurtsevert al., 2003) and from the Lower Turonian of the Crimean Moun-ains (Bragina, 2004).
Material: 16 specimens.
Genus Becus Wu, 1986
Type species: Becus gemmatus Wu, 1986
Becus regius O’Dogherty, 1994Plate 6, Fig. 11
2 de mi
F
F
M(wka2
P
F
F
p
Ult
P
iotf2
p
p
c
O
M
a
p
MOsTT
F
(
F
1oa
F
p
P
P
U
84 B. Musavu-Moussavou et al. / Revue
1994. Becus regius nov. sp. – O’Dogherty, p. 318, Pl. 58,igs. 18–26.
2004. Becus regius O’Dogherty – Bragina, p. S439, Pl. 24,igs. 1 and 10.
Occurrence: In the UMB, this species is known to occur fromiddle Albian to Upper Cenomanian strata underlying the BL
U.A. 12–19 of O’Dogherty, 1994). During this study B. regiusas found to occur in the lower part of the BL. This species is alsonown to occur in the Upper Cenomanian of northern Turkeynd the Lower Turonian of the Crimean Mountains (Bragina,004).
Material: Two specimens.
Genus Dactyliodiscus Squinabol, 1903bType species: Dactyliodiscus cayeuxi Squinabol, 1903b
Dactyliodiscus longispinus (Squinabol, 1904)Plate 6, Fig. 121904. Spongolonche diversispina n. sp. – Squinabol, p. 206,
l. 6, Fig. 6.1904. Stylotrochus euganeus n. sp. – Squinabol, p. 207, Pl. 6,
ig. 8.1904. Stylotrochus euganeus n. sp. – Squinabol, p. 207, Pl. 6,
. 333, Pl. 62, Figs. 6–11.Occurrence: Upper Albian to Middle Cenomanian of the
MB (O’Dogherty, 1994). The presence of D. longispinus inimestones underlying the BL extends its known age range tohe Uppermost Cenomanian.
Material: Five specimens.
Dactilyodiscus spinosus Bragina, 2004Plate 6, Fig. 132004. Dactilyodiscus spinosus n. sp. – Bragina, p. S431,
l. 25, Fig. 9.Occurrence: During this study, D. spinosus was found
n only one sample (BTT 564) coming from limestonesverlying the BL. To our knowledge, this is the first thathis species is reported from the UMB. It is also knownrom Upper Cenomanian strata of northern Turkey (Bragina,004).
Material: One specimen.
Genus Pseudoaulophacus Pessagno, 1963Type species: Pseudoaulophacus floresensis Pessagno, 1963
. S425, Pl. 36, Fig. 3.Occurrence: In the UMB, this species is reported from
iddle Albian to Lower Turonian levels (Erbacher, 1994;’Dogherty, 1994; Salvini and Marcucci Passerini, 1998; this
tudy). P. putahensis is also reported from the Cenomanian touronian of Turkey (Yurtsever et al., 2003) and from Loweruronian strata of the Crimean Mountains (Bragina, 2004).
Material: Nine specimens.
Family SPONGURIDAE Haeckel, 1862Genus Patellula Petrushevskaya and Kozlova, 1972Type species: Stylospongia planoconvexa Pessagno, 1963
Patellula cognata O’Dogherty, 1994Plate 6, Fig. 151994. Patellula cognata n. sp. – O’Dogherty, p. 326, Pl. 60,
igs. 6–12.Occurrence: Lower to Middle Cenomanian of the UMB
O’Dogherty, 1994; this study).Material: One specimen.
Patellula ecliptica O’Dogherty, 1994Plate 6, Figs. 16 and 171994. Patellula ecliptica nov. sp. – O’Dogherty, p. 329, Pl. 61,
igs. 1–5.Occurrence: Lower Turonian of the UMB (O’Dogherty,
994). The presence of this species within the lower partf the BL (Uppermost Cenomanian) extends its knownge.
Material: 12 specimens.
Patellula helios (Squinabol, 1903b)Plate 6, Fig. 181903b. Stylotrochus helios n. sp. – Squinabol, p. 124, Pl. 10,
igs. 23 and 23a.1982. Pseudoaulophacus lenticulatus (White) – Taketani,
’Dogherty, p. 292, Pl. 52, Figs. 6–10.2004. Staurosphaeretta longispina (Squinabol) – Bragina,
. S396, Pl. 16, Figs. 2, 4, 5; Pl. 37, Fig. 14; Pl. 38, Fig. 2.Occurrence: In the UMB, this species is reported from Upper
ptian to Upper Cenomanian strata underlying the BL (U.A.–18 of O’Dogherty, 1994). During this study the species wasound in only one sample (BG 54, Middle Cenomanian). S.ongispina is also reported from the Middle to Upper Cenoma-ian of northern Turkey and the Lower Turonian of the Crimeanountains (Bragina, 2004).Material: Five specimens.
Genus Triactoma Rust, 1885 emend. Pessagno and Yang inessagno et al., 1989
Type species: Triactoma tithonianum Rust, 1885
Triactoma hexeris O’Dogherty, 1994Plate 6, Figs. 20 and 211994. Triactoma cf. aff. T. echioides Foreman – Marcucci
asserini et al. Pl. 1, Fig. 12.1994. Triactoma hexeris nov. sp. – O’Dogherty, p. 303, Pl. 55,
igs. 24–27.1998. Triactoma hexeris O’Dogherty – Salvini and Marcucci
ig. 6B.Occurrence: In the UMB, this species is known to occur
hroughout the Lower Cenomanian to Lower Turonian interval
O’Dogherty, 1994; Salvini and Marcucci Passerini, 1998; thistudy). It is also known from Cenomanian to Turonian strata ofurkey (Yurtsever et al., 2003).
Material: 13 specimens.
C
cropaleontologie 50 (2007) 253–287 285
cknowledgements
Constructive remarks by P. Dumitrica and L. O’Doghertymproved the initial version of the manuscript. C. Abrial and. Lethiers helped with drawing and plate preparation. This isublication no. 18 of the Centro di Geobiologia of the Universityf Urbino.
eferences
liev, K.S., 1965. Radiolarians of the Lower Cretaceous deposits of northeast-ern Azerbaidzhan and their stratigraphic significance. Izdatel’stvo akademiiNauk, Azerbaidzhanskoi SSR, Baku, p. 124.
rthur, M.A., Premoli Silva, I., 1982. Development of widespread organic car-bon rich strata in the Mediterranean Tethys. In: Schlanger, S.O., Cita, M.B.(Eds.), Nature and Origin of Cretaceous Carbon-Rich Facies. AcademicPress, London, pp. 7–54.
accelle, L., Bosellini, A., 1965. Diagrammi per la stima visiva della com-posizione percentuale nelle rocce sedimentarie. Annali dell’Universita diFerrara (Nuovo Serie). Sezione IX, Scienze Geologiche e Paleontologiche1, 117–153.
ak, K., Bak, M., Paul, Z., 2001. Barnasiowka radiolarian shale formation—anew lithostratigraphic unit in the Upper Cenomanian–Lowermost Turonianof the Polish outer Carpathians (Silesian series). Annales Societatis Geolo-gorum Poloniae 71, 75–103.
ak, M., 1993. Late Albian–Early Cenomanian Radiolaria from the Czorstynsuccession, Pienny Klippen belt, Carpathians. Studia Geologica Polonica102, 177–207.
ak, M., 1995. Mid Cretaceous Radiolaria from the Pienny Klippen Belt,Carpathians, Poland. Cretaceous Research 16, 1–23.
ak, M., 1999. Cretaceous radiolarian zonation in the Polish part of the PiennyKlippen belt (Western Carpathians). Geologica Carpathica 50, 21–31.
aumgartner, P., 1980. Late Jurassic Hagiastridae and Patulibracchiidae(Radiolaria) from the Argolis Peninsula (Peloponnesus, Greece). Micropa-leontology 26, 274–322.
aumgartner, P.O., Bjorklund, K.R., Caulet, J.P., De Wever, P., Kelogg, D.,Labracherie, M., Nakaseko, K., Nishimura, A., Schaaf, A., Schmidt-Effing,R., Yao, A., 1981. Second European meeting of radiolarian paleontologists:current research on Cenozoic and Mesozoic radiolarians. Eclogae Geologi-cae Helvetiae 74, 1027–1061.
ampbell, A.S., 1954. Radiolaria. In: Moore, R.C. (Ed.), Treatise on Inverte-brate Paleontology, Part D, Protista 3. Geological Society of America andUniversity of Kansas Press, Lawrence/Kansas, pp. 11–195.
ampbell, A.S., Clark, B.L., 1944. Radiolaria from upper Cretaceous of middleCalifornia. Geological Society of America Special Papers 57, 1–61.
aron, M., Robaszynski, F., Amedo, F., Baudin, F., Deconinck, J.-F., Hochuli, P.,Von Salis-Perch, Nielsen, Tribovillard, N., 1999. Estimation de la duree del’evenement anoxique global au passage Cenomanien/Turonien. Approchecyclostratigraphique dans la formation Bahloul en Tunisie centrale. Bulletinde la Societe Geologique de France 2, 145–160.
ayeux, L., 1897. Contribution a l’etude micrographique des terrainssedimentaires. 1. Etude de quelques depots siliceux secondaires et tertiairesdu Bassin de Paris et de la Belgique. 2. Craie du Bassin de Paris. Memoiresde la Societe geologique du Nord 4 (2), 1–591.
occioni, R., Erba, E., Premoli Silva, I., 1991. Litho- and biostratigraphyof the Livello Bonarelli close to the Cenomanian/Turonian bound-ary (Umbria-Marche Apennines, Italy) and possible paleoceanographicsignificance. Colloque International sur les Evenements de la LimiteCenomanien/Turonien, Grenoble, 24–26 Mai 1991. Geologie Alpine,
Memoire H.S. 17, 25–26.
occioni, R., Erba, E., Premoli Silva, I., 1992. Barremian–Aptian calcareousplankton biostratigraphy from the Gorgo a Cerbara section (Marche, cen-tral Italy) and implications for plankton evolution. Cretaceous Research 13,517–537.
2 de mi
C
D
D
D
D
D
D
E
E
E
E
E
E
E
E
F
F
F
F
F
F
G
G
G
H
H
H
H
H
J
K
K
K
K
K
K
K
K
L
L
M
M
86 B. Musavu-Moussavou et al. / Revue
occioni, R., Luciani, V., 2004. Planktonic foraminifera and environmentalchanges across the Bonarelli event (OAE2, latest Cenomanian) in its typearea: A high-resolution study from the Tethyan reference Bottaccione section(Gubbio, central Italy). Journal of Foraminiferal Research 3, 109–129.
eflandre, G., 1953. Radiolaires fossiles. In: Grasse, P.P. (Ed.), Traite de Zoolo-gie 1 (2). Masson, Paris, pp. 389–436.
e Wever, P., 1982. Radiolaires du Trias et du Lias de la Tethys (systematiquestratigraphie). Societe Geologique du Nord 7, 1–600.
e Wever, P., Dumitrica, P., Caulet, J.P., Nigrini, C., Caridroit, M., 2001. Radio-larians in the Sedimentary Record. Gordon and Breach Science Publishers,pp. 1–533.
e Wever, P., Thiebault, F., 1981. Les radiolaires d’age Jurassique superieura Cretace superieur dans les radiolarites du Pinde–Olonos (presqu’ıle deKoroni ; Peloponnese meridional, Grece). Geobios 14, 577–609.
umitrica, P., 1970. Cryptocephalic and cryptothoracic Nassellaria in someMesozoic deposits of Romania. Revue roumaine de Geologie. Geophysiqueet Geographie 14, 45–124.
umitrica, P., 1975. Cenomanian radiolaria at Podul Dımbovitei. Micropaleon-tological guide to the Mesozoic and Tertiary of the Romanian Carpathians.In: 14th European Micropaleontological Colloquium: Institute of Geologyand Geophysics, Bucharest, pp. 87–89.
hrenberg, C.G., 1847. Ueber die mikroskopischen Kieselschaligen Polycystienals machtige Gebirgsmasse von Barbados und uber das Verhaltniss deraus mehr als 300 neuen Arten bestehenden ganz eigenthumlichen Formen-gruppe jener Felsmasse zu den jetzt lebenden Thieren und zur kreidebildungEine neue Anregung zur Erforscung des Erdlebens. Koniglische PreussischeAkademie der Wissenschaften zu Berlin, Bericht, 40–60.
hrenberg, C.G., 1875. Fortsetzung der mikrogeologischen Studien alsGesammt-Uebersichtder mikrokopischen Palaontologie gleichartiganalysirter Gebirgsarten der Erde, mit specieller Rucksicht auf denPolycystinen-Mergel von Barbados. Konigliche Akademie der Wissen-schaften zu Berlin, Abhandlungen, Jahre 1875, 1–225.
mpson-Morin, K.M., 1981. Campanian Radiolaria from DSDP Site 313, Mid-Pacific Mountains. Micropaleontology 27, 249–292.
rbacher, J., 1994. Entwicklung und Palaoozeoanographie mittelkretazischerRadiolarien der westlichen Tethys (Italien) und des Nordatlantiks. TubingerMikropalaontologische Mitteilungen, Institut und Museum fur Geologie undPalaontologie der Universitat Tubingen 12, 1–120.
rbacher, J., 1998. Mid-Cretaceous radiolarians from the Eastern equatorialAtlantic and their paleoceanography. In: Mascle, J., Lohmann, G.P., Moul-lade, M. (Eds.), Proccedings of the Ocean Drilling Program, ScientificResults 159, pp. 363–373.
rbacher, J., Thurow, J., 1997. Influence of oceanic anoxic events on the evo-lution of mid-Cretaceous radiolaria in the North Atlantic and west Tethys.Marine Micropaleontology 30, 139–158.
rbacher, J., Thurow, J., 1998. Mid-Cretaceous radiolarian zonation for theNorth Atlantic: an example of oceanographically controlled evolutionaryprocesses in the marine biosphere? In: Cramp, A., Macleod, C.J., Lee,S.V., Jones, E.J.W. (Eds.), Geological Evolution of Ocean Basins Resultsfrom Ocean Drilling Program, 131. Geological Society, London SpecialPublications, pp. 71–82.
spitalie, J., Deroo, G., Marquis, F., 1985. La pyrolyse Rock-Eval et ses appli-cations. Revue de l’Institut Francais du Petrole 40, 563–579.
oreman, H.P., 1968. Upper Maastrichtian Radiolaria of California. Thepalaeontological association. Special Papers in Palaeontology 3, 1–82.
oreman, H.P., 1973. Radiolaria from DSDP Leg 20. In: Heezen, B.C., Mac-Gregor, J.D., et al. (Eds.), Initial Reports of the Deep Sea Drilling Project20. US Government Printing Office, Washington, pp. 249–305.
oreman, H.P., 1975. Radiolaria from the North Pacific, Deep Sea DrillingProject, Leg 32. In: Larson, R.L., Moberdy, R., et al. (Eds.), Initial Reportsof the Deep Sea Drilling Project 32. US Government Printing Office, Wash-ington, pp. 579–676.
oreman, H.P., 1978. Cretaceous Radiolaria in the Eastern South Atlantic, deep
sea drilling project, Leg 40. In: Bolli, H.M., Ryan, W.B.F., et al. (Eds.),Initial Reports of the Deep Sea Drilling Project 40. US Government PrintingOffice, Washington, pp. 839–843.
rohlich, F., 1989. Deep-sea biogenic silica: new structural and analytical datafrom infrared analysis - geological implications. Terra Nova 1, 267–273.
M
cropaleontologie 50 (2007) 253–287
rohlich, F., Gendron-Badou, A., 2003. La spectroscopie infrarouge, un outilpolyvalent. In: Miskovsky, J.C. (Ed.), Geologie de la Prehistoire. AEEGP,editeur, Paris, pp 663–667.
allicchio, S., Marcucci, M., Pieri, P., Premoli Silva, I., Sabato, L., Salvini,G., 1996. Stratigraphical data from a Cretaceous claystones sequence of the“Argile Varicolori” in the Southern Apennines (Basilicata, Italy). Palaeopela-gos 6, 261–272.
orican, S., 1994. Jurassic and Cretaceous radiolarian biostratigraphy and sed-imentary evolution of the Budva Zone (Dinarides, Montenegro). Memoiresde Geologie (Lausanne) 18, 1–177.
orican, S., Smuc, A., 2004. Albian Radiolaria and Cretaceous stratigraphy ofMt Mangart (Western Slovenia). Razprave IV. Razereda sazu XLV-3, 29–49.
aeckel, E., 1862. Die Radiolarien (Rhizopoda Radiolaria). Eine monographie.Reimer, Berlin.
aeckel, E., 1881. Entwurf eines radiolarien-systems auf grund von studiender challenger-radiolarien. Jenaische Zeitschrift fur Naturwissenschaft 15,418–472.
allam, A., Wignall, P.B., 1997. Mass Extinctions and their aftermath. OxfordUniversity Press, Oxford.
arries, P., Little, C.T.S., 1999. The early Toarcian (Early Jurassic) and theCenomanian–Turonian (Late Cretaceous) mass extinction: similarities andcontrasts. Palaeogeography, Palaeoclimatology, Palaeoecology 154, 39–66.
olmes, W.M., 1900. On Radiolaria from the Upper Chalk at Coulsdon (Surrey).Quarterly Journal of the Geological Society of London 56, 694–704.
enkyns, H.C., 1980. Cretaceous anoxic events from continents to oceans. Jour-nal of the Geological Society of London 137, 171–188.
han, S., Urquhart, E., Thurow, J., 1999. Radiolaria and Cenomanian–Turonianenvironments of Hacho de Montejaque, Penibetic, southern Spain. In:Farinacci, A., Lord, A.R. (Eds.), Depositional Episodes and Bioevents.Palaeopelagos Special Publication 2, pp. 123–143.
iessling, W., 1996. Facies characterization of Mid-Mesozoic deep-water sedi-ments by quantitative analysis of siliceous microfaunas. Facies 35, 237–274.
ozlova, G.E., Gorbovetz, A.N., 1966. Radiolarians of the Upper Eocenedeposits of West Siberian Lowland, Proceedings of the All Union PetroleumScientific Research Institute for Geological Survey (VNIGRI) 248, pp.1–159.
ozur, H., Mostler, H., 1978. Beitrage zur Erforschung der mesozoischenRadiolarien. Teil II: Oberfamilie Trematodiscacea Haeckel, 1862 emend.und Beschreibung ihrer triassischen Vertreter. Geologische PalaontologischeMitteillungen Innsbruck 8, 123–182.
ozur, H., Mostler, H., 1982. Entactinaria subordo nov., a new radiolarian sub-order. Geologische Palaontologische Mitteilungen Innsbruck 11, 399–414.
uhnt, W., 1990. Agglutinated foraminifera of western Mediterranean UpperCretaceous pelagic limestones (Umbria Apennines, Italy, and BeticCordillera, Southern Spain). Micropaleontology 36, 133–160.
uhnt, W., Thurow, J., Wiedmann, J., Herbin, J.P., 1986. Oceanic anoxic condi-tions around the Cenomanian/Turonian boundary and the response of biota.Mitteilungen Geologisch-Palaontologische, Institut Universitet Hamburg60, 205–246.
uroda, J., Ohkouchi, N., Ishii, T., Tokuyama, H., Taira, A., 2005. Lamina-scaleanalysis of sedimentary components in Cretaceous black shales by chemi-cal compositional mapping: Implications for paleoenvironmental changesduring the oceanic anoxic events. Geochimica, Cosmochimica Acta 69,1479–1494.
eckie, R.M., Bralower, T.J., Casham, R., 2002. Oceanic anoxic events andplankton evolution: Biotic response to tectonic forcing the mid-Cretaceous.Paleoceanography 17, 1–29.
ipman, R.Kh., 1969. Novy rod i novyye vidy eotsenovykh radiolaryariy SSSR.Vsesyuz. Nauchno-Issled. Geol. Inst. Trudy 130, 181–200.
arcucci Passerini, M., Bettini, P., Dainelli, J., Sirugo, A., 1991. The “BonarelliHorizon” in the central Apennines (Italy): radiolarian biostratigraphy. Cre-taceous Research 12, 321–331.
arcucci Passerini, M., Cabella, R., Passerini, P., 1994. Early–late Cretaceous
radiolarian deposits in the Northern Apennines: biostratigraphy and min-eralogical data from the “Scisti Policromi” in the Tuscan succession nearMonsummanto, Tuscany. Palaeopelagos 4, 23–34.
ekik, F., 2000. Early Cretaceous Pantanelliidae (Radiolaria) from northwestTurkey. Micropaleontology 46, 1–30.
de mi
M
M
M
N
O
O
P
P
P
P
P
P
P
P
P
P
P
P
P
R
R
R
R
S
S
S
S
S
S
S
S
S
S
S
T
T
T
T
T
U
V
W
Y
Z
B. Musavu-Moussavou et al. / Revue
orel, L., 1998. Stratigraphie a haute resolution de passage Cenomanien–Turo-nien. Academie de Paris, These de l’Universite Pierre-et-Marie-Curie, Paris.
unasri, K.W., Bambang, W., 1994. Cretaceous radiolarians from the Luk-Ulo Melange Complex in the Karangsambung area, central Java, Indonesia.Journal of Southeast Asian Earth Sciences 9, 29–43.
usavu-Moussavou, B., Danelian, T., 2006. The radiolarian response to oceanicanoxic event 2 in the southern part of the Northern proto-Atlantic (DemeraraRise, ODP leg 207). Revue de Micropaleontologie 49, 141–163.
akaseko, K., Nishimura, A., Sugano, K., 1979. Cretaceous Radiolaria in theShimanto Belt, Japan. News of Osaka Micropaleontologist special publica-tion 2, 1–49.
’Dogherty, L., 1994. Biochronology and paleontology of Mid-Cretaceousradiolarians from Northern Apennines (Italy) and Betic Cordillera (Spain).Memoires de Geologie (Lausanne) 21, 1–411.
’Dogherty, L., Guex, J., 2002. Rates and pattern of evolution among Cretaceousradiolarians: relations with global paleoceanographic events. Micropaleon-tology 48 (supplement 1), 1–22.
essagno Jr., E.A., 1963. Upper Cretaceous Radiolaria from Puerto Rico.Micropaleontology 9, 197–214.
essagno Jr., E.A., 1969. The Neosciadiocapsidae, a new family of Upper Cre-taceous Radiolaria. Bulletin of American Paleontology 56, 377–439.
essagno Jr., E.A., 1971a. Jurassic and Cretaceous Hagiastridae from Blake-Bahama basin (Site 5A, JOIDES Leg 1) and the Great Valley sequence,California Coast ranges. Bulletins of American Paleontology 60, 5–83.
essagno Jr., E.A., 1971b. A new radiolarian from the Upper Cretaceous of theCalifornia Coast Ranges. Micropaleontology 17, 361–364.
essagno Jr., E.A., 1972. Cretaceous Radiolaria. Part I: the Phaseliformidae, newfamily, and other Spongodiscacea from the Upper Cretaceous portion of theGreat Valley Sequence. Part II: Pseudoaulophacidae Riedel from the Creta-ceous of California and the Blake-Bahama basin (JOIDES leg 1). Bulletinsof American Paleontology 61, 269–328.
essagno Jr., E.A., 1973. Upper Cretaceous Spumellarian from the Great Valleysequence California coast ranges. Bulletins of American Paleontology 63,49–102.
essagno Jr., E.A., 1976. Radiolarian zonation and stratigraphy of the UpperCretaceous of the Great Valley sequence, California coast ranges. Micropa-leontology special publication 2, 1–95.
essagno Jr., E.A., 1977a. Upper Jurassic Radiolaria and radiolarian biostratig-raphy of California coast ranges. Micropaleontology 23, 56–113.
essagno Jr., E.A., 1977b. Lower Cretaceous radiolarian biostratigraphy of GreatValley Sequence and Franciscan complex, California coast ranges. Cushmanfoundation for foraminiferal research, special publication 15, 1–87.
essagno Jr., E.A., Six, W.M., Yang, Q., 1989. The Xiphostylidae Haeckeland Parvivaccidae, n. fam., (Radiolaria) from the North American Jurassic.Micropaleontology 35, 193–255.
etrushevskaya, M.G., Kozlova, G.E., 1972. Radiolaria: leg 14, deep drillingproject. In: Hayes, D.E., Pimm, A.C., et al. (Eds.), Initial Reports of DeepSea Drilling Project 14. US Government Printing Office, Washington, pp.495–648.
ignotti, L., 1994. The Cenomanian–Turonian siliceous-anoxic event in theScisti Policromi (Tuscan succession, Northern Apennines): data on radi-olarian and foraminiferal biostratigraphy. Palaeopelagos 4, 141–153.
remoli Silva, I., Erba, E., Salvini, G., Locatelli, C., Verga, D., 1999. Bioticchanges in Cretaceous oceanic anoxic events of the Tethys. Journal ofForaminiferal Research 29, 352–370.
iedel, W.R, 1967. Subclass Radiolaria. In: Harland, W.B., et al. (Eds.), TheFossil Record. A Symposium with Documentation. Geological Society ofLondon, pp. 291–298.
iedel, W.R., 1971. Systematic classification of polycystine Radiolaria. In:Funnell, B.M., Riedel, W.R. (Eds.), The micropaleontology of the oceans.Cambridge University Press, Cambridge, pp. 649–661.
ust, D., 1885. Beitrage zur Kenntniss der fossilen Radiolarien aus Gesteinendes Jura. Palaeontographica 31, 269–321.
ust, D., 1898. Neue Beitrage zur Kenntniss der fossilen Radiolarien ausGesteinen des Jura und der Kreide. Palaeontographica 45, 1–67.
alvini, G., Marcucci Passerini, M., 1998. The radiolarian assemblages of theBonarelli Horizon in the Umbria-Marches Apennines and Southern Alps,Italy. Cretaceous Research 19, 777–804.
Z
cropaleontologie 50 (2007) 253–287 287
anfilippo, A., Riedel, W.R., 1985. Cretaceous Radiolaria. In: Bolli, H.M.,Saunders, J.B., Perch-Nielsen, K. (Eds.), Plankton Stratigraphy. CambridgeUniversity Press, Cambridge/New York, pp. 573–630.
chaaf, A., 1981. Late Cretaceous Radiolaria from Deep Sea Drilling ProjectLeg 621. In: Thiede, J., Vaillier, T.L., et al. (Eds.), Initial Reports of the DeepSea Drilling Project 62. US Government Printing Office, Washington, pp.419–470.
copelliti, G., Bellanca, A., Coccioni, R., Luciani, V., Neri, R., Baudin, F.,Chiari, M., Marcucci, M., 2004. High-resolution geochemical and bioticrecords of the Tethyan ‘Bonarelli level’ (OAE2, Latest Cenomanian) from theCalabianca–Guidaloca composite section, northwestern Sicily, Italy. Palaeo-geography, Palaeoclimatology, Palaeoecology 208, 293–317.
copelliti, G., Bellanca, A., Neri, R., Baudin, F., Coccioni, R., 2006. Com-parative high-resolution chemostratigraphy of the Bonarelli level from thereference Bottaccione section (Umbria-Marche Apennines) and from anequivalent section in NW Sicily: consistent and contrasting responses toOAE2. Chemical Geology 228, 266–285.
quinabol, S., 1903a. Radiolarie fossile di Teolo (Euganei). Atti e memoriedell’Accademia di Scienze di Lettere ed Arti Padova, n. ser. 19, 127–130.
quinabol, S., 1903b. Le Radiolarie dei noduli selciosi nella Scaglia degliEuganei. Contribuzione I. Rivista Italiana di Paleontologia 19, 127–130.
quinabol, S., 1904. Radiolarie cretacee degli Euganei. Atti e memoriedell’Accademia di Scienze, Lettere ed Arti, Padova, n. ser. 20, 171–244.
quinabol, S., 1914. Contributo alla conoscenza dei Radiolarii fossili del Veneto.Appendice di un genere di Radiolari caratteristico del Secondario. Memoriedell’Istituto Geologico della R. Universita di Padova 2, 249–306.
uzuki, H., Yamaguchi, M., Setoguchi, T., 2002. Cretaceous radiolaria fromchert pebbles of the Miocene Honda Group of Huila, central Colombia.Neues Jahrbuch fur Geologie und Palaontologie Monatshefte 1, 48–64.
aketani, Y., 1982. Cretaceous radiolarian biostratigraphy of the Urakawa andObira Areas, Hokkaido. Science Reports of the Tohuku University, Sendai52, 1–76.
an, S.H., 1927. Over de samenstelling en het onstaan van krijten mergel-gesteenten van de Molukken. In: Brouwer, H.A. (Ed.), Jaarboek van hetmijnwezen in Nederlandsch Oost-Indie, Jaargan 55, 1926, verhandelingen,3rd gedeelte, pp. 5–165.
hurow, J., 1998. Cretaceous radolarians of the North Atlantic Ocean: ODPLeg 103 (Sites 638, 640 and 641). In: Boillot, G., Winterer, E.L. (Eds.),Proceedings of the Ocean Drilling Program, Scientific Results, pp. 379–418.
sikos, H., Jenkyns, H.C., Walsworth-Bell, B., Petrizzo, M.R., Forster, A.,Kolonic, S., Erba, E., Premoli Silva, I., Baas, M., Wagner, T., SinningheDamste, J.S., 2004. Carbon-isotope stratigraphy recorded by the Ceno-manian/Turonian oceanic anoxic event: correlation and implications basedon three key-localities. Journal of the Geological Society of London 161,711–719.
umanda, F.P., 1989. Cretaceous Radiolaria biostratigraphy in the Esashi Moun-tain area, Northern Hokkaido, Japan. Science Reports of the Institute ofGeoscience, University of Tsukuba, section B10, 1–44.
rquhart, E., 1994. New data on the ranges of some Cretaceous Tethyan radi-olaria. Comptes Rendus de l’Academie des Sciences de Paris (2) 318,1401–1407.
ishnevskaya, V., De Wever, P., 1998. Upper Cretaceous Radiolaria fromthe Russian Platform (Moscow Basin). Revue de Micropaleontologie 41,235–265.
u, H.R., 1986. Some new genera and species of Cenomanian Radiolarian fromsouthern Xizang (Tibet). Acta Micropaleontologica Sinica 3, 347–360.
urtsever, T.S., Tekin, U.K., Demirel, I.H., 2003. First evidence of the Cenoma-nian/Turonian boundary event (CTBE) in the Alakircay Nappe of AntalyaNappes, southwest Turkey. Cretaceous Research 24, 41–53.
iabrev, S.V., Aitchison, J.C., Abrajevitch, A.V., Badengzhu, Davis, A.M., Luo,H., 2003. Precise radiolarian age constraints on the timing of ophiolite
generation and sedimentation in the Dazhuqu terrane, Yarlung-Tsangposuture zone, Tibet. Journal of the Geological Society of London 160,591–599.
ittel, K.A., 1876. Uber einige fossile Radiolarien aus der norddeutschen Krei-den. Zeitschrift der Deutschen Geologischen Gesellschaft 28, 75–87.