Aalenian carbon-isotope stratigraphy: Calibration with ammonite, radiolarian andnannofossil events in the Western Tethys
José Sandoval a,⁎, Luis O'Dogherty b, Roque Aguado c, Annachiara Bartolini d,Sébastien Bruchez e, Markus Bill f
a Dpto Estratigrafía y Paleontología, Universidad de Granada, 18002 Granada, Spainb Dpto. Ciencias de la Tierra; CASEM, 11510, Puerto Real, Spainc Dpto de Geología, Universidad de Jaén, Alfonso X EL Sabio 28, 23700 Linares, Spaind CNRS UMR 5143, MNHN 8 rue Buffon, 75231, Paris, Francee Institut de Géologie et Paléontologie, Bâtiment Anthropole, 1015 Lausanne, Switzerlandf Center for Isotope Geochemistry, Lawrence Berkeley National Laboratory, Berkeley CA 94720, United States
bon-isotope stratigraphy of four Aalenian sections well-dated with ammonitesfrom the Median Subbetic palaeogeographic domain (external zones of the Betic Cordillera, southern Spain)is presented. Two of these represent the most complete Aalenian sections of the southern Iberianpalaeomargin (western Tethys). These sections are rhythmic sequences made of marls and marly limestonescontaining rich ammonite fauna and nannofossils that enable an accurate biostratigraphic control at thesubzone level. Moreover, the lower and upper Aalenian boundaries are well defined in these sections andtherefore represent optimal sites to link the carbon-isotope curves to ammonite zones and subzones andnannofossil events.The uppermost Toarcian–lowermost Bajocian δ13C values of bulk carbonates show evident fluctuations. Thevalues are low (~1‰) in the Upper Toarcian (Mactra Subzone), increase slightly to ~2‰ in Lower Aalenian(Comptum Subzone), fall to a relative minimum (~1–1.5‰) in the Middle Aalenian (Bradfordensis Zone),reach a marked positive peak of 2.5–2.7‰ in the Upper Aalenian (Concavum Subzone), and then the valuesdecrease again to 1.5–1.7‰ near the Aalenian/Bajocian boundary. Marine carbonate δ13C curves in Subbeticand the Apennines shows similar variations but higher δ13C values in the Apennines sections. A slight offsetis also present between the sections from the same basin. This drift is interpreted by variations in carbonisotopic composition of sea-water DIC associated with different palaeoceanographic settings.Comparison of the δ13C values of bulk carbonates and biotic events (ammonite diversity, and nannofossil andradiolarian abundances) reveals relationships between the carbon cycle and biotic evolution. The latestToarcian–Aalenian turnovers (Toarcian/Aalenian boundary and Middle/Late Aalenian) follow low δ13C values,where extinct taxa are replaced by new ones. Radiation episodes are associated with increasing δ13C valuesand with sea-level rise. In the earliest Late Aalenian, concomitantly with a major δ13C positive excursion andwith the transgressive cycle, it is noted a major increase of the eutrophic calcareous nannofossil genusBiscutum and a sharp peak in radiolarian abundance. Although the Aalenian interval seems not to becharacterized by major large amplitude negative or positive excursions, as observed in other Phanerozoicintervals, these data point to a close connection between perturbations in the global carbon cycle and thepalaeoenvironmental changes that occurred, which in turn had important interactions with the evolution ofpelagic biota.
In this study is presented a detailed carbon-isotope stratigraphy forthe Aalenian (Middle Jurassic) pelagic carbonates in the Betic Cordillera
l rights reserved.
(southern Spain), which represents an ideal region to directly tie thestable carbon-isotope curves to ammonite zones and subzones, andthereby provide an accurate chronostratigraphic calibration.
Aalenian sediments are well-represented in the Mesozoic basinsbordering the Iberian Meseta: Lusitanian Basin and Cantabrian,Iberian and Betic Cordilleras. The stratotype of the lower boundariesof the Aalenian and Bajocian stages (the Fuentelsalz Section in theIberian Cordillera and the Cabo Mondego section in the LusitanianBasin) were established in these basins (Rocha et al., 1990; Pavia and
116 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
Enay, 1997; Cresta et al., 2001). Although the Aalenian and Bajocianstratotype boundaries are located in the Iberian Cordillera andLusitanian Basin, the Median Subbetic domain of the Betic Cordillerahas possibly the most complete and expanded Aalenian stratigraphicsections of the Iberian Palaeomargin, with pelagic or hemipelagiccontinuous sedimentation throughout the complete stage. Moreover,some Subbetic sections contain abundant and well preservedammonites, permitting the establishment of a detailed biostrati-graphic scheme in which all the Mediterranean Aalenian ammonitezones and subzones are represented (Linares and Sandoval, 1993;Henriques et al., 1996; Sandoval et al., 2001a). Also, calcareousnannofossils and radiolarians are common and, the former arerelatively well preserved, allowing good biostratigraphic correlations.Therefore, the Subbetic constitutes an ideal area to study chemos-tratigraphy of the Aalenian and to establish the possible relationshipsbetween chemical cycle variations and biotic changes of marinepelagic organisms.
During the last 40 yr, several specific works on the Aalenian of theBetic Cordillera have been published (García-Dueñas et al., 1967;Linares, 1979; Linares and Sandoval, 1986, 1988, 1990, 1992, 1993,1996; Hernández-Molina et al., 1991; García-Gómez et al., 1995;Henriques et al., 1996; Sandoval and Chandler, 2000; Sandoval et al.,2001a). These articles give a comprehensive account of the strati-graphy, palaeobiogeography, ammonite fossil content, and biochro-nology of the Aalenian of the Betic Cordillera, and thus a frameworkfor chemostratigraphic studies.
Stable carbon-isotopes of marine carbonates have been successfullyused to constrain variations of the global carbon cycle through theMesozoic (e.g. Scholle and Arthur, 1980; Weissert, 1989; Jenkyns et al.,2002; Dromart et al., 2003; Hesselbo et al., 2007, etc.). Some studiescontaining data of Aalenian carbon-isotope stratigraphy have beenpublished (Bartolini and Cecca,1999; Bartolini et al., 1999; Jenkyns et al.,2002; Morettini et al., 2002; Hesselbo et al., 2003; O'Dogherty et al.,2006; Sandoval et al., 2004, 2006), most being carried out onstratigraphic sections without good biostratigraphic data or on sectionswhich do not include the lower and upper Aalenian boundaries.Therefore carbon-isotope studies in complete, well developed, andwell-dated sections are necessary.
The aims of this work are multiple: (a) to construct a δ13C curvebased on marine carbonate for the uppermost Toarcian–Aalenianinterval to be considered as a reference for Mediterranean regions;(b) to correlate the δ13C values with highly detailed ammonitebiostratigraphy (standard zones and subzones); (c) to analyse possibleconnections between fluctuations in the δ13C of marine carbonate andbiotic events (ammonite, radiolarians and calcareous nannofossils).For these purposes four sections were selected (Barranco de AguaLarga, Cerro de Mahoma, Cerro Méndez 1 and Cerro Méndez 2), thatrepresent basinal deposits of the western Tethys located on thesouthern Iberian palaeomargin. In these sections ammonites arecommon and relatively well preserved. The biostratigraphic analysisenabled us to recognize the uppermost Toarcian (Aalensis Zone), thecomplete Aalenian (Opalinum, Murchisonae, Bradfordensis and Con-cavum Zones) and the lowermost Bajocian (Discites Zone). Calcareousnannofossils and radiolarians (in discrete beds of the Upper Aalenianand Lower Bajocian) are also common in these Median Subbetichemipelagic sections.
2. Geographic and geologic setting
The sections studied, currently separated by approximately220 km, are located in southern Spain, provinces of Jaén (Barrancode Agua Larga), Granada (Cerro Méndez) and Murcia (Cerro deMahoma). Geologically, they are located in the External Zones of theBetic Cordillera, which extends across S and SE Spain, with a WSW–
ENE trend (Fig. 1). In the Betic External Zones, two main geologicalunits are differentiated: the Prebetic (platform facies) and the
Subbetic (pelagic facies). The physiography of the Subbetic basinwas irregular due to severe Jurassic intracontinental rifting, whichcaused two high swell areas (External and Internal Subbetic) and onerelatively deep trough (Median Subbetic) in the Southern Iberianpalaeomargin (Aurell et al., 2002). Concretely, the Median Subbetic –
where the four studied sections are located – is the only Betic domainwith continuous pelagic or hemipelagic sedimentation throughoutthe stratigraphic interval analysed.
3. Material and analytical laboratory procedure(chemostratigraphical and micropalaeontological)
3.1. Isotopes
A total of 387 samples (202 from Agua Larga, 124 from Cerro deMahoma, 27 from Cerro Méndez 1 and 34 from Cerro Méndez 2) arefrom calcareous beds, covering the uppermost Toarcian–lowermostBajocian interval were analysed for their δ13C. 326 samples (fromAgua Larga and Cerro de Mahoma) were analysed for their δ18O. Allcarbon and oxygen stable isotopic analyses were carried on bulkcarbonate samples. Only the limestones or marly limestones (N80%CaCO3) were selected for analyses. The samples were analysed at theLaboratory of Isotope Geochemistry of the University of Lausanne(Switzerland) following the standard procedure of McCrea (1950).Rock powder samples were reacted at 50 °C for a half-hour. Afractionation factor of 1.00931 between CaCO3 and H3PO4 wasapplied (Swart et al., 1991). Carbonate δ13C values were determinedwith a Finningan MAT 251 multi collector Mass Spectrometer andwith a Finningan Mat delta-S Mass Spectrometer. All δ13C arereported relative to PDB. The working standard was calibratedrelative to NBS-19 δ13CPDB=1.95‰ (Coplen et al., 1983). The standarddeviation (1σ) associated to the analytical error of δ13C values is±0.05‰.
3.2. Calcareous nannofossils
Smear slides were prepared for 134 samples of marls (80 fromAgua Larga section and 54 from Cerro de Mahoma section), taken atirregularly spaced along the interval studied. Sample preparation wasperformed as a simple procedure to retain the original composition ofcalcareous nannofossil assemblages. It consisted of a simple mechan-ical breakage of a small piece of rock, its dilution with water andspreading onto a slide, drying in an oven, and finally mounting withcoverslips using Canada balsam as the adhesive. Special care wastaken to prepare the samples as homogeneously as possible so thatparticle density in different slides would be comparable.
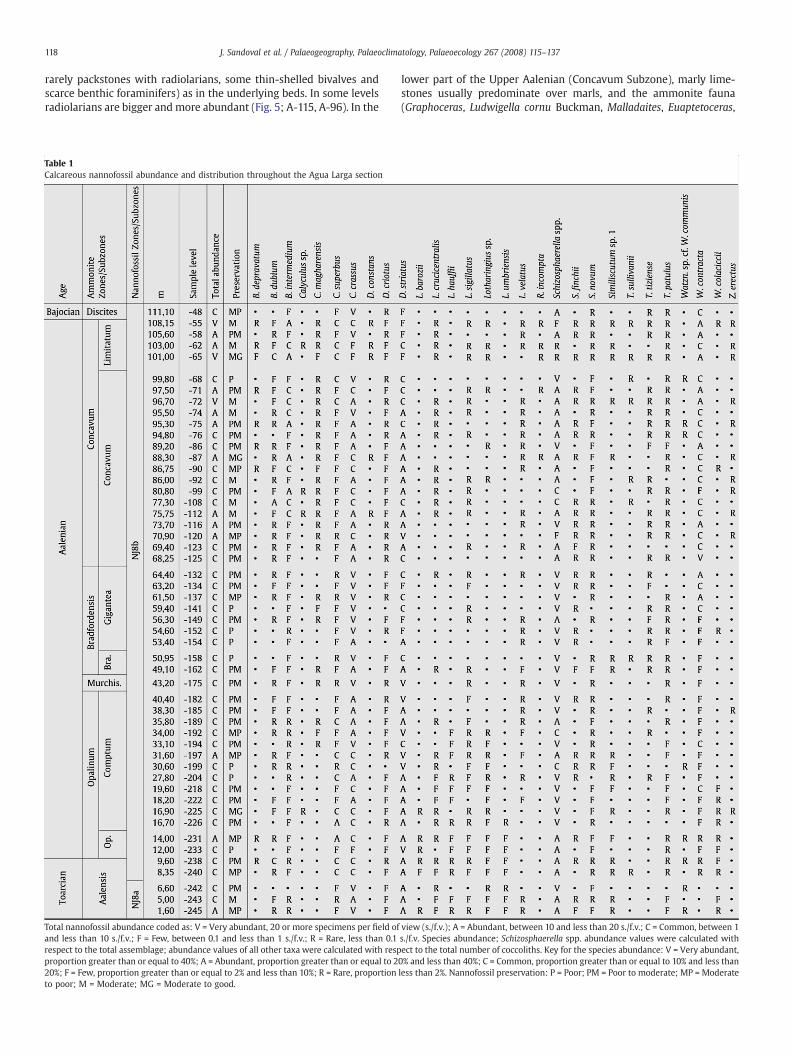
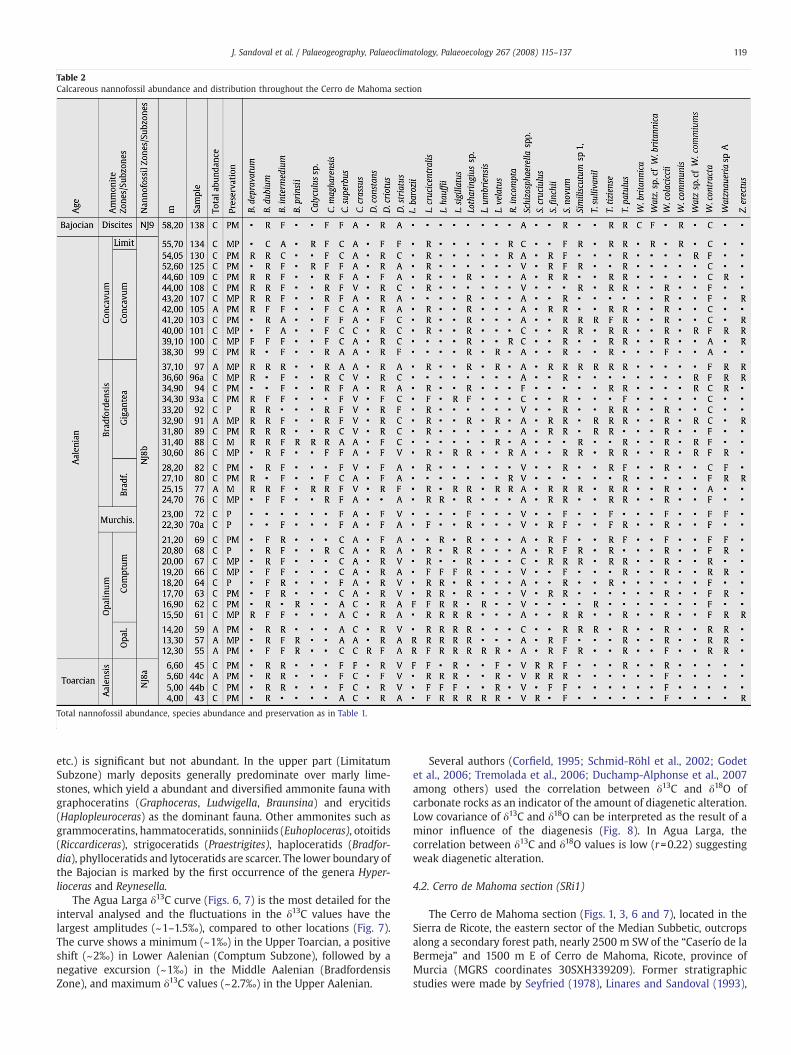
The smear slides were investigated for nannofossil content using apolarizing light microscope at 1200× magnification. The preservationof nannofossils in each sample was evaluated by examination underthe light microscope using an index based on the criteria proposed byRoth and Thierstein (1972). Only 91 samples (51 from Barranco deAgua Larga and 40 from Cerro de Mahoma), representing the bestpreserved and abundant assemblages were retained to perform asemiquantitative study. For this, the nannofossils were counted alonga longitudinal traverse of the smear slide (1 traverse=200 fields ofview). In 5 samples, due to the scarcity of nannofossils, two longi-tudinal traverses were examined to reach an adequate amount forthe correct calculation of percentage. A minimum of 107 (only in 2samples) and up to 1180 nannofossil specimens were counted in eachsample. For Schizosphaerella, only complete isolated valves orfragments representing more than half a valve were counted. Thetotal nannofossil abundance in each sample was calculated bydividing the number of specimens by the number of fields of viewnecessary to count them, and finally expressed as specimens per fieldof view. Percentage values of individual coccolith species werecalculated with respect to the total number of coccoliths. Percentage
Fig. 1. General sketch of the Western Mediterranean region with approximate location of the sections studied in the Betic Cordillera (1, Barranco de Agua Larga; 2, Cerro Méndez;3, Cerro de Mahoma) and in the Umbria–Marche Apennines (4, Presale; 5, Terminilleto).
of Schizosphaerella spp. was calculated based on the total of nan-nofossil specimens (Tables 1 and 2).
4. Analysed sections: lithology, ammonite biostratigraphy andcarbon-isotope stratigraphy
MarI–marly limestone rhythmites were sampled for isotopeanalyses in sections from uppermost Toarcian to lowermost Bajocian.In the sections studied the ammonite fauna has been revised, derivedmainly from the calcareous beds and, studied calcareous nannofossilsof marly beds for the first time in Barranco de Agua Larga and Cerro deMahoma sections. Moreover, in these sections we analysed radiolarianabundances, mainly from calcareous strata.
4.1. Barranco de Agua Larga section (AQ1)
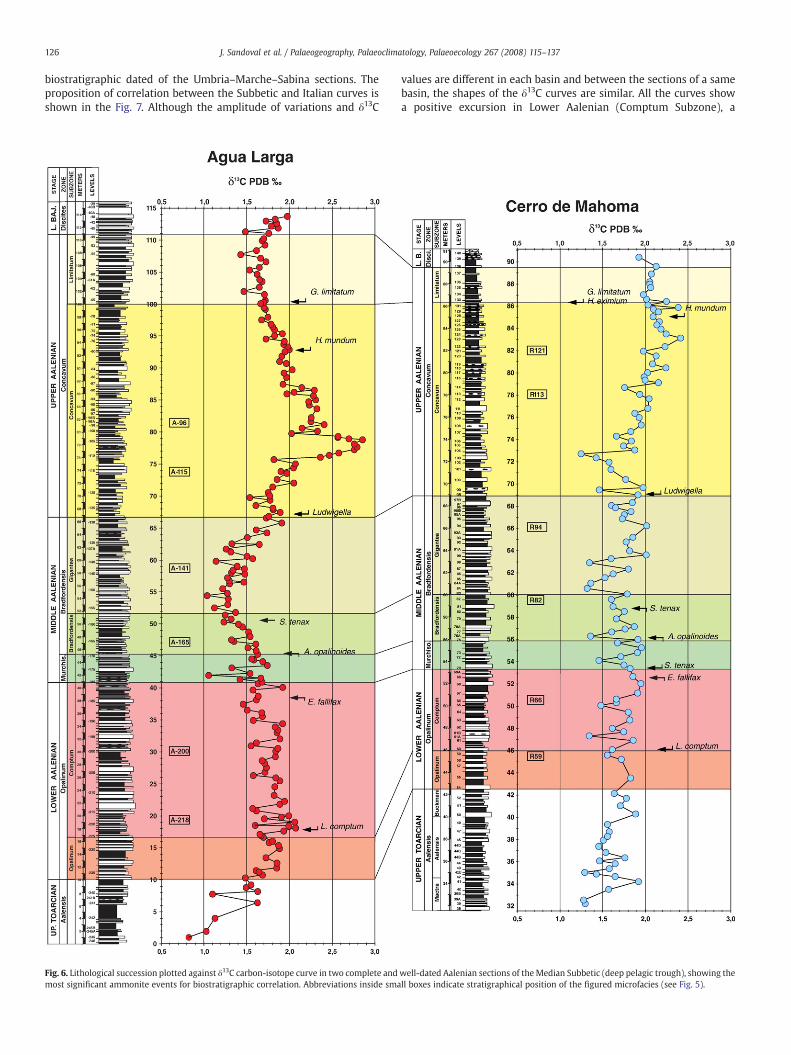
This section (Figs. 1, 2, 6 and 7), located in the central sector of theMedian Subbetic, outcrops along the Agua Larga stream, 500 m NWofthe Cortijo Alorí, near of the village of Campillo de Arenas, Province ofJaén (MGRS coordinates 30SVG514601). Uppermost Aalenian andLower Bajocian sediments of this section were previously studied bySandoval (1983), Linares and Sandoval (1990, 1996), Sandoval andChandler (2000), and by O'Dogherty et al. (2006), but the rest of thesection has not been studied hitherto. In the “Barranco de Agua Larga”we report data of the uppermost Toarcian–lowermost Bajocian(Aalensis–Discites zones) interval. The Toarcian/Aalenian and theAalenian/Bajocian boundaries are well exposed.
The uppermost Toarcian (Aalensis Zone) is 10 m thick (0–10 m),comprising marls with marly limestone interbeds (mudstones to
wackestones with some radiolarians, scarce thin-shelled bivalves,benthic foraminifers, ostracods and embryos of ammonites). Tracefossil genus Chondrites is abundant. Although ammonites are scarce,some typical Late Toarcian taxa – Pleydellia aalensis (Zieten), Cottes-woldia sp., Osperlioceras sp. – give accurate ages.
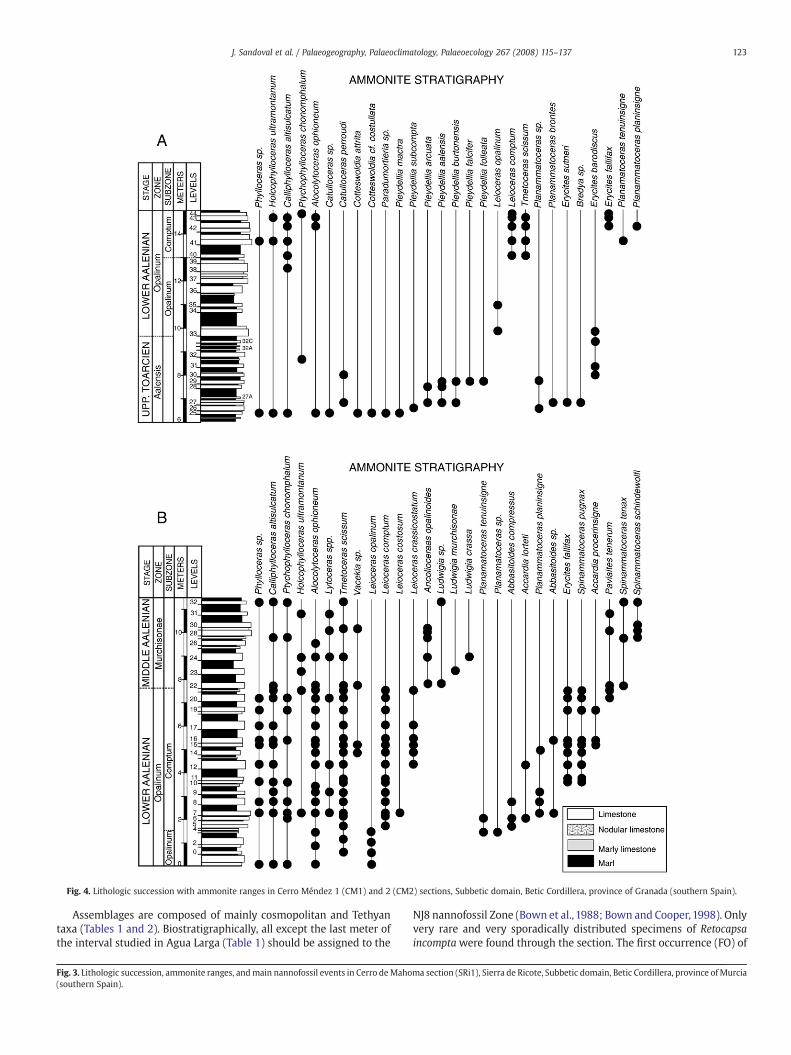
The Lower Aalenian, Opalinum Zone, (~31 m thick; 10–41 m) iscomposed of a rhythmic alternation of darker grey micritic marlylimestones and marls (mudstones and wackestones with radiolarians,some thin-shelled bivalves and scarce benthic foraminifers (Fig. 5;A-218, A-200). Trace fossils, especially Zoophycos and Chondrites, arelocally abundant. Individual thicknesses of the marly limestone andmarly beds range from 10 to 90 cm. Ammonites, although scarce, arehighly representative – Leioceras lineatum Buckman, L. comptum Buck-man, L. crassicostatum Buckman, Tmetoceras scissum (Benecke), Erycitesfallifax Arkell, Vacekia stephensi Buckman – providing accurate biostrati-graphy. Some phylloceratids and lytoceratids also occur.
The Middle Aalenian is approximately 25.8 m thick (41–66.8 m)and shows lithology and microfacies similar to those of the under-lying beds (Fig. 5; A-165, A-141). Radiolarians can be more abundantand better preserved; juvenile ammonites are common. In thisinterval darker marls locally predominate over marly limestones. Theammonites are more abundant and diversified than in the LowerAalenian, showing graphoceratids (Ancolioceras, Ludwigia, Brasilia)together with Tmetoceras, Spinammatoceras, Malladaites, Abbasi-toides, Ambersites, etc. Scarce lytoceratids and phylloceratids are alsopresent. The Middle Aalenian standard ammonite zones of Murchi-sonae and Bradfordensis are present.
The Upper Aalenian (Concavum Zone) is ~44 m thick (66.8–111 m)with similar lithology and microfacies (mudstones to wackestones or
118 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
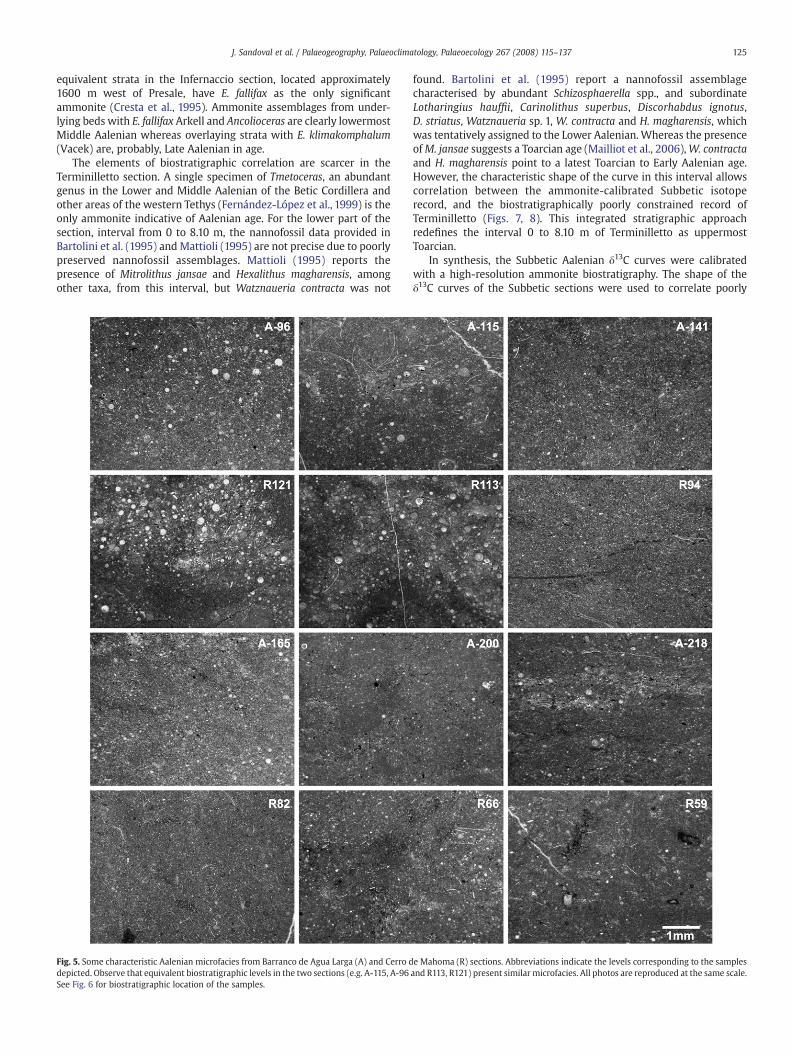
rarely packstones with radiolarians, some thin-shelled bivalves andscarce benthic foraminifers) as in the underlying beds. In some levelsradiolarians are bigger and more abundant (Fig. 5; A-115, A-96). In the
Table 1Calcareous nannofossil abundance and distribution throughout the Agua Larga section
Total nannofossil abundance coded as: V = Very abundant, 20 ormore specimens per field of vless than 10 s./f.v.; F = Few, between 0.1 and less than 1 s./f.v.; R = Rare, less than 0.1 s./f.v. Spectotal assemblage; abundance values of all other taxa were calculated with respect to the togreater than or equal to 40%; A = Abundant, proportion greater than or equal to 20% and less tproportion greater than or equal to 2% and less than 10%; R = Rare, proportion less than 2%.
Table 1Calcareous nannofossil abundance and distribution throughout the Agua Larga section
Total nannofossil abundance coded as: V = Very abundant, 20 or more specimens per field ofand less than 10 s./f.v.; F = Few, between 0.1 and less than 1 s./f.v.; R = Rare, less than 0.1respect to the total assemblage; abundance values of all other taxa were calculated with respproportion greater than or equal to 40%; A = Abundant, proportion greater than or equal to 220%; F = Few, proportion greater than or equal to 2% and less than 10%; R = Rare, proportionto poor; M = Moderate; MG = Moderate to good.
lower part of the Upper Aalenian (Concavum Subzone), marly lime-stones usually predominate over marls, and the ammonite fauna(Graphoceras, Ludwigella cornu Buckman, Malladaites, Euaptetoceras,
iew (s./f.v.); A = Abundant, between 10 and less than 20 s./f.v.; C = Common, between 1 aies abundance; Schizosphaerella spp. abundance values were calculatedwith respect to ttal number of coccoliths. Key for the species abundance: V = Very abundant, proportihan 40%; C = Common, proportion greater than or equal to 10% and less than 20%; F = Fe ,Nannofossil preservation: P = Poor; PM = Poor to moderate; MP = Moderate to poor; M
view (s./f.v.); A = Abundant, between 10 and less than 20 s./f.v.; C = Common, betweens./f.v. Species abundance; Schizosphaerella spp. abundance values were calculated witect to the total number of coccoliths. Key for the species abundance: V = Very abundan0% and less than 40%; C = Common, proportion greater than or equal to 10% and less thaless than 2%. Nannofossil preservation: P = Poor; PM = Poor to moderate; MP = Modera
ndheonw=
1ht,nte
Table 2Calcareous nannofossil abundance and distribution throughout the Cerro de Mahoma section
Total nannofossil abundance, species abundance and preservation as in Table 1.
Table 2Calcareous nannofossil abundance and distribution throughout the Cerro de Mahoma section
Total nannofossil abundance, species abundance and preservation as in Table 1.
etc.) is significant but not abundant. In the upper part (LimitatumSubzone) marly deposits generally predominate over marly lime-stones, which yield a abundant and diversified ammonite fauna withgraphoceratins (Graphoceras, Ludwigella, Braunsina) and erycitids(Haplopleuroceras) as the dominant fauna. Other ammonites such asgrammoceratins, hammatoceratids, sonniniids (Euhoploceras), otoitids(Riccardiceras), strigoceratids (Praestrigites), haploceratids (Bradfor-dia), phylloceratids and lytoceratids are scarcer. The lower boundary ofthe Bajocian is marked by the first occurrence of the genera Hyper-lioceras and Reynesella.
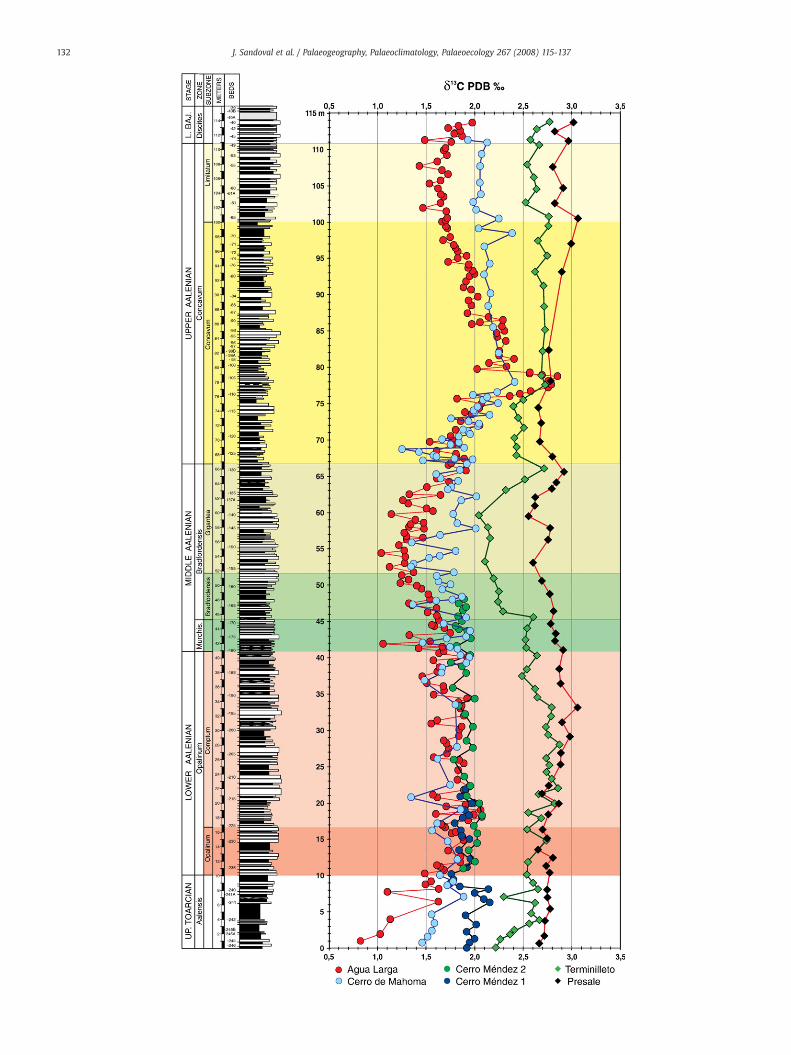
The Agua Larga δ13C curve (Figs. 6, 7) is the most detailed for theinterval analysed and the fluctuations in the δ13C values have thelargest amplitudes (~1–1.5‰), compared to other locations (Fig. 7).The curve shows a minimum (~1‰) in the Upper Toarcian, a positiveshift (~2‰) in Lower Aalenian (Comptum Subzone), followed by anegative excursion (~1‰) in the Middle Aalenian (BradfordensisZone), and maximum δ13C values (~2.7‰) in the Upper Aalenian.
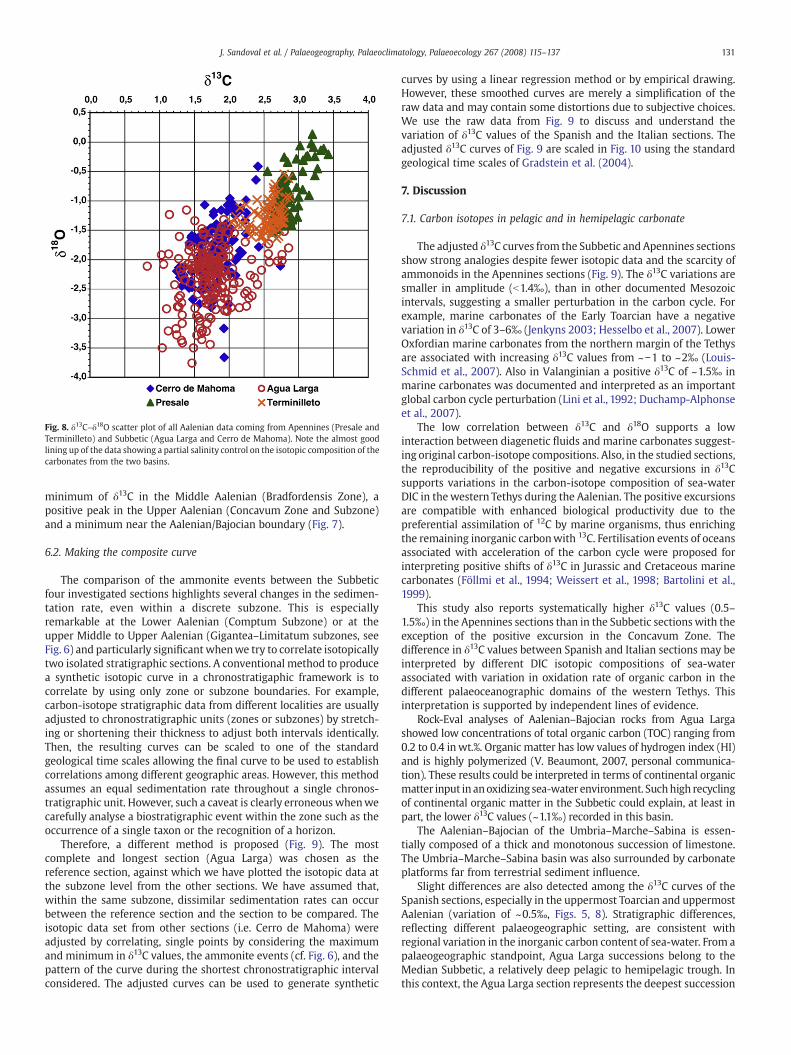
Several authors (Corfield, 1995; Schmid-Röhl et al., 2002; Godetet al., 2006; Tremolada et al., 2006; Duchamp-Alphonse et al., 2007among others) used the correlation between δ13C and δ18O ofcarbonate rocks as an indicator of the amount of diagenetic alteration.Low covariance of δ13C and δ18O can be interpreted as the result of aminor influence of the diagenesis (Fig. 8). In Agua Larga, thecorrelation between δ13C and δ18O values is low (r=0.22) suggestingweak diagenetic alteration.
4.2. Cerro de Mahoma section (SRi1)
The Cerro de Mahoma section (Figs. 1, 3, 6 and 7), located in theSierra de Ricote, the eastern sector of the Median Subbetic, outcropsalong a secondary forest path, nearly 2500 m SW of the “Caserío de laBermeja” and 1500 m E of Cerro de Mahoma, Ricote, province ofMurcia (MGRS coordinates 30SXH339209). Former stratigraphicstudies were made by Seyfried (1978), Linares and Sandoval (1993),
120 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
García-Gómez et al. (1995), O'Dogherty et al. (2001) and Sandovalet al. (2001a). These biostratigraphic studies have been complimentedwith additional sampling and a review of the biostratigraphic data.
The uppermost Toarcian (Aalensis Zone) materials are 11 m (31.5–42.5) thick composed of grey-white marls and marly limestones(mudstones and wackestones with mainly radiolarians, thin-shelledpelagic bivalves, benthic foraminifers and ostracods). Bed thicknessvaries from 5 to 60 cm. Trace fossils, especially Zoophycos and Chon-drites are common. Ammonites, abundant and diversified, aredumortieriins (Pleydellia, Cotteswoldia and Catulloceras), grammocer-atins, harpoceratins, hammatoceratids, erycitids, phylloceratids andlytoceratids (Fig. 3).
Lower Aalenian materials, 10.8 m thick (42.5–53.3 m), are repre-sented by grey marls and marly limestones. The Opalinum Zone andSubzone has similar facies and microfacies to those of the uppermostToarcian (Fig. 5; R59). However, bioturbation (Zoophycos) is morecommon and the ammonites (Leioceras,Holcophylloceras) are scarcer. IntheComptumSubzone the stratigraphic succession ismore calcareous atthe base, with the dominance of compact marly limestones – withradiolarians, thin-shelled pelagic bivalves, ostracods, and benthicforaminifers (Fig. 5; R66) – becoming more marly in the upper part ofthe Lower Aalenian. The ammonite assemblages are dominated byLeioceras (very abundant) and Tmetoceras. Erycitids, hammatoceratids,phylloceratids and lytoceratids are also common forms.
TheMiddle Aalenian rocks (Murchisonae and Bradfordensis zones)are 15.7 m (53.3–69 m) of grey marly limestones and marls(mudstones andwackestones with some radiolarians and thin-shelledpelagic bivalves (Fig. 5; R82, R94). In the Murchisonae Zone, marlyhorizons predominate over marly limestones, which show a bedthickness variation from 10 to 70 cm. The materials of theBradfordensis Zone are slightly more calcareous than in the under-lying levels. The fossil trace Thalassinoides is common in some bedstogether with Chondrites. Ammonites (Fig. 3) such as graphoceratins(Ancolioceras, Ludwigia and Brasilia), erycitids (Erycites, Spinammato-ceras, and Abbasitoides), Tmetoceras, phylloceratids and lytoceratidsare relatively abundant and characteristic.
The Upper Aalenian (Concavum Zone) is 20.3 m thick (69–89.5)and shows similar lithofacies to those recognized in the BradfordensisZone, although limestone beds are more compact, chert nodules arecommon, and Thalassinoides scarcer. Microfacies are wackestones or,rarely, packstones with abundant and well preserved radiolarians andthin-shelled pelagic bivalves (Fig. 5; R113, R121). Ammonites are notabundant, but characteristic taxa (mostly Graphoceras and Haplo-pleuroceras) allow good biostratigraphic results; the Concavum andLimitatum subzones are recognized (Fig. 3). Hyperlioceras sp., bed 138,indicates Lower Bajocian (Discites Zone).
In the lower part of this interval, a sample (bed 108; 75.5 m) hasyielded a rich radiolarian fauna (Laxtorum? jurassicum Isozaki andMatsuda, Palinandromedad sognoensis Baumgartner, Parahsuum oloriziEl Kadiri, and Ristola praemirifusus Baumgartner and Bartolini),allowing recognition of the UAZone 2 of Baumgartner et al. (1995).Likewise, in the upper part (around the boundary of the Concavum–
Limitatum Subzone), a good sample (bed 129; 85.9 m) presents a richand diversified radiolarian content characteristic of UAZone 3 ofBaumgartner et al. (1995) with Parasaturnalis diplocyclis (Yao), Hexa-saturnalis suboblongus (Yao), Eucyrtidiellum unumaense (Yao) and Sellabeniderkoulensis (El Kadiri) among others.
The general pattern of Cerro de Mahoma δ13C curve parallels thoseof Agua Larga, albeit with smaller amplitude of fluctuations (0.8–1.1‰)and more scattered values are observed (Figs. 6, 7 and 8).
For the Cerro de Mahoma data, the correlation between δ13C andδ18O (Fig. 8) is moderate (r=0.47), suggesting some influence of
Fig. 2. Lithologic succession, ammonite stratigraphy, and major nannofossil events in the Ba(southern Spain).
diagenesis (Corfield, 1995; Schmid-Röhl et al., 2002; Godet et al.,2006; Tremolada et al., 2006; Duchamp-Alphonse et al., 2007).
4.3. Cerro Méndez sections (CM1 and CM2)
Cerro Méndez, as Barranco de Agua Larga, is located in the centralsector of the Median Subbetic (Fig. 1).
The Cerro Méndez 1 section (Fig. 4A) outcrops along a secondaryravine of the Guadahortuna River, 3 km E of Alamedilla village andapproximately 700 m S of the peak of Cerro Méndez, Province ofGranada (MGRS coordinates 30SVG813603). A revision of previousbiostratigraphic data of Linares and Sandoval (1993) and of García-Gómez et al. (1995) has been performed. Upper Toarcian–lowermostBajocian (Fallaciosum Zone–Discites zones) interval is well-repre-sented. The only interval having accurate stratigraphic data isanalysed (Fig. 4A). The uppermost Toarcian (Aalensis Zone) isapproximately 3.7 m (6.0–9.7) thick, and the lithology consists ofhemipelagic grey marl–marly limestone alternations with twointercalations of red nodular limestones (mudstones and wackestoneswith mainly thin-shelled bivalves, radiolarians, benthic foraminifers,ammonite embryos, and ostracods). Bed thickness varies from 5 to30 cm. Trace fossils, especially Zoophycos and Chondrites are common.Ammonites are abundant, with diversified and characteristic Dumor-tieriinae (Pleydellia, Cotteswoldia and Catulloceras), hammatoceratids,erycitids, phylloceratids, and lytoceratids. The Lower Aaleniancomprises 5.3 m thick (9.7–15 m) grey marls and marly limestones(wackestones with abundant fine shelled bivalves, radiolarians andscarce benthic foraminifers); bed thickness varies from 10 to 70 cm. Inthe lower part of the Opalinum Subzone, detrital materials predomi-nate, whereas in the upper part of this subzone and in the ComptumSubzone, limestone beds are dominant. In the Opalinum Subzone,bioturbation (Zoophycos) is common and the ammonites are scarce(Leioceras, Erycites, Holcophylloceras). In the Comptum Subzone,ammonites, more abundant, are dominated by Leioceras and Tmeto-ceras, and erycitids, hammatoceratids, phylloceratids, and lytoceratidsare also commons.
The Cerro Méndez 2 section (Fig. 4B) is located along theGuadahortuna River, approximately 700 m N of Cerro Méndez, 1(MGRS coordinates 30S814608). Previous biostratigraphic datareported by Linares and Sandoval (1993) are here revised. The LowerAalenian (Opalinum Zone) and the Middle Aalenian (MurchisonaeZone) are represented. Lithology and microfacies are similar to thoseof CM1, and ammonites are more abundant, especially in theComptum Subzone.
In the Upper Toarcian of Cerro Mendez 1, δ13C values are higherthan those of Agua Larga and Cerro de Mahoma (Fig. 7). The valuesdecrease from 2.2 to 1.7‰ in the uppermost part of the Toarcian. In theLower and Middle Aalenian, the Cerro Méndez 1 and 2 δ13C values(Fig. 7) correlate well with those of Agua Larga and Cerro de Mahoma.The δ13C values fluctuate less and remain around 2‰ in the lowerAalenian and slightly decrease to ~1.7‰ in the Middle Aalenian.
5. Nannofossil and radiolarian biostratigraphy
5.1. Nannofossils
Calcareous nannofossils, studied only from Barranco de Agua Largaand Cerro de Mahoma, are irregularly distributed, being rare toabundant, throughout the two sections. Measured abundances in theBarranco de Agua Larga section vary from 59.2 to 2.8 (average 12.7)nannoliths/10 fields of view at 1200×, and from 39.7 to 2.8 (average14.4) in Cerro de Mahoma.
rranco de Agua Larga section (AQ1), Subbetic domain, Betic Cordillera, province of Jaén
122 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
Fig. 4. Lithologic succession with ammonite ranges in Cerro Méndez 1 (CM1) and 2 (CM2) sections, Subbetic domain, Betic Cordillera, province of Granada (southern Spain).
Assemblages are composed of mainly cosmopolitan and Tethyantaxa (Tables 1 and 2). Biostratigraphically, all except the last meter ofthe interval studied in Agua Larga (Table 1) should be assigned to the
Fig. 3. Lithologic succession, ammonite ranges, andmain nannofossil events in Cerro deMaho(southern Spain).
NJ8 nannofossil Zone (Bown et al., 1988; Bown and Cooper,1998). Onlyvery rare and very sporadically distributed specimens of Retocapsaincomptawere found through the section. The first occurrence (FO) of
ma section (SRi1), Sierra de Ricote, Subbetic domain, Betic Cordillera, province ofMurcia
124 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
Watznaueria contracta (sample −240) was detected near the base of theinterval studied, within the uppermost Toarcian (Fig. 2). Consequently,mostof the interval studied inAgua Larga shouldbe assigned to theNJ8bSubzone (Bown et al., 1988). The successive FOs of Triscutum tiziense(sample −204) and Carinolithus magharensis (sample −194) wereregistered in the Lower Aalenian, within the Comptum Subzone(Fig. 2). The uppermost part of the Limitatum Subzone and lowermostpart of the Discites Zone (Aalenian–Bajocian transition) is characterizedby the record of some forms ofWatznaueria transitional toW. britannica,although the trueW. britannica specimenswere found in a sample (−40)only 1 m below the top of the interval (Fig. 2). This last 1 m should thenbe assigned to the NJ9 nannofossil Zone.
The biostratigraphic results from the Cerro de Mahoma section(Table 2) are highly consistent with those from Agua Larga. AsR. incompta is an extremely rare species also in Cerro de Mahoma, thelowermost part of the interval studied was, tentatively, assigned to theNJ8a nannofossil Subzone. The FO of W. contracta (base of the NBJ8bnannofossil Zone) was observed in sample 55, coinciding approxi-mately with the Toarcian/Aalenian boundary. However, a slightlylower (uppermost Aalenian) FO of this species in Cerro de Mahomacannot be ruled out because of the very poor preservation of thesamples from this interval. Within the Lower Aalenian (ComptumSubzone), the successive FOs of T. tiziense (sample 64) andC. magharensis (sample 68), were observed. Finally, the FO of true W.britannica (sample 138) was observed at the base of the Discites Zone,nearly coinciding with the Aalenian/Bajocian boundary (Fig. 3). As aresult, the last 1 m of the succession studied was assigned to the NJ9nannofossil Zone. As in Agua Larga, nearly the whole interval studiedof the Cerro de Mahoma section should be assigned to the NJ8bnannofossil Subzone.
The Tethyan zonation proposed by Mattioli and Erba (1999) couldonly be partially applied owing to the fact that C. margerelii was notfound in the Aalenian of Agua Larga section.
5.2. Radiolarians
A complete analysis of microfacies in the Barranco de Agua Largasection (samples A) and in the Cerro de Mahoma section (samples R)was performed. About one hundred thin sections were examined forsedimentological and palaeontological features. Radiolarians andthin-shelled pelagic bivalves (probably Bositra sp.) are the two maincomponents of the microfacies throughout the Aalenian. Bioturbation(mainly Chondrites) is quite common throughout the samples. Inparticular, preservation of radiolarian skeletons is enhanced inside themicro-bioturbated areas (see Fig. 5, sample R121). Levels of concen-tration by winnowing due to bottom currents or turbiditic processeswere not recognized. Only a few finely laminated levels wereidentified in the Lower Aalenian.
Thin section analysis also shows regular distribution of radiolar-ians through the studied interval, with increasing upward abundancefrom the upper part of the Middle Aalenian in both sections (Fig. 10).Radiolarian abundances were established from visual percentageestimation by means of Baccelle and Bosellini (1965) comparisoncharts. The values range from 1 to 40%. For the Aalenian interval in theSubbetic basin radiolarians represent less than 10% of the originalsediment volume, except for the Gigantea to Concavum subzones,where their represent up to an estimated average of 30–40%. Theradiolarian abundance increase coincides with the positive excursionrecorded in all the investigated sections (Fig. 10).
In general, the Lower Aalenian shows a dominance of thin-shelledbivalves over radiolarians (Fig. 5). This trend is reversed progressivelythroughout the Middle Aalenian, where radiolarians are the dominantskeletal component. Above this level, radiolarians are abundant excepton those discrete levels where radiolarian abundance falls and thin-shelled pelagic bivalves occur again (i.e. at the boundary of theBradfordensis–Gigantea subzones).
Despite the large abundance of radiolarians, the preservation israther poor because the skeletons are commonly calcified. Some-times, especially in the Middle Aalenian, the skeletons are socalcified that they occur as radiolarian ghosts, making it difficult todistinguish them from the lime matrix. Only in the upper part ofthe Cerro de Mahoma section do the radiolarians become quiteabundant. The occurrence of chert nodules (Fig. 3) improvespreservation, allowing an acid processing treatment for taxonomicdetermination. In the Barranco de Agua Larga section, no chertnodules were visible (Fig. 2), and, consequently radiolarian extrac-tion was unsuccessful; however, a similar trend in the radiolarianabundance was noted (see comparison on Fig. 10). Such observa-tions, together the similar pattern of the abundance curves betweenthe two sections clearly point to a primary control in the abundancefluctuation recorded.
6. Biostratigraphic correlations and carbon-isotope curves inwestern Tethys
6.1. Stratigraphic correlations between Subbetic and Apennines sections
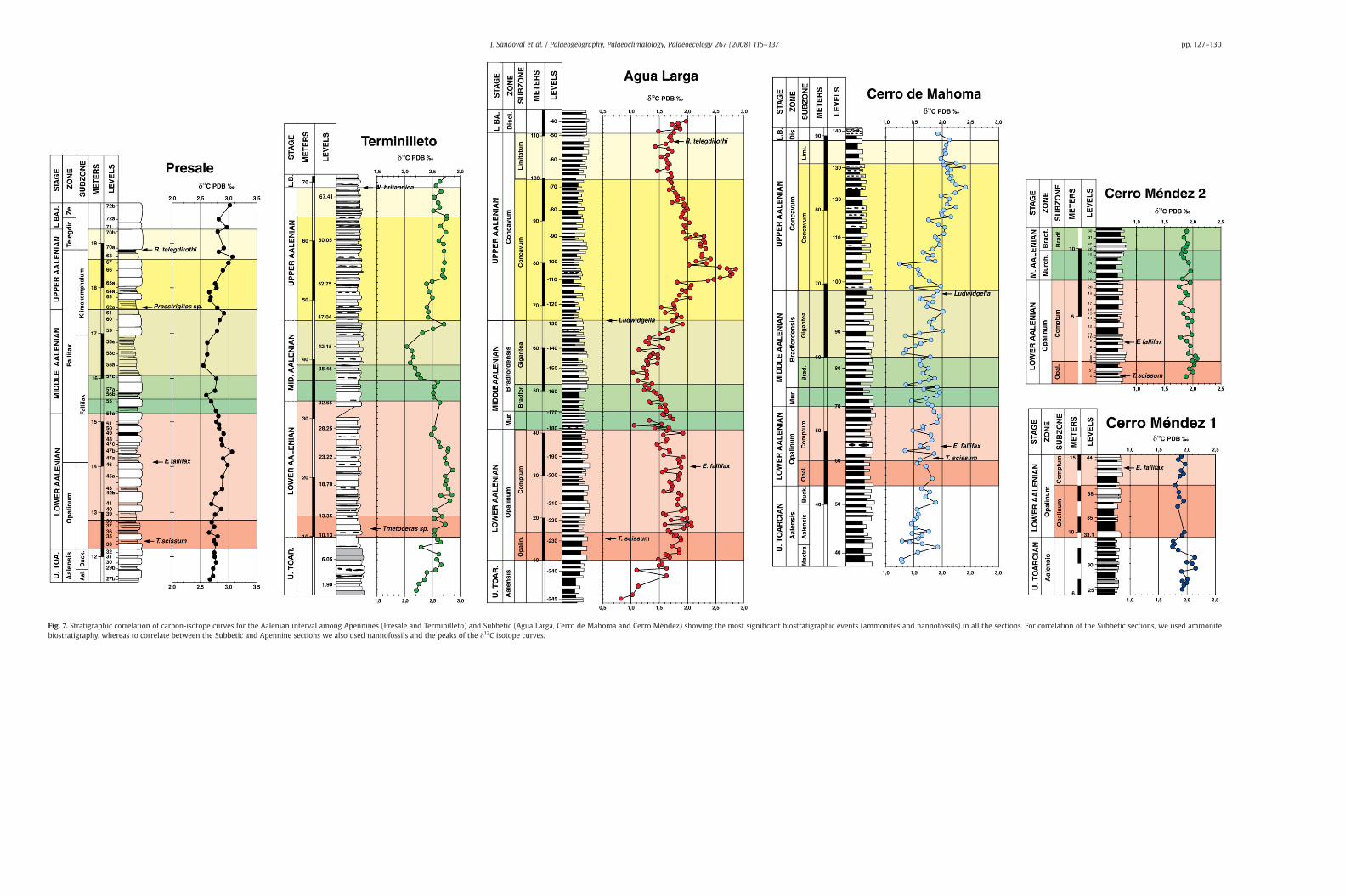
The Betic sections have an accurate ammonite biostratigraphy (seeSection 4) which enables the correlation of isotopic data with highconfidence throughout the studied sections. Fig. 6 shows thebiostratigraphic correlation between the most complete records ofthe Aalenian of the Median Subbetic. In this figure the correlation isdefined at the ammonite subzone level and the most importantammonite events are also indicated. These data are used to correlatethe biostratigraphic and isotopic correlations among the four Spanishsections analysed and to compare our result to those reported for theAalenian in the Umbria–Marche Apennines (Bartolini et al., 1995;Bartolini and Cecca 1999).
Stratigraphic correlation of the Subbetic carbon-isotope curveswith others previously established in the Aalenian of western Tethys(Presale and Terminilletto, Central Italy) presents some problems,primarily due to poor biostratigraphic markers in the Italian sections,mainly in the Terminilletto. In the Presale section, Cresta et al. (1995)identified and proposed five biohorizons for this interval: the bio-zones of Leioceras opalinum, Erycites fallifax – this divided into twosubzones, E. fallifax and Euaptetoceras klimacomphallum – and Doci-doceras telegdirothi; however the Aalenian stage was not divided insub-stages.
Linares and Sandoval (1993) and our observations (Figs. 2–4) showthat the typical Erycites fallifax Arkell, which occurs in the beds 47 and48 of the Presale section, is a species abundant in many sections of theBetic Cordillera, where it is documented from the uppermost LowerAalenian (Comptum Subzone) and, possibly, does not surpass thelowermost Middle Aalenian. By contract, other Erycites species extendto the Bradfordensis Zone (Linares and Sandoval, 1993). Euaptetocerasklimakomphalum (Vacek), which occurs in bed 59 of Presale, wasrecorded from the Middle Aalenian, but this species is particularlyabundant in the Upper Aalenian and lowermost Lower Bajocian ofsome Subbetic sections (Linares and Sandoval, 1993). Praestrigites,from the bed 62 of Presale, is a typical genus of the Upper Aalenian andLower Bajocian of some west Mediterranean localities. Riccardicerastelegdirothi (Prinz), which occurs in beds 70–72 of the Presale section,is common in the uppermost Aalenian and lowermost Bajocian ofmany well-documented Mediterranean localities (Cresta and Galácz,1990; Linares and Sandoval, 1993; Sandoval and Chandler, 2000;Sandoval et al., 2000).
Another problem is the possible stratigraphic unconformitysuspected at the Presale section near the Middle/Upper Aalenianboundary. In bed 58 (see Fig. 7) a sharp change in the lithology isobserved, changing from limestones to centimetre alternations ofnodular marly limestones and marls (“unita multistrato” in Crestaet al., 1995), which do not contain significant ammonites. However,
equivalent strata in the Infernaccio section, located approximately1600 m west of Presale, have E. fallifax as the only significantammonite (Cresta et al., 1995). Ammonite assemblages from under-lying beds with E. fallifax Arkell and Ancolioceras are clearly lowermostMiddle Aalenian whereas overlaying strata with E. klimakomphalum(Vacek) are, probably, Late Aalenian in age.
The elements of biostratigraphic correlation are scarcer in theTerminilletto section. A single specimen of Tmetoceras, an abundantgenus in the Lower and Middle Aalenian of the Betic Cordillera andother areas of the western Tethys (Fernández-López et al., 1999) is theonly ammonite indicative of Aalenian age. For the lower part of thesection, interval from 0 to 8.10 m, the nannofossil data provided inBartolini et al. (1995) and Mattioli (1995) are not precise due to poorlypreserved nannofossil assemblages. Mattioli (1995) reports thepresence of Mitrolithus jansae and Hexalithus magharensis, amongother taxa, from this interval, but Watznaueria contracta was not
Fig. 5. Some characteristic Aalenian microfacies from Barranco de Agua Larga (A) and Cerro ddepicted. Observe that equivalent biostratigraphic levels in the two sections (e.g. A-115, A-96See Fig. 6 for biostratigraphic location of the samples.
found. Bartolini et al. (1995) report a nannofossil assemblagecharacterised by abundant Schizosphaerella spp., and subordinateLotharingius hauffii, Carinolithus superbus, Discorhabdus ignotus,D. striatus, Watznaueria sp. 1, W. contracta and H. magharensis, whichwas tentatively assigned to the Lower Aalenian.Whereas the presenceofM. jansae suggests a Toarcian age (Mailliot et al., 2006),W. contractaand H. magharensis point to a latest Toarcian to Early Aalenian age.However, the characteristic shape of the curve in this interval allowscorrelation between the ammonite-calibrated Subbetic isotoperecord, and the biostratigraphically poorly constrained record ofTerminilletto (Figs. 7, 8). This integrated stratigraphic approachredefines the interval 0 to 8.10 m of Terminilletto as uppermostToarcian.
In synthesis, the Subbetic Aalenian δ13C curves were calibratedwith a high-resolution ammonite biostratigraphy. The shape of theδ13C curves of the Subbetic sections were used to correlate poorly
e Mahoma (R) sections. Abbreviations indicate the levels corresponding to the samplesand R113, R121) present similar microfacies. All photos are reproduced at the same scale.
126 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
biostratigraphic dated of the Umbria–Marche–Sabina sections. Theproposition of correlation between the Subbetic and Italian curves isshown in the Fig. 7. Although the amplitude of variations and δ13C
Fig. 6. Lithological succession plotted against δ13C carbon-isotope curve in two complete andmost significant ammonite events for biostratigraphic correlation. Abbreviations inside sma
values are different in each basin and between the sections of a samebasin, the shapes of the δ13C curves are similar. All the curves showa positive excursion in Lower Aalenian (Comptum Subzone), a
well-dated Aalenian sections of theMedian Subbetic (deep pelagic trough), showing thell boxes indicate stratigraphical position of the figured microfacies (see Fig. 5).
Fig. 7. Stratigraphic correlation of carbon-isotope curves for the Aalenian interval among Apennines (Presale and Terminilleto) and Subbetic (Agua Larga, Cerro de Mahoma and Cerro Méndez) showing the most significant biostratigraphic events (ammonites and nannofossils) in all the sections. For correlation of the Subbetic sections, we used ammonitebiostratigraphy, whereas to correlate between the Subbetic and Apennine sections we also used nannofossils and the peaks of the δ13C isotope curves.
pp. 127–130J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115–137
Fig. 8. δ13C–δ18O scatter plot of all Aalenian data coming from Apennines (Presale andTerminilleto) and Subbetic (Agua Larga and Cerro de Mahoma). Note the almost goodlining up of the data showing a partial salinity control on the isotopic composition of thecarbonates from the two basins.
minimum of δ13C in the Middle Aalenian (Bradfordensis Zone), apositive peak in the Upper Aalenian (Concavum Zone and Subzone)and a minimum near the Aalenian/Bajocian boundary (Fig. 7).
6.2. Making the composite curve
The comparison of the ammonite events between the Subbeticfour investigated sections highlights several changes in the sedimen-tation rate, even within a discrete subzone. This is especiallyremarkable at the Lower Aalenian (Comptum Subzone) or at theupper Middle to Upper Aalenian (Gigantea–Limitatum subzones, seeFig. 6) and particularly significant whenwe try to correlate isotopicallytwo isolated stratigraphic sections. A conventional method to producea synthetic isotopic curve in a chronostratigaphic framework is tocorrelate by using only zone or subzone boundaries. For example,carbon-isotope stratigraphic data from different localities are usuallyadjusted to chronostratigraphic units (zones or subzones) by stretch-ing or shortening their thickness to adjust both intervals identically.Then, the resulting curves can be scaled to one of the standardgeological time scales allowing the final curve to be used to establishcorrelations among different geographic areas. However, this methodassumes an equal sedimentation rate throughout a single chronos-tratigraphic unit. However, such a caveat is clearly erroneous whenwecarefully analyse a biostratigraphic event within the zone such as theoccurrence of a single taxon or the recognition of a horizon.
Therefore, a different method is proposed (Fig. 9). The mostcomplete and longest section (Agua Larga) was chosen as thereference section, against which we have plotted the isotopic data atthe subzone level from the other sections. We have assumed that,within the same subzone, dissimilar sedimentation rates can occurbetween the reference section and the section to be compared. Theisotopic data set from other sections (i.e. Cerro de Mahoma) wereadjusted by correlating, single points by considering the maximumand minimum in δ13C values, the ammonite events (cf. Fig. 6), and thepattern of the curve during the shortest chronostratigraphic intervalconsidered. The adjusted curves can be used to generate synthetic
curves by using a linear regression method or by empirical drawing.However, these smoothed curves are merely a simplification of theraw data and may contain some distortions due to subjective choices.We use the raw data from Fig. 9 to discuss and understand thevariation of δ13C values of the Spanish and the Italian sections. Theadjusted δ13C curves of Fig. 9 are scaled in Fig. 10 using the standardgeological time scales of Gradstein et al. (2004).
7. Discussion
7.1. Carbon isotopes in pelagic and in hemipelagic carbonate
The adjusted δ13C curves from the Subbetic and Apennines sectionsshow strong analogies despite fewer isotopic data and the scarcity ofammonoids in the Apennines sections (Fig. 9). The δ13C variations aresmaller in amplitude (b1.4‰), than in other documented Mesozoicintervals, suggesting a smaller perturbation in the carbon cycle. Forexample, marine carbonates of the Early Toarcian have a negativevariation in δ13C of 3–6‰ (Jenkyns 2003; Hesselbo et al., 2007). LowerOxfordian marine carbonates from the northern margin of the Tethysare associated with increasing δ13C values from ~−1 to ~2‰ (Louis-Schmid et al., 2007). Also in Valanginian a positive δ13C of ~1.5‰ inmarine carbonates was documented and interpreted as an importantglobal carbon cycle perturbation (Lini et al., 1992; Duchamp-Alphonseet al., 2007).
The low correlation between δ13C and δ18O supports a lowinteraction between diagenetic fluids and marine carbonates suggest-ing original carbon-isotope compositions. Also, in the studied sections,the reproducibility of the positive and negative excursions in δ13Csupports variations in the carbon-isotope composition of sea-waterDIC in thewestern Tethys during the Aalenian. The positive excursionsare compatible with enhanced biological productivity due to thepreferential assimilation of 12C by marine organisms, thus enrichingthe remaining inorganic carbonwith 13C. Fertilisation events of oceansassociated with acceleration of the carbon cycle were proposed forinterpreting positive shifts of δ13C in Jurassic and Cretaceous marinecarbonates (Föllmi et al., 1994; Weissert et al., 1998; Bartolini et al.,1999).
This study also reports systematically higher δ13C values (0.5–1.5‰) in the Apennines sections than in the Subbetic sections with theexception of the positive excursion in the Concavum Zone. Thedifference in δ13C values between Spanish and Italian sections may beinterpreted by different DIC isotopic compositions of sea-waterassociated with variation in oxidation rate of organic carbon in thedifferent palaeoceanographic domains of the western Tethys. Thisinterpretation is supported by independent lines of evidence.
Rock-Eval analyses of Aalenian–Bajocian rocks from Agua Largashowed low concentrations of total organic carbon (TOC) ranging from0.2 to 0.4 inwt.%. Organic matter has low values of hydrogen index (HI)and is highly polymerized (V. Beaumont, 2007, personal communica-tion). These results could be interpreted in terms of continental organicmatter input in anoxidizing sea-water environment. Suchhigh recyclingof continental organic matter in the Subbetic could explain, at least inpart, the lower δ13C values (~1.1‰) recorded in this basin.
The Aalenian–Bajocian of the Umbria–Marche–Sabina is essen-tially composed of a thick and monotonous succession of limestone.The Umbria–Marche–Sabina basin was also surrounded by carbonateplatforms far from terrestrial sediment influence.
Slight differences are also detected among the δ13C curves of theSpanish sections, especially in the uppermost Toarcian and uppermostAalenian (variation of ~0.5‰, Figs. 5, 8). Stratigraphic differences,reflecting different palaeogeographic setting, are consistent withregional variation in the inorganic carbon content of sea-water. From apalaeogeographic standpoint, Agua Larga successions belong to theMedian Subbetic, a relatively deep pelagic to hemipelagic trough. Inthis context, the Agua Larga section represents the deepest succession
132 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115-137
with high sedimentation rates and was located in a central position ofthe basin. The Cerro de Mahoma is also a deep succession locatedcloser to the external Subbetic, characterized by lower thickness andlime fraction dominating rather than marls. In addition, chertylimestones are quite frequent in the Upper Aalenian. Thus, palaeogeo-graphic, stratigraphic successions and higher δ13C values are con-sistent with higher biological productivity and/or a lower oxidationrate of organic carbon in Cerro de Mahoma than in Agua Larga.
The offset in the δ13C values between the studied sectionscorresponds to variation of carbon-isotope composition of sea-waterDIC, probably associated with changes in the oxidation rate of organiccarbon. During the Upper Aalenian, δ13C values of marine carbonatesin the Subbetic and Apennines become similar and correspond to amajor transgressive phase and high biological productivity (Fig. 10).Both the δ13C values and the major transgressive phase are compatiblewith a mixing of sea-water DIC reservoirs, probably associated with achange in oceanic circulation and a better mixing of sea-water massesin the western Tethys.
7.2. Carbon-isotope changes, sea-level and biotic turnovers
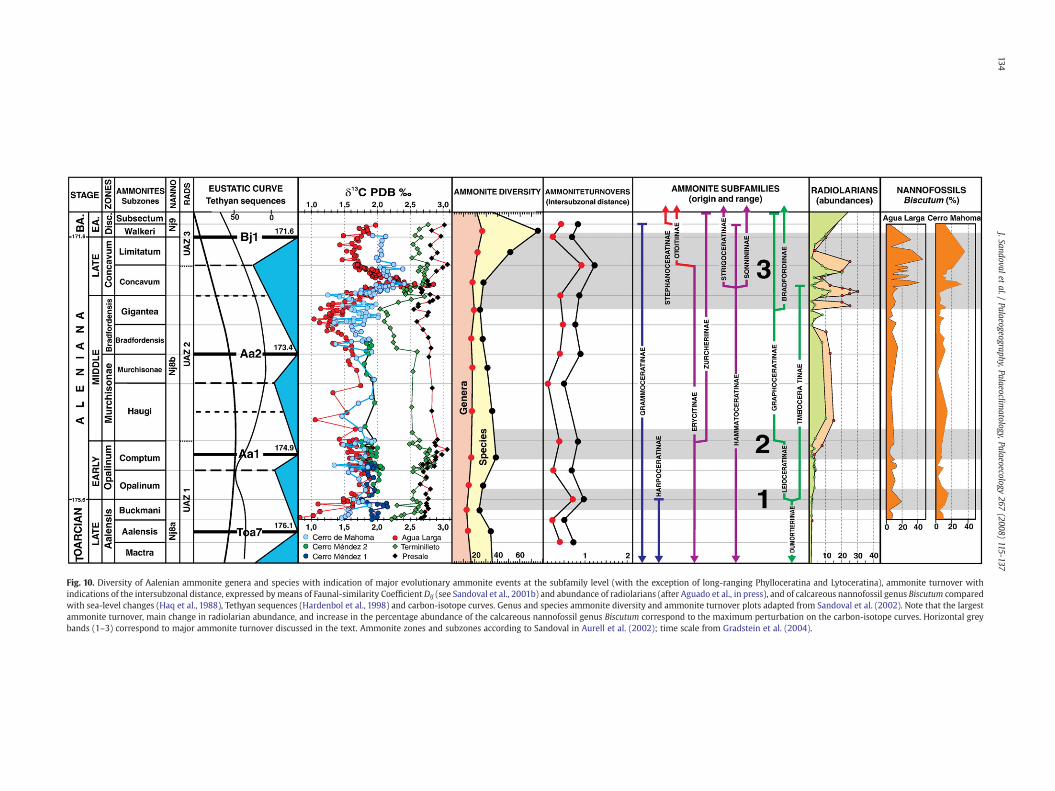
7.2.1. AmmonitesSandoval et al. (2001b, 2002) reported a detailed analysis of the
ammonite diversity and turnover during Toarcian–Early Bajocianinterval, with data from Betic Cordillera and other western Tethyslocalities. In these papers, ammonite diversity and faunal-turnoverwere correlated with sea-level changes (Haq et al., 1988; Hallam,1988,1990) and Tethyan eustatic cycles (Hardenbol et al., 1998). Thesestudies suggest that sea-levels changes and corresponding marinetransgressions and regressions were the principal cause for ammoniteturnover. Likewise, O'Dogherty et al. (2006) analysed carbon cycleperturbations andmajor evolutionary events that took place inMiddleJurassic ammonites. They showed that the main Middle Jurassicammonite turnover coincided with intervals of rapid shifts to low δ13Cvalues.
Data from Sandoval et al. (2002) on ammonite turnover (inter-subzonal distance, expressed by means of the Faunal-similarityCoefficient, Dij; see Sandoval et al., 2001b, p. 316) and diversity atthe generic and specific levels are revised and discussed here. Somesignificant ammonite turnover events took place during the intervalstudied (Fig. 10). The first significant turnover place during the latestToarcian–Early Aalenian. In this interval, most typical Late Toarcianammonites (the subfamilies Harpoceratinae and Dumortieriinae andmost of Grammoceratinae) went extinct. These were replaced byLeioceratinae and Tmetoceratinae, which had their first records in theearliest Aalenian and which together with Phylloceratina andLytoceratinawere the dominant ammonites during the Early Aalenian.The Dij values are relatively high at generic (Dij~0.7) and specific(Dij~1) levels between the Buckmani and Opalinum subzones (Fig.10).The extinction events dominated during the Late Toarcian, coincidingwith relatively low δ13C values, whereas maximum diversification oftypical Early Aalenian forms took place in the Comptum Subzone,coinciding with a moderate positive excursion of the δ13C (Fig. 10).
In the Early–Middle Aalenian transition the Leioceratinae give wayto Graphoceratinae and, moreover, are the first significant records oftypical Mediterranean Erycitidae (Zurcheriinae and Abbasitoides). Thisminor turnover (Dij~0.8 at a specific level between the Comptum andHaugi Subzones) coincides with an interval of fluctuation in the δ13Ccurve and with the beginning of a negative excursion (Fig. 10).
Fig. 9. Synthetic set of correlated δ13C isotopic curve. We present a set of isotopic curves calibrframework of reference (see procedure in the text). If we consider the isotopic signal within ainterval from another section by moving the stratigraphic positions of their isotopic values wcorrelation calibrated by ammonite events. This method minimizes miscorrelation of the issedimentation rate.
The major ammonite turnover of the interval studied is recogniz-able between the latest Middle Aalenian and the earliest LateAalenian. Many Early–Middle Aalenian genera (Vacekia, Tmetoceras,Ludwigia, Brasilia, Planammatoceras, Paviaites, Erycites, Spinammato-ceras, Malladaites, etc.) went extinct whereas new groups such asbradfordiins, strigoceratids, sonniniids, otoitids, as well as newgraphoceratids, erycitids and hammatoceratids originated and spread.This turnover is especially significant between the Concavum andLimitatum subzones with high Dij values at generic (Dij~1,) andspecific (Dij~1.3) levels. Extinctions dominated over originations inthe latest Middle Aalenian, coinciding with low δ13C values, andoriginations proliferated in the Late Aalenian which led to maximumammonite diversity in the Limitatum Subzone (Fig. 10). Low δ13Cvalues near the Aalenian/Bajocian boundary and the latest EarlyBajocian δ13C positive excursion coincided with most important Mid-Jurassic ammonite replacement (O'Dogherty et al., 2006).
7.2.2. NannofossilsAnalysis of the fluctuations in the abundance of some selected taxa
with special palaeoceanographic significance (mainly Biscutum, Cre-pidolithus crassus and Schizosphaerella spp.), together with thevariations in the δ13C values provide evidence (Aguado et al., inpress) that during the Early–Middle Aalenian, stable mesotrophic tooligotrophic conditions prevailed in surface waters of the Subbeticbasin. According to Bown et al. (2004) and Erba (2006), calcareousnannoplankton are mesotrophic to oligotrophic, dominating warmand stable environments. Diversification should then be favored bylow-nutrient availability and stable conditions. During the LateToarcian–Early Aalenian, the nannofossil records of the two sectionsshow an origination phase, characterized by the successive FOs ofWatznaueria contracta, Triscutum tiziense, and Carinolithus maghar-ensis, which probably was a consequence of this period of stability andoligotrophic conditions in surface waters. Also, the most outstandingfeature within the nannofossil assemblages throughout the LateAalenian–Early Bajocian interval, the evolutionary transition betweenLotharingius and Watznaueria (Bown et al., 1988; Cobianchi et al.,1992; Mattioli, 1995; Bown and Cooper, 1998; Mattioli and Erba, 1999;Perilli et al., 2002; Erba, 2006; Sandoval et al., 2006, Aguado et al., inpress), could be related to this period of oligotrophic and stableconditions. According to Roth (1987, 1989) scarcity of nutrientsinduces competition and probably some Lotharingius species andW. contracta occupied similar ecologic niches.
During the Late Aalenian, the nannofossil assemblages arecharacterized by high proportions of Biscutum (Fig. 10), which iscommonly regarded as an eutrophic taxon, and low proportions ofC. crassus and Schizosphaerella, that are considered oligotrophic(Aguado et al., in press). This indicates that the palaeoceanographicconditions changed in the Late Aalenian, when meso- to eutrophicconditions developed in the surface waters of South IberianPalaeomargin, probably as a consequence of more vigorous upwelling.This upwelling could be a consequence of palaeotectonic modifica-tions in the palaeogeography of the western Tethys in relation to theAtlantic opening (Bill et al., 2001). According to Roth (1987, 1989),nutrient availability was an important factor in the evolution ofcalcareous nannoplankton in Mesozoic oceans. Availability of limitingnutrients related with high fertility and more vigorous water mixing,probably resulted in low-diversity nannoplankton assemblages andextinctions (Roth, 1987, 1989; Bown et al., 2004; Erba, 2006). Duringthis interval, the Agua Larga and Cerro de Mahoma nannofossil
ated subzone by subzone by using the isotopic signal at the Agua Larga section as a basicsubzone in the reference section to be locked values, we proceed to adjust the equivalentith respect to the reference section. The translation of points is based on a peak to peakotopic signal within a same subzone when we compare a set of sections with different
Fig. 10. Diversity of Aalenian ammonite genera and species with indication of major evolutionary ammonite events at the subfamily level (with the exception of long-ranging Phylloceratina and Lytoceratina), ammonite turnover withindications of the intersubzonal distance, expressed bymeans of Faunal-similarity Coefficient Dij (see Sandoval et al., 2001b) and abundance of radiolarians (after Aguado et al., in press), and of calcareous nannofossil genus Biscutum comparedwith sea-level changes (Haq et al., 1988), Tethyan sequences (Hardenbol et al., 1998) and carbon-isotope curves. Genus and species ammonite diversity and ammonite turnover plots adapted from Sandoval et al. (2002). Note that the largestammonite turnover, main change in radiolarian abundance, and increase in the percentage abundance of the calcareous nannofossil genus Biscutum correspond to the maximum perturbation on the carbon-isotope curves. Horizontal greybands (1–3) correspond to major ammonite turnover discussed in the text. Ammonite zones and subzones according to Sandoval in Aurell et al. (2002); time scale from Gradstein et al. (2004).
records are characterized by the LO of S. finchii, although the LO ofother taxa (L. hauffi, L. barozii) is not discarded.
7.2.3. RadiolariansThe current knowledge on Aalenian radiolarian is quite fragmen-
tary for both biostratigraphic and palaeobiological aspects. In fact,there is no coherent taxonomic base to establish global radiolarianzonation for the Aalenian, despite recent effort at taxonomic revisionof Early Jurassic radiolarians (Baumgartner et al., 1995; Gorican et al.,2006). The only published paper dealing with the faunal changes thatoccurred at the Early/Middle Jurassic transition is based on very richand well preserved fauna from manganese nodules (Yao, 1997). Thisinformation, though based on discrete levels covering the Toarcian–Aalenian interval, constrains the mechanism and the frameworkgiving rise to faunal change. According to Yao (1997), the majorturnover is located in Middle–Late Aalenian times; a quasi-completereplacement of Early Jurassic fauna is achieved during the Aalenian.Through the Late Aalenian new and important phylogenetic lineagesdeveloped and these were to become the sources for the MiddleJurassic–Early Cretaceous radiolarian genera.
This major turnover is located at the time of an increase ofabundance of radiolarians, as well as a major perturbation in thecarbon cycle (Fig. 10). This drastic faunal change can be interpreted asa major biological response of the siliceous microplankton (as alsooccurs in ammonites) to the opening of new ecological niches due tomajor palaeotectonic alterations in the palaeogeography of thewestern Tethys as consequence of the acceleration of central Atlanticspreading (Bill et al., 2001). The increasing δ13C values in the LateAalenian–Bajocian may be a consequence of a greater oceanicfertilization, as shown by the faunal change that occurred in theradiolarian assemblages (Bartolini et al., 1999). Upwelling played amajor role by enhancing siliceous production (O'Dogherty et al.,2006), in a particular scenario that resulted from the opening of theAlpine Tethys Ocean, which in turn caused a rearrangement, or even anew model of oceanic circulation (Bill et al., 2001).
8. Conclusions
Highly detailed δ13C curves are presented in four Aalenian sectionsin the Subbetic domain. The δ13C curves are well calibrated with high-resolution (subzone) ammonite biostratigraphy and with the maincalcareous nannofossil events.
Correlation between ammonite zonation and nannofossil eventsindicates that the nannofossil zone NJ8 coincides roughly with theentire Aalenian stage. Also, the boundary between radiolarian UAZone2 and UAZone 3 is herein modified and now is placed at theConcavum–Limitatum Subzone boundary.
δ13C values of marine carbonate in Subbetic and in the Umbria–Marche–Sabina sections are consistent with variations of carbon-isotope compositions of sea-water DIC inwestern Tethys. Positive δ13Cexcursions are associated with high abundances of eutrophic calcar-eous nannofossil taxa and strong increases in radiolarian abundancesuggest a high biological productivity and thus enhanced biologicalpump.
During the Upper Aalenian, Subbetic and Apennines marinecarbonate have similar δ13C values suggesting a mixing of DIC sea-water reservoirs in the western Tethys, probably related to a general-ised high productivity, change in oceanic circulation and a bettermixing of sea-water masses.
Stratigraphic differences associated with different paleoceanographicsettings and slight offsets in δ13C values in Subbetic and in Umbria–Marche–Sabina sections point to differences in oxidation rate of organiccarbon in thewesternTethys. Lowerδ13C values in the Subbetic basinwereprobably associated with higher oxidation rate of continental organiccarbon than inUmbrian–Marche–Sabinabasin characterizedbycarbonatestratigraphic sequences far from the influence of terrestrial sediments.
In general, it is found a good correlation between ammoniteextinctions and δ13C decreases, their radiations with δ13C increasing,which also correlate with regressive and transgressive intervalsrespectively
Although the Aalenian time interval is not characterized by majorcarbon perturbations and major biological crisis, a general analysis ofpelagic faunal–floral turnovers reveals a strong interaction betweenvariations in the global carbon cycle rate and the biotic evolution.
Acknowledgments
Wewould like to express our gratitude to Dr. J. Guex, University ofLausanne, for comments on the manuscript and Dr. S. Hesselbo,University of Oxford, for a critical riding, helpful suggestions andEnglish corrections that have improved the manuscript. Also we thankFinn Surlyk and an anonymous reviewer for their comments on thismanuscript. This study forms part of the results obtained in ResearchProjects: CGL2005-02500, financed by the DGI (Dirección General deInvestigación, Spain), by the EMMI and RNM-200 Research Groups,Junta de Andalucía, Spain) and by the Project no. 2000-052555financed by the Swiss National Science Foundation.
References
Aguado, R., O'Dogherty, L., Sandoval, J. (in press). Fertility changes of surface watersduring the Aalenian of the Western Tethys as revealed by calcareous nannofossilsand carbon-cycle perturbations. Marine Micropaleontology. doi:10.1016/j.marmicro.2008.06.001.
Aurell, M., Meléndez, G., Olóriz, F., Bádenas, B., Caracuel, J., García-Ramos, J.C., Goy, A.,Linares, A., Quesada, S., Robles, S., Rodríguez-Tovar, F.J., Rosales, I., Sandoval, J.,Suarez de Centi, C., Tavera, J.M., Valenzuela, M., 2002. Jurassic. In: Gibbons, W.,Moreno, T. (Eds.), The Geology of Spain. The Geological Society, London,pp. 213–253.
Baccelle, L., Bosellini, A., 1965. Diagrammi per la stima visiva della composizionepercentuale nelle rocce sedimentarie. Annali dell'Università di Ferrara Sezione9 (1/3), 59–62.
Bartolini, A., Cecca, F., 1999. 20 My hiatus in the Jurassic of Umbria–Marche Apennines(Italy): carbonate crisis due to eutrophication. Comptes rendus de l'Académie dessciences de Paris, Sciences de la Terre et des Planétes 329, 587–595.
Bartolini, A., Baumgartner, P.O., Mattioli, E., 1995. Middle and Late Jurassic radiolarianbiostratigraphy of the Colle Bertone and Terminilletto sections (Umbria–Marche–Sabina Apennines, Central Italy): an integrated stratigraphical approach. In:Baumgartner, P.O., O'Dogherty, L., Gorican, S., Urquhart, E., Pillevuit, A., De Wever,P. (Eds.), Middle Jurassic to Lower Cretaceous Radiolaria of Tethys: Occur-rences, Systematics, Biochronology. Mémoires de Géologie (Lausanne), vol. 23,pp. 817–831.
Bartolini, A., Baumgartner, P.O., Guex, J., 1999. Middle and Late Jurassic radiolarianpalaeoecology versus carbo-isotope stratigraphy. Palaeogeography, Palaeoclimatol-ogy, Palaeoecology 145, 43–60.
Baumgartner, P.O., O'Dogherty, L., Gorican, S., Urquhart, E., Pillevuit, A., De Wever, P.(Eds.), 1995. Middle Jurassic to Lower Cretaceous Radiolaria of Tethys: occurrences,systematics, biochronology. Mémoires de Géologie (Lausanne) 23, 1–1172.
Bill, M., O'Dogherty, L., Guex, J., Baumgartner, P.O., Masson, H., 2001. Radiolarite ages inAlpine–Mediterranean ophiolites: constraints on the oceanic spreading and theTethys–Atlantic connection. Bulletin of the Geological Society of America 113 (1),129–143.
Bown, P.R., Cooper, M.K.E., Lord, A.R., 1988. A calcareous nannofossil biozonationscheme for the early to mid Mesozoic. Newsletters on Stratigraphy 20, 91–114.
Bown, P.R., Lees, J.A., Young, J.R., 2004. Calcareous nannoplankton evolution anddiversity through time. In: Thierstein, H.R., Young, J.R. (Eds.), Coccolithophores.From Molecular Processes to Global Impact. Springer-Verlag, Berlin, pp. 481–508.
Cobianchi, M., Erba, E., Pirini-Radrizzani, C., 1992. Evolutionary trends of calcareousnannofossil genera Lotharingius and Watznaueria during the Early and MiddleJurassic. Memorie di Scienze Geologiche. Univ. Padova 43, 19–25.
Corfield, R.M., 1995. An introduction of the techniques, limitations and landmarks ofcarbonate oxygen isotope palaeothermometry. In: Boscence, D.W.J., Allison, P.A.(Eds.), Marine Palaeoenvironmental Analysis from Fossils, vol. 83. GeologicalSociety, pp. 27–42 (Special Publication).
Cresta, S., Galácz, A., 1990. Mediterranean basal Bajocian ammonite faunas. examplesfrom Hungary and Italy. Memorie descrittive della Carta Geologica D' Italia 40,165–198.
Cresta, S., Basilici, G., Bussoletti, A., Comas, M.J., Elmi, S., Fernández-López, S., Ferretti, A.,Goy, A., Henriques, M.H., Kalin, O., Jiménez, A., Martínez, G., Sandoval, J., Ureta, S.,Venturi, F., 1995. Relazione sul Workshop “Paleontologia e Biostratigrafia deisedimenti Toarciano–Bajociani in successioni di altomorfostructturale dell'Appennino
136 J. Sandoval et al. / Palaeogeography, Palaeoclimatology, Palaeoecology 267 (2008) 115-137
Umbro–Marchigiano: l'area di Monte Nerone. Correlazioni con altre regioni dellaTetide Occidentale”. (Piobbico, 2–14 agosto 1993). Bolletino Servizio Geologico d'Italia112, 129–139.
Cresta, S., Goy, A., Ureta, S., Arias, C., Barrón, E., Bernad, J., Canales, F., García-Joral, E.,García-Romero, P.R., Gialanella, J.J., Gómez, M.L., González, J.A., Herrero, C.,Martínez, G., Osete, M.L., Perilli, N., Villalaín, J.J., 2001. Definition of the GlobalBoundary Stratotype Section and Point (GSSP) of the Aalenian (Middle Jurassic) andthe Toarcian–Aalenian Boundary. Episodes 24, 166–175.
Dromart, G., Garcia, J.P., Gaumet, F., Picard, S., Roussseau, M., Atrops, F., Lecuyer, C.,Sheppard, M.F., 2003. Perturbation of the carbon cycle at the Middle/Late Jurassictransition: geological and geochemical evidence. American Journal of Science 303,667–707.
Duchamp-Alphonse, S., Gardin, S., Fiet, N., Bartolini, A., Blamart, D., Pagel, M., 2007.Fertilization of the northwestern Tethys (Vocontian Basin, SE France) during theValanginian carbon-isotope perturbation: evidence from calcareous nannofossils andtrace element data. Palaeogeography, Palaeoclimatology, Palaeoecology, 243,132–151.
Erba, E., 2006. The first 150 million years history of calcareous nannoplankton:biosphere–geosphere interactions. Palaeogeography, Palaeoclimatology, Palaeoe-cology 232, 237–250.
Fernández-López, S.R., Henriques, M.H., Linares, A., Sandoval, J., Ureta, M.S., 1999.Aalenian Tmetoceras (Ammonoidea) from Iberia: taxonomy, habitats and evolution.In: Olóriz, F., Rodríguez-Tovar, F.J. (Eds.), Advancing Research on Living and FossilCephalopods. Kluwer Academic/Plenum Publishers, New York, pp. 91–108.
Föllmi, K.B., Weissert, H., Bisping, M., Funk, H., 1994. Phosphogenesis, carbon-isotopestratigraphy, and carbonate-platform evolution along the lower Cretaceous north-ern Tethyan margin. Geological Society of America Bulletin 106, 729–746.
García-Dueñas, V., Linares, A., Mouterde, R., 1967. Datos estratigráficos sobre la seriemesozoica del Río de las Juntas (Montillana, Zona Subbética, Granada). Cuadernosde Geología, Universidad de Granada 1, 193–211.
García-Gómez, R., Jiménez, A.P., Linares, A., Rivas, P., Sandoval, J., 1995. The Toarcian–Aalenian boundary in the Betic Cordillera (southern Spain). Geobios, MémoireSpecial 17, 211–222.
Godet, A., Bodin, S., Föllmi, K.B., Vermeulen, J., Gardin, S., Fiet, N., Adatte, T., Zsolt, B.,Stüben, D., van de Schootbrugge, B., 2006. Evolution of the marine stable carbon-isotope record during the Early Cretaceous: a focus on the late Hauterivian andBarremian in the Tethyan realm. Earth and Planetary Science Letters 242, 254–271.
Gorican, S., Carter, E.S., Dumitrica, P., Whalen, P.A., Hori, R.S., DeWever, P., O'Dogherty, L.,Matsuoka, A., Guex, J., 2006. Catalogue and Systematics of Pliensbachian, Toarcianand Aalenian Radiolarian Genera and Species. ZRC Publishing, Scientific ResearchCentre of the Slovenian Academy of Sciences and Arts, Ljubljana. (Ljubljana).
Gradstein, F.M., Ogg, J.G., Smith, A.G., Agterberg, F.P., Bleeker, W., Cooper, R.A., Davydov,V., Gibbard, P., Hinnov, L., House, M.R., Lourens, L., Luterbacher, H.-P., McArthur, J.,Melchin, M.J., Robb, L.J., Shergold, J., Villeneuve, M., Wardlaw, B.R., Ali, J., Brinkhuis,H., Hilgen, F.J., Hooker, J., Howarth, R.J., Knoll, A.H., Laskar, J., Monechi, S., Powell, J.,Plumb, K.A., Raffi, I., Röhl, U., Sadler, P., Sanfilippo, A., Schmitz, B., Shackleton, N.J.,Shields, G.A., Strauss, H., Van Dam, J., Veizer, J., van Kolfschoten, Th., Wilson, D.,2004. A Geologic Time Scale 2004. Cambridge University Press, Cambridge.
Hallam, A., 1988. A re-evaluation of Jurassic eustasy in the light of new data and therevised Exxon curve. In: Wilgus, C.K., et al. (Ed.), Sea-level Changes: An IntegratedApproach, vol. 42. S.E.P.M, pp. 261–273 (Special Publications).
Hallam, A., 1990. Biotic and abiotic factors in the evolution of Early Mesozoic marinemollusks. In: Ross, R.M., Allmon, W.D. (Eds.), Cause of Evolution. A PaleontologicalPerspective. University Chicago Press, Chicago, pp. 249–260.
Haq, B.U., Hardenbol, J., Vail, P.R., 1988. Mesozoic and Cenozoic chronostratigraphy andcycles of sea-level change. In: Wilgus, C.K., et al. (Ed.), Sea-level Changes: AnIntegrated Approach, vol. 42. S.E.P.M., pp. 71–108 (Special Publication).
Hardenbol, J., Thierry, J., Farley, M.B., Jacquin, T., De Graciansky, P.C., Vail, P.R., 1998.Mesozoic and Cenozoic sequence chronostratigraphic framework of EuropeanBasins. Jurassic sequence chronostratigraphy. In: De Graciansky, P.C., Hardenbol, J.,Jacquin, J., Vail, P.R. (Eds.), Mesozoic and Cenozoic Sequence Stratigraphy ofEuropean Basins. S.E.P.M. (Special Publication 60, chart 6).
Henriques, M.H., Linares, A., Sandoval, J., Ureta, M.S., 1996. The Aalenian in the Ibéria(Betic, Lusitanian and Iberian Basins). GeoResearch Forum 1 (2), 139–150.
Hernández-Molina, F.J., Sandoval, J., Aguado, R., O'Dogherty, L., Comas, M.C., Linares, A.,1991. Olistoliths from the Middle Jurassic in Cretaceous materials of the Fardesformation. Biostratigraphy (Subbetic Zone, Betic Cordillera). Revista de la Sociedadgeológica de España 4, 79–104.
Hesselbo, S.P., Morgans-Bell, H.S., McElwain, J.C., Rees, P.M., Robinson, S.A., Ross, C.E.,2003. Chemostratigraphy of the Jurassic System: applications, limitations andimplications for palaeoceanography. Journal of Geology 111, 259–286.
Hesselbo, S.P., Jenkyns, H.C., Duarte, L.V., Oliveira, L.C.V., 2007. Carbon-isotope record of theEarly Jurassic (Toarcian) Oceanic Anoxic Event from fossil wood andmarine carbonate(Lusitanian Basin, Portugal). Earth and Planetary Science Letters 253, 455–470.
Jenkyns, H.C., 2003. Evidence for rapid climate change in the Mesozoic–Palaeogenegreenhouseworld. Philosophical Translations Royal Society London, A 361,1885–1916.
Jenkyns, H.C., Jones, C.E., Gröcke, D.R., Hesselbo, S.P., Parkinson, D.N., 2002.Chemostratigraphy of the Jurassic System: applications, limitations and implica-tions for palaeoceanography. Journal of Geological Society, London 159, 351–378.
Linares, A., 1979. El Aalenense en la Zona Subbética. Cuadernos de Geología, Universidadde Granada 10, 413–424.
Linares, A., Sandoval, J., 1986. Malladaites nov. gen. et Spinammatoceras (Hammatocer-atidae, Ammonitina) de l'Aalénien de la Zone Subbétique, Sud de l'Espagne.Geobios, 19, 207–224.
Linares, A., Sandoval, J., 1988. Asthenoceras y Fontannesia (Grammoceratinae?,Ammonitina) del tránsito Aalenense–Bajocense de la Zona Subbética (Sur deEspaña). Revista Española de Paleontología 3, 3–11.
Linares, A., Sandoval, J., 1990. The lower boundary of the Bajocian in the “Barranco deAgua Larga” section, (Subbetic Domain, Southern Spain). Memorie descrittive dellaCarta Geologica D' Italia 40, 13–22.
Linares, A., Sandoval, J., 1992. El género Vacekia (Grammoceratinae, Ammonitina) delAaleniense de la Cordillera Bética. Revista Española de Paleontología 91–99 (Extra).
Linares, A., Sandoval, J., 1993. El Aaleniense de la Cordillera Bética (Sur de España).Análisis bioestratigráfico y caracterización paleobiogeográfica. Revista de laSociedad geológica de España 6, 177–206.
Linares, A., Sandoval, J., 1996. The genus Haplopleuroceras (Erycitidae, Ammonitina) inthe Betic Cordillera, Southern Spain. Geobios 29, 287–305.
Lini, A., Weissert, H., Erba, E., 1992. The Valanginian carbon isotope event: a first episodeof greenhouse climate conditions during the Cretaceous (Global Change SpecialIssue) Terra Nova 4, 374–384.
Louis-Schmid, B., Rais, P., Bernasconi, S.M., Pellenard, P., Collin, P.-Y., Weissert, H., 2007.Detailed record of the mid-Oxfordian (Late Jurassic) positive carbon-isotopeexcursion in two hemipelagic sections (France and Switzerland): a plate tectonictrigger? Palaeogeography, Palaeoclimatology, Palaeoecology 459, 459–472.
Mailliot, S., Mattioli, E., Guex, J., Pittet, B., 2006. The early Toarcian anoxia, a synchronousevent in the Western Tethys? An approach by quantitative biochronology (UnitaryAssociations), applied on calcareous nannofossils. Palaeogeography, Palaeoclima-tology, Palaeoecology 240, 562–586.
Mattioli, E., 1995. Bacino Umbro Marchigiano: Produttività primaria, preservazione edeffetti della diagenesi. Ph.D. Thesis. Università di Perugia (Unpublished).
Mattioli, E., Erba, E., 1999. Synthesis of calcareous nannofossil events in Tethyan Lowerand Middle Jurassic successions. Rivista Italiana di Paleontologia e di Stratigrafia105, 349–376.
McCrea, J.M., 1950. On isotopic chemistry of carbonates and paleotemperature scale.Journal of Chemical Physics 18, 849–857.
Morettini, E., Santantonio, M., Bartolini, A., Cecca, F., Baumgartner, P.O., Hunziker, J.C.,2002. Carbon-isotope stratigraphy and carbonate production during the Early–Middle Jurassic: examples from the Umbria–Marche–Sabina Apennines (centralItaly). Palaeogeography, Palaeoclimatology, Palaeoecology 184, 251–273.
O'Dogherty, L., Nieto, L.M., Sandoval, J., 2001. Depósitos de brechas intraformacionales yslumps en el Jurásico Medio del Subbético Medio oriental (Sierra de Ricote,provincia de Murcia). Geotemas 3, 249–253.
O'Dogherty, L., Sandoval, J., Bartolini, A., Bruchez, S., Bill, M., Guex, J., 2006. Carbon-isotope stratigraphy and ammonite faunal turnover for the Middle Jurassic in theSouthern Iberian palaeomargin. Palaeogeography, Palaeoclimatology, Palaeoecol-ogy 239, 311–333.
Pavia, G., Enay, R., 1997. Definition of the Aalenian–Bajocian Stage boundary. Episodes20, 16–22.
Perilli, N., Henriques, M.H., Giannetti, M., 2002. Aalenian calcareous nannofossilchanges and Lotharingius/Watznaueria turnover: evidence from the LusitanianBasin (Portugal). 9th International Nannoplankton Association Conference. Parma,vol. 1, p. 145.
Rocha, R., Henriques, M.H., Soares, A.F., Mouterde, R., Caloo, B., Ruget, C., Fernandez-Lopez, S., 1990. The Cabo Mondego section as a possible Bajocian boundarystratotype. Memorie descrittive della Carta Geologica d' Italia 40, 49–60.
Roth, P.H., 1989. Ocean circulation and calcareous nannoplankton evolution during theJurassic and Cretaceous. Palaeogeography, Palaeoclimatology, Palaeoecology 74,111–126.
Roth, P.H., Thierstein, H.,1972. Calcareous nannoplankton: Leg 14 of theDeep SeaDrillingProject. In: Hayes, D.E., Pimm, A.C., et al. (Eds.), Init. Repts. DSDP 14, 421–485.
Sandoval, J., 1983. Bioestratigrafía y Paleontología (Stephanoceratoidea y Perisphinctoi-dea) del Bajocense y Bathonense de las Cordilleras Béticas. Ph.D. thesis, Servicio dePublicaciones, Univ. Granada, Granada, Spain.
Sandoval, J., Chandler, R.B., 2000. The Sonniniid ammonite Euhoploceras from theMiddle Jurassic of south-west England and southern Spain. Palaeontology 43,495–533.
Sandoval, J., Linares, A., Henriques, M.H., 2000. The Middle Jurassic genus Riccardiceras(Otoitidae, Ammoniotina) in the westernmost Tethys: Betic Cordillera andLusitanian Basin. Revue de Paléobiologie 8, 29–44.
Sandoval, J., Henriques, M.H., Ureta, M.S., Goy, A., Rivas, P., 2001a. The Lias/Doggerboundary in Iberia: Betic and Iberian cordilleras and Lusitanian Basin. Bulletin de laSociété géologique de France 172, 18–21.
Sandoval, J., O'Dogherty, L., Guex, J., 2001b. Evolutionary rates of Jurassic ammonites inrelation to sea-level fluctuations. Palaios 16, 330–363.
Sandoval, J., O'Dogherty, L., Vera, J.A., Guex, J., 2002. Sea level changes and ammonitefaunal turnover during the Lias/Dogger transition in the western Tethys. Bulletin dela Société géologique de France 173, 57–66.
Sandoval, J., Bartolini, A., Bill, M., O'Dogherty, L., 2004. Carbon-isotope stratigraphy andammonite faunal turnover for the Middle Jurassic in the South Iberian Margin. 24thIAS Meeting, Coimbra, Portugal, p. 240.
Sandoval, J., Aguado, R., Bartolini, A., Bill, M., Bruchez, S., O'Dogherty, L., Perilli, N., 2006.Aalenian carbon-isotope stratigraphy: a tool for ammonite, radiolarian andnannofossil zones calibration in Western Tethys. Volumina Jurassica 4, 212–213.
Schmid-Röhl, A., Röhl, H.J., Oschmann, W., Frimmel, A., Schwark, L., 2002. Paleoenvir-onmental reconstruction of Lower Toarcian epicontinental black shales (PosidoniaShale, SW Germany): global versus regional control. Geobios 35, 13–20.
Scholle, P.A., Arthur, M.A., 1980. Carbon-isotope fluctuations in Cretaceous pelagiclimestones; potential stratigraphic and petroleum exploration tool. AAPG Bulletin64 (1), 67–87.
Seyfried, H., 1978. Der subbetische Jura von Murcia (Súdost–Spanien). GeologischesJahrbuch 29, 3–204.
Swart, P.K., Burns, S.J., Leder, J.J., 1991. Fractionation of the stable isotopes of oxygen andcarbon dioxide during the reaction of calcite with phosphoric acid as a function oftemperature and technique. Chemical Geology 86, 89–96.
Tremolada, F., Erba, E., Van de Schootbrugge, B., Mattioli, E., 2006. Calcareousnannofossil changes during the late Callovian–Early Oxfordian cooling phase.Marine Micropaleontology 59, 197–209.
Weissert, H., 1989. C-isotope stratigraphy, a monitor for palaeoenvironmental change: astudy from the early Cretaceous. Surveys in Geophysics 10, 1–61.
Weissert, H., Lini, A., Föllmi, K.B., Kuhn, O., 1998. Correlation of early Cretaceous carbonisotope stratigraphy and platform drowning events: a possible link? Palaeogeo-graphy, Palaeoclimatology, Palaeoecology 137, 189–203.
Yao, A., 1997. Faunal change of Early–Middle Jurassic radiolarians. Proceedings of theFifth Radiolarian Symposium. In: A. Yao (Ed.), News of Osaka Micropaleontologists,special Volume 10, 155–182.