Altered phenotype and function of NKcells infiltrating Human Papillomavirus(HPV)-associated genital warts duringHIV infection☆
Alfred Berea,1, Shahila Tayibb,1, Jean-Mari Krieka,Lindi Massona, Shameem Z. Jaumdally a, Shaun L. Barnabas a,c,William H. Carrd, Bruce Allana, Anna-Lise Williamsona,e,Lynette Denny f, Jo-Ann S. Passmorea,e,⁎
a Institute of Infectious Disease and Molecular Medicine and Division of Medical Virology, University of Cape Town,Cape Town, South Africab Dept. Obstetrics and Gynaecology, Jalan Taming Sari, Taiping Hospital, Perak, Malaysiac Desmond Tutu HIV Foundation, Cape Town, South Africad Department of Biology, Medgar Evers College, The City University of New York, NY, USAe National Health Laboratory Services, Cape Town, South Africaf Dept Obstetrics and Gynaecology, University of Cape Town and Groote Schuur Hospital, Cape Town, South Africa
Received 10 September 2013; accepted with revision 5 December 2013Available online 14 December 2013
☆ This work was funded by the Cancetraining in the USA as part of the ColumInfectious disease research initiativerecipient of the National Research Fouthe HIV Vacccine Trials Network (HVTSouth African Research Chairs Initiative⁎ Corresponding author at: Institute
University of Cape Town, Anzio Road,E-mail address: Jo-ann.Passmore@
Abstract HIV-infected individuals experience more persistent HPV infections and are less likely toresolve genital warts. This study compared phenotype and functions of NK and T cells from genitalwarts and blood from 67 women. We compared in vitro functional responses of NK and T cells bymultiparametric flow cytometry. HIV+ women had significantly lower frequencies of CD4 T cells inwarts (p = 0.001) and blood (p = 0.001). While the distribution of NK cell subsets was similar, HIV+women tended to have lower frequencies of CD56Dim NK cells in both blood (p = 0.0001) and warts(p = 0.006) than HIV−women. Wart NK cells from HIV+ women expressed significantly lower CD107a
and produced IFN-γ. HAART status was not associated with differences in NK cell functionality. We
r Association of South Africa (CANSA) and Poliomyelitis Research Foundation (PRF). AB and JP receivedbia University-Southern African Fogarty AITRP Program. AB is a recipient of the Wellcome Trust Clinical(CIDRI) fellowship from UCT. NIH FIC K01-TW00703-04A1 grant provided support to WHC. JMK is andation (NRF) of SA Scarce Skills Post-Graduate Scholarship. SLB is a recipient of a SHAPe fellowship ofN) and the Desmond Tutu HIV Foundation. This work is partially based on research supported by theof the Department of Science and Technology and National Research Foundation (NRF) of South Africa.
of Infectious Disease and Molecular Medicine, Division of Medical Virology, Faculty of Health Sciences,Observatory 7925, Cape Town, South Africa. Fax: +27 21 4066681.uct.ac.za (J.-A.S. Passmore).ibuted equally to this work.
Human Papillomaviruses (HPVs) are a highly diverse groupof DNA viruses replicating in differentiating epithelialcells. Some HPV sub-types preferentially infect the genitalmucosa. Many of these viruses cause benign genital warts andare referred to as low-risk types (including HPV-11 and -6)[1,2]. Whereas others, such as HPV-16 and HPV-18, have thepotential to cause cervical cancer and are referred to ashigh-risk HPV types [3–5]. In the absence of HIV, infection byeither high or low risk HPV sub-types commonly regresseswithout treatment. Typically, 23% of warts are cleared bytwo months following infection and 66% are cleared by twoyears [6]. Host defences that are responsible for resolvingsuch HPV infections rely on an intact immune responseinvolving both T cells and natural killer (NK) cells [7,8]. BecauseHPV-associated genital warts are typically self-limiting orself-resolving, their persistence is often indicative of immunedysfunction.
Impaired NK cell function and skewed NK cell subset dis-tributions have been reported in HIV+ individuals. The ability ofthese NK cells to control secondary viral infections differsdepending on which viral sub-type initiates the secondaryinfection (9). Chronic HPV infections, which are often charac-terized by high HPV viral loads, may be facilitated by HIVco-infection that reduces the likelihood of spontaneous HPVclearance [10–12]. Also, it is likely that shared transmissionroutes via the genital mucosa increase the probability ofindividuals acquiring HPV and HIV simultaneously. Co-infectionwith HIV and HPV can present major challenges to an infectedindividual's immune system as both viruses interferewith innateand adaptive immunity.
NK cells can be subdivided into distinct populationsbased on the expression of CD16 and CD56 cell-surfaceantigens, with the two main NK cell subsets being definedas CD56Dim CD16+ and CD56Bright CD16−. During chronic HIVdisease, there is an expansion of a functionally-impairedCD56− CD16+ NK cell subset [13]. CD16, the FcRIII receptor, isinvolved in the antibody-dependent cellular cytotoxic (ADCC)function of NK cells [14]. The CD56Dim NK cells representthe major cytotoxic subset responsible for the direct lysis ofvirally-infected cells (8, 14). CD107a expression is used as asurrogatemarker of cytotoxic degranulation that is upregulatedon the surface of NK cells when stimulated [15]. CD56Bright NKcells, in contrast, are reported to have low cytotoxic capacitybut produce cytokines abundantly [14,16]. The CD56−CD16+ NK cell subset, which increases in frequency duringHIV disease progression, exhibits low cytotoxicity and poorcytokine secretion potential [13].
While many studies have clearly described the phenotypicand functional characteristics of NK cells circulating inperipheral blood, fewer have compared these to NK cellsfound at effector sites and in tissues. Because of their rolein placental formation and fertility, several important
studies on uterine NK cells in the upper reproductive tractsuggest that their phenotypes differ significantly fromthose of systemic NK cells, and likely depend on theirtissue location and the cytokine environment in which theyexist [17]. While uterine NK cells express CD56, fewexpress CD16. In comparison, NK cells derived from theupper female genital tract tend to be CD56Bright whilethose in the lower genital tract tend to be CD56Dim [17].
Better understanding of the specific role(s) of, anddefects in, NK cell function in HPV-associated genital wartregression during HIV infection is significantly hindered by alack of data on NK cell function at sites in the genitalmucosae where HPV infection and replication primarily takeplace. The aim of this study was to compare the phenotypes(CD16 and CD56 expression) and functions (CD107a degranu-lation and IFN-γ production) of NK cells isolated from pairedHPV-associated genital warts and blood obtained from bothHIV+ and HIV− women. Since CD4 T cells that produce IFN-γare the best-characterized correlate of wart regression [18]depletion and loss of function of CD4 T cells during HIV diseaseprogression may influence HPV pathogenesis. Thus, we alsocompared IFN-γ production of in vitro stimulated CD4 T cellsderived from warts and blood. We hypothesised that HIV+women would have systemic NK and T cell defects that wouldbe evident by HPV-associated genital warts containing wart-infiltrating NK and T cells with reduced function compared tothose found in HIV− women.
2. Materials and methods
2.1. Study participants and sample collection
Sixty-seven women with genital warts were recruited from theColposcopy Outpatients clinic at Groote Schuur Hospital. TheHuman Research Ethics Committee of the University of CapeTown approved all aspects of the study andwritten consent wasobtained from each of the women participating in this study.Genital warts were removed from each woman under speculumexamination in surgical theatre and immediately placed into a15 ml conical tube containing 3 ml of R10 (10% FBS in RPMI-1640medium (GIBCO®) containing 50 ug/ml Penicillin, 50 mg/mlStreptomycin, 0.8 mg/ml Fungin and 50 mg/ml L-glutamine)for transport to the laboratory. Tubes containing genitalwarts were transported to the laboratory at UCT within 1–2 hof collection. A portion of each of the warts was placedimmediately into a Digene Transport tube for isolation of DNAfor HPV genotyping. In addition, 40 ml ACD anti-coagulatedwhole blood was collected from each woman by venepuncturefor the isolation of peripheral bloodmononuclear cells (PBMCs).
2.2. Disaggregation of genital warts
Genital warts were transferred onto a stainless steel 35micron cell strainer together with the 3 ml of R10 transport
212 A. Bere et al.
medium. The tissue was cut into 2–3 mm pieces with asterile, disposable scalpel and the wart was disaggregatedmanually by agitating the tissue chunks through the stainlesssteel strainer using a plastic plunger to separate the lympho-cytes from the tissue matrix. The strainer was flushed withanother 3 ml of R10 medium to remove all the cells and thehomogenized suspension was pooled into a 15 ml conical tubeand centrifuged at 1200 rpm (280 g) for 10 min. Cells werere-suspended in 6 ml of R10 medium.
2.3. PBMC isolation
PBMCs were isolated by density gradient centrifugation usingFicoll-Histopaque (Sigma–Aldrich, Egham, Runnymede, UK) andLeucoSep® centrifuge tubes (Greiner Bio-one, Frickenhausen,Germany). Plasma was stored at −80 °C for measurement ofHIV viral loads.
2.4. Flow cytometry
Genital wart- and blood-derived NK and T cell phenotypesand functional responses were measured using the follow-ing panel of fluorescently-conjugated monoclonal anti-bodies: CD3-APC-H7, CD4-PerCp-Cy5.5, CD16-PE, CD56-PECy7, CD107-FITC and IFN-γ-Alexa fluor 700 (all BD Biosciences,San Diego, CA, USA), CD8-quantum dot 605 (Invitrogen,Carlsbad, CA, USA), CD14-PacBlue (BD Biosciences San Diego,CA, USA), and CD19-PacBlue (Invitrogen, Carlsbad, CA, USA).All antibodies were pre-titered to optimal concentrations.Violet viability reactive dye (“Vivid”; Invitrogen) was includedto differentiate live from dead cells. To investigate thefunction of genital wart and blood NK cells, these werestimulated with (a) the tumour cell line K562 (1 × 105 K562cells in 100 μl); (b) PMA (0.1 μg/ml)/ionomycin (0.1 μg/ml)[positive control]; and (c) media alone (negative control). Toeach stimulation, 10 μl of R10 stimulation mix (4.5 ml R10,500 μl DNase I, anti-CD49d (1 μg/ml), anti-CD28 (1 μg/ml),BFA (0.5 μg/ml) and monensin (10 μg/ml) (Sigma Aldrich)was added. In addition, anti-CD107a-FITC antibody (5 μl) wasincluded in each stimulation tube. For PMA/ionomycinstimulation, cells were incubated for 3 h. For the remainingstimulations with K562 or no antigen (negative control), cellswere cultured for 6 h at 37 °C, 5% CO2 (Thermo ElectronCorporation). Cells were subsequently washedwith FACSwashbuffer (1% FBS and 0.01% NaN3, GIBCO® PBS, Invitogen™,Carlsbad, CA, USA) and centrifuged at 838 g (2300 rpm) for3 min at 4 °C. Cells were re-suspended in 50 μl PBS containingVivid dye (Invitrogen) and incubated for 20 min at roomtemperature. For cell-surface staining, a mixture of antibod-ies, consisting of anti-CD19-Pacific Blue, anti-CD14-PacificBlue, anti-CD8-Qdot 605, anti-CD4-PerCP-Cy5.5, CD16-PE andCD56-PE Cy7 in PBS, was prepared and cells were stained for20 min at room temperature. Cells were fixed and perme-abilized at room temperature for 20 min using Cytofix/Cytoperm buffer (BD Biosciences) and stained intracellularlywith anti-CD3-APC-H7 and anti-IFN-γ-Alexa fluor 700. Allstaining was done in PBS at a final volume of 50 μl. Cellswerewashed, and finally resuspended in 1% paraformaldehyde(Cell fix BD Biosciences) and stored at 4 °C until acquisition(within 24 h). Samples were acquired on a BD Fortessa (BDBiosciences) with FACSDiva software version 6.0 (BD
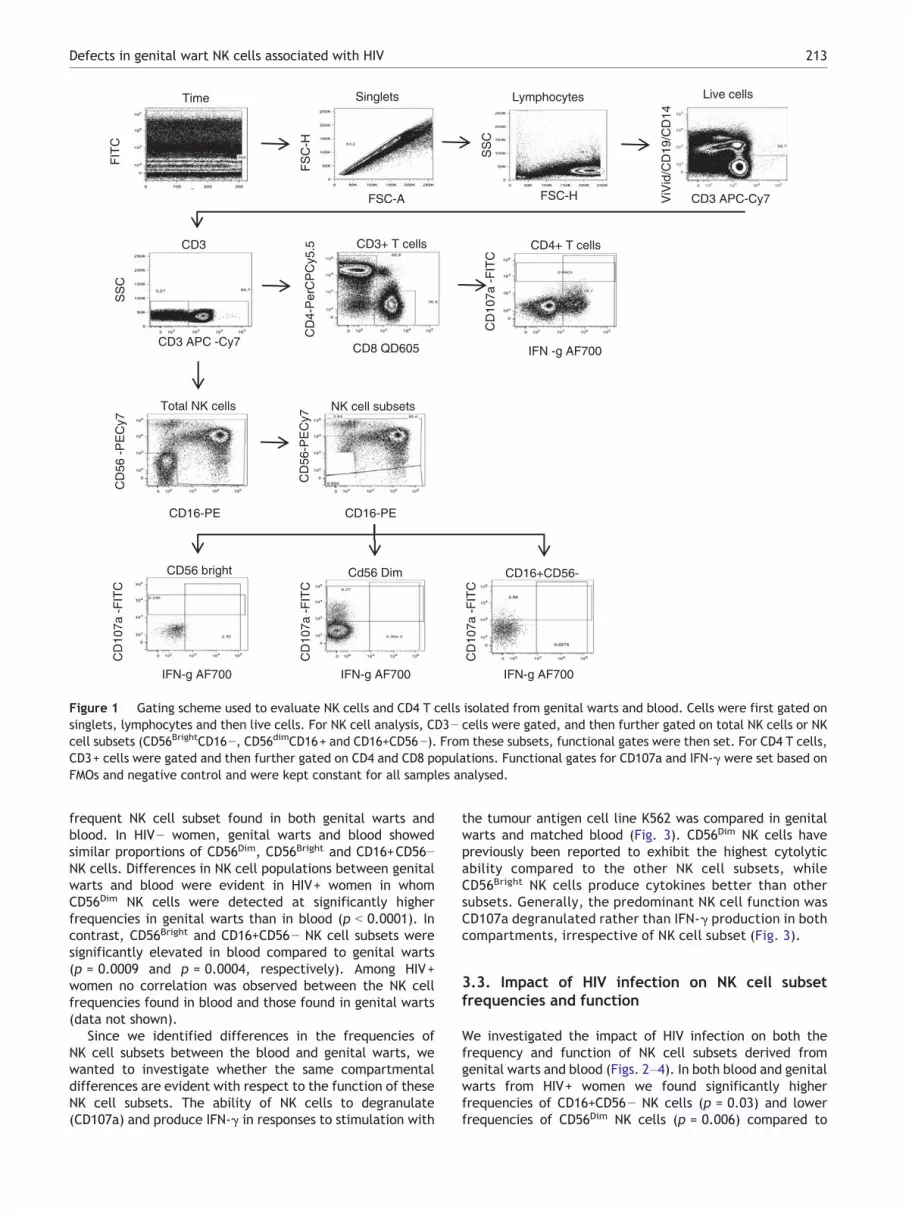
Biosciences), with approximately 0.5 × 106 events beingacquired per sample. Data analysis was performed usingFlowJo software v8.5.3 (Tree Star, Inc; Ashland, Oregon,OR, USA). Dead cells (ViVid+), monocytes (CD14+), and B cells(CD19+) were excluded from the analysis. Fluorescence minusone (FMO) controls were used to set gates. For the gatingstrategy, doublets were excluded based on forward scatter(FSC) height and FSC area (Fig. 1). A broad PBMC gate wasthen defined based on FSC height and side light scatter.Monocytes and B cells were excluded based on CD14 andCD19 gating, respectively. NK cells were identified withinthe CD3− gate based on the expression of CD16 and CD56.NK cells were then subdivided into CD56Bright, CD56Dim, andCD16+CD56− populations and subsequently analyzed forthe expression of IFN-γ and CD107a as summarised onFig. 1.
2.5. Statistical analysis
Statistical analyses were performed using GraphPad Prism5® (GraphPad Software, San Diego California USA). TheMann–Whitney U test was applied for independent samplecomparisons, the Wilcoxon Ranks Test was used for matchednon-parametric comparisons and Spearman Ranks correlationwas applied for assessing associations. P-values of≤0.05 wereconsidered significant.
3. Results
3.1. Clinical characteristics of women
Sixty seven women with genital warts were included inthis study to investigate the impact of HIV-infection on thephenotype and function of NK and T cells resident in HPV-associated genital warts and how these compare with cellscirculating in blood. Of the 67 women included, 51/67 (76.1%)were HIV+ while 16/67 (23.9%) were HIV− (Table 1). Of the 51HIV+ women, 37/51 (72.5%) were on HAART while 14/51(27.5%) were not. HIV+HAART+ women tended to be older thanboth HIV+HAART− and HIV− women (p b 0.01), had lower HIVviral loads than HIV+HAART− women (p b 0.001), were morelikely to have normal Pap smear results (p = 0.04), and weremore likely to have milder and smaller warts (p = 0.006)compared to HIV+HAART−women. HIV+HAART+women did notdiffer significantly from HIV− women in terms of their Papsmear results and the sizes of their genital warts.
Despite HIV+HAART+ women having a higher proportionof mild warts than HIV+HAART− women, CD4 counts amongHIV+ women, irrespective of HAART status, did not differ.Irrespective of HIV or HAART status, HPV-11 and then HPV-6,were found to be the most common HPV types associatedwith genital warts in this study, with HPV-16 and -45 beingthe next most common types detected.
3.2. Comparison of NK cell phenotype and functionin genital warts and blood
The frequencies of different NK cell subsets in genital wartswere compared to those found in blood (Fig. 2). Irrespectiveof HIV status, CD56Dim CD16+ NK cells were the most
FSC-A
Time
FSC-H CD3 APC-Cy7
CD3 APC -Cy7
Total NK cells NK cell subsets
CD3+ T cells CD4+ T cells
CD56 bright Cd56 Dim CD16+CD56-
Singlets Lymphocytes Live cells
CD3
FIT
C
FS
C-H
SS
C
ViV
id/C
D19
/CD
14
SS
C
CD
4-P
erC
PC
y5.5
CD8 QD605
CD16-PE
IFN -g AF700
CD
107a
-F
ITC
CD
107a
-F
ITC
CD
107a
-F
ITC
CD
107a
-F
ITC
IFN-g AF700 IFN-g AF700 IFN-g AF700
CD
56 -
PE
Cy7
CD
56-P
EC
y7
CD16-PE
Figure 1 Gating scheme used to evaluate NK cells and CD4 T cells isolated from genital warts and blood. Cells were first gated onsinglets, lymphocytes and then live cells. For NK cell analysis, CD3− cells were gated, and then further gated on total NK cells or NKcell subsets (CD56BrightCD16−, CD56dimCD16+ and CD16+CD56−). From these subsets, functional gates were then set. For CD4 T cells,CD3+ cells were gated and then further gated on CD4 and CD8 populations. Functional gates for CD107a and IFN-γ were set based onFMOs and negative control and were kept constant for all samples analysed.
213Defects in genital wart NK cells associated with HIV
frequent NK cell subset found in both genital warts andblood. In HIV− women, genital warts and blood showedsimilar proportions of CD56Dim, CD56Bright and CD16+CD56−NK cells. Differences in NK cell populations between genitalwarts and blood were evident in HIV+ women in whomCD56Dim NK cells were detected at significantly higherfrequencies in genital warts than in blood (p b 0.0001). Incontrast, CD56Bright and CD16+CD56− NK cell subsets weresignificantly elevated in blood compared to genital warts(p = 0.0009 and p = 0.0004, respectively). Among HIV+women no correlation was observed between the NK cellfrequencies found in blood and those found in genital warts(data not shown).
Since we identified differences in the frequencies ofNK cell subsets between the blood and genital warts, wewanted to investigate whether the same compartmentaldifferences are evident with respect to the function of theseNK cell subsets. The ability of NK cells to degranulate(CD107a) and produce IFN-γ in responses to stimulation with
the tumour antigen cell line K562 was compared in genitalwarts and matched blood (Fig. 3). CD56Dim NK cells havepreviously been reported to exhibit the highest cytolyticability compared to the other NK cell subsets, whileCD56Bright NK cells produce cytokines better than othersubsets. Generally, the predominant NK cell function wasCD107a degranulated rather than IFN-γ production in bothcompartments, irrespective of NK cell subset (Fig. 3).
3.3. Impact of HIV infection on NK cell subsetfrequencies and function
We investigated the impact of HIV infection on both thefrequency and function of NK cell subsets derived fromgenital warts and blood (Figs. 2–4). In both blood and genitalwarts from HIV+ women we found significantly higherfrequencies of CD16+CD56− NK cells (p = 0.03) and lowerfrequencies of CD56Dim NK cells (p = 0.006) compared to
Table 1 Clinical characteristics of women involved in this study.
Characteristic HIV+HAART+ HIV+HAART− HIV−
N 37 14 16Age [years; median (IQR)] 28 (24–36) 23 (21–25) 24 (20–35)CD4 count [cell/μl; median (IQR)] 328 (218–481) 299 (299–415) N/APlasma viral load [RNA copies/ml; median (IQR)] 0 (0–360) 33356 (11000–75000) a N/APap smear resultsNormal 9/30 a (30.0%) 8/14 (57.1%) 6/9 a (66.7%)LSIL 19/30 a (63.3%) 6/14 (42.9%) 2/9 a (22.2%)HSIL 2/30 a (6.7%) 0/14 (0.0%) 1/9 a (11.1%)Size of genital wartsMild 25/31 a (80.6%) 5/14 (35.7%) 12/16 (75.0%)Moderate 5/31 a (16.1%) 8/14 (57.1%) ⁎ 3/16 (18.8%)Severe 1/31 a (3.2%) 1/14 (7.1%) 1/16 (6.3%)Weight of genital wart (g) 0.10 (0.06–0.3) 0.35 (0.13–0.36) ⁎ 0.16 (0.08–0.22)a Pap smear cytology and wart size data were only available for a subset of the women in the study.⁎ p b 0.01.
214 A. Bere et al.
HIV− women (Fig. 2). Interestingly, the distribution of genitalwart NK cell subsets did not differ significantly between HIV+women who were HAART therapy naïve compared to thoseusing HAART (data not shown).
We investigated whether blood- and genital wart derived NKcells from HIV+ women had detectable functional defectsrelative to HIV− women. Based on CD107a expression (as asurrogate for degranulation) and intracellular IFN-γ production,NK cells from HIV−womenwere generally more functional thanthose from HIV+ subjects (Fig. 3). Warts from HIV+ women(naïve to HAART) contained significantly lower frequencies ofCD56Bright and CD56Dim NK cells that expressed CD107a (p =0.001 for CD56Bright and p = 0.003 for CD56Dim) and producedIFN-γ (p = 0.001 for CD56Bright and p = 0.0002 for CD56Dim) thanwarts from HIV− women. HAART did not improve thesefunctional defects in wart NK cells because HIV+ women onHAART had similar frequencies of NK cells expressing CD107 orproducing IFN-γ as HIV+ women naïve to HAART. Although HIV+women on HAART tended to have smaller warts than theirHAART naïve counterparts (Table 1), the NK cells of HAART+women had similar functionality (based on CD107a expressionand IFN-γ production) to those of HIV+HAART− negative women(Figs. 3A-B). Functional defects (CD107a expression or IFN-production) were not similarly evident in blood derived NK cellsfrom these same HIV+ women, who had higher CD56Bright andCD56Dim NK responses than HIV− women (Figs. 3C–D).
We stratified these functional responses based on theclinical characteristics of genital warts (mild, moderateand severe; Table 2). Among HIV− women, moderatewarts from HIV− women had 9-fold higher frequencies ofCD107a + CD56Dim (p = 0.005) and 7-fold higher frequencies ofCD56Bright NK cells (p = 0.008) than mild warts. While CD56Dim
CD107a + NK cells were similarly 12-fold elevated in moderatecompared to mild warts of HIV+ women, CD56Bright NK cellswere absent from both moderate and mild found in thesewomen. IFN-g + NK cell subsets were present at similarly lowfrequencies in the moderate and mild warts of both HIV− andHIV+ women, and were not obviously associated with wartseverity (Table 2).
The ability of blood NK cells to produce IFN-γ was lowerthan their ability to express CD107a. Despite this, during
HIV-infection, IFN-γ production by all subsets of NK cells inblood was significantly higher than those detected in genitalwarts, with genital wart IFN-γ production only being evidentat low frequencies in a few HIV+ women (Fig. 4).
3.4. Defects in CD4 T cell frequencies rather thanfunction during HIV disease
CD4 T cell frequencies (of CD3+ cells) were significantlylower in the blood of HIV+ women than they were in theblood of HIV− women (Fig. 5A; p = 0.002 for HAART− andp = 0.007 for HAART+). There were lower frequencies of CD4T cells in genital warts than in blood. Similarly, CD4 T cellfrequencies were also significantly lower in the genital wartsof HIV+ women than HIV− women, regardless of treatmentstatus (p b 0.01). HAART+ women had significantly higherCD4 T cell frequencies in warts compared to HAART− women(p = 0.0001), although these still present at lower frequen-cies than those measured in HIV− women. While CD4 T cellsaccounted for ~17% of all CD3 T cells isolated from the genitalwarts of the HIV−women, this helper T cell subset was almostcompletely absent from the genital warts of HIV+ women, inwhich CD8 T cells made up the major population. Despitereduced numbers, their ability to produce IFN-γ in response toPMA/ionomycin stimulation was similar (Fig. 5B). This indi-cates that reduced numbers of CD4 T cells rather than thesecells being functionally defective in the genital warts andblood of HIV+ women may underlie their observed inability tospontaneously clear HPV-associated genital warts.
4. Discussion
HIV+ women generally have more persistent HPV-infectionsthan those occurring in HIV− women. This increasedpersistence places HPV-infected women at greater risk ofdeveloping cervical disease [10–12]. Although HPV typesthat cause genital warts are typically benign, failure tospontaneously clear such warts during HIV infection hasimportant socio-economic consequences. In this study, wehave investigated the impact of HIV-infection on both the
A) Blood B) Genital warts
CD56Bright CD56Dim CD16+CD56-
0
20
40
60
80
100
p=0.0001
p=0.0007%
NK
cel
l exp
ress
ion
0
20
40
60
80
100
HIV+HIV-
CD56Bright CD56Dim CD16+CD56-
p=0.006
p=0.03
0
20
40
60
80
100
% o
f NK
cel
ls
0
20
40
60
80
100
P=0.001
P<0.0001
P<0.0001Genital wartsBlood
C) HIV- D) HIV+
Figure 2 Comparison of NK cell phenotypes in (A) blood versus (B) genital warts. HIV− women (grey boxes) and HIV+ women (whiteboxes). Each box and whisker plot shows the median (central line), IQR (outer lines of box) and 10–90% range (error bars) of theindividuals. Wilcoxon Rank Test was used to compare groups and p b 0.05 was considered significant.
215Defects in genital wart NK cells associated with HIV
phenotype and function of NK and T cell subsets isolatedfrom HPV-associated genital warts and blood of women.Genital warts were larger in HIV-infected than uninfectedwomen, containing similar frequencies of CD56Dim, CD56Bright
and CD16+CD56− NK cells despite containing significantlylower frequencies of CD4 T cells. Despite being present atsimilar frequencies in HIV-infected and HIV negative warts,significantly lower frequencies of CD56Dim and CD56Bright NKcell subsets from HIV-infected women expressed CD107a andproduced IFN-γ compared to HIV negative women, suggestingimportant functional defects in these anti-viral cells. WhileCD4 T cells were present at significantly lower frequencies inboth blood and genital warts in HIV infected compared touninfected women, the ability of these cells to produce IFN-γin response to PMA/ionomycin stimulation was similar.
The best known correlate of HPV-associated wart regres-sion, described in the canine oral papillomavirus model, isCD4 T cell infiltration into warts [7,19]. Nicholls et al. (7)reported an influx of lymphocytes just prior to the regressionof lesions, and that this influx reached a peak during theresolution of lesions. Nicholls et al., [7] noted that the denseinfiltration of T cells happened at the basal region of theepidermis, and that the most abundant population was thatof intraepithelial CD4 T cells. They hypothesized that theinfiltration of CD4 T cells would facilitate lesion clearancethrough cytokine-induced activation of macrophages thatsubsequently kill HPV-infected keratinocytes. However,
Nicholls et al. [7] did not address how HPV-associatedgenital warts are resolved during CD4 T cell lymphopenia,such as that which occurs during chronic HIV disease.
To our knowledge, no comparable studies identifyingcorrelates of wart clearance or persistence have beencarried out in humans. Whereas our cross-sectional studycannot infer causality, we discovered differences in func-tional responses of NK cells derived from genital warts ofHIV+ women compared to HIV− women. We also found lowerfrequencies of CD4 T cells among warts of HIV+ womencompared to those of HIV− women. Taken together, theseresults imply that it is entirely plausible that these defectsmay underlie the failure of HIV+ women to clear warts.Nicolls et al. [7] suggested that CD4 T cells trafficked intocanine oral warts and produced IFN-γ to resolve wartlesions. We found that among HIV+ women lower frequen-cies of CD4 T cells in genital warts, rather than intrinsicdecreased functionality, were associated with larger warts.Nicolls et al. [7] did not measure NK cell frequencies norfunctionality, thus they could not comment on the role of NKcells in HPV pathogenesis during HIV co-infection. Ourfinding that HIV+ women have multiple NK cell subset andfunctional deficiencies suggests that multiple factors mayunderlie wart persistence.
It was interesting to note that many of the HIV− womendeveloping warts in this study had CD4+ T-cells percentages(of CD3+ cells) in blood and genital warts of ~50% (suggesting a
Figure 3 Comparison of distinct NK cell subset functions in genital warts (A–B) and blood (C–D) genital warts. PBMC anddisaggregated wart-infiltrating cells were stimulated by the tumour cell line K562 to measure NK cell function (CD107a degranulationor IFN-γ production). HIV+ women (HAART−: white boxes; HAART+: black boxes) and HIV− women (grey boxes). Each box and whiskerplot shows the median (central line), IQR (outer lines of box) and 10–90% range (error bars) of the individuals. Wilcoxon Rank Test wasused to compare groups and p b 0.05 was considered significant.
216 A. Bere et al.
CD4:CD8 ratio of 1:1) which is lower than CD4+ cell frequencieswhich report typical healthy HIV− CD4:CD8 of N2:1 [20,21].This finding possibly suggests that HIV− women developinggenital warts in our setting had lower CD4 counts than
0
10
20
30 Genital wartBlood
% N
K c
ell r
espo
nse
CD107a+ IFN- + CD107a+ IFN- + CD107a+ IFN- +
p=0.01
p=0.01
p=0.01
CD56Bright CD56Dim CD16+CD56-
Figure 4 Comparison of genital wart (white boxes) andmatched blood responses (grey boxes) by distinct NK cellsubsets during HIV infection. Each box and whisker plot showsthe median (central line), IQR (outer lines of box) and 10–90%range (error bars) of the individuals. Wilcoxon Rank Test wasused to compare NK cells in warts to those in blood and p b 0.05was considered significant.
previously published studies that may account for theirinability to control wart development. It would be interest-ing in future studies to include genital tissue from healthywomen without warts for comparison, to further investigatethe impact of non-HIV associated CD4 deficiency on suscepti-bility to HPV wart progression.
Impaired NK cell function and the expansion of dysfunc-tional NK cell subsets have been noted in HIV+ individuals,with the extent of impairment being associated with plasmaHIV loads [9,13,14]. While several studies have reported asignificant reduction in the frequencies of CD56+ NK cellsduring HIV infections [22–24], the emergence of CD56−CD16+ NK cells (particularly in individuals with high HIVloads in blood) has been shown to account for this apparentreduction in CD56+ cells [13,25]. We also noted both thatCD56− CD16+ NK cells were significantly higher in frequencyin HIV+ than HIV− women (particularly in blood, but also ingenital warts), and we found that enrichment of this subsetwas associated with reduced frequencies of CD56Dim NKcells. Interestingly, as others have shown, we did not findthat initiation of HAART normalized CD16+CD56− NK cellsubset frequencies in either blood or warts to those seen inHIV−women [26]. In our cohort, the insignificant differences inCD4 count between HAART-experienced and HAART-naïveHIV-infected women suggest either full immuno-competencewas not restored by HAART or that the duration of HAARTwas not sufficient to enable restoration of full immuno-competence.
Table 2 NK subset function according to wart size.
NK phenotype and function Frequency of NK function according to wart size[Median % (IQR)]
*p = 0.008; **p = 0.005; ***Wart size data was available for 45 HIV+ women (31/37 HIV+HAART+ and 14/14 HIV+HAART−).
217Defects in genital wart NK cells associated with HIV
Our observation that higher proportions of functionallyimpaired NK cells are present in the genital warts of HIV+women has important biological implications. PreviouslyCD56− CD16+ NK cells have been shown to lack most NK cellfunctions [27]. Although we did observe that CD56− CD16+NK cells from HIV+ women had significantly reduced IFN-γproducing capacity compared to HIV− women, they retainedthe ability to degranulate (i.e. they were CD107a+). We alsofound that wart-derived CD56dimCD16+ NK cells from HIV+women showed impaired degranulation compared to thosefrom HIV− women.
Following HAART initiation, most HIV+ individuals showa gradual rise in CD4 T cell counts, although many do notattain pre-infection CD4 counts and remain immune-suppressed[28,29]. Approximately 7–20% of HIV+ individuals who haveinitiated HAART have been reported to have suboptimal
0
20
40
60
80
HIV- HIV+HAART-
HIV+HAART+
p=0.009
p=0,002
p=0.007
p=0.0001
p=0.001
% C
D4
T c
ells
(of
CD
3)
% C
D4
T c
ells
IFN
-
A B
1
2
Figure 5 Comparison of CD4 T cell frequencies and the abilities oblood (grey boxes) taken from HIV− and HIV+ women (HAART− or Hline), IQR (outer lines of box) and 10–90% range (error bars) of the ip b 0.05 was considered significant.
immune reconstitution despite almost complete viral suppres-sion [30–32] and this variability is dependent on the duration ofHAART [33]. Poorer recovery of CD4+ T cell counts is associatedwith delayed initiation of HAART (especially in those with CD4+T cell counts b200 cells/ml).
The frequency distribution of NK cell subsets found ateffector sites such as the genital mucosa is likely to differfrom those found in blood because of both their tissuelocation and the cytokine environment in which they exist[17]. CD56Dim NK cells are the predominant NK cell subset inthe spleen and peripheral blood, whereas CD56Bright NK cellsare predominant in lymph nodes and tonsils [34,35]. Previousstudies have shown that NK cells are relatively abundant inthe uterus during pregnancy, expressing CD56 but not CD16[36,37]. Sentman et al. [17] have further differentiated NKcell phenotypes based on their position within the female
HIV- HIV+HAART-
HIV+HAART+
0
1
2
3
4
5
0
0Genital wart
Blood
f these cells to produce IFN-γ in genital warts (white boxes) andAART+). Each box and whisker plot shows the median (centralndividuals. Wilcoxon Rank Test was used to compare groups and
218 A. Bere et al.
genital tract: upper genital tract NK cells were described asbeing CD56Bright while those in the lower genital tract beingCD56Dim. We found that NK cells infiltrating genital wartswere predominantly CD56Dim and that this reflected thedominant subset in blood.
CD56Dim NK cells, which predominated in our study in bothgenital warts and blood, irrespective of HIV status, havepreviously been shown primarily to be cytotoxic killers ratherthan cytokine producers [14]. Consistent with this, we foundboth that NK cell degranulation (as evidenced by CD107aexpression) was the predominant effector mechanism by thisNK cell subset, and that frequencies of CD107a + NK cells weresimilar in genital warts to those observed in blood. We showedthat CD107a expression by CD56Dim NK cells was positivelyassociated with the severity of warts in both HIV+ and HIV−women, with NK cells from moderate warts degranulatingmore than those from smaller warts. Unlike CD56Dim NK cellsor CD56Bright cells from HIV− women, we found that wartCD56Bright cells from HIV+ women lacked CD107a expressionand were therefore apparently lacking in their ability todegranulate in response to stimulation with K562 cells. This isinteresting because CD56Bright NK cells are reported primarilyto produce cytokines rather than exhibit cytolytic activity[14]. Among HIV+ women in general, IFN-γ production wassignificantly lower in all wart-derived NK cell subsets relativeto that observed by blood-derived NK cells. However, followingin vitro stimulation, frequencies of IFN-γ-expressing NK cellsderived from blood of HIV+ women were similar to or greaterthan those derived fromwarts of HIV−women. Others have alsofound that HIV infection reduces the likelihood of spontaneousHPV clearance and wart regression [10,11]. Thus, based on ourdatawe conclude that this enables the establishment of chronicHPV infections through multiple dysfunctions in cytolyticactivity and cytokine production by several NK cell subsets atthe genital mucosa. This finding provides new insights intopotential causes of HPV persistence.
Although the initiation of HAART in HIV+ women has beenreported to offer some reconstitution of functional deficitsamong dysfunctional NK subsets (CD56− CD16+) [14,26],we did not observe this in our study. We observed noquantitative differences in NK cell function among womenon HAART compared to those who were HAART−. Althoughwe do not know the duration of HAART in this cohort or theCD4 counts at the time of HAART initiation, the fact that CD4counts in women on HAART was not significantly higher thanHAART naïve women suggests that CD4 reconstitution inHAART+ women was not complete. It is possible thatprolonged HAART or earlier HAART initiation would resultin reversal of some of the NK cell defects we report in thisstudy. The finding that IFN-γ responses by CD4 T cells weresimilar while absolute frequencies of CD4 T cells weresignificantly lower in warts from HIV+ compared to HIV−women suggests that wart size or regression was notdependent on CD4+ IFN-γ producing cells. It is clear thatwomen with persistent HPV infections who are unable tocontain growth of potentially disfiguring genital warts are atthe highest risk of developing cervical disease progressing tocancer. Given that HIV+ women frequently fail to containthe proliferation of HPV-associated warts, it is imperativethat we understand the underlying reasons for persistentHPV infections in these woman so that we can reduce thelikelihood of them developing cervical cancer. The multiple
defects we have identified within the genital wart associat-ed NK cell subsets of HIV+ women is a small but importantstep towards the development of strategies combattingpersistent secondary high risk HPV infections in immune-compromised women.
Conflict of interest statement
The author(s) declare that there no conflicts of interest.
Acknowledgments
This study was supported by the Cancer Association of SouthAfrica (CANSA) and Poliomyelitis Research Foundation (PRF).
References
[1] A.N. Burchell, P. Tellier, J. Hanley, F. Coutlée, E.L. Franco,Influence of partner's infection status on prevalent humanpapillomavirus among persons with a new sex partner, Sex.Transm. Dis. 37 (2010) 34–40.
[2] N. Muñoz, F.X. Bosch, S. de Sanjosé, R. Herrero, X.Castellsagué, K.V. Shah, P.J. Snijders, C.J. Meijer, Interna-tional Agency for Research on Cancer Multicenter CervicalCancer Study Group, Epidemiologic classification of humanpapillomavirus types associated with cervical cancer, N. Engl.J. Med. 348 (2003) 518–527.
[3] L. Denny, M. Quinn, R. Sankaranarayanan, Chapter 8: Screen-ing for cervical cancer in developing countries, Vaccine 24(Suppl. 3: S3) (2006) 71–77.
[4] J. Ferlay, H.R. Shin, F. Bray, D. Forman, C. Mathers, D.M.Parkin, Estimates of worldwide burden of cancer in 2008:GLOBOCAN 2008, Int. J. Cancer 127 (2010) 2893–2917.
[5] A. Jemal, F. Bray, M.M. Center, J. Ferlay, E. Ward, D. Forman,Global cancer statistics, CA Cancer J. Clin. 61 (2011) 69–90.
[6] A.M. Massing, W.L. Epstein, Natural history of warts. Atwo-year study, Arch. Dermatol. 87 (1963) 306–310.
[7] P.K. Nicholls, P.F. Moore, D.M. Anderson, R.A. Moore, N.R.Parry, G.W. Gough, M.A. Stanley, Regression of canine oralpapillomas is associated with infiltration of CD4+ and CD8+Lymphocytes, J. Virol. 283 (2001) 31–39.
[8] M. Claus, J. Greil, C. Watzl, Comprehensive analysis of NK cellfunction inwhole blood samples, J. Immunol. Methods 341 (2009)154–164.
[10] I. Heard, J.M. Tassie, V. Schmitz, L. Mandelbrot, M.D.Kazatchkine, G. Orth, Increased risk of cervical disease amonghuman immunodeficiency virus-infected women with severeimmunosuppression and high human papillomavirus load, Obstet.Gynecol. 96 (2000) 403–409.
[11] P.H. Smits, R. Bakker, E. Jong, J.W. Mulder, P.L. Meenhorst, B.Kleter, L.J. van Doorn, W.G. Quint, High prevalence of Humanpapillomavirus infections in urine samples from human immu-nodeficiency virus-infected men, J. Clin. Microbiol. 43 (2005)5936–5939.
[12] Z.Z.A. Mbulawa, D.J. Marais, L.F. Johnson, D. Coetzee, A.Williamson, Impact of Human immunodeficiency virus onnatural history of human papillomavirus genital infection onSouth African men and women, J. Infect. Dis. 206 (2012) 15–27.
[13] D. Mavilio, G. Lombardo, J. Benjamin, D. Kim, D. Follman, E.Marcenaro, M.A. O’Shea, A. Kinter, C. Kovacs, A. Moretta, A.S.Fauci, Characterization of CD56–/CD16 natural killer (NK)
219Defects in genital wart NK cells associated with HIV
cells: a highly dysfunctional NK subset expanded in HIV-infectedviremic individuals, Proc. Natl. Acad. Sci. U. S. A. 102 (2005)2886–2891.
[14] E. Brunetta, K.L. Hudspeth, D. Mavilio, Pathologic naturalkiller cell subset redistribution in HIV-1 infection: new insightsin pathophysiology and clinical outcomes, J. Leukoc. Biol. 88(2010) 1119–1130.
[15] G. Alter, J.M. Malenfant, M. Altfeld, CD107a as a functionalmarker for the identification of natural killer cell activity, J.Immunol. Methods 294 (2004) 15–22.
[16] A. Poli, T. Michel, M. Thérésine, E. Andrès, F. Hentges, J.Zimmer, CD56bright natural killer (NK) cells: an important NKcell subset, J. Immunol. 126 (2009) 458–465.
[17] C.L. Sentman, T.F. Mselle, S. Basu, Human mucosal NK cells,in: L. Brossay (Ed.), Everything you always wanted to knowabout NK cells but were afraid to ask, Transworld ResearchNetwork, Kerala, India, 2007, pp. 99–115.
[18] M.A. Stanley, Immune responses to human papilloma viruses,Indian J. Med. Res. 130 (2009) 266–276.
[19] M.A. Stanley, Epithelial cell responses to infection with humanpapillomavirus, Clin. Microbiol. Rev. 25 (2012) 215–222.
[20] M. Prakash, S. Patterson, M.S. Kapembwa, Evaluation of thecervical cytobrush sampling technique for the preparation ofCD45+ mononuclear cells from the human cervix, J. Immunol.Methods 258 (2001) 37–46.
[21] N.N. Nkwanyana, P.P. Gumbi, L. Roberts, L. Denny, W.Hanekom, A. Soares, B. Allan, A.L. Williamson, D. Coetzee,A.J. Olivier, W.A. Burgers, J.A. Passmore, Impact of HIV-1infection and inflammation on the composition and yield ofcervical mononuclear cells in the female genital tract, Immunol-ogy 128 (2009) e746–e757.
[22] B.G. Brenner, A. Dascal, R.G. Margolese, M.A. Wainberg, Naturalkiller cell function in patients with acquired immunodeficiencysyndrome and related diseases, J. Leukoc. Biol. 46 (1989) 75–83.
[23] M.R. Goodier, N. Imami, G. Moyle, B. Gazzard, F. Gotch, Lossof the CD56hiCD16- NK cell subset and NK cell interferon-gammaproduction during antiretroviral therapy for HIV-1: partial recoveryby human growth hormone, Clin. Exp. Immunol. 134 (2003)470–476.
[24] M.B. Lucia, N. Froio, E. Tacconelli, M. Tubarello, S. Rutella, C.Rumi, R. Cauda, CD16 + CD56 + CD8+ natural killer (NK) cellsare decreased during HIV infection, Eur. J. Histochem. 41(Suppl. 2) (1997) 197–198.
[25] G. Alter, N. Teigen, B.T. Davis, M.M. Addo, T.J. Suscovich,M.T. Waring, H. Streeck, M.N. Johnston, K.D. Staller, M.T.Zaman, X.G. Yu, M. Lichterfeld, N. Basgoz, E.S. Rosenboer, M.Altfeld, Sequential deregulation of NK cell subset distributionand function starting in acute HIV-1 infection, Blood 106 (2005)3366–3369.
[26] G. Alter, M. Altfeld, NK cells in HIV‐1 infection: evidence fortheir role in the control of HIV‐1 infection, J. Intern. Med. 265(2008) 29–42.
[27] D. Mavilio, G. Lombardo, A. Kinter, M. Fogli, A. La Sala, S.Ortolano, A. Farschi, D. Follmann, R. Gregg, C. Kovacs, E.Marcenaro, D. Pende, A. Moretta, A.S. Fauci, Characterization of
the defective interaction between a subset of natural killer cellsand dendritic cells in HIV-1 infection, J. Exp. Med. 203 (2006)2339–2350.
[28] C.A. Sabin, C.J. Smith, A.D. Monforte, M. Battegay, C.Gabiano, L. Galli, S. Geelen, D. Gibb, M. Guiguet, A. Judd, C.Leport, F. Dabis, N. Pantazis, K. Porter, F. Raffi, C. Thorne, C.Torti, S. Walker, J. Warszawski, U. Wintergerst, G. Chene, J.Lundgren, I. Weller, D. Costagliola, B. Ledergerber, J.Lundgren, G. Chene, G. Touloumi, J. Warszawski, L. Meyer,F. Dabis, M.M. Krause, C. Goujard, C. Leport, F. de Wolf, P.Reiss, K. Porter, M. Dorrucci, C. Sabin, J. Amo, N. Obel, A.Mocroft, O. Kirk, S. Staszewski, S. Perez-Hoyos, J. Almeda, A.Antinori, P.A. Tovo, B. Salzberger, G. Fatkenheuer, J. Ramos,M. Battegay, C. Mussini, P. Tookey, J. Casabona, J.M. Miro, A.Castagna, S. Wit, R. Teira, M. Garrido, N. Dedes, C. Sabin, A.Phillips, H. Furrer, M. Egger, M.L. Newell, J. Sterne, A. Telenti,Response to combination antiretroviral therapy: variation byage, AIDS 22 (2008) 1463–1473.
[29] P.J. Weidle, S. Malamba, R. Mwebaze, C. Sozi, G. Rukundo, R.Downing, D. Hanson, D. Ochola, P. Mugyenyi, J. Mermin, B.Samb, E. Lackritz, Assessment of a pilot antiretroviral drugtherapy programme in Uganda: patients' response, survival,and drug resistance, Lancet 360 (2002) 34–40.
[30] S. Grabar, V. Le Moing, C. Goujard, C. Leport, M.D. Kazatchkine,D. Costagliola, L. Weiss, Clinical outcome of patients with HIV-1infection according to immunologic and virologic response after 6months of highly active antiretroviral therapy, Ann. Intern. Med.133 (2000) 401–410.
[31] S.H. Tuboi, M.W.G. Brinkhof, M. Egger, R.A. Stone, P. Braitstein,D. Nash, E. Sprinz, F. Dabis, L.H. Harrison, M. Schechter,Discordant responses to potent antiretroviral treatment inpreviously naive HIV-1–infected adults initiating treatment inresource-constrained countries, J. Acquir. Immune Defic. Syndr.45 (2007) 52–59.
[32] S.D. Lawn, L. Myer, G. Harling, C. Orrell, L.G. Bekker, R.Wood, Determinants of mortality and nondeath losses froman antiretroviral treatment service in South Africa: implica-tions for program evaluation, Clin. Infect. Dis. 43 (2006)770–776.
[33] R. Tan, A.O. Westfall, J.H. Willig, M.J. Mugavero, M.S. Saag,R.A. Kaslow, M.C. Kempf, Clinical outcome of HIV-infectedantiretroviral-naive patients with discordant immunologic andvirologic responses to highly active antiretroviral therapy, J.Acquir. Immune Defic. Syndr. 47 (2008) 553–558.
[34] E. Vivier, E. Tomasello, M. Baratin, T. Walzer, S. Ugolini,Function of natural killer cells, Nat. Immunol. 9 (2008)503–510.
[35] M.A. Cooper, T.A. Fehniger, S.C. Turner, K.S. Chen, B.A.Ghaheri, T. Ghayur, W.E. Carson, M.A. Caligiuri, Humannatural killer cells: a unique innate immunoregulatory rolefor the CD56 (bright) subset, Blood 97 (2001) 3146–3151.
[36] A. Moffet-King, Natural killer cells and pregnancy, Nat. Rev.Immunol. 2 (2002) 656–663.