Page 1
Chapter 8
Cornea
MICHAEL K. SMOLEK and STEPHEN D. KLYCE
Main Menu Table Of Contents
Search
SURFACE ANATOMY
TEAR FILM
EPITHELIUM
ANCHORING STRUCTURES
BOWMAN'S LAYER
STROMA
DESCEMET'S MEMBRANE
ENDOTHELIUM
CONCLUSION
ACKNOWLEDGMENT
REFERENCES
As a portion of the ocular tunic, the cornea
protects the delicate intraocular contents with
its tough, yet pliable, collagen structure. It is
remarkable that a tissue with this ability to
resist injury can provide the essential optics
and transparency to focus an image on the retina.
In this chapter the gross, microscopic, and
ultramicroscopic anatomic structure of the cornea
is described. Corneal physiology is not
discussed, except when structural features can be
more clearly defined by a description of their
physiologic significance. An effort has been made
Page 2
to cite references primarily for normal adult
human corneas; references to other species are
made when human information is lacking or
ambiguous. Citations to embryonic development are
included when they provide better insight into
the adult anatomy of the cornea.
Back to Top
SURFACE ANATOMY
The anterior and posterior surfaces of the human
cornea are often approximated in schematic eye
calculations by radii of curvature of 7.8 and 6.5
mm, respectively, compared to the external
surface of the scleral globe, which has a radius
of approximately 11.5 mm.1 The anterior corneal
surface becomes slightly flatter in the
periphery, giving the overall cornea a naturally
prolate shape. If the cornea were perfectly
spherical, it would suffer from considerable
spherical aberration in which the rays passing
through the peripheral parts of an optical system
do not come to the same focal point as the rays
near the central axis of the system. Peripheral
corneal flattening reduces, but does not entirely
eliminate, the corneal contribution of spherical
aberration to the optical system of the eye. The
crystalline lens provides additional correction
of the residual spherical aberration of the
Page 3
cornea, depending on the accommodative power of
the lens. The radius of curvature of the anterior
surface translates into a vergence power of
approximately 48.8 diopters (D), which accounts
for roughly three quarters of the total
refractive power of the eye's optical system. The
posterior corneal surface adds negative power so
that the total power of the typical, normal
cornea is approximately 43 D.
The difference in curvature between the
relatively flatter anterior surface and the
relatively steeper posterior surface is
associated with the central cornea being thinner
than the periphery. Maurice reported a central
thickness value of 0.52 mm and a peripheral value
of approximately 0.65 mm in adult humans when
measured with an optical pachometer.2 Abnormal
tissue thinning may be indicative of a corneal
dystrophy, such as keratoconus, Terrien's
marginal degeneration, or pellucid marginal
degeneration.
The cornea protrudes slightly beyond the limits
of the scleral globe because of the difference in
curvature between the relatively steep cornea and
the relatively flat globe. A shallow sulcus is
formed at the intersection of the corneal and
scleral surfaces, which roughly demarcates a
Page 4
region called the limbus. The limbus is typically
defined in one of two ways. The histologic limbus is
the full-thickness annular interface that
separates the optically transparent corneal
stroma from the opaque sclera. The surgical limbus
is the annular region bound by a line from the
anterior surface termination of Bowman's layer to
the posterior surface termination of Descemet's
membrane and by a line oriented perpendicular to
the external scleral surface that intersects
Schlemm's canal in the angle of the anterior
chamber.
Externally, the cornea appears elliptical with
its vertical chord shorter than its horizontal
chord (10.6 versus 11.7 mm for males and 9.6
versus 10.7 mm for females).2 This difference
arises from opaque scleral tissue extending over
the anterior corneal margin slightly more along
its superior and inferior aspects. When viewed
from within the dissected globe, the posterior
cornea appears circular with a diameter of 11.7
mm. The external corneal surface area is
approximately 120 mm2, or one-fourteenth the
total area of the ocular globe.3 Surprisingly,
the surface area is not significantly increased
by the localized distension found in keratoconus,
although keratoglobus conditions do appear to
Page 5
have a greater surface area.
Back to Top
TEAR FILM
The precorneal tear film is approximately 7 μm
thick with a volume of 6.2 ± 2 μL during normal
tear production.4 Tear fluid is typically
produced at a rate of 1.2 μL/minute, with a major
portion drained from the palpebral fissure
through the nasolacrimal duct and a smaller
volume lost through evaporation from the ocular
surface (approximately 3 μL/hour at 30% relative
humidity).5 Tear chemistry is complex;
ingredients include various electrolytes,
metabolites, proteins, enzymes, and lipids.6 The
functional significance of the tear film is
broad. It provides lubricating qualities and a
smooth optical interface with the air. It also
protects the epithelium from airborne
contaminants and provides natural immunity to
infectious agents through secretory
immunoglobulin molecules.7
The tear film is composed of the cellular
exudates of separate structures of the ocular
adnexa and has been difficult to measure
precisely because of its delicate fluid
nature.8 The anterior-most layer is the lipid or
Page 6
oily layer derived from secretions of the
meibomian glands located in the eyelid and
caruncle and is between 0.1 and 0.5 μm thick. The
aqueous lacrimal tear layer is at least 5 μm
thick and often thicker, depending on tear
production by the lacrimal glands located in the
superiotemporal margin of the orbit. The
posterior mucous layer is approximately 1 μm
thick and is derived from secretions of
conjunctival goblet cells. The hydrophilic nature
of mucus substantially reduces surface tension
and provides a smooth, wettable surface for the
aqueous tear layer. The separation between the
middle lacrimal tear layer and the posterior
mucous layer is actually indistinct, making
definitive thickness measurements of the two
components difficult. It is highly likely that
these two components form a graded mixture, with
the posterior mucous component gradually blending
into the anterior aqueous lacrimal tear
component.
Back to Top
EPITHELIUM
The corneal epithelium is the anterior-most cell
layer of the cornea (Fig. 1). It is typically
several cell layers thick, consisting of the
Page 7
apical cell squamous layer, the multilayered,
polygonal-shaped wing cells beneath the apical
layer, and the posterior-most layer of basal
cells (Fig. 2). The wing cell layer is two or
three cells thick in the central cornea, but
tends to be four to five cells thick in the
periphery. In total, the epithelium is
approximately 50 μm thick in the central human
cornea.
Fig. 1. Full-thickness view of the normal
human cornea. From top: epithelium,
Bowman's layer (arrow), stroma,
Descemet's membrane (arrow), and endothelium
(hematoxylin-eosin stain, 80×). (Courtesy of Drs.
Rodrigues, Waring, Hackett, and Donohoo.)
Fig. 2. Full-thickness electron
micrograph of corneal epithelium. Note
the cell shape change with depth, the
variation of cell membrane
interdigitation, and the intracellular
differences between cell types. S, apical surface
cells; W, wing cells; and B, basal cells. Also
note the microvilli seen along the apical
membrane of the surface cells (3,620×). Inset:
Epithelium (E) overlies a thin, dense basement
Page 8
membrane (arrow) with no discernible laminar
appearance (periodic acid-Schiff [PAS] stain,
330×). (Courtesy of Drs. Rodrigues, Waring,
Hackett, and Donohoo.)The epithelium is in a constant state of
turnover, with exfoliating apical cells being
replaced by underlying wing cells. During normal
exfoliation, desquamating cells are released only
after the replacement cell has established new
tight junctions with neighboring cells and the
new apical membrane is capable of maintaining
continuity of the tear film.9 Studies of induced
exfoliation of a monolayer of epithelial cells
with a biologic detergent indicate recovery of
the paracellular barriers and transepithelial
electric resistance in approximately 1 hour.10 The
epithelium completely turns over in approximately
7 days.11 Once injured, a high degree of motility
ensures coverage of a denuded area by adjacent
basal cells, followed by replacement of the
normal complement of cell layers. Basal cells are
the only epithelial cells capable of mitosis;
however, many epithelial cells originate as the
progeny of limbal stem cells and migrate
centripetally to supplement or replace cells lost
through normal desquamation or injury.12–14 Using
immunohistochemical staining for antibodies to
keratins, Wiley and associates found regional
Page 9
heterogeneity indicating that the superior
corneal periphery and limbus have the greatest
numbers of stem cells producing replacement
epithelial cells.15 Limbal stem cell deficiency
may result in conjunctival epithelium invasion of
the cornea, leading to vascularization, the
appearance of goblet cells, and an irregular or
unstable epithelium that reduces visual acuity
and may produce pain or discomfort.16
The epithelium is known to chemically interact
with keratocyte cells of the stroma. These
interactions appear to be dominated by cytokines
such as interleukin-1 (IL-1) and soluble Fas
ligand that are released by injured epithelial
cells. It would appear that IL-1 is a master
regulator for corneal wound healing given its
effect on keratocyte apoptosis and the modulation
of matrix metalloproteinase and growth factors
such as keratinocyte growth factor (KGF) and
hepatocyte growth factor (HGF). The Fas ligand
system is known to influence the immune
privileged state of the cornea. In addition to
the epithelial-to-keratocyte communication,
keratocytes influence the state of the epithelium
via HGF and KGF, which affect cell turnover,
motility, and proliferation.17
APICAL CELLS
Page 10
Apical surface cells appear broad and flattened:
4 to 5 μm thick and 40 to 50 μm in diameter.
Freshly emerged surface cells appear bright
during specular microscopy and have relatively
small numbers of microvilli covering their apical
membrane (Fig. 3).18
Fig. 3. Scanning electron micrograph
of corneal epithelium surface. Note
the numerous microvilli and the
cellular margins (12,900×).(Courtesy of Drs.
Rodrigues, Waring, Hackett, and Donohoo.)As the cell matures, its specular microscopic
appearance tends to darken (Fig. 4). This may be
due to changes in surface texture, because
microvilli densely cover the apical membrane at
this stage of development.4,19 Prior to cell
exfoliation, apical surface margins tend to
appear smooth, with microplicae clustered only
near the center. In its final stages the
biomicroscopic appearance of the cell surface
appears darker than in its earlier stages (Fig.
5).
Fig. 4. Confocal microscopic transverse
image of the human surface epithelium in
vivo. The squamous cell nuclei and some cell
Page 11
margins are visible (500×). (Courtesy of Nidek
Technologies.)
Fig. 5. Scanning electron micrograph of
the anterior surface of the corneal
epithelium. Note that the darker cells
have fewer microvilli or microplicae near the
cell margins (1,500×). (Courtesy of Drs.
Rodrigues, Waring, Hackett, and Donohoo.)The 300-nm thick glycocalyx (buffy cell coat) of
the apical membrane can be preserved intact for
histologic evaluation.7,8,19 The glycocalyx is
composed of glycoprotein material, and numerous
separate fine filaments become visible on the
apical surface after tannic acid staining (Fig.
6). These filaments cover the tips and sides of
the microplicae and microvilli extensively,
inserting into the cell membrane. Angular bends
and filament branching are evident, as well as a
beaded substructure.19 The shortest filaments are
150 nm in length in the central cornea, while
filaments 300 nm long are found on the
conjunctival cell surfaces.19 The glycocalyx binds
loosely with the overlying mucous layer and
provides binding sites for immunoglobulin present
in the tears.7
Page 12
Fig. 6. Transmission electron micrograph
showing the apical membrane of the
surface epithelium and intercellular
junctions along the lateral membranes.
Arrows indicate fine, branching filaments of the
glycocalyx emerging from the surface microvilli.
A tight junction (box) and densely staining
desmosomes are seen along the lateral membranes.
Glycogen is evident (circle), as well as numerous
vesicles (23,000×). Inset: Arrows point to
microvilli from two adjacent surface cells
(64,500×). (Courtesy of Drs. Rodrigues, Waring,
Hackett, and Donohoo.)The margins of the apical cell membrane possess
the important tight junctions surrounding the
cell circumference near the apical margin.20 This
junction complex is the correlate of the
paracellular pathway of high resistance to ion
flow. The lateral and basal membranes of the
apical cells have gap junctions, numerous
desmosomal junctions, and numerous membrane-bound
vesicles. The cytoplasm contains a flattened
nucleus (which probably disintegrates prior to
desquamation), few organelles, and a notable
increase in tonofilaments compared with the
underlying cells. There are some aggregates of
glycogen granules, small mitochondria, sparsely
distributed free ribosomes, and poorly developed
Page 13
Golgi's complexes. Cytoplasmic vesicles are
fairly numerous.21
WING CELLS
Wing cells are distinguished by a variety of
polygonal shapes and by their large ovoid nuclei.
The cells are roughly 12 to 15 μm in diameter,
and their cytoplasm contains few rough
endoplasmic reticulum cisternae, mitochondria, or
Golgi's complexes. The large numbers of
cytoskeletal tonofilaments are approximately 8 nm
in length, and numerous interdigitations exist
along the cell membranes.21 Desmosomal and gap
junctions are seen between adjacent wing cells
and between basal and apical cells (Fig. 7).
Fig. 7. Transmission electron
micrograph of elaborate
interdigitations between epithelial
wing cell membranes. Desmosomes are
seen along the cell walls, as well as
intracellular tonofilaments (arrow).
Intracellular rough endoplasmic reticulum,
scattered mitochondria, and ribosomes are seen in
addition to the prominent nuclei (13,800×).
Inset: Desmosome junction (49,500×). (Courtesy of
Drs. Rodrigues, Waring, Hackett, and Donohoo.)BASAL CELLS
Page 14
Basal cells appear as elongated polygonal cells
approximately 10 μm in width and 15 to 20 μm in
height, with prominent ovoid nuclei. The
polygonal nature can be readily discerned in
confocal microscopy images of the basal cell
layer (Fig. 8). Buschke and associates22 showed
that cell mitosis occurs in only 1 of 250 basal
cells of the rat epithelium. In their study,
mitosis occurred in irregular clumps of three to
six cells, and mitotic cells were much more
numerous in the periphery. The basal cell
cytoplasm appears similar to that of the wing
cells, as do the anterior and lateral cell
membranes with their complement of desmosomal
attachments. However, the basal membrane is
notable for the presence of hemidesmosomes, which
are discussed below.
Fig. 8. Confocal microscopic transverse
image of the human basal epithelium in
vivo. The cells are outlined by a fine polygonal
mesh which is the source of haloes around bright
lights at night (Sattler's veil) that occurs with
epithelial edema (500×). (Courtesy of Nidek
Technologies).
Back to Top
Page 15
ANCHORING STRUCTURES
HEMIDESMOSOMES
The epithelial basal cell hemidesmosome is a
multilayered junctional structure associated with
the basal membrane, with intracellular keratin
filaments (intermediate filaments) entering along
its cytoplasmic aspect. As basal epithelial cells
migrate in response to wound healing,
hemidesmosome junctions are observed to
disassemble and their integrin components
diffusely distribute across the basal cell
membrane.23 In vitamin A-deficient rat models, the
severity of epithelial sloughing was found to
correspond to the size and frequency of residual
hemidesmosomes.24
Kurpakus and colleagues25 found a monoclonal
antibody, mAb6A5, that was directed against a
200-kd polypeptide component on the cytoplasmic
exposure of the hemidesmosome. This component may
be associated with the electron-dense plate that
is seen crossing through the intermediate
filaments just proximal to the hemidesmosome
plaque.26 Short filaments pass from the dense
plate into the plaque. Polypeptides of 180 and
230 kd were isolated by Owaribe and co-
workers26 and localized to the hemidesmosome
plaque. The 230-kd component was found to be the
Page 16
bullous pemphigoid antigen (BPA), while the 180-
kd component was heavily glycosylated, making it
a candidate for a transmembrane glycoprotein. In
addition, major polypeptides of 120, 200, and 480
kd were isolated from the region and may be
hemidesmosome structural components as well.26
Stepp and associates,27 using direct
immunofluorescence of frozen sections, found β1,
β4, and β5 integrin subunits and α3, α5, α6, and αv
subunits localized in the epithelium.
Specifically, α5, α6, and β4 subunits of integrin
heterodimers were found to be localized on the
basal membrane side of the hemidesmosome site,
while β1, β5, α2, α3, and αv were localized to
sites associated with cell-t-cell contact.27 A
125-kd polypeptide was isolated to the basal side
of the hemidesmosome by Kurpakus and co-
workers28 with the monoclonal antibody mAbHD. This
polypeptide may be a protein associated with the
anchoring filaments that span the lamina lucida
from the hemidesmosome plaque to the basal
lamina.
Fluorescence confocal imaging of the integrin
subunits α6 and β4 indicated that the α6 subunits
were localized to both the basal and lateral
surfaces of the basal epithelial cell, but the
Page 17
β4 subunits were present only along the basal
floor of the cell. The number and distribution of
hemidesmosomes do not appear to vary with age;
however, discontinuous staining for the
α6 subunits in corneal epithelium of individuals
over the age of 70 has been noted.29
BASAL LAMINA
Basement membrane is secreted extracellularly by
the epithelial cells and forms one of several
structural components associated with cell
adhesion to Bowman's layer or stroma. With light
microscopy, a densely stained basement membrane
approximately 75 to 100 nm thick is
visible.21 Under the high magnifications of
electron microscopy, this layer appears
distinctly laminated. Anteriorly, the lamina
lucida appears as a clear, electron-lucent zone
approximately 23 nm thick, while the lamina densa
that lies apposed to Bowman's layer is an
electron-dense region approximately 48 nm
thick.21 It has not been established how basal
lamina adheres to Bowman's layer. A third
component of the basement membrane is the
reticular lamina, which lies distal to the lamina
densa and is within Bowman's layer. This region
includes the anchoring fibrils and plaques and
Page 18
other electron dense materials associated with
the lamina densa (Fig. 9).
Fig. 9. Transmission electron
micrograph of epithelial basal
lamina in the human cornea,
Bowman's layer is seen below,
with basal epithelial cells
above. Note the desmosome junction seen centrally
between two basal cells (arrowhead), Magnified
view appears in upper right box. Multiple
hemidesmosomes are seen along the basal lamina
(curved arrows) with platelike features seen
within the lamina lucida. Magnified view of a
hemidesmosome is seen in the box, lower right,
Bar = 0.5 μm. (Courtesy of Roger Beuerman, Ph.D.,
New Orleans, Louisiana.)Numerous electron-dense, fine filaments are
observed clustered within the lamina lucida; they
course from the hemidesmosomes of the basal
epithelium and insert into neighboring regions on
the lamina densa. Within the lamina lucida, a
dense plate often can be seen interposed along
the filaments and lying parallel to the
hemidesmosome plaque. The precise composition of
these filaments and the dense plate is uncertain,
but the 125-kd polypeptide described previously
Page 19
may be involved.28 In addition, the antihuman
monoclonal antibody, 19-DEJ-l, was found to be
reactive to the antigenic epitope of either the
anchoring filaments or the dense plate through
the filaments.30
ANCHORING FIBRILS AND PLAQUES
Within the anterior aspect of Bowman's layer,
fine filaments travel distally from the lamina
densa near the proximal insertion of the
anchoring filaments and immediately coalesce to
form striated anchoring fibrils approximately
0.15 μm in width (Fig. 10).31 These anchoring
fibrils then course posteriorly to an average
depth of 0.60 μm in the adult human and terminate
in electron-dense regions called anchoring
plaques.31,32 Maximum penetration depths of 2.05 μm
were reported for the anchoring fibrils in
humans. Some anchoring fibrils may terminate
among the type I collagen fibrils31 or return
proximally to reinsert into the lamina densa at a
neighboring site.32 There also appear to be
plaque-to-plaque anchoring fibrils. Anchoring
fibrils found in Bowman's layer appear similar to
those found in the stroma of the rabbit, a
species that lacks Bowman's layer.31
Page 20
Fig. 10. Transmission electron
micrograph of the epithelial
basal lamina in rabbit cornea.
Large micrograph shows anchoring
filaments within the lamina lucida, oriented
between the hemidesmosomes of the basal
epithelial cells (HD) and the lamina densa.
Anchoring fibrils (AF) travel distally (large
arrows) from the lamina densa to insert into
electron-dense anchoring plaques (small arrows)
(73,900×). Inset right: Cross-banding on the
anchoring fibril. Inset left: An anchoring fibril
inserting into two adjacent lamina densa sites.
(From Gipson IK, Spurr-Michaud SJ, Tisdale AS:
Anchoring fibrils form a complex network in human
and rabbit cornea. Invest Ophthalmol Vis Sci
28:212, 1987.)Anchoring fibrils were found to be type VII
collagen filaments in bovine32 and human
corneas31 using immunogold labeling and
immunofluorescence staining techniques,
respectively. Type VII collagen is a dimer of
high molecular weight.33 Each monomer has a
helical domain and a globular, carboxyl domain.
The dimer is created by disulfide bonds linking
the tails of the monomers together, and striated
fibrils are formed by the nonstaggered
arrangement of the helical filaments. The
Page 21
globular domains were found localized within the
anchoring plaques, which also label for type IV
collagen.30 Gipson and co-workers31 and Keene and
colleagues32 presented similar models for the
architecture of the anchoring fibril network and
indicated that the striated type I collagen
fibrils in Bowman's layer must be interwoven and
trapped by the anchoring fibril network (Fig.
11).
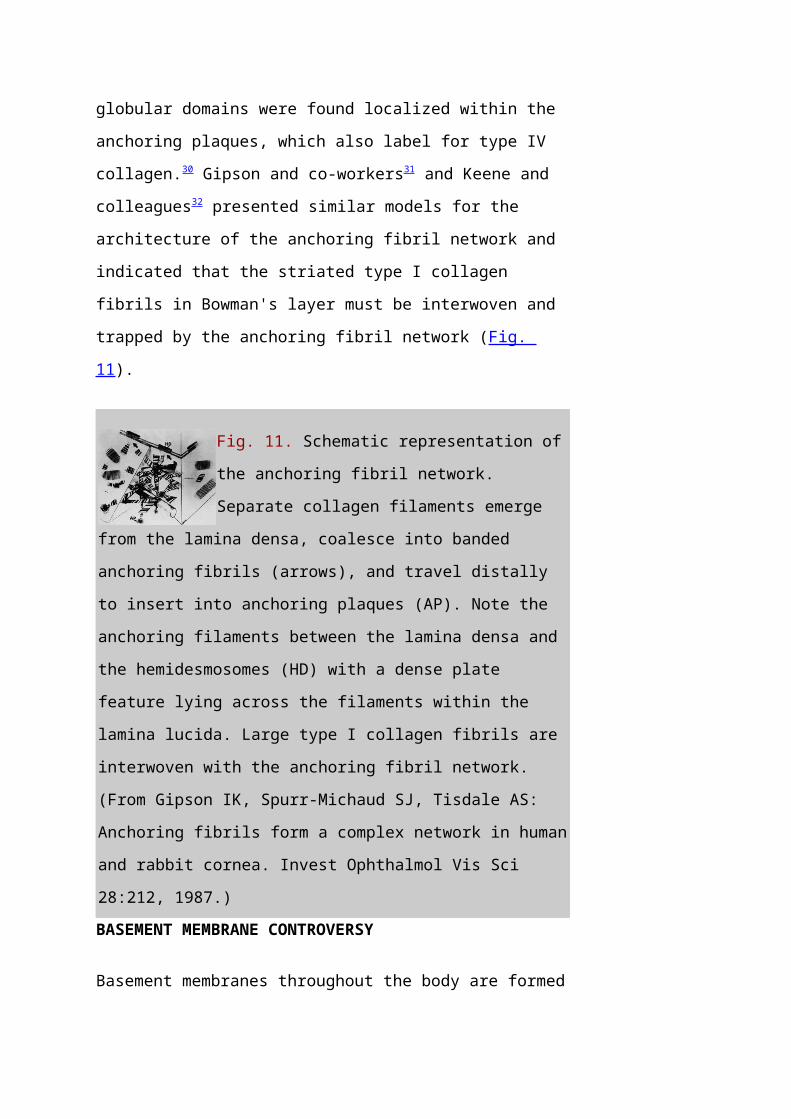
Fig. 11. Schematic representation of
the anchoring fibril network.
Separate collagen filaments emerge
from the lamina densa, coalesce into banded
anchoring fibrils (arrows), and travel distally
to insert into anchoring plaques (AP). Note the
anchoring filaments between the lamina densa and
the hemidesmosomes (HD) with a dense plate
feature lying across the filaments within the
lamina lucida. Large type I collagen fibrils are
interwoven with the anchoring fibril network.
(From Gipson IK, Spurr-Michaud SJ, Tisdale AS:
Anchoring fibrils form a complex network in human
and rabbit cornea. Invest Ophthalmol Vis Sci
28:212, 1987.)BASEMENT MEMBRANE CONTROVERSY
Basement membranes throughout the body are formed
Page 22
primarily from type IV collagen, yet evidence for
type IV collagen within corneal lamina densa
remains sketchy and controversial. Marshall and
associates were unable to demonstrate the
presence of type IV collagen in the human basal
lamina using immunoelectronmicroscopy labeling
with colloidal gold.34 Indirect immunofluorescence
labeling for anti-type IV antigens was shown
within a subepithelial band by Konomi and
colleagues,35 but the layer did not appear
strongly labeled. Newsome and co-
workers36 indicated strong immunofluorescence
labeling, but the corneal location was not
specified. Nakayasu and associates37 showed strong
immunofluorescence labeling of a subepithelial
layer near the limbus but did not report type IV
labeling for the concomitant
immunoelectronmicroscopy portion of the study.
In a project designed to resolve this
controversy, Kolega and colleagues38 found that
basement membrane type IV collagen was
differentially stained according to location in
the adult human. There was no immunofluorescent
staining for type IV collagen in the central
cornea using three different monoclonal
antibodies, and only weak staining in the corneal
periphery. However, there was intense staining in
Page 23
the conjunctiva and across the limbal region.
Cleutjens and colleagues39 and others40,41 also
found no type IV collagen in the central cornea
of the adult but did find it in the periphery.
Furthermore, type IV collagen was found to be
strongly labeled in the fetal central cornea.39
Linsenmayer and associates42 had previously
discussed findings of heterogeneous type IV
collagen distribution in a variety of basement
membranes throughout the body, including the
crystalline lens capsule.42 Thus, it would be
unusual, but not unexpected, for corneal
epithelial basement membrane to exhibit a
heterogeneous distribution. If type IV collagen
is missing, binding mechanisms in the lamina
densa would have to be reconsidered. This also
would have implications for how laminin, heparan
sulfate proteoglycan, and fibronectin binding
operate in the absence of type IV collagen.
It is possible that the type IV collagen present
in the human cornea does not react to any anti-
type IV antibodies known to date.43 A second
possibility may be that type IV collagen is
masked to specific immunolabels by proteoglycans
surrounding the collagen.40,41,44 Finally, type IV
collagen that is seen in the embryologic basement
Page 24
membrane may diminish with maturation of the
tissue. Cleutjens and co-workers39 have stated
that the underlying Bowman's layer, because it is
acellular, may not provide the appropriate
signals to the epithelium to stimulate type IV
production. Any type IV that is present could be
either remnant embryonic material or produced by
progeny cells of the limbal stem cells. This
would explain the differential localization of
type IV and indicate that progeny cells of the
stem cells lose their ability to produce type IV
collagen during migration toward the central
cornea.
To further complicate the issue, Cleutjens and
colleagues45 noted an absence or thinning of
lamina densa within the central human adult
cornea, despite electron-dense plaques, anchoring
fibers, and so forth, being present. They
speculate that there may be a correlation between
epithelial migration, the thinned appearance of
the basal lamina, and the missing type IV
collagen. However, Kolega and associates38 found
that the loss of type IV labeling was not
spatially correlated with the labeling of the
monoclonal antibody AE-5, a marker of 64-kd basic
keratin differentiation in epithelium. Finally,
it has been suggested that type VII collagen
Page 25
anchoring fibrils may not even need a complete
basal lamina structure as part of the anchoring
complex.45
Back to Top
BOWMAN'S LAYER
When viewed with electron microscopy, Bowman's
layer appears as a felt-like composite of
randomly oriented, striated collagen fibrils
dispersed throughout an amorphous matrix (Fig.
12). In an adult, this layer is approximately 8
to 12 μm thick, being slightly thicker in the
corneal periphery.21 Bowman's layer is acellular,
except for nerve axons coursing toward the
epithelium.46 Historically, electron microscopy
has suggested a lack of keratocyte (fibroblast)
cells within Bowman's layer, which called into
question its ability to regenerate after injury
or that keratocytes could migrate into Bowman's
layer. In confocal microscopy views of the living
eye, numerous keratocytes are observed at the
level of Bowman's layer (Fig. 13).
Fig. 12. Bowman's layer of the
human cornea. Stromal collagen
fibers (S) insert into the
posterior aspect of Bowman's
layer (B). The anterior aspect of Bowman's layer
Page 26
appears smooth but slightly irregular. The lamina
densa appears as a continuous, dense band
adjacent to numerous hemidesinosomes along the
basal epithelium (E). Bar = 2 μm. (Courtesy of
Roger Beuerman, Ph.D., New Orleans, Louisiana.)
Fig. 13. Confocal microscopic transverse
image of the human Bowman's layer in
vivo. Nerve axons course freely through
the structure and keratocyte nuclei appear
numerous and more refractile than deeper in the
stroma. (500×). (Courtesy of Nidek Technologies.)Bowman's layer exists in primates and a few
additional species (most notably in birds). This
is of interest, particularly in terms of the
biomechanical strength and stiffness that
Bowman's layer appears to contribute to the
cornea. Nonprimate animals are poor models
compared to primates when predicting clinical
results with keratorefractive surgical
procedures. Biomechanical computer modeling of
the cornea with anisotropic material properties
is still in its infancy, but recent efforts may
be extended for a fuller appreciation of the
contribution of Bowman's layer to corneal shape
after surgery.47
Bowman's layer is composed of collagen types I,
Page 27
III, V, and VI as shown by
immunoelectronmicroscopy34,37,48 and
immunofluorescence microscopy,35,36 with type I
constituting the bulk of collagen present. Type
IV and VII collagens are present in association
with the anchoring complexes described in the
previous section. While type I collagen is absent
from the amorphous background matrix,34 it has
been localized with immunogold labeling to
striated fibrils having uniform diameters of 20
to 25 nm and 67-nm banding.46 Use of
immunofluorescence microscopy for antigens
specific to type I collagen has proven less
reliable, because there have been reports of both
its presence36 and absence37 in Bowman's layer.
The existence of type III collagen anywhere in
the eye has been controversial, but recent
immunoelectronmicroscopy findings support its
presence in Bowman's layer in the normal human
adult.34,36 Type III collagen was confirmed in the
embryonic chick cornea with immunofluorescence
labeling, but was found lacking in adult avian
tissue.49 It has been found in diseased and
wounded human corneas.36,37 The function of type
III collagen has been linked to control of fibril
diameter and uniformity when hybridized with type
Page 28
I collagen.37
Type V collagen also has been implicated as a
fibril diameter-controlling collagen in
heterotypic association with type I in the
embryonic chick cornea.50,51However, type V
labeling sites on the predominantly type I fibril
could be found only after considerable mechanical
disruption of these fibrils. Conversely,
immunofluorescence labeling for type V collagen
occurred throughout Bowman's layer in the human
adult without mechanical disruption37 and was
confirmed by immunoelectronmicroscopy.37,48 The
dichotomy between species may be explained by
labeling sites being more exposed on the human
fibrils than on avian fibrils, and this also may
depend on masking by the surrounding
proteoglycans.
Using immunogold labeling, type VI collagen was
found abundantly distributed as fine filaments
throughout Bowman's layer.48 It was not associated
with type I collagen striated fibrils. As with
many other collagens, the precise function of
type VI collagen is unknown, but it is ubiquitous
in connective tissues throughout the body33 and
may contribute to the collagen matrix in which
striated collagens are embedded. It is not found
Page 29
in the primary stroma of the embryonic avian eye
until after fibroblasts arrive to lay down the
secondary stroma.52
The anterior surface of Bowman's layer is sharply
defined by its interface with the lamina densa of
the overlying basal lamina; however, this
interface is not necessarily smooth.46 When viewed
with scanning electron microscopy, the surface
appears undulating, with a woven texture and
occasional pores. These pores are 0.5 to 1.5 μm
in diameter and are likely to be the channels
through which corneal nerve axons travel to enter
the epithelium.46
In the posterior aspect of Bowman's layer,
striated collagen fibrils from the underlying
stroma become contiguous with Bowman's layer.
Individual fibrils splay outward into the
amorphous matrix.46 Occasionally, relatively large
bundles of stromal fibrils course obliquely from
the midstromal regions and terminate eventually
in Bowman's layer.53 These fibril bundles
presumably provide considerable cohesion between
the underlying stroma and Bowman's layer;
Bowman's layer cannot be stripped away from
stroma as a continuous sheet as can Descemet's
membrane. As a consequence of the random
Page 30
insertions by the stromal fibrils, there is only
a roughly defined interface between the two
layers when viewed under high magnification.
Back to Top
STROMA
The bulk of the cornea consists almost entirely
of the corneal stroma, a fibrous tissue layer
approximately 450 μm thick in the central cornea
(Fig. 14). As determined by biochemical and
immunohistologic methods, the stroma is composed
predominantly of type I collagen with types III,
V, and VI also in evidence.34,37,48 Immunogold
labeling was intense for collagen type VI, which
is associated with the interfibrillar matrix only
and not localized to the striated fibrils.48 Type
III and V collagens are codistributed with
striated type I collagen. This codistribution is
similar to the codistribution of types III and V
in Bowman's layer. Type V and III collagens also
have been labeled at the interfacial matrix
separating stroma and Descemet's membrane.34,48
Fig. 14. Stromal collagen fibrils (c) in
uniform spatial arrangement within
orthogonal lamellae. A portion of a
keratocyte is seen within the interlamellar
space. Granular material (asterisk) is visible
Page 31
adjacent to the cell body (30,000×). (Courtesy of
Drs. Rodrigues, Waring, Hackett, and Donohoo.)Type I collagen is organized into striated
fibrils 25 to 35 nm in diameter with periodic
banding according to electron microscopy.46 Type V
collagen appears colocalized with type I in the
striated fibers.54 Using atomic force microscopy,
corneal collagen was reported to have a D-
periodicity of 63.9 to 68.5 nm.55,56Scleral
periodicity banding occurs at 67-nm intervals.
Fibrils are composed of 4-nm microfibril
components arranged in a right-hand helix and
tilted at 15 degrees to the fibril axis in the
cornea and 5 degrees in the scleral. This
striated banding is the result of the offset,
stacked arrangement of individual procollagen
triple-helix molecules into units of five (the
microfibril), which are further combined to form
the basic unit of the collagen fibril. The 67-nm
axial repeat was shown to correspond to the N–
and C- telopeptids and the D-band periodic gap
zone.57
These fibrils are combined into highly ordered,
sheet-like bundles called lamellae, which lie
essentially parallel to the corneal surfaces.
Lamellae vary in width and thickness throughout
the stroma, with a tendency to have smaller
Page 32
dimensions anteriorly (0.5 to 30 μm wide and 0.2
to 1.2 μm thick) and larger dimensions
posteriorly (100 to 200 μm wide and 1 to 2.5 μm
thick).46 Hundreds of individual lamellae can be
discerned in a cross-sectional slice of the full-
thickness central cornea.
LAMELLAR ORGANIZATION
Fibrils within a given stromal lamellae appear to
run without interruption along the length of the
lamellae presumably to become contiguous with
scleral fibrils at the limbus. Normally occurring
fibril terminations are observed only in
association with extracellular compartments on
the processes of fibroblasts in which collagen
molecules are organized into fibril strands
during stromal remodeling.58
Fibrils in adjacent lamellae tend to be oriented
at highly oblique angles relative to one another.
The orientation of lamellae as a function of
depth has been well studied in the developing
chick cornea in which the orientation is nearly
orthogonal.59 There have been reports of nonrandom
orientations of lamellae in the human. At
midcentral stroma, lamellae tend to orient along
the vertical and horizontal axes.60 In the far
periphery evidence exists for circumferential
Page 33
annular orientation that is parallel to the
limbus.61 Because it is impractical to track
single bundles of fibrils for any great distance
in the stroma, these fibrils may actually be
organized with swirling arc-like orientations
near the limbus that collectively appear to be a
continuous belt of fibrils.62
Cross-sectional views of the central cornea give
a first impression of a highly ordered structure
with little interweaving among lamellae. Closer
examination of the histologic evidence, combined
with mechanical strength tests of lamellar
organization, indicates that a much more complex
network of interweaving and lamellar bifurcations
does exist.63 In particular, the anterior one-
third of the stroma appears more disorganized
than the posterior two-thirds when seen with
light microscopy. Similarly, the peripheral
regions of the stroma are more disorganized than
the central regions in the human, as shown by
interlamellar cohesive strength tests and
concomitant histology.63 Polarized light
micrographs of relatively large expanses of the
stroma in cross section reveal the extent of
interweaving and bifurcation more readily.53
Some collagen fibril bundles are not truly
Page 34
lamellar but are oriented at oblique angles to
the surface-parallel lamellae.53,63 These bundles
also appear to bifurcate frequently, with
portions becoming contiguous with surface-
parallel lamellae at various depths. A measure of
cohesive strength that structurally ties the
anterior stroma to more posterior regions of
stroma is imparted by these depth-varying fibril
bundles. These oblique bundles have not been
observed in rabbit stroma, which may account for
the greater stiffness and shearing strength seen
with isolated human stroma compared with the
rabbit.2 However, when swollen stroma is examined
in thick sections, fine collagen bundles are
found between the lamellar bundles in the rabbit,
but not in the human. In the corneas of the
elasmobranchs, suture fibers oriented
perpendicularly to the corneal surface are seen.
These observations of depth-varying collagen in
diverse species may indicate analogous structural
features that resist shearing forces, but they
are not homologous structures.
PROTEOGLYCANS
The hydrophilic mucopolysaccharide ground
substance in which collagen fibrils are embedded
takes the form of the proteoglycan, polypeptide
Page 35
protein cores to which glycosaminoglycans (GAGs)
are covalently bonded. GAGs are large
polysaccharide groups consisting of repeating
disaccharide units, with glucosamine or
galactosamine on the first monosaccharide and
galactose, glucuroninc acid, or iduronic acid on
the second monosaccharide. Proteoglycans can
assume a wide variety of forms depending on the
number and types of GAGs per molecule, as well as
the inclusion of additional side chains such as
oligosaccharides. Some of the GAGs found in the
stroma are keratan sulfate, dermatan sulfate,
chondroitin sulfate, chondroitin, and the
atypical, noncovalently bound hyaluronic acid. A
discussion about the complexity and subtle
species distinctions of corneal proteoglycans is
beyond the scope of this chapter, but the subject
has been reviewed elsewhere.64,65
The most abundant corneal stromal proteoglycans
are lumican with keratan sulfate GAG side chains
and decorin with chondroitin/dermatan sulfate GAG
side chains. In vitro analysis of fibril
formation indicates that both lumican and decorin
appear to have an inhibitory effect on collagen
fibrillogenesis because of the core proteins of
the PGs and not the GAG side chains.66
Page 36
Keratan sulfate and dermatan sulfate GAGs appear
to bind along the collagen fibrils at regularly
spaced sites and tend to be oriented
perpendicularly to the fibril. The less prevalent
chondroitin sulfate and hyaluronic acid are
localized within interfibrillar spaces without
evidence of binding to the fibril collagen.67 In
the developing cornea, proteoglycan-GAG complexes
vary in distribution and orientation with
time.67 The primary role of GAGs appears to be the
maintenance of interfibrillar spacing.
GAGs are heterogeneously dispersed throughout the
cornea. Castoro and co-workers found dermatan
sulfate to be more prevalent in the anterior
portion of the bovine stroma, and keratan sulfate
more prevalent in the posterior portion.68 This
distinction was based on differential water
content within the stroma. It was theorized that
more total water was found in the posterior
stroma because keratan sulfate readily absorbs
and releases water. Conversely, the anterior
stroma contained relatively less extractable
water, presumably because dermatan sulfate GAGs
bind to water molecules more tightly than do
keratan sulfate GAGs. However, Klyce and Russell
have shown that this anterior-posterior hydration
gradient can be predicted entirely by
Page 37
consideration of the transport and permeability
characteristics of the epithelium and
endothelium.69 Borcherding and associates63 found a
reduction of keratan sulfate at the corneal
limbus of the human stroma with a corresponding
rise in dermatan sulfate. Dermatan sulfate was
not found centrally but was a major GAG within
the sclera. Chondroitin was found centrally but
not peripherally, while chondroitin sulfate was
found only in the periphery and limbus.70
TRANSPARENCY
Recently, Müller and colleagues71 reported that
the distance between adjacent collagen fibrils is
20 nm on average, and given a fibril diameter of
23 nm, the interfibrillar spacing by her protocol
using fresh, unswollen tissue is only 43 nm on
average. They also found that the ring-like
structures of proteoglycans surrounding each
fibril (when viewed in cross section) have a mean
diameter of 45 nm, while the distance between
proteoglycans along the axial length of each
fibril is 42 nm. Each proteoglycan has an average
length of 54 nm based on her measurements and a
thickness of 11 nm.71
The small, regular diameter of the individual
collagen fibrils and their highly ordered spatial
Page 38
arrangement with interfibrillar spacing much
smaller than that of visible light (400 to 700
nm) has been implicated as the basis for corneal
transparency.72 In contrast, the sclera has
collagen fibrils that are not uniformly arranged
and have varying diameters, which results in an
opaque tissue.
Maurice72 recognized that because of the
difference in refractive index between the
stromal collagen fibrils and ground substance, up
to 94% of incident light on the cornea would be
scattered, making the cornea virtually opaque. He
proposed that collagen fibrils are in a lattice-
like arrangement with perfect dimensional order
and that each fibril scatters an individual
wavelet of light. By the process of destructive
wavefront interference, the light scatter from
individual fibrils would be cancelled by one
another, and the cornea would remain transparent
as long as the lattice arrangement was
maintained. This theory is an example of long-
range spatial order in which even fibrils
relatively distant from one another are precisely
spaced in accordance with the spatial dimensions
of the lattice.
Histologic views of the stromal fibrils in the
Page 39
normal cornea suggest that they possess a quasi-
regular spatial arrangement with short-range
interfibrillar order but not long-range
order.73 It is generally believed that short-range
order must be accounted for in any description of
corneal transparency and light scatter because of
the unique arrangement of the fibrils. Of the
short-range order models, that of Hart and
Farrell is among the most rigorous.73
In the case of physiologically induced light
scatter, Benedek74 noted the histologic appearance
of regions devoid of fibrils, which he called
"lakes." Light scatter could be generated by a
difference in the local refractive index of the
stroma, for example by a pocket of localized
edema, as compared to the surrounding tissue.
Lakes were suspected to be artifacts of
histologic processing, but Farrell and associates
have shown that these lakes theoretically could
induce light scatter equivalent to observed
empiric data.75 This supports the theory that
edematous light scatter is generated by lakes
with critical dimensions approaching one-half
wavelength of light and not merely by a random
disruption of fibrillar order. However, models
are subject to limitations, and edema may induce
a continuum of disruption, including lakes and
Page 40
short-range order disruption. Also, Bowman's
layer, which has randomly distributed fibrils, is
transparent. The mechanisms responsible for this
clarity appear to be the lack of scattering
elements spatially arranged with dimensions that
approach the critical distance of one-half
wavelength of light and the fact that the overall
thickness of the layer is relatively small
compared to the stroma.
KERATOCYTES
Occupying 3% to 5% of the total stromal volume,
keratocytes are interspersed throughout the
corneal stroma and form a communicating network
through their branching stellate processes. With
confocal microscopy, keratocytes appear to be
less densely dispersed than in Bowman's layer
(Fig. 15). In the deep stroma, keratocytes appear
spindle-shaped and more numerous than in the
midstroma (Fig. 16).
Fig. 15. Confocal microscopic transverse
image of the human mid-stroma in vivo.
The keratocyte nuclei appear less dense than in
Bowman's layer and less refractile. Note the
highly refractile nucleus in the upper part of
the image. This may be an activated keratocyte
Page 41
(500×). (Courtesy of Nidek Technologies.)
Fig. 16. Confocal microscopic transverse
image of the human deep-stroma in vivo.
Many of the keratocyte nuclei at this level are
spindle-shaped and appear dense in numbers
compared to the central cornea (500×). (Courtesy
of Nidek Technologies.)Tight junctions have been seen between tips of
processes, although usually a 20-nm intercellular
space is seen.21 Gap junctions are functional
based on microinjections of fluorescent dyes that
pass from cell to cell.76 Occasionally, portions
of keratocyte processes appear to lie within a
lamellar bundle; however, the majority of
keratocytes reside within the potential
interlamellar spaces. The stromal density of the
keratocytes demonstrates a linear loss as a
function of age, which tends to parallel the loss
of endothelial cells over time.77
The keratocyte is approximately 2 μm thick at the
cell body with a disproportionately large nucleus
compared to its cytoplasmic volume. Free
ribosomes, rough endoplasmic reticulum, and
Golgi's complexes are found in the cytoplasm.
Immunoelectronmicroscopic antigen labeling
specific for types III, V, and VI collagen was
Page 42
found localized to keratocytes; however, type III
labeling was weak.34,48 Keratocytes have a
remarkable presence of a network of fenestrations
on their cell walls that appear to be
functionally related to the diffusion and
mechanical attachment of collagen fibers to the
cell body.78
Keratocytes are normally spatially quiescent,
albeit constantly maintaining the extracellular
matrix. They appear to have a clockwise,
corkscrew-like appearance in terms of their
distribution with depth in the stroma. It is
speculated this has some function with respect to
communication in depth, while maintaining optimum
corneal transparency.78
However, keratocytes have a considerable degree
of mobility; when activated they are able to
migrate into wound margins rapidly to synthesize
new collagen and glycoproteins for tissue repair.
In vitro results indicate that a wide variety of
growth factors increase keratocyte chemotaxis.79 A
major difference between activated and quiescent
keratocytes is the organization of the
contractile cytoskeletal proteins. In the
quiescent form, contractile proteins appear to be
associated with f-actin for the maintenance of
Page 43
cell shape and interconnectivity. In the
activated form, a putative contractile apparatus
comprised of f-actin, myosin, and α-actinin is
organized into muscle-like stress fiber
bundles.80 Chondroitin sulfate and collagen types
V and VI were shown to have an inhibitory effect
on activated keratocyte migration, while
fibronectin increased migration significantly.
Integrins had a mediating effect on migration,
presumably affecting cell–matrix interactions.81
Keratocytes can vacate the anterior stroma
rapidly after de-epithelialization.82 The signal
for this response is unknown, but the observation
underscores the potential responsiveness of this
usually unremarkable cell. When keratocytes do
respond to a stromal insult, they are more
appropriately termed fibroblasts to reflect the
diminution in their stellate appearance and their
increased ability to generate procollagen for
subsequent fibril construction. There is strong
circumstantial evidence that keratocyte
activation is correlated to a higher level of
reflected light scatter from the stroma,
specifically from the cell bodies themselves, and
this may constitute much of the haze observed
after photorefractive keratectomy (PRK).83 It has
been well documented that tears are cytotoxic to
Page 44
keratocyte cells, and it has been shown that
apoptosis is reduced by surgical removal of the
extraorbital lacrimal gland, although some
compensatory cytotoxicity appears to be derived
from the intraorbital lacrimal gland in the mouse
model.84
Birk and Trelstad85 investigated the mechanisms
and regulation of fibrillogenesis in embryonic
chicken cornea. They found that extracellular
fibroblast cell surface compartmentalization was
responsible for the organization of the fibrils
into lamellar bundles. Furthermore, the
orthogonal organization of lamellae in the chick
was due to the orthogonality of the surface
compartments of the fibroblast processes.
Fibroblasts apparently orient themselves during
the deposition of the secondary stroma by taking
spatial cues existing in the primary stroma.
Thus, a single fibroblast cell is able to
dispense bundles of collagen fibrils along axes
that are oriented at or near 90 degrees to one
another; these bundles then form into lamellae.
Back to Top
DESCEMET'S MEMBRANE
Descemet's membrane can be thought of as the
basal lamina of the endothelium, and it varies in
Page 45
thickness in the human from approximately 3 μm at
birth to 8 to 10 μm in adulthood (Fig. 17).86 The
age-related growth and renewal of the membrane
after injury indicate that it is an extracellular
secretion of the endothelium. Descemet's membrane
is stratified into distinct layers according to
histologic appearance and immunohistochemical
labeling: a thin, unbanded zone immediately
adjacent to the interfacial matrix of the stroma
(approximately 0.3 μm); a banded anterior zone (2
to 4 μm); and an unbanded posterior zone that may
be as much as two-thirds the total thickness of
Descemet's membrane in adults (> 4 μm). The
banded and unbanded zone thicknesses vary,
depending on the age of development and the
species.66 Because the unbanded posterior zone is
laid down by the endothelium over the course of a
lifetime, it is a temporal record of the
physiologic state of the endothelium. For
example, unusual striated collagen banding occurs
in the posterior zone in association with corneal
disease such as Fuchs' dystrophy.87
Fig. 17. Full-thickness view of
Descemet's membrane. Stromal collagen
(C) with a keratocyte (F) is seen at
top. Endothelium (E) is seen at bottom.
Large box indicates anterior banded zone
Page 46
of 100-nm spaced collagen. Smaller boxes indicate
occasional foci in the amorphous posterior
unbanded zone. Arrows point to vesicles on the
endothelial basal membrane. (Courtesy of Drs.
Rodrigues, Waring, Hackett, and Donohoo.)As noted previously, Descemet's membrane does not
adhere strongly to the stroma, and it can be
surgically dissected as a sheet. The randomly
oriented collagen fibers in the interfacial
matrix of the stroma have a densely matted
appearance,40 and 22-nm thick fibers arising from
the matrix penetrate Descemet's membrane to a
depth of only 0.16 to 0.21 μm.88
The banded anterior zone is composed of numerous
precisely aligned sheets forming a three-
dimensional array or lattice.21 Each sheet of the
array is constructed from triangular elements
with densely staining nodes at the apices. The
separation between nodes is approximately 100 to
110 nm. The highly ordered arrangement of the
nodes causes the banded appearance seen in cross
section. In the posterior unbanded zone, the
appearance is of a homogeneous, fibrillogranular
region. The anterior unbanded zone also has a
fibrillogranular appearance but is extremely
difficult to discern visually (Fig. 18).86
Page 47
Fig. 18. Descemet's membrane is a
composite of a nonbanded posterior
portion with a granular appearance (DM)
and an anterior portion with a banded
appearance (arrows). At the interface between the
stromal collagen (above) and Descemet's membrane
is a narrow, unbanded, granular-appearing region.
The endothelium (E) is seen below (37,000×).
(Courtesy of Drs. Rodrigues, Waring, Hackett, and
Donohoo.)The composition of Descemet's membrane is
primarily collagen. Marshall and
colleagues34,48 found immunogold labeling for
collagen types III and IV in the posterior
unbanded zone, type IV in the anterior banded
zone, and types V and VI in the anterior unbanded
zone and at the interfacial matrix of the stroma.
In addition, long striated collagen V labeled
filaments were seen within Descemet's membrane
immediately adjacent to the endothelial cells.
These filaments may be the same as those
occasionally seen with specular microscopy2 and
by electron microscopy86 but do not appear to be
associated with the extensive striated
distribution found with disease-related states.
Tamura and associates89 also found collagen type
VIII strongly labeled in the anterior banded zone
using immunohistochemical localization for
Page 48
monoclonal antibodies.
Back to Top
ENDOTHELIUM
The human corneal endothelium is a single layer
of 400,000 to 500,000 cells. Confocal microscopy
provides views of this cell layer that surpass
the details seen under specular microscopy (Figs.
19 and 20). Cells are 4 to 6 μm in height and 20
μm in width, and their posterior surfaces are
predominantly hexagonal when viewed under
specular microscopy (Fig. 21). Cross-sectional
views with electron microscopy show that cell
lateral walls are extremely tortuous and
interdigitate with extensive folds and finger-
like projections. It has been estimated that the
total paracellular path length may be 10 times
longer than the total height of the
cell.90 Numerous gap junctions along the lateral
membranes provide cell-to-cell cytoplasmic
communication as evidenced by the presence of
connexin 43 and the spreading of fluorescent dye
from an injected cell to surrounding cells (Fig.
22).91
Fig. 19. Confocal microscopic transverse
image of the human corneal
endothelium in vivo. In the young normal cornea,
Page 49
the majority of the cells will have a hexagonal
outline and they will be fairly uniform in size.
The dark spots near the center in many of the
cells may represent the central endothelial
cilium (500×). (Courtesy of Nidek Technologies.)
Fig. 20. Confocal microscopic transverse
image of the human corneal endothelium
in vivo after penetrating keratoplasty. Note
cellular enlargement, polymegathism, and
polymorphism. Cell nuclei are clearly visible.
Normally non-dividing, endothelial cells enlarge
slowly with age to compensate for cell loss,
maintaining a continuous lining on Descemet's
membrane. Cataract surgery and transplants
generally exhibit cell loss. Below about 400
cells per mm2 endothelial decompensation can
occur with ensuing edema (500×). (Courtesy of
Nidek Technologies.)
Fig. 21. Scanning electron micrograph of
corneal endothelium. Note the regular
hexagonal arrangement of the cells
(1,170×). (Courtesy of Drs. Rodrigues,
Waring, Hackett, and Donohoo.)
Fig. 22. Scanning electron micrograph of
endothelial cell intracellular junctions
(8,100×). (Courtesy of Drs. Rodrigues,
Page 50
Waring, Hackett, and Donohoo.)The apical portion of the lateral cell membranes
facing the anterior chamber possesses small flap-
like or leaflet-like features that overlap
adjacent cells and harbor segmented tight
junctions that form an incomplete seal around the
apical cell margin (Fig. 23). Megamolecules
(greater than 80 kd) are essentially prevented
from penetrating the paracellular pathway, but
molecules such as horseradish peroxidase and
lanthanum are able to cross the apical
junctions.92,93While these tight junctions appear
dimensionally larger than those found in the
apical cells of the epithelium, they are not as
efficient.90 Nevertheless, these junctions,
combined with the closely apposed paracellular
pathway, prevent excessive passive transport of
anterior chamber fluid into the stroma. Any
normal leakage that may occur around the
endothelial cells tends to be counterbalanced by
the active ion transport processes in the cell
membranes. With pathologic cellular damage or
substantial reduction in cell density from a
normal value between 1,400 and 2,500 cells/mm2 to
a critical value of approximately 400 to 700
cells/mm2, the endothelial transport capability
becomes overwhelmed, and chronic stromal edema
can ensue.90 Endothelial cell density normally
Page 51
increases from the center to peripheral cornea by
approximately 10%, with the superior peripheral
quadrant increasing by nearly 16%.94
Fig. 23. Extensively interdigitated
lateral membranes of endothelial
cells. Near the apical margin, a
tight junction is formed (arrow). Bar = 0.5 ×m.
(Courtesy of Drs. Rodrigues, Waring, Hackett, and
Donohoo.)The normal central corneal endothelium has been
shown to have a reduction in the cell density
over a 10-year period. Repeated specular
microscopy in the same individuals over a 10-year
period with the same camera and processing method
indicates that cell density decreased from a mean
of 2,715 ± 301 to 2,539 ± 284 cells per square
millimeter (p < 0.0001). The percentage of
hexagonal cells dropped over the same time period
from 67% to 64% (p = 0.003), while the
coefficient of variation increased from 0.26 ±
0.05 to 0.29 ± 0.06 (p < 0.001).95
A single cilium often can be seen on the apical
endothelial membrane in a location geographically
associated with the intracellular centriole
pair.96 It is unclear whether this cilium is a
vestigial artifact or if it has an important
Page 52
regulatory or receptive function. Small numbers
of short microvilli also are seen on the apical
surface.
It has often been reported that the endothelial
cell monolayer does not proliferate in vivo.
However, endothelium does possess the ability to
proliferate, but this capacity is normally turned
off in the natural state due to the arrest of the
G1 phase of the cell cycle. The endothelial cells
appear unable to respond to autocrine or
paracrine stimulation even though they express
mRNA and the protein for growth factors and their
receptors. Importantly, cell-to-cell contact
appears to inhibit endothelial cell proliferation
during development and later maintains the
nonproliferative monolayer state of the adult
cornea. Ethylenediaminetetraacetic acid (EDTA)
releases cell-to-cell contact inhibition and
promotes proliferation in cell cultures of older
corneas when exposed to appropriate growth
factors.97–99
It was once believed that the basal cell surface
facing Descemet's membrane was irregular in
appearance due to the complexity suggested by the
lateral walls. In recent views provided by
scanning electron micrographs of stripped bovine
Page 53
and human endothelium, the cell mosaic facing
Descemet's membrane appeared hexagonal in a
manner similar to the mosaic facing the anterior
chamber, although the cell outlines appeared
broader and less distinct.100 The cell margins
adjacent to Descemet's membrane were ruffled,
with overlapping, flattened processes, some of
which appeared to insert into adjacent cell
membranes, while many were free and damaged
presumably during histologic preparation.
It has been shown that the glycosaminoglycans
keratan sulfate (KS) and chondroitin sulfate (CS)
are associated with the endothelial cell surface
in a variegated pattern (some cells have high
levels of KS and adjacent cells may have none,
while CS levels tend to show an inverse
relationship to KS levels) and likely play a role
in the ability of the cells to adhere and migrate
across the basal lamina.101
The intracellular contents of the endothelial
cell are dominated by a large nucleus and
numerous organelles, including mitochondria,
smooth and rough endoplasmic reticulum, and
Golgi's apparatus.21 The obvious metabolic
activity of these amitotic cells indicated by
their morphology is related to the energy
Page 54
requirement of the endothelial ion transport
system that plays a major role in the regulation
of corneal hydration.
INNERVATION
Sensory innervation of the cornea occurs
primarily through the ophthalmic branch of the
trigeminal nerve, which inserts into the
posterior globe as the long ciliary nerve fibers
and as a portion of the short ciliary nerves
through the ciliary ganglion. The limbal region
is supplied by these nerve fibers passing
anteriorly within the suprachoroidal space with
possible branching connections between the fibers
of the long and short ciliary fibers.90 These
fiber trunks begin branching near the ora serrata
region to form a circumferential plexus near the
corneoscleral junction. Branches from this plexus
travel anteriorly to innervate the adjacent
conjunctival and limbal epithelium.102 Some minor
innervation of the peripheral cornea also occurs
through superficial branches in the episcleral
and subconjunctival regions.
Primary branching from the plexus occurs as 60 to
70 nerve fibers radially enter the midstromal
cornea with approximately half containing 15 to
30 axons and the remainder having fewer than 15
Page 55
axons per fiber.103 Most fibers lose their myelin
sheaths 2 to 3 mm within the cornea; however, a
few branches may retain a Schwann cell covering
for some distance into the stroma. Loss of myelin
sheathing would aid in making the cornea
optically transparent. Considerable branching,
including recurrent branching, occurs among the
midstromal fibers. Relatively few nerve fibers
have been found to branch into the posterior
third of the stroma, and no innervation of the
endothelium or Descemet's membrane has been found
in humans.104
As the midstromal fibers travel toward the
central cornea, axons become finer, and beaded
features are seen along the filaments prior to
their termination. Collaterals from the
midstromal fibers branch anteriorly at 90 degrees
to create an extensive sub-Bowman's layer plexus.
Fibers from the plexus travel anteriorly through
Bowman's layer toward the epithelium, where they
again turn at 90 degrees and travel parallel to
the corneal surface just posterior to the
epithelium. Based on confocal microscopy, the
majority of fibers in the subbasal plexus of the
central cornea appear to be oriented along the
superior-inferior axis. There are approximately
5400 to 7200 nerve bundles in the subbasal
Page 56
plexus; because each bundle may contain several
axons, the total number of axonal fibers may be
as high as 44,000.105 Each fiber immediately forms
numerous, elaborate leashes with up to several
dozen beaded axon terminals per leash and some
additional unbeaded fibers. The beaded appearance
was at one time considered an artifact of tissue
processing or trauma; however, confocal
microscopy has shown these to be genuine features
(Fig. 24). The unbeaded straight terminals are
not seen entering the epithelium; however, the
beaded terminals of the leashes diverge
vertically and obliquely among the epithelial
cells in a complex manner before terminating at
the level of the apical cells (Fig. 25).102 The
diameter of the individual nerve fibers in the
subbasal plexus are between 0.05 and 2.5 μm with
most being in the range of 0.1 to 0.5 μm.
Fig. 24. Confocal microscopic transverse
image of the human basement epithelial
membrane in vivo demonstrates
unmyelinated beaded nerve leashes. These send off
nerve terminals within the corneal epithelium
(500×; courtesy of Nidek Technologies).
Page 57
Fig. 25. Nerve axons (N) are visible
in the basal cell layer of the
epithelium (49,500×). (Courtesy of
Drs. Rodrigues, Waring, Hackett, and
Donohoo.)Sympathetic innervation of the cornea occurs
through fibers with cell bodies in the superior
cervical ganglion.106,107 It has been demonstrated
in some species such as the rabbit by using
neuroanatomic mapping methods and observing the
functional changes in epithelial ion transport
induced by sympathectomy. Sympathetic innervation
is considered rare in all primates, including
humans. Parasympathetic innervation has not been
demonstrated in humans.
The corneal epithelium is one of the most highly
innervated structures in the body. Sensitivities
are 300 to 600 times that of skin, and a corneal
surface area of 0.01 mm2 may contain as many as
100 terminal endings.102 For the human cornea with
a surface area of approximately 120 mm2, there
may be as many as 1.2 million terminal endings.
Back to Top
CONCLUSION
The understanding of corneal anatomy is being
refined continuously. Studies of epithelial wound
Page 58
healing have led to a clearer picture of the
hemidesmosome-anchoring fibril complex.
Similarly, refractive surgery has generated the
need for more information on stromal collagen
structure and fibroblast activity. The use of
ophthalmic lasers and the continued efforts to
improve corneal storage media have stimulated
more research in endothelial cell morphology.
Clinical research has been, and will continue to
be, a significant motivator in the study of
corneal anatomy. It is likely that the
relationship between physiologic function and
anatomic diversity will become increasingly
important as attempts are made to alter the
cornea in very subtle ways to ultimately improve
clinical results.
Back to Top
ACKNOWLEDGMENT
This study was supported in part by National
Institutes of Health grants EY-014162 (M.K.S.),
EY-13311 (S.D.K.), and EY-02377 (L.S.U.) from the
National Eye Institute, Bethesda, Maryland.
Back to Top
REFERENCES
1. Tripathi RC, Tripathi BJ: Anatomy, orbit, and
adnexa of the human eye. In Davson H (ed): The
Page 59
Eye. 3rd ed. New York: Academic Press, 1984:1–268
2. Maurice DM: The cornea and sclera. In Davson H
(ed): The Eye. 3rd ed. New York, Academic Press,
1984: 1–158
3. Smolek MK, Klyce SD: Is Keratoconus a true
ectasia? An evaluation of corneal surface area.
Arch Ophthalmol 118:1179, 2000
4. Mishima S, Gassett A, Klyce SD: Determination
of tear volume and tear flow. Invest Ophthalmol
5:264, 1966
5. Rolando M, Refojo MF: Tear evaporimeter for
measuring water evaporation rate from the tear
film under controlled conditions in humans. Exp
Eye Res 36:25, 1983
6. Van Haeringen NJ: Clinical biochemistry of
tears. Surv Ophthalmol 26:84, 1981
7. Gipson IK, Yankauchas M, Spurr-Michaud SJ, et
al: Characteristics of a glycoprotein in the
ocular surface glycocalyx. Invest Ophthalmol Vis
Sci 33:218, 1992
8. Nichols BA, Chiappino ML, Dawson CB:
Demonstration of the mucous layer of the tear
film by electron microscopy. Invest Ophthalmol
Page 60
Vis Sci 26:464, 1985
9. Hazlett LD, Wells P, Spann B, et al:
Epithelial desquamation in the adult mouse
cornea: a correlative TEM-SEM study. Ophthalmic
Res 12:315, 1980
10. Wolosin JM: Regeneration of resistance and
ion transport in rabbit corneal epithelium after
induced surface cell exfoliation. J Membr Biol
104:45, 1988
11. Hanna C, Bicknell DS, O'Brien JE: Cell
turnover in the adult human eye. Arch Ophthalmol
65:695, 1961
12. Davenger M, Evenson A: Role of the
pericorneal papillary structure in renewal of
corneal epithelium. Nature 229:560, 1971
13. Alldredge OC, Krachmer JH: Clinical types of
corneal transplant rejection. Their
manifestations, frequency preoperative
correlates, and treatment. Arch Ophthalmol
99:599, 1981
14. Thoft RA, Friend J: The X, Y, Z hypothesis of
corneal epithelial maintenance. Invest Ophthalmol
Vis Sci 24:1442, 1983
15. Wiley L, SunderRaj N, Sun TT, et al: Regional
Page 61
heterogeneity in human corneal and limbal
epithelia: an immunohistochemical evaluation.
Invest Ophthalmol Vis Sci 32:594, 1991
16. Dua HS, Azuara-Blanco A: Limbal stem cells of
the corneal epithelium. Surv Ophthalmol 44:415,
2000
17. Wilson SE, Liu JJ, Mohan RR: Stromal-
epithelial interactions in the cornea. Prog Retin
Eye Res 18:293, 1999
18. Doughty MJ: Morphometric analysis of the
surface cells of rabbit corneal epithelium by
scanning electron microscopy. Am J Anat 189:316,
1990
19. Nichols B, Dawson CR, Tongi B: Surface
features of the conjunctiva and cornea. Invest
Ophthalmol Vis Sci 24:570, 1983
20. McLaughlin BJ, Cadwell RB, Sasaki Y, et al:
Freeze-fracture quantitative comparison of rabbit
corneal epithelial and endothelial membranes.
Curr Eye Res 4:951, 1985
21. Hogan MJ, Alvarado JA, Weddell E: Histology
of the Human Eye. Philadelphia: WB Saunders, 1971
22. Buschke W, Friedenwald JS, Fleischmann W:
Studies on the mitotic activity of the corneal
Page 62
epithelium: methods- The effects of colchicine,
ether, cocaine, and ephedrin. Bull Johns Hopkins
Hosp 73:143, 1943
23. Gipson IK, Spurr-Machaud S, Tisdale A, et al:
Redistribution of the hemidesmosome components
alpha 6 beta 4 integrin and bullous pemphigoid
antigens during epithelial wound healing. Exp
Cell Res 207:86, 1993
24. Shams NB, Hanninen LA, Chaves HV, et al:
Effect of vitamin A deficiency on the adhesion of
rat corneal epithelium and the basement membrane
complex. Invest Ophthalmol Vis Sci 34:2646, 1993
25. Kurpakus MA, Jones JC: A novel hemidesmosomal
plaque component tissue distribution and
incorporation into assembling hemidesmosomes in
an in vitro model. Exp Cell Res 194:139, 1991
26. Owaribe K, Nishizawa Y, Franke WW: Isolation
and characterization of hemidesmosomes from
bovine corneal epithelial cells. Exp Cell Res
192:622, 1991
27. Stepp MA, Spurr-Michaud S, Gipson IK:
Integrins in the wounded and unwounded stratified
squamous epithelium of the cornea. Invest
Ophthalmol Vis Sci 34:1829, 1993
Page 63
28. Kurpakus MA, Stock EL, Jones JCR: Analysis of
wound healing in an in vitro model: Early
appearance of laminin and a 125 ×
103 Mr polypeptide during adhesion complex
formation. J Cell Sci 96:651, 1990
29. Trinkhaus-Randall V, Tong M, Thomas P, et al:
Confocal imaging of the alpha 6 and beta 4
integrin subunits in the human cornea with aging.
Invest Ophthalmol Vis Sci 34:3103, 1993
30. Fine JD, Horiguchi Y, Jester J, et al:
Detection and partial characterization of a
midlamina lucida-hemidesmosome-associated antigen
(19-DEJ-1) present within human skin. J Invest
Dermatol 92:825, 1989
31. Gipson IK, Spurr-Michaud SJ, Tisdale AS:
Anchoring fibrils form a complex network in human
and rabbit cornea. Invest Ophthalmol Vis Sci
28:212, 1987
32. Keene DR, Sakai L Y, Lunstrum GP et al: Type
VII collagen forms an extended network of
anchoring fibrils. J Cell BioI 104:611, 1987
33. Burgeson RE: Type VII collagen. In Mayne R,
Burgeson R (eds): Structure and Function of
Collagen Types. New York: Academic Press,
Page 64
1987:145–172
34. Marshall GE, Konstas AG, Lee WR: Immunogold
fine structural localization of extracellular
matrix compounds in aged human cornea: I. Types
I–IV collagen and laminin. Graefes Arch Clin Exp
Ophthalmol 229:157, 1991
35. Konomi H, Hayashi T, Nakayasu K, et al:
Localization of type V collagen and type IV
collagen in human cornea, lung, and skin. Am J
PathoI 116:417, 1984
36. Newsome DA, Foidart JM, Hassell JH, et al:
Detection of specific collagen types for normal
and keratoconus corneas. Invest Ophthalmol Vis
Sci 20:738, 1981
37. Nakayasu K, Tanaka M, Konomi H, et al:
Distribution of types I, II, III, IV, and V
collagen in normal and keratoconus corneas.
Ophthalmic Res 18:1, 1986
38. Kolega J, Manabe M, Sun T-T: Basement
membrane heterogeneity and variation in corneal
epithelial differentiation. Differentiation
42:54, 1989
39. Cleutjens JP, Havenith MG, Kasper M, et al:
Absence of type IV collagen in the centre of the
Page 65
corneal epithelial basement membrane. Histochem J
22:688, 1990
40. Fujikawa LS, Foster CS, Gipson IK, et al:
Basement membrane components in healing rabbit
corneal epithelial wounds: Immunofluorescence and
ultrastructural studies. J Cell Biol 98:128, 1984
41. Scheinman JI, Tsai C: Monoclonal antibody to
type IV collagen with selective basement membrane
localization. Lab Invest 50:101, 1984
42. Linsenmayer TF, Fitch JM, Mayne R: Basement
membrane structure and assembly: Inferences from
immunological studies with monoclonal antibodies.
In Trelstad R (ed): The Role of Extracellular
Matrix in Development. New York: Alan R Liss,
1984:145–172
43. Fitch JM, Linsenmayer TF: Monoclonal antibody
analysis of ocular basement membranes during
development. Dev Biol 95:137, 1983
44. Yurchenco PD, Schittny JC: Molecular
architecture of basement membranes. FASEB J
4:1577, 1990
45. Cleutjens JP, Havenith MG, Vallinga M, et al:
Monoclonal antibodies to native basement
membranes reveal heterogeneous immunoreactivity
Page 66
patterns. Histochemistry 92:407, 1989
46. Komai Y, Ushiki T: The three dimensional
organization of collagen fibrils in the human
cornea and sclera. Invest Ophthalmol Vis Sci
32:2244, 1991
47. Hanna KD, Jouve FE, Waring GO: Preliminary
computer simulation of the effects of radial
keratotomy. Arch OphthalmoI 107:911, 1986
48. Marshall GE, Konstas AG, Lee WR: Immunogold
fine structural localization of extracellular
matrix compounds in aged human cornea: II.
Collagen types V–VI. Graefes Arch Clin Exp
Ophthalmol 229:164, 1991
49. Von der Mark K, Von der Mark H, Timpl R, et
al: Immunofluorescent localization of collagen
types I, II, and III in the embryonic chick eye.
Dev Biol 59:75, 1977
50. Birk DE, Fitch JM, Linsenmayer TF:
Organization of collagen types I and V in the
embryonic chicken cornea. Invest Ophthalmol Vis
Sci 27:1470, 1986
51. Fitch JM, Birk DE, Mentzer A, et al: Corneal
collagen fibrils: dissection with specific
collagenases and monoclonal antibodies. Invest
Page 67
Ophthalmol Vis Sci 29:1125, 1988
52. Linsenmayer TF, Bruns RR, Mentzer A, et al:
Type VI collagen: immunohistochemical
identification as a filamentous component of the
extracellular matrix of the developing avian
corneal stroma. Dev Biol 118:425, 1986
53. Tripathi RC, Bron AJ: Secondary anterior
crocodile shagreen of Vogt. Br J Ophthalmol
59:59, 1975
54. Linsenmayer TF, Gibney E, Igoe F, et al: Type
V collagen: molecular structure and fibrillar
organization of the chicken alpha 1 (V) NH2 -
terminal domain, a putative regulator of corneal
fibrillogenesis. J Cell Biol 121:1181, 1993
55. Yamamato S, Hashizume H, Hitome J, et al: The
subfibrillar arrangement of corneal and scleral
collagen fibrils as revealed by scanning electron
and atomic force microscopy. Arch Histol Cytol
63:127, 2000
56. Meller D, Peters K, Meller K: Human cornea
and sclera studied by atomic force microscopy.
Cell Tissue Res 288:111, 1977
57. Holmes DF, Gilpin CJ, Baldock C, et al:
Corneal collagen fibril structure in three-
Page 68
dimensions: structural insights in fibril
assembly, mechanical properties, and tissue
organization. Proc Natl Acad Sci USA 98:7307,
2001
58. Ploetz C, Zycband EI, Birk DE: Collagen
fibril assembly and deposition in the developing
dermis: segmental deposition in extracellular
compartments. J Struct Biol 106:73, 1991
59. Trelstad RL, Coulombre AJ: Morphogenesis of
the collagenous stroma in the chick cornea. J
Cell Biol 50:840, 1971
60. Meek KM, Blamires T, Elliott GF, et al: The
organisation of collagen fibrils in the human
corneal stroma: a synchrotron x-ray diffraction
study. Curr Eye Res 6:841, 1987
61. Meek KM, Newton RH: Organization of collagen
fibrils in the corneal stroma in relation to
mechanical properties and surgical practice. J
Refract Surg 15:695, 1999
62. Maurice DM: Mechanics of the cornea. In
Cavanagh HD (ed): The Cornea: Transactions of the
World Congress on the Cornea III. New York: Raven
Press, 1988:1987–1993
63. Smolek MK, McCarey BE: Interlamellar adhesive
Page 69
strength. Invest Ophthalmol Vis Sci 31:1087, 1990
64. Scott JE: Proteoglycan-fibrillar collagen
interactions. Biochem J 252:313, 1988
65. Scott JE, Bosworth TR: A comparative
biochemical and ultrastructural study of
proteoglycan-collagen interactions in corneal
stroma. Biochem J 270:491, 1990
66. Rada JA, Cornuet PK, Hassell JR: Regulation
of corneal collagen fibrillogenesis in vitro by
corneal proteoglycan (lumican and decorin) core
proteins. Exp Eye Res 56:635, 1993
67. Cintron C, Covington HI: Proteoglycan
distribution in developing rabbit cornea. J
Histochem Cytochem 38:675, 1990
68. Castoro JA, Bettelheim AA, Bettelheim FA:
Water concentration gradients across bovine
cornea. Invest Ophthalmol Vis Sci 29:963, 1988
69. Klyce SD, Russell SR: Numerical solution of
coupled transport equations applied to corneal
hydration dynamics. J Physiol 292:107, 1979
70. Borcherding MS, Blacik LJ, Sittig RA, et al:
Proteoglycans and collagen fibre organization in
human corneoscleral tissue. Exp Eye Res 21:59,
Page 70
1975
71. Muller LJ, Pels E, Schurmans LRHM, et al: A
new three-dimensional model of the organization
of proteoglycans and collagen fibrils in the
human corneal stroma. Exp Eye Res 78:493, 2004
72. Maurice DM: The structure and transparency of
the cornea. J Physiol 136:263, 1957
73. Hart RW, Farrell RA: Light scattering in the
cornea. J Opt Soc Am 59:766, 1969
74. Benedek GB: The theory of transparency of the
eye. Appl Optics 10:459, 1971
75. Farrell RA, McCally RL, Tatham PER:
Wavelength dependencies of light scattering in
normal and cold swollen rabbit corneas and their
structural implications. J Physiol 233:589, 1973
76. Watsky MA: Keratocyte gap junctional
communication in normal and wounded rabbit
corneas and human corneas. Invest Ophthalmol Vis
Sci 36:2568, 1995
77. Moller-Pedersen T: A comparative study of
human corneal keratocyte and endothelial cell
density during aging. Cornea 16:333, 1997
78. Muller LJ, Pels L, Vrensen GF: Novel aspects
Page 71
of the ultrastructural organization of human
corneal keratocytes. Invest Ophthal Vis Sci
36:2557, 1995
79. Andresen JL, Ehlers N: Chemotaxis of human
keratocytes is increased by platelet-derived
growth factor-BB, epidermal growth factor,
transforming growth factor-alpha, acidic
fibroblast growth factor, insulin-like growth
factor-I, and transforming growth factor-beta.
Curr Eye Res 17:79, 1998
80. Jester JV, Barry PA, Lind GJ, et al: Corneal
keratocytes: in situ and in vitro organization of
cytoskeletal contractile proteins. Invest
Ophthalmol Vis Sci 35:730. 1994
81. Andresen JL, Ledet T, Hager H, et al: The
influence of corneal stromal matrix proteins on
the migration of human corneal fibroblasts. Exp
Eye Res 71:33, 2000
82. Crosson CE: Cellular changes following
epithelial abrasion. In Beuerman RW, Crosson CE,
Kaufman HE (eds): Healing Processes in the
Cornea, Vol 1. Advances in Applied Biotechnology
Series. Houston, TX: Gulf Publishing, 1989: 3–14
83. Moller-Pedersen T: Keratocyte reflectivity
Page 72
and corneal haze. Exp Eye Res 78:553, 2004
84. Zhao J, Nagasaki T: Lacrimal gland as the
major source of mouse tear factors that are
cytotoxic to corneal keratocytes. Exp Eye Res
77:297, 2003
85. Birk DE, Trelstad RL: Extracellular
compartments in matrix morphogenesis: collagen
fibril, bundle, and lamellar formation by corneal
fibroblasts. J Cell Bioi 99:2024, 1984
86. Johnson DH, Bourne WM, Campbell RJ: The
ultrastructure of Descemet's membrane. I. Changes
with age in normal cornea. Arch Ophthalmol
100:1942, 1982
87. Bourne WM, Johnson DH, Campbell RJ: The
ultrastructure of Descemet's membrane. III.
Fuchs' dystrophy. Arch Ophthalmol 100:1952, 1982
88. Binder PS, Rock ME, Coalwell Schmidt K, et
al: High-voltage electron microscopy of normal
human cornea. Invest Ophthalmol Vis Sci 32:2234,
1991
89. Tamura Y, Konomi H, Sawada H, et al: Tissue
distribution of type VIII collagen in human adult
and fetal eyes. Invest Ophthalmol Vis Sci
Page 73
32:2636, 1991
90. Klyce SD, Beuerman RW: Structure and function
of the cornea. In Kaufman HE, Barron BA, McDonald
MB, Waltman SR (eds): The Cornea. New York:
Churchill Livingstone, 1988:3–54.
91. Williams K, Watsky M: Gap junctional
communication in the human corneal endothelium
and epithelium. Curr Eye Res 25:29, 2002
92. Waring GO, Bourne WM, Edelhauser HF, et al:
The corneal endothelium: Normal and pathological
structure and function. Ophthalmology 89:531,
1982
93. Kreutziger GO: Lateral membrane morphology
and gap junction structure in rabbit corneal
endothelium. Exp Eye Res 23:285, 1976
94. Amann J, Holley GP, Lee SB, et al: Increased
endothelial cell density in the paracentral and
peripheral regions of the human cornea. Am J
Ophthalmol 135:584, 2003
95. Bourne WM, Nelson LR, Hodge DO: Central
corneal endothelial cell changes over a ten-year
period. Invest Ophthalmol Vis Sci 38:779, 1997
96. Gallagher B: Primary cilia of the corneal
Page 74
endothelium. Am J Anat 159:475, 1980
97. Senoo T, Obara Y, Joyce N: EDTA. a promoter
of proliferation in human corneal endothelium.
Invest Ophthal Vis Sci 41:2930, 2000
98. Senoo T, Joyce NC: Cell cycle kinetics in
corneal endothelium from old and young donors.
Invest Ophthal Vis Sci 41:660, 2000
99. Joyce N: Proliferative capacity of the
corneal endothelium. Prog Retin Eye Res 22:359,
2003
100. Sherrard ES, Ng YL: The other side of the
corneal endothelium. Cornea 9:48, 1990
101. Rae JL, Watsky MA: Ionic channels in corneal
endothelium. Am J Physiology 270:975, 1996
102. Rozsa AJ, Beuerman RW: Density and
organization of free nerve endings in the corneal
epithelium of the rabbit. Pain 14:105, 1982
103. Zander E, Weddell G: Observations on the
innervation of the cornea. J Anat 85:68, 1951
104. Mawas MJ: L'innervation de la cornee
humaine. Bull Soc Ophtalmol Paris 2:162, 1951
105. Muller LJ, Vrensen GF, Pels L, et al:
Page 75
Architecture of human corneal nerves. Invest
Ophthalmol Vis Sci 38:985, 1997
106. Klyce SD, Beuerman RW, Crosson CE:
Alteration of corneal epithelial ion transport by
sympathectomy. Invest Ophthalmol Vis Sci 26:434,
1985
107. Klyce SD, Jenison GL, Crosson CE, et al:
Distribution of sympathetic nerves in the rabbit
cornea. ARVO Abstract. Invest Ophthalmol Vis Sci
27:354, 1986
Back to Top