Review Maize drought tolerance: Potential improvements through arbuscular mycorrhizal symbiosis? Christopher R. Boomsma, Tony J. Vyn * Department of Agronomy, Purdue University, Lilly Hall of Life Sciences, 915 West State Street, West Lafayette, IN 47907, United States Contents 1. Introduction ...................................................................................................... 15 2. Mycorrhizae and water relations ..................................................................................... 16 2.1. Physiological effects .......................................................................................... 16 2.2. Potential mechanisms ........................................................................................ 17 3. Arbuscular mycorrhizae and maize drought tolerance .................................................................... 17 3.1. Pre-flowering morpho-physiological responses and associated mechanisms ............................................. 17 3.2. Flowering and post-flowering morpho-physiological responses and associated mechanisms ................................ 21 3.3. In-field morpho-physiological responses.......................................................................... 23 4. Future research ................................................................................................... 24 4.1. Treatments, field research, and cultural practices .................................................................. 24 4.2. Secondary traits, conventional breeding, and QTL analysis ........................................................... 24 4.3. AM colonization ............................................................................................. 25 4.4. Molecular and genomic techniques .............................................................................. 26 5. Considerations .................................................................................................... 27 6. Conclusions ...................................................................................................... 28 Acknowledgements ................................................................................................ 28 References ....................................................................................................... 28 Field Crops Research 108 (2008) 14–31 ARTICLE INFO Article history: Received 7 August 2007 Received in revised form 27 December 2007 Accepted 10 March 2008 Keywords: Maize Arbuscular mycorrhizae Drought tolerance Water relations Anthesis-silking interval Leaf water potential ABSTRACT Due to long-term trends in global climate change and the expansion of maize production in drought- prone regions, the development of drought-tolerant maize varieties is of high importance, particularly for maize producers in developing nations where plant breeding improvements are more easily adopted than high-input agronomic practices. Prior advances in breeding for drought-tolerant maize have often involved selection for beneficial morpho-physiological traits such as a reduced anthesis-silking interval (ASI) and an improved harvest index (HI). Although maize is an effective host of arbuscular mycorrhizae (AM) in infertile and drought conditions, few maize breeding and physiological research programs have examined the potential of AM symbiosis to improve maize drought tolerance. The objective of this review is to highlight the morpho-physiological responses and potential mechanisms through which AM colonization might improve maize drought tolerance and, in doing so, to examine the potential of this symbiotic relationship to serve as an effective tool for improving cultivar drought tolerance. In this review, we briefly describe the general effects of AM symbiosis on plant water relations, and then extensively examine the effects of AM colonization on pre-flowering, flowering, and post-flowering maize morpho-physiology under drought-stress conditions in greenhouse and field settings. We then suggest potential areas for future research related to (a) the adoption of cropping practices promoting AM colonization and survival; (b) the further understanding of AM effects on maize morpho-physiology; and (c) the creation of AM-colonized, drought-tolerant maize cultivars through conventional breeding as well as molecular and genomic techniques. We conclude by discussing practical considerations associated with the use of AM for maize drought-stress research and large-scale, commercial maize production. ß 2008 Elsevier B.V. All rights reserved. * Corresponding author. Tel.: +1 765 496 3757; fax: +1 765 496 2926. E-mail address: [email protected](T.J. Vyn). Contents lists available at ScienceDirect Field Crops Research journal homepage: www.elsevier.com/locate/fcr 0378-4290/$ – see front matter ß 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.fcr.2008.03.002
Transcript
Field Crops Research 108 (2008) 14–31
Contents l is ts ava i lab le at ScienceDirec t
Field Crops Research
journa l homepage: www.e lsev ier .com/ locate / fcr
Review
Maize drought tolerance: Potential improvements through arbuscularmycorrhizal symbiosis?
Christopher R. Boomsma, Tony J. Vyn *
Department of Agronomy, Purdue University, Lilly Hall of Life Sciences, 915 West State Street, West Lafayette, IN 47907, United States
A R T I C L E I N F O
Article history:
Received 7 August 2007
Received in revised form 27 December 2007
Accepted 10 March 2008
Keywords:
Maize
Arbuscular mycorrhizae
Drought tolerance
Water relations
Anthesis-silking interval
Leaf water potential
A B S T R A C T
Due to long-term trends in global climate change and the expansion of maize production in drought-
prone regions, the development of drought-tolerant maize varieties is of high importance, particularly for
maize producers in developing nations where plant breeding improvements are more easily adopted
than high-input agronomic practices. Prior advances in breeding for drought-tolerant maize have often
involved selection for beneficial morpho-physiological traits such as a reduced anthesis-silking interval
(ASI) and an improved harvest index (HI). Although maize is an effective host of arbuscular mycorrhizae
(AM) in infertile and drought conditions, few maize breeding and physiological research programs have
examined the potential of AM symbiosis to improve maize drought tolerance. The objective of this review
is to highlight the morpho-physiological responses and potential mechanisms through which AM
colonization might improve maize drought tolerance and, in doing so, to examine the potential of this
symbiotic relationship to serve as an effective tool for improving cultivar drought tolerance. In this
review, we briefly describe the general effects of AM symbiosis on plant water relations, and then
extensively examine the effects of AM colonization on pre-flowering, flowering, and post-flowering
maize morpho-physiology under drought-stress conditions in greenhouse and field settings. We then
suggest potential areas for future research related to (a) the adoption of cropping practices promoting AM
colonization and survival; (b) the further understanding of AM effects on maize morpho-physiology; and
(c) the creation of AM-colonized, drought-tolerant maize cultivars through conventional breeding as well
as molecular and genomic techniques. We conclude by discussing practical considerations associated
with the use of AM for maize drought-stress research and large-scale, commercial maize production.
2 At the leaf level, WUE can be defined as the ratio of energy captured by
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 15
1. Introduction
Across the globe today, maize (Zea mays L.) is a direct staple foodfor millions of individuals and, through indirect consumption as afeed crop, is an essential component of global food security.Currently, the temperate cropping systems of North Americaaccount for over 40% of the world’s temperate maize production(Campos et al., 2004). Despite high maize production levels, duringmost growing seasons in North America, significant portions of themaize crop suffer from drought-induced yield losses (Jensen, 1995).Significant yield losses due to water stress are present in bothtemperate and tropical environments of other continents that alsoprovide maize for local and global consumption (Campos et al.,2004). Observed and predicted changes to the Earth’s atmosphere,climate, and biophysical system (e.g. higher greenhouse gasconcentrations, increased mean surface temperatures, greaterevapotranspiration, rainfall pattern redistributions) have thepotential to increase the occurrence of drought at locations aroundthe globe (IPCC, 2001, 2007). Long-term trends in global climatechange, along with the ongoing expansion of maize production intomarginal production regions, have the potential to generate agreater number of drought-prone maize production environmentsthat can lead to yield instability and, subsequently, food supplyinsecurity at local and even global levels (Edmeades et al., 1997a;IPCC, 2001, 2007; Hillel and Rosenzweig, 2002; Campos et al., 2004).
In order to stabilize and increase global maize production for aburgeoning world population, the development of maize varietieswith enhanced drought tolerance continues to be an importantobjective. Maize producers in drought-prone areas of developingnations can often adopt plant breeding improvements moreefficiently and effectively than high-input agronomic practiceswhich often depend on input availability, appropriate infrastruc-ture, market access, and requisite crop and soil management skills(Campos et al., 2004). Additionally, in many regions such as thelowland tropics, the use of drought-tolerant cultivars may be theonly economical option for many small-scale farmers (Bolanos andEdmeades, 1993a). In the United States and other industrializednations, improvements in hybrid drought tolerance could poten-tially benefit growers in numerous ways. These include lowerirrigation costs through improved water use efficiency (WUE) andenhanced productivity during drought episodes in non-irrigatedsystems. Since water availability is variable across fields andproducers typically grow only one hybrid in a particular field, amoderate amount of drought tolerance is necessary in all maizehybrids (Bruce et al., 2002). Thus, the development and adoption ofdrought-tolerant varieties is seen as a long-term solution to manyof the problems plaguing drought-prone maize production regionsaround the globe (Logrono and Lothrop, 1997).
Drought stress affects the morpho-physiology of maize at boththe cellular and whole-plant levels. Cellular-level responses towater stress1 include (a) abscisic acid (ABA) accumulation; (b)decreased cell expansion and division; (c) osmotic adjustment(although limited in maize relative to other species [Bolanos andEdmeades, 1991]); (d) proline accumulation; (e) chlorophyllphotooxidation; and (f) reduced enzyme activity. Whole-plant-level responses to drought stress include (a) reduced leaf, silk,stem, root, and grain kernel expansion; (b) stomatal closure; (c)decreased photosynthesis and respiration; (d) reduced assimilateflux to growing organs; (e) accelerated leaf senescence; (f) delayedsilk growth and greater ear and kernel abortion; (g) increased root/shoot ratio; and (h) stem reserve (i.e. photoassimilate) remobiliza-tion and subsequent lodging (Banziger et al., 2000). These and
1 The terms ‘drought stress’ and ‘water stress’ are used interchangeably in this
article.
other morpho-physiological responses to drought stress have beendetailed in multiple review and research articles (e.g. Schussler andWestgate, 1995; Zinselmeier et al., 1995; Gutierrez et al., 1997;Setter, 1997; Westgate, 1997; Banziger et al., 2000), so readers areencouraged to consult these works for further information.
The past development of drought-tolerant maize varieties inbreeding programs has often involved the identification of andselection for beneficial morpho-physiological traits such as areduced anthesis-silking interval (ASI) and improved WUE2
(Campos et al., 2004). However, despite maize being an effectivehost of arbuscular mycorrhizae (AM), few maize breeding andphysiological research programs have examined the morpho-physiological effects of AM infection on maize drought tolerance.Furthermore, to the best of the authors’ knowledge, no maizebreeding programs have selected for AM colonization and itsbeneficial effects when developing drought-tolerant varieties.
The symbiotic relationship between AM and the roots of higherplants contributes significantly to plant nutrition and growth (Auge,2001), and has been shown to increase the productivity of a varietyof agronomic crops including maize (Sylvia et al., 1993). Thesepositive responses in productivity to AM colonization have mainlybeen attributed to the enhanced uptake by AM of relativelyimmobile soil ions such as phosphorus (P), potassium (K), calcium(Ca), magnesium (Mg), sulfur (S), iron (Fe), zinc (Zn), copper (Cu), andmanganese (Mn) (Marschner and Dell, 1994; Marschner, 1995; Liuet al., 2000a,b; Liu et al., 2007), but also involve the enhanced uptakeand transport of far more mobile nitrogen (N) ions, particularlyunder drought conditions (Tobar et al., 1994a; Azcon et al., 1996; Liuet al., 2007). In maize and other species, the most widely recognizedcontribution of AM fungi to host-plant nutrition involves theirability to extract P from outside the P depletion zone around plantroots (Marschner, 1995; Miller, 2000; Liu et al., 2003; Smith et al.,2003; Liu et al., 2007). Enhanced plant growth might also result fromsoil-borne pathogen (e.g. nematodes, pathogenic fungi, bacteria)protection (Cardoso and Kuyper, 2006; St-Arnaud and Vujanovic,2007), improved soil structural development, aggregate stabiliza-tion (Bethlenfalvay and Schuepp, 1994; Miller and Jastrow, 2000;Picone, 2003; Rillig, 2004; Wright, 2005), and non-AM-host weedsuppression (Picone, 2003). Yet the positive effects of AM fungi onhost-plant growth and development are not only noticeable in lowsoil fertility conditions (Jeffries, 1987) but also in droughtenvironments (Sylvia et al., 1993; Picone, 2003; Liu et al., 2007).Since nutrient mobility is limited under drought conditions, AM mayhave a larger impact on overall plant growth and development in dryrelative to well-watered conditions (Sanchez-Dıaz and Honrubia,1994).
Although AM symbiosis may be of benefit to maize productivityin both infertile and drought environments, the metabolic cost ofimproved nutrient and water acquisition through AM symbiosis is asignificant component of the overall carbon economy of a host plant(Zhu et al., 2005). Varying by plant and fungal species, plant age, andAM developmental stage, AM can consume between 2 and 20% ofdaily host photosynthate production (Graham, 2000; Bryla andEissenstat, 2005). The respiratory cost of AM symbiosis can vary withenvironmental conditions such as soil fertility, temperature, andmoisture; light intensity; and elevated atmospheric CO2 (Bryla andEissenstat, 2005). In fertile agricultural soils, colonization by AM canactually reduce crop productivity, as carbon costs associated withAM colonization exceed carbon-production benefits derived from
photosynthesis per unit of water transpired. However, in crop production, it often
refers to the ratio of biomass accumulation (expressed as CO2 assimilation), total
biomass, or grain yield to water consumed (expressed as transpiration (T),
evapotranspiration (ET), or total water input to the system) (Sinclair et al., 1984).
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3116
the symbiotic relationship (Ryan and Graham, 2002; Zhu et al.,2005). The net benefit (excluding ancillary benefits such asimproved soil structure) of AM colonization therefore depends onthe advantages derived from the acquisition of limiting nutrientsand water versus the metabolic cost of maintaining the fungalsymbiont (Azcon-Aguilar and Bago, 1994; Zhu et al., 2005). In orderto enhance plant growth and productivity in drought-stressenvironments, AM symbiosis would, therefore, ideally (a) resultin a positive net carbon budget (i.e. AM-mediated increases in hostphotoassimilate production outweigh AM requirements for plant-derived carbohydrates) and (b) be minimized when water andnutrients are not limiting (thus reducing unnecessary carbohydrateconsumption).
While a small number of studies have reported no improve-ment in drought tolerance in maize (e.g. Hetrick et al., 1984;Hetrick et al., 1987; Simpson and Daft, 1990) and other species(Auge, 2001) as a result of AM colonization, numerous others havedemonstrated significant enhancements in various crop species(Auge, 2001). Past research in multiple plant species has shownthat many of these mechanisms involve traits associated withhost-plant water relations (Auge, 2001). Although the effects of AMsymbiosis on plant water relations have been studied extensivelyin a broad array of plants, relatively few of these studies havefocused on these mechanisms and their associated effects in maize,particularly under drought conditions. Some studies have found noeffect of AM symbiosis on maize water-relation parameters orgrowth responses during drought (e.g. Kothari et al., 1990;Simpson and Daft, 1990). Yet as extensively detailed by Auge(2001), numerous other studies in AM-colonized maize within thepast 20 years have found alterations in host-plant gas exchange,water relations, and general drought tolerance under water-limiting conditions (Table 1). However, this review extensivelyexamines only a number of these studies since (a) an exhaustiveexamination of all pertinent literature is beyond the scope of thisreview and (b) the remaining articles not discussed in detail herehave been sufficiently reviewed by Auge (2001) and are discussedin other recent investigations (i.e. Sylvia et al., 1993; Subramanianand Charest, 1995, 1997, 1998, 1999; Subramanian et al., 1995,1997; Schellenbaum et al., 1998; Amerian et al., 2001).
The objective of this review is to highlight the morpho-physiological responses and potential mechanisms through which
Table 1Summary of AM effects on host maize gas exchange, water relations, and drought tole
Fungus species Physiological parameter(s) aff
Effects potentially related to AM influences on greater plant size or improved P nutriti
AM colonization might improve maize drought tolerance, and, indoing so, to examine the potential of this symbiotic relationship toserve as an effective tool in improving cultivar drought tolerance.We begin by briefly describing the general effects of AM symbiosison plant water relations. Next we extensively examine the effectsof AM colonization on pre-flowering, flowering, and post-floweringmaize morpho-physiology under drought-stress conditions ingreenhouse and field settings. We then suggest potential areas forfuture research related to (a) the adoption of cropping practicespromoting AM colonization and survival; (b) the further under-standing of AM effects on maize morpho-physiology; and (c) thecreation of AM-colonized, drought-tolerant maize cultivarsthrough conventional breeding as well as molecular and genomictechniques. We conclude by discussing practical considerationsassociated with the use of AM for maize drought-stress researchand large-scale, commercial maize production.
2. Mycorrhizae and water relations
AM fungi have the ability to affect plant water relations in bothwater-limiting and well-watered conditions. Auge (2001) hasprovided an extensive review on the effects of AM symbiosis onplant water relations in numerous host species colonized byvarious fungal symbionts, with a particular emphasis on theseeffects under drought conditions. The following section is a briefsummary of the physiological effects of AM fungi on general plantwater relations under drought conditions, and the possiblemechanisms that underlie these effects. For further discussion ofthese mechanisms and other AM influences on host water relationsin a variety of plant species, refer to Auge (2001) and articles citedwithin.
2.1. Physiological effects
AM fungi often alter rates of water influx and efflux in hostplants, thus affecting tissue water content and leaf physiology. Oneprimary impact of AM symbiosis involves changes in stomatalconductance (gs) and transpiration (T), with T typically higher andgs frequently unaffected or greater during drought stress in AMrelative to non-AM plants. At times, AM fungi also postponereductions in leaf water potential (Cw) during periods of drought
rance under water-limiting conditions [adapted from Auge (2001)]
ected Reference
on
ssed plants Ramakrishnan et al. (1988a)
Ramakrishnan et al. (1988b)
] Muller and Hofner (1991)
elopment, >[P, Cu], Sylvia et al. (1993)
Acosta-Avalos et al. (1996)
mino acids] Subramanian and Charest (1995)
ergence of silks, <ASI Subramanian and Charest (1997)
ing enzymes, >[amino acids], Subramanian and Charest (1998)
er potential; GLA, green leaf area; SMC, soil moisture content.
3 Further information on the recurrent selection of Tuxpeno Sequıa can be found
in articles by Bolanos et al. (1993) and Bolanos and Edmeades (1996).
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 17
stress and hasten returns to control levels upon the alleviation ofwater-limiting conditions (Auge, 2001).
Measured reductions in soil moisture content (SMC) indicatethat root systems of AM plants often dry soils at a faster rate andmore thoroughly than root systems of non-AM plants. This effectlikely results from either the greater evaporative surface area(i.e. larger above-ground biomass) or more extensive rootsystems observed in AM relative to non-AM plants. However,it may also result from the adherence of AM hyphae to soilparticles, thus improving contact with the soil solution (Auge,2001). Enhanced drying by AM plants may also be associatedwith the access of hyphae to small pore spaces inaccessible tohost roots and root hairs (Sylvia et al., 1993; Ruiz-Lozano, 2003)and the subsequent uptake of water by AM mycelia for themaintenance of physiological activities during water stress(Sanchez-Dıaz and Honrubia, 1994). However, symbiosis doesnot typically result in improved root hydraulic conductivity(Auge, 2001).
AM symbiosis also influences plant metabolism, primarily byaffecting carbon allocation and metabolite concentrations in bothshoot and root tissues. In particular, AM colonization increasesfoliar concentrations of soluble carbohydrates and chlorophyll,with enhanced carbohydrate content the result of enhancedphotosynthetic capacity. Colonized plants often show eitherincreased or decreased accumulation of amino acids and nitro-genous compounds in addition to greater enzymatic activity (e.g.superoxide dismutase (SOD), see Ruiz-Lozano et al., 1996) underdrought-stress conditions (Auge, 2001).
2.2. Potential mechanisms
Through investigations of AM symbiosis in multiple plantspecies, a variety of mechanisms have been proposed to explainthe general effects of AM colonization on drought tolerance andoverall plant water relations (Auge, 2001). The most wellunderstood mechanism involves the effect of AM fungi on plantsize, with AM symbiosis generally leading to greater plantbiomass and altered within-plant size ratios (e.g. root–shootand root length–leaf area ratios). Early studies examining theeffects of AM symbiosis on plant water relations generallyconcluded that improved drought tolerance results from enhan-ced P nutrition (e.g. Nelsen and Safir, 1982). Yet as discussed byAuge (2001), further studies revealed the existence of othermechanisms either only partially correlated with, or unrelated to,plant nutrition or size (e.g. Auge et al., 1994). One proposedmechanism primarily focuses upon the impact of AM colonizationon water absorption rates, which further involves alterations ofplant gas exchange parameters and, subsequently, overall leafhydration. Other mechanisms involve changes in plant hydraulicconductance (e.g. enhanced stele tissue size), soil water relations(e.g. increased aggregate stability, greater soil available water),soil–root Cw gradients (e.g. enhanced soil drying), plant Cw
components (e.g. gs-leaf Cw relationship alterations), and non-hydraulic root signals (e.g. differing cytokinin and auxin (IAA)concentrations). Yet as suggested by Sanchez-Dıaz and Honrubia(1994), AM-induced changes in water relations may involvecomplex interactions among multiple mechanisms.
3. Arbuscular mycorrhizae and maize drought tolerance
Similar to possible responses and mechanisms discussed byAuge (2001), Subramanian et al. (1995, 1997) suggest that AMpotentially confer drought tolerance in maize through the complexinteraction of a variety of mechanisms. These include (a) improvedhydraulic conductivity; (b) direct AM water uptake; (c) altered
water relations (i.e. greater leaf Cw, gs and T); (d) enhanced rootingdepth and length; (e) improved plant nutrition; and (f) alteredplant metabolism. Since the early 1990s, a number of studies haveexamined one or more of the above mechanisms during the pre-flowering, flowering, and post-flowering developmental periodsusing maize cultivars inoculated with Glomus etunicatum (Sylviaet al., 1993), Glomus intraradices (Subramanian and Charest, 1995,1997, 1998, 1999; Subramanian et al., 1995, 1997; Amerian et al.,2001), and Glomus mosseae (Schellenbaum et al., 1998; Amerianet al., 2001). A majority of these studies (i.e. Subramanian andCharest, 1995, 1997, 1998, 1999; Subramanian et al., 1995, 1997;Schellenbaum et al., 1998) have involved the open-pollinated,tropical maize cultivar Tuxpeno Sequıa, which was selected foreight cycles of drought tolerance (C0, drought-sensitive–C8,drought-tolerant) by breeders at CIMMYT (Bolanos et al.,1993)3. In this section, we present a detailed summary of themorpho-physiological responses observed in these mostly pot-culture, greenhouse studies during the pre-flowering, flowering,and post-flowering periods and discuss how these responses arerelated to the mechanisms reviewed by Auge (2001) andSubramanian et al. (1995, 1997). Readers should note that dueto the relative paucity of studies examining drought responses inAM-colonized maize, a heavy emphasis is placed on the results ofthese particular pot-culture, greenhouse experiments (i.e. Sub-ramanian and Charest, 1995, 1997, 1998, 1999; Subramanian et al.,1995, 1997; Schellenbaum et al., 1998; Amerian et al., 2001).Although relatively few in number, these studies presently providethe most comprehensive view of AM effects on maize droughttolerance.
3.1. Pre-flowering morpho-physiological responses and associated
mechanisms
Water stress at any growth stage can reduce the grain yieldpotential of maize. In particular, drought stress during the pre-flowering period can reduce leaf area development, photosynth-esis, and ear and floret establishment (Banziger et al., 2000). Yetonly a handful of studies (Subramanian et al., 1997; Schellenbaumet al., 1998; Subramanian and Charest, 1998, 1999) have examinedpre-flowering drought responses in maize colonized by AM. Here,we summarize and discuss the findings of these studies.
In a pot-culture, greenhouse experiment, Subramanian et al.(1997) examined the pre-flowering drought tolerance of selectioncycle 0 and 8 plants of the cultivar Tuxpeno Sequıa inoculated withG. intraradices. AM colonization in plants from both selection cyclesenhanced leaf Cw and relative water content (RWC). Uponrewatering (66–86 days after sowing) after the removal ofirrigation for 3 weeks (45–65 days after sowing), both C0 and C8
AM plants took less time than non-AM plants to attain leaf Cw
values comparable to well-watered plants. Declines in maize leafCw in response to drought stress can increase membrane andprotein damage, enhance enzyme denaturation, and reduce CO2
assimilation (Foyer et al., 1998; Banziger et al., 2000). The higherCw and faster recovery rate of AM-infected plants during pre-flowering growth and development suggest AM significantly affectpre-flowering water relations in both drought-sensitive anddrought-tolerant plants of Tuxpeno Sequıa.
Greater pre-flowering P concentrations were also present inAM-colonized plants (Subramanian et al., 1997). Symbiosis withAM is consistently associated with a higher P status in colonizedhosts, which typically contributes to greater biomass production.Greater P status has been suggested to directly improve drought
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3118
tolerance through improved stomatal ABA4 sensitivity, greater gs,and increased T (Auge, 2001). Yet as detailed by Auge (2001), AMcan alter host water relations independently of enhanced Pacquisition. The mechanism(s) by which enhanced P uptakedirectly contribute(s) to improved drought tolerance in AM-colonized maize remains unknown.
Relative to non-AM plants of their respective selection cycles, C0
and C8 AM plants possessed greater root biomass under droughtconditions (Subramanian et al., 1997). Yet during eight cycles ofrecurrent selection (C0–C8) for drought tolerance in TuxpenoSequıa in the absence of AM, Bolanos et al. (1993) found an overallreduction in per-plant root biomass. Edmeades et al. (1999) andBanziger et al. (2000) therefore suggest that drought tolerance inmaize is not associated with total biomass production but instead agreater partitioning of biomass to the developing ear (i.e. improvedharvest index (HI)) and a better distribution of roots in the soilprofile rather than a greater root biomass per se.
In order to maintain an adequate Cw during periods of waterstress, maize roots typically extract water at greater soil depths(Sharp and Davies, 1985; Lorens et al., 1987; Ludlow and Muchow,1990; Kondo et al., 2000) by forsaking superficial root developmentin favor of root growth deep into the soil profile (Bolanos et al.,1993). Yet this rooting response to water stress is not universal(e.g. Prystupa and Lemcoff, 1998), and strongly depends upon theamount and timing of water availability to the crop (Bolanos et al.,1993)5. Enhanced superficial root development can therefore beimportant for sustaining maize growth and development duringshort periods of water stress (Dekankova et al., 2004). For example,substantial drying at greater soil depths later in the growing season(i.e. during late-vegetative and early-reproductive development)increases the importance of water uptake from upper portions ofthe soil profile following light rainfalls. A root system that is highlybranched at shallow soil depths while extending into deeper soilhorizons has the potential to ensure the rapid resumption of waterand nutrient uptake after a dry period (Sharp and Davies, 1985;Kuchenbuch et al., 2006) while simultaneously extracting waterfrom lower portions of the soil profile during prolonged periods ofwater stress. With mycorrhizal spore abundance, hyphal density,and root colonization shown to be greatest in the first 15-cm soillayer when maize is grown with reduced or conventional tillage(Kabir et al., 1998; Oehl et al., 2005), extensively branched, AM-colonized roots in the upper soil profile may significantlycontribute to water extraction from that soil horizon.
Colonization by AM often induces root morphological changesin host plants, with these responses varying by plant species andgenotype, fungal inoculum/symbiont, and nutrient and wateravailability (Berta et al., 1993; Atkinson et al., 1994; Yano et al.,1996; Mickelson and Kaeppler, 2005; Zhu et al., 2005)6. A relativelyfew number of studies suggest that AM colonization similarlyalters maize root morphology in both low and high P soils, withthese alterations primarily involving an increase in lateral rootgrowth during early host development (Kaldorf and Ludwig-Muller, 2000; Bressan and Vasconcellos, 2002; Paszkowski andBoller, 2002) or a modification of root shallowness (Zhu et al.,
4 The phytohormone ABA plays numerous roles in stimulating both cellular-level
and whole-plant physiological and metabolic responses to drought stress in maize.
These roles and responses are not covered in this review, primarily since they have
not been examined in water-stressed, AM-colonized maize. Readers are encouraged
to consult other articles for further information on ABA and drought stress (e.g.
Setter, 1997; Tuberosa et al., 2002; Giuliani et al., 2005; Zhang et al., 2006).5 Bolanos et al. (1993) suggest that if selections in their study been done under
frequent but light sprinkler irrigations, superficial rooting would likely have
increased at the expense of deeper rooting.6 For further discussion on the effects of AM symbiosis on root morphology, refer
to Atkinson et al. (1994).
2005). Unfortunately, none of these studies examined rootmorphological changes in drought conditions. Still, AM-mediatedalterations to root morphology, particularly in superficial soilhorizons, may improve maize drought tolerance and/or P acqui-sition (Zhu et al., 2005). More specifically, maize cultivars withenhanced lateral root proliferation and a high root/shoot ratio inresponse to AM could improve water uptake, inorganic N capture,and P acquisition after light rainfall events during droughtygrowing seasons (Edmeades et al., 1997b; Oikeh et al., 1999).However, given that maize roots are typically highly branched inthe surface soil layers, enhanced root branching resulting from AMcolonization may only be beneficial when deeper root growth isstrongly encouraged by early-season drought stress and/or when Pavailability is limited.
Drought stress inhibits both cell expansion and cell division,resulting in reduced leaf area development during the pre-flowering period (Banziger et al., 2000). Subramanian et al.(1997) observed AM-mediated increases in shoot biomass thatlikely involved improved pre-flowering leaf area (though thisparameter was not directly measured in this study). Potentiallygreater leaf area would result in increased photosyntheticcapacity per plant and, concomitantly, a greater accumulationof photoassimilates in above- and below-ground biomass(Subramanian et al., 1995). Subramanian et al. (1997) foundthat drought reduced sugar accumulation by 66% and 50% in theshoots of non-AM plants and by only 30% and 32% in the shootsof AM plants of C0 and C8 selection cycles, respectively. Greatersugar production may have resulted from not only potentiallyimproved leaf area, but also reduced chlorophyll photooxidation(Banziger et al., 2000), higher leaf Cw values, and, concomitantly,higher photosynthetic rates. Enhanced shoot accumulation ofphotoassimilates during the pre-flowering period in AM plantscould improve kernel set at flowering and increase the amount ofassimilates available for remobilization to developing kernels 2–3 weeks after pollination (reducing source limitations in thepost-flowering period). Higher sugar accumulation in roottissue could improve root exploration during periods of waterstress and provide sufficient carbohydrates for AM survival andfunctioning.
Subramanian et al. (1997) also report that SMC values werehigher in soils occupied by AM versus non-AM plants of bothselection cycles throughout the 3 weeks of water stress. Theycontend that this may have resulted from improved soilaggregation in the AM-treatment soils, which consequentlyincreased the soil’s water-holding capacity. These results contrastwith studies in maize (Auge et al., 1994) and other host species(Auge, 2001), which either found a negligible or negative effect ofAM fungi on SMC. Soil moisture levels during water stress aretypically lower in soils occupied by AM plants as a result of morethorough water acquisition and greater above-ground biomass(Auge, 2001). The higher SMC values recorded by Subramanianet al. (1997) are therefore intriguing and difficult to explainwithout further investigation.
During periods of drought, N availability is reduced resulting indecreased N uptake and lower rates of N assimilation (i.e. nitrate(NO3
�) reduction and ammonium (NH4+) assimilation). In separate
pot-culture, greenhouse studies using the previously describedselection cycle 0 maize cultivar inoculated with G. intraradices,Subramanian and Charest (1998, 1999) examined the effects ofpre-flowering water stress and AM symbiosis on N acquisition,assimilation, and metabolism. The cultivar from selection cycle 8was also used in the 1998 study. The irrigation regimes for the1998 study were identical to those of Subramanian et al. (1997);however, the 1999 study involved the imposition of water stressfor 2 weeks at 56 days after sowing.
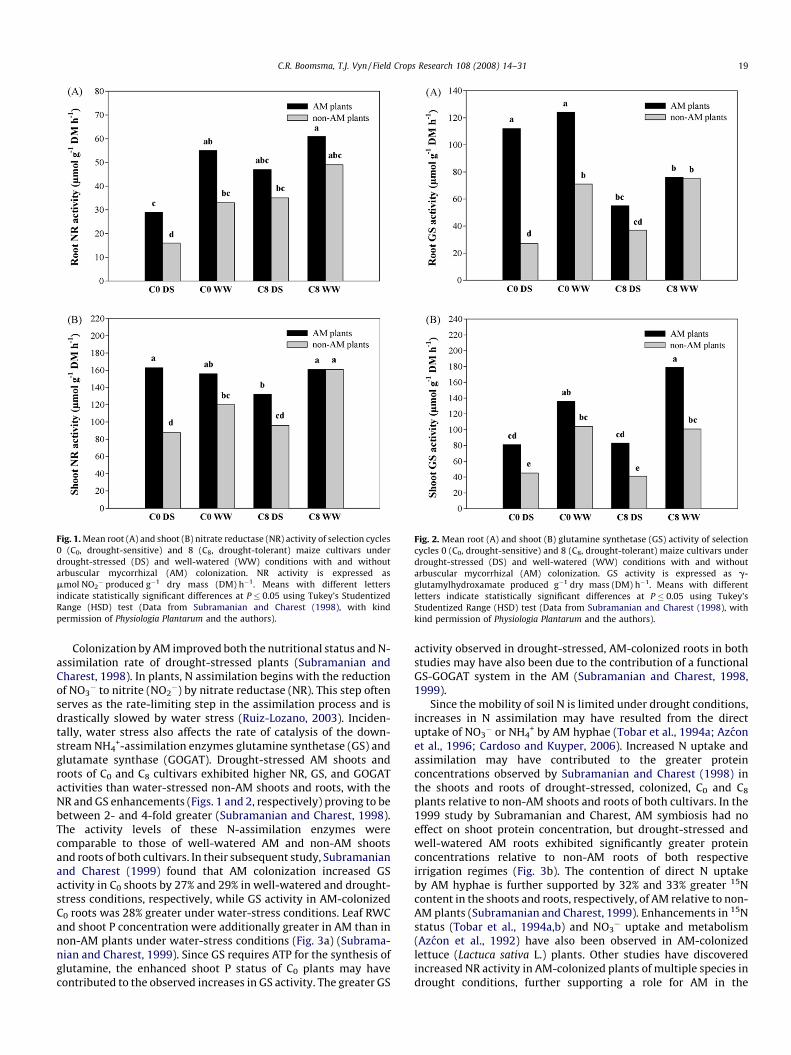
Fig. 1. Mean root (A) and shoot (B) nitrate reductase (NR) activity of selection cycles
0 (C0, drought-sensitive) and 8 (C8, drought-tolerant) maize cultivars under
drought-stressed (DS) and well-watered (WW) conditions with and without
arbuscular mycorrhizal (AM) colonization. NR activity is expressed as
mmol NO2� produced g�1 dry mass (DM) h�1. Means with different letters
indicate statistically significant differences at P � 0.05 using Tukey’s Studentized
Range (HSD) test (Data from Subramanian and Charest (1998), with kind
permission of Physiologia Plantarum and the authors).
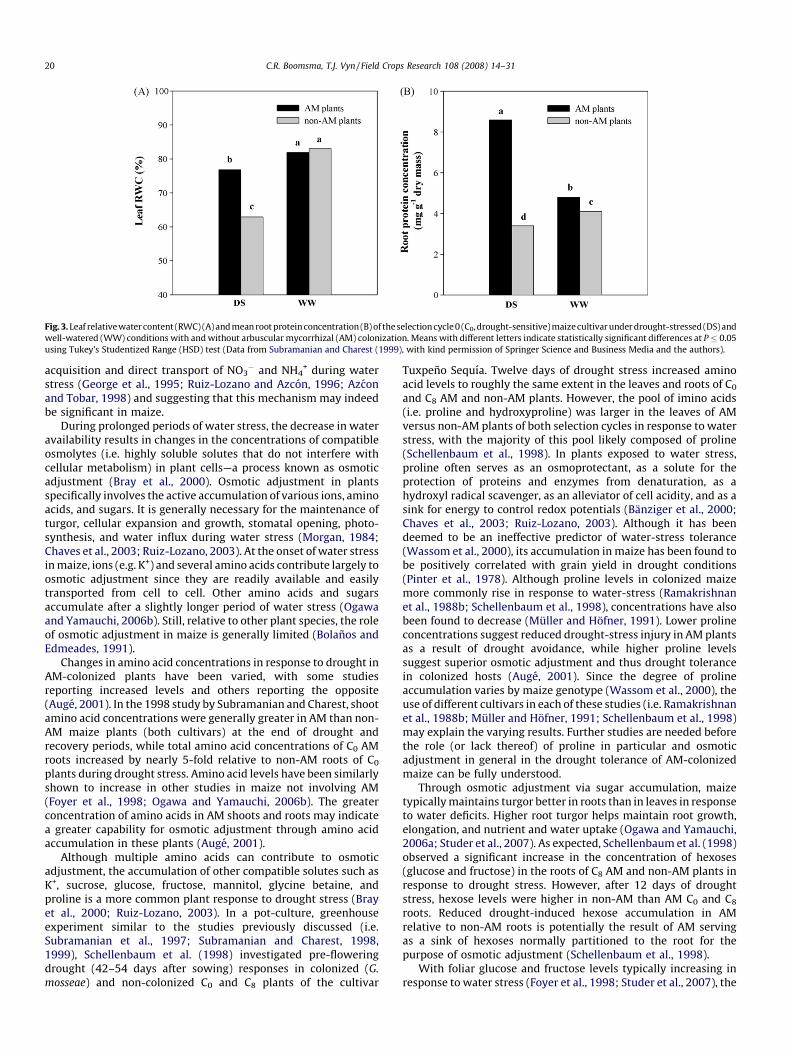
Fig. 2. Mean root (A) and shoot (B) glutamine synthetase (GS) activity of selection
cycles 0 (C0, drought-sensitive) and 8 (C8, drought-tolerant) maize cultivars under
drought-stressed (DS) and well-watered (WW) conditions with and without
arbuscular mycorrhizal (AM) colonization. GS activity is expressed as g-
glutamylhydroxamate produced g�1 dry mass (DM) h�1. Means with different
letters indicate statistically significant differences at P � 0.05 using Tukey’s
Studentized Range (HSD) test (Data from Subramanian and Charest (1998), with
kind permission of Physiologia Plantarum and the authors).
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 19
Colonization by AM improved both the nutritional status and N-assimilation rate of drought-stressed plants (Subramanian andCharest, 1998). In plants, N assimilation begins with the reductionof NO3
� to nitrite (NO2�) by nitrate reductase (NR). This step often
serves as the rate-limiting step in the assimilation process and isdrastically slowed by water stress (Ruiz-Lozano, 2003). Inciden-tally, water stress also affects the rate of catalysis of the down-stream NH4
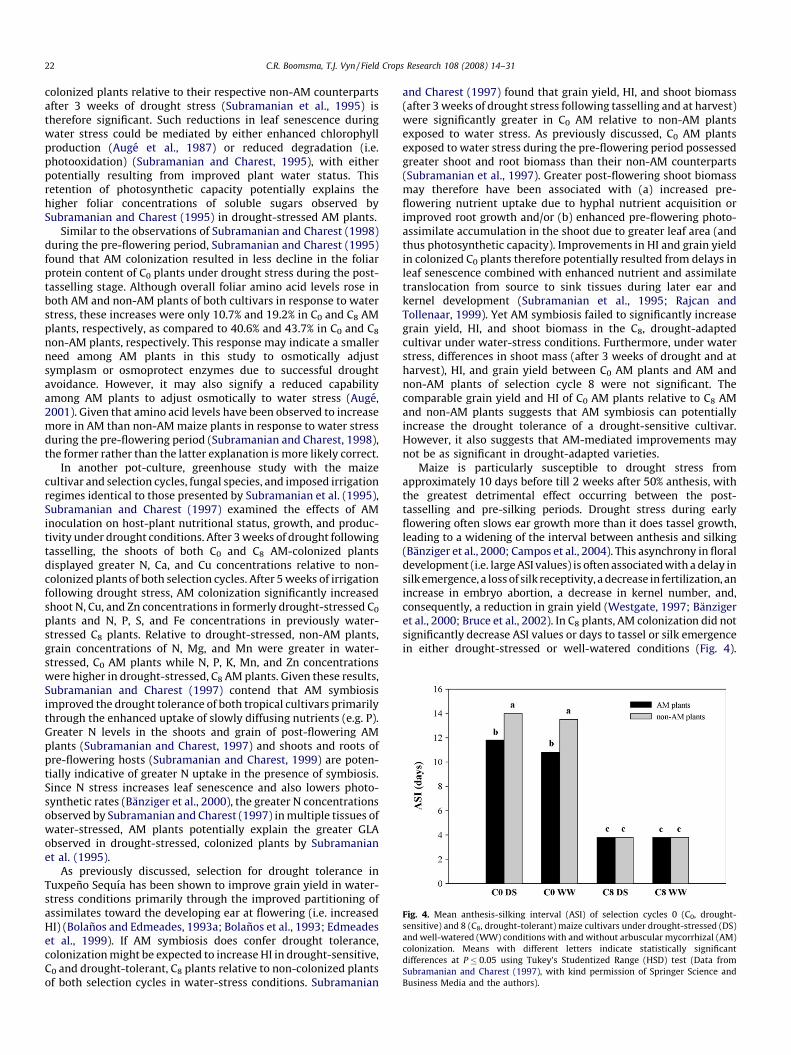
+-assimilation enzymes glutamine synthetase (GS) andglutamate synthase (GOGAT). Drought-stressed AM shoots androots of C0 and C8 cultivars exhibited higher NR, GS, and GOGATactivities than water-stressed non-AM shoots and roots, with theNR and GS enhancements (Figs. 1 and 2, respectively) proving to bebetween 2- and 4-fold greater (Subramanian and Charest, 1998).The activity levels of these N-assimilation enzymes werecomparable to those of well-watered AM and non-AM shootsand roots of both cultivars. In their subsequent study, Subramanianand Charest (1999) found that AM colonization increased GSactivity in C0 shoots by 27% and 29% in well-watered and drought-stress conditions, respectively, while GS activity in AM-colonizedC0 roots was 28% greater under water-stress conditions. Leaf RWCand shoot P concentration were additionally greater in AM than innon-AM plants under water-stress conditions (Fig. 3a) (Subrama-nian and Charest, 1999). Since GS requires ATP for the synthesis ofglutamine, the enhanced shoot P status of C0 plants may havecontributed to the observed increases in GS activity. The greater GS
activity observed in drought-stressed, AM-colonized roots in bothstudies may have also been due to the contribution of a functionalGS-GOGAT system in the AM (Subramanian and Charest, 1998,1999).
Since the mobility of soil N is limited under drought conditions,increases in N assimilation may have resulted from the directuptake of NO3
� or NH4+ by AM hyphae (Tobar et al., 1994a; Azcon
et al., 1996; Cardoso and Kuyper, 2006). Increased N uptake andassimilation may have contributed to the greater proteinconcentrations observed by Subramanian and Charest (1998) inthe shoots and roots of drought-stressed, colonized, C0 and C8
plants relative to non-AM shoots and roots of both cultivars. In the1999 study by Subramanian and Charest, AM symbiosis had noeffect on shoot protein concentration, but drought-stressed andwell-watered AM roots exhibited significantly greater proteinconcentrations relative to non-AM roots of both respectiveirrigation regimes (Fig. 3b). The contention of direct N uptakeby AM hyphae is further supported by 32% and 33% greater 15Ncontent in the shoots and roots, respectively, of AM relative to non-AM plants (Subramanian and Charest, 1999). Enhancements in 15Nstatus (Tobar et al., 1994a,b) and NO3
� uptake and metabolism(Azcon et al., 1992) have also been observed in AM-colonizedlettuce (Lactuca sativa L.) plants. Other studies have discoveredincreased NR activity in AM-colonized plants of multiple species indrought conditions, further supporting a role for AM in the
Fig. 3. Leaf relative water content (RWC) (A) and mean root protein concentration (B) of the selection cycle 0 (C0, drought-sensitive) maizecultivar under drought-stressed (DS) and
well-watered (WW) conditions with and without arbuscular mycorrhizal (AM) colonization. Means with different letters indicate statistically significant differences at P� 0.05
using Tukey’s Studentized Range (HSD) test (Data from Subramanian and Charest (1999), with kind permission of Springer Science and Business Media and the authors).
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3120
acquisition and direct transport of NO3� and NH4
+ during waterstress (George et al., 1995; Ruiz-Lozano and Azcon, 1996; Azconand Tobar, 1998) and suggesting that this mechanism may indeedbe significant in maize.
During prolonged periods of water stress, the decrease in wateravailability results in changes in the concentrations of compatibleosmolytes (i.e. highly soluble solutes that do not interfere withcellular metabolism) in plant cells—a process known as osmoticadjustment (Bray et al., 2000). Osmotic adjustment in plantsspecifically involves the active accumulation of various ions, aminoacids, and sugars. It is generally necessary for the maintenance ofturgor, cellular expansion and growth, stomatal opening, photo-synthesis, and water influx during water stress (Morgan, 1984;Chaves et al., 2003; Ruiz-Lozano, 2003). At the onset of water stressin maize, ions (e.g. K+) and several amino acids contribute largely toosmotic adjustment since they are readily available and easilytransported from cell to cell. Other amino acids and sugarsaccumulate after a slightly longer period of water stress (Ogawaand Yamauchi, 2006b). Still, relative to other plant species, the roleof osmotic adjustment in maize is generally limited (Bolanos andEdmeades, 1991).
Changes in amino acid concentrations in response to drought inAM-colonized plants have been varied, with some studiesreporting increased levels and others reporting the opposite(Auge, 2001). In the 1998 study by Subramanian and Charest, shootamino acid concentrations were generally greater in AM than non-AM maize plants (both cultivars) at the end of drought andrecovery periods, while total amino acid concentrations of C0 AMroots increased by nearly 5-fold relative to non-AM roots of C0
plants during drought stress. Amino acid levels have been similarlyshown to increase in other studies in maize not involving AM(Foyer et al., 1998; Ogawa and Yamauchi, 2006b). The greaterconcentration of amino acids in AM shoots and roots may indicatea greater capability for osmotic adjustment through amino acidaccumulation in these plants (Auge, 2001).
Although multiple amino acids can contribute to osmoticadjustment, the accumulation of other compatible solutes such asK+, sucrose, glucose, fructose, mannitol, glycine betaine, andproline is a more common plant response to drought stress (Brayet al., 2000; Ruiz-Lozano, 2003). In a pot-culture, greenhouseexperiment similar to the studies previously discussed (i.e.Subramanian et al., 1997; Subramanian and Charest, 1998,1999), Schellenbaum et al. (1998) investigated pre-floweringdrought (42–54 days after sowing) responses in colonized (G.
mosseae) and non-colonized C0 and C8 plants of the cultivar
Tuxpeno Sequıa. Twelve days of drought stress increased aminoacid levels to roughly the same extent in the leaves and roots of C0
and C8 AM and non-AM plants. However, the pool of imino acids(i.e. proline and hydroxyproline) was larger in the leaves of AMversus non-AM plants of both selection cycles in response to waterstress, with the majority of this pool likely composed of proline(Schellenbaum et al., 1998). In plants exposed to water stress,proline often serves as an osmoprotectant, as a solute for theprotection of proteins and enzymes from denaturation, as ahydroxyl radical scavenger, as an alleviator of cell acidity, and as asink for energy to control redox potentials (Banziger et al., 2000;Chaves et al., 2003; Ruiz-Lozano, 2003). Although it has beendeemed to be an ineffective predictor of water-stress tolerance(Wassom et al., 2000), its accumulation in maize has been found tobe positively correlated with grain yield in drought conditions(Pinter et al., 1978). Although proline levels in colonized maizemore commonly rise in response to water-stress (Ramakrishnanet al., 1988b; Schellenbaum et al., 1998), concentrations have alsobeen found to decrease (Muller and Hofner, 1991). Lower prolineconcentrations suggest reduced drought-stress injury in AM plantsas a result of drought avoidance, while higher proline levelssuggest superior osmotic adjustment and thus drought tolerancein colonized hosts (Auge, 2001). Since the degree of prolineaccumulation varies by maize genotype (Wassom et al., 2000), theuse of different cultivars in each of these studies (i.e. Ramakrishnanet al., 1988b; Muller and Hofner, 1991; Schellenbaum et al., 1998)may explain the varying results. Further studies are needed beforethe role (or lack thereof) of proline in particular and osmoticadjustment in general in the drought tolerance of AM-colonizedmaize can be fully understood.
Through osmotic adjustment via sugar accumulation, maizetypically maintains turgor better in roots than in leaves in responseto water deficits. Higher root turgor helps maintain root growth,elongation, and nutrient and water uptake (Ogawa and Yamauchi,2006a; Studer et al., 2007). As expected, Schellenbaum et al. (1998)observed a significant increase in the concentration of hexoses(glucose and fructose) in the roots of C8 AM and non-AM plants inresponse to drought stress. However, after 12 days of droughtstress, hexose levels were higher in non-AM than AM C0 and C8
roots. Reduced drought-induced hexose accumulation in AMrelative to non-AM roots is potentially the result of AM servingas a sink of hexoses normally partitioned to the root for thepurpose of osmotic adjustment (Schellenbaum et al., 1998).
With foliar glucose and fructose levels typically increasing inresponse to water stress (Foyer et al., 1998; Studer et al., 2007), the
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 21
contribution of sugars to foliar osmotic adjustment can besubstantial in maize (Ogawa and Yamauchi, 2006b). While foliarhexose content significantly increased in response to droughtstress in C8 non-AM plants, it decreased slightly in C8 AM plants.This phenomenon might be explained by AM and the roots whichthey colonize acting as stronger sink of photosynthates than leavesduring drought stress rather than the inability of AM plants to usesugars for foliar osmotic adjustment.
3.2. Flowering and post-flowering morpho-physiological responses
and associated mechanisms
Although drought affects maize grain yield potential during thepre-flowering period, the crop is most susceptible to water stressduring and shortly after flowering (Banziger et al., 2000). As withthe pre-flowering period, AM effects on maize drought toleranceduring the flowering and post-flowering periods have been limitedto a relatively small number of studies (Subramanian et al., 1995;Subramanian and Charest, 1995, 1997; Amerian et al., 2001). In thissection, we elaborate in some detail on the findings of this limitednumber of investigations.
Subramanian et al. (1995) and Subramanian and Charest (1995)examined flowering and post-flowering responses to droughtstress in a pot-culture, greenhouse setting using G. intraradices-colonized plants from selection cycles 0 and 8 of the cultivarTuxpeno Sequıa. In both studies, irrigation was withheld for 3weeks after tasselling (75–95 days after sowing) and then resumedfor 5 weeks until harvest.
After 1 week of drought stress, AM plants of both selectioncycles exhibited leaf Cw values significantly greater than non-AMplants. However, during the 2nd and 3rd weeks, improvements inleaf Cw associated with AM symbiosis were less pronounced in thedrought-resistant relative to drought-sensitive cultivar (Subrama-nian et al., 1995). Similar trends were observed by Subramanianet al. (1997) when drought stress was imposed during the pre-flowering period, although pre-flowering leaf Cw values weregenerally lower than post-flowering values. Lower pre-floweringCw values may have resulted from enhanced rates of T or reducedwater acquisition during the pre-flowering period or may havebeen associated with differing physiological responses (e.g.stomatal closure) to water stress during each period.
After 8 cycles (C0–C8) of recurrent selection for droughttolerance in Tuxpeno Sequıa in the absence of AM, Bolanos et al.(1993) found no difference in leaf Cw between the drought-sensitive C0 and drought-tolerant C8 cultivars when measuringplants in the field. This suggests that the greater Cw observed bySubramanian et al. (1995, 1997) in AM plants may not beassociated with enhanced drought tolerance. However, it should benoted that Bolanos et al. (1993) measured plants in the field whileSubramanian et al. (1995, 1997) measured maize in a greenhousesetting. Other studies examining cultivar differences have foundincreased HI, biomass, and grain yield to be correlated with greaterleaf Cw in drought conditions (e.g. Bennett et al., 1986; Lorenset al., 1987). Given that leaf Cw can be used as an indirect measureof a maize plant’s ability to explore the soil volume (Bolanos et al.,1993), the higher leaf Cw of AM plants may have resulted fromenhanced proliferation of roots or access by AM hyphae tootherwise unavailable water in drought conditions (Kothari et al.,1990), thus conferring some degree of improved drought tolerance.
In addition to higher leaf Cw values, both C0 and C8 AM-colonized plants possessed higher gs values7 under drought-stress
7 Subramanian et al. (1995) report values for stomatal resistance (rs) instead of gs.
However, since gs = 1/rs and gs appears to be more commonly used in the literature
(e.g. Auge, 2001), we have chosen to interpret these results using the term gs.
conditions, indicating that the plants were capable of keepingstomata open for longer periods than non-AM plants (Subrama-nian et al., 1995) and, as a result, could maintain lower leaftemperatures (Gutierrez et al., 1997). Because of greater leaf Cw
and gs values, AM plants typically exhibit greater photosyntheticrates than non-AM plants (Auge, 2001). Enhancements inphotosynthesis may explain the greater concentrations of totaland reducing sugars observed in AM relative to non-AM maizefoliar growth of both selection cycles by Subramanian and Charest(1995) at the end of 3 weeks of drought. Although greater sugarcontent in drought-stressed, AM leaves contrasts with the findingsof Schellenbaum et al. (1998) previously discussed, differencesmay have resulted from G. mosseae acting as a stronger sink foravailable sugars in the pre-flowering study by Schellenbaum et al.(1998) combined with more extensive colonization by G. mosseae
than G. intraradices (Amerian et al., 2001).Along with greater leaf Cw and gs values, AM plants possessed
higher T rates in drought-stress conditions, with T rates 29% and19% greater in C0 and C8 AM plants, respectively, relative to non-AM plants (Subramanian et al., 1995). The drought-resistant, AM-colonized cultivar maintained significantly greater T rates relativeto the drought-sensitive, AM-colonized cultivar throughout thedrought episode. However, examining physiological differencesbetween C0 and C8 plants of Tuxpeno Sequıa in the absence of AM,Pena-Valdivia et al. (1997) discovered that the mean T rate was 30%lower in C8 plants. Given the higher grain yield and HI (Bolanos andEdmeades, 1993a; Bolanos et al., 1993) and reduced T (Pena-Valdivia et al., 1997) of C8 plants, it appears that eight cycles of full-sib recurrent selection in Tuxpeno Sequıa have improved the WUEof the cultivar (Pena-Valdivia et al., 1997). Any improvements inWUE conferred through selection are likely to be similarly presentin AM-colonized C8 plants, and thus observed increases in leaf Cw,gs, and T resulting from symbiosis likely resulted from greaterutilization of available water.
After 3 weeks of drought stress following tasselling, selectioncycle 0 and 8 AM plants exhibited greater green leaf area (GLA)than their non-AM counterparts. Drought-sensitive and drought-tolerant AM-colonized cultivars retained 27.5% and 2.5% greaterGLA, respectively, relative to non-AM plants, and non-AM C8 plantsretained significantly more GLA relative to non-colonized C0 plants(Subramanian et al., 1995). The maintenance of GLA is importantduring the flowering and post-flowering periods, since currentphotosynthesis rather than the remobilization of photoassimilatescreated during the pre-flowering period generally support silk andearly ear growth and development (Westgate, 1997; Banzigeret al., 2000). When examining differences in drought toleranceamong C0 and C8 plants of Tuxpeno Sequıa in the absence of AM,Bolanos et al. (1993) found no difference in GLA retention betweenthe two selection cycles. Although this suggests that the improveddrought tolerance of C8 plants may not be due to greater retentionof GLA, the greater HI of C8 plants of Tuxpeno Sequıa must also beconsidered (Bolanos and Edmeades, 1993a; Bolanos et al., 1993). InC8 plants, increased demands for N from a larger developing ear arelikely met by increased remobilization of N from leaves. Thus, theabsence of accelerated leaf senescence observed by Bolanos et al.(1993) in C8 relative to C0 plants indicates significant progress fromselection for reduced senescence (Chapman and Edmeades, 1999).Maize genotypes that maintain a greater GLA are more capable offilling developing kernels in the presence of water stress since theycapture solar radiation for a longer portion of the grain-fillingperiod. Incidentally, delayed senescence has been shown to bepositively correlated with grain yield under drought stress (Betranet al., 2003) and has been recommended as a secondary trait forselection (Aparicio-Tejo and Boyer, 1983; Chapman and Edmeades,1999). The enhanced GLA of C0 and (to a lesser extent) C8 AM-
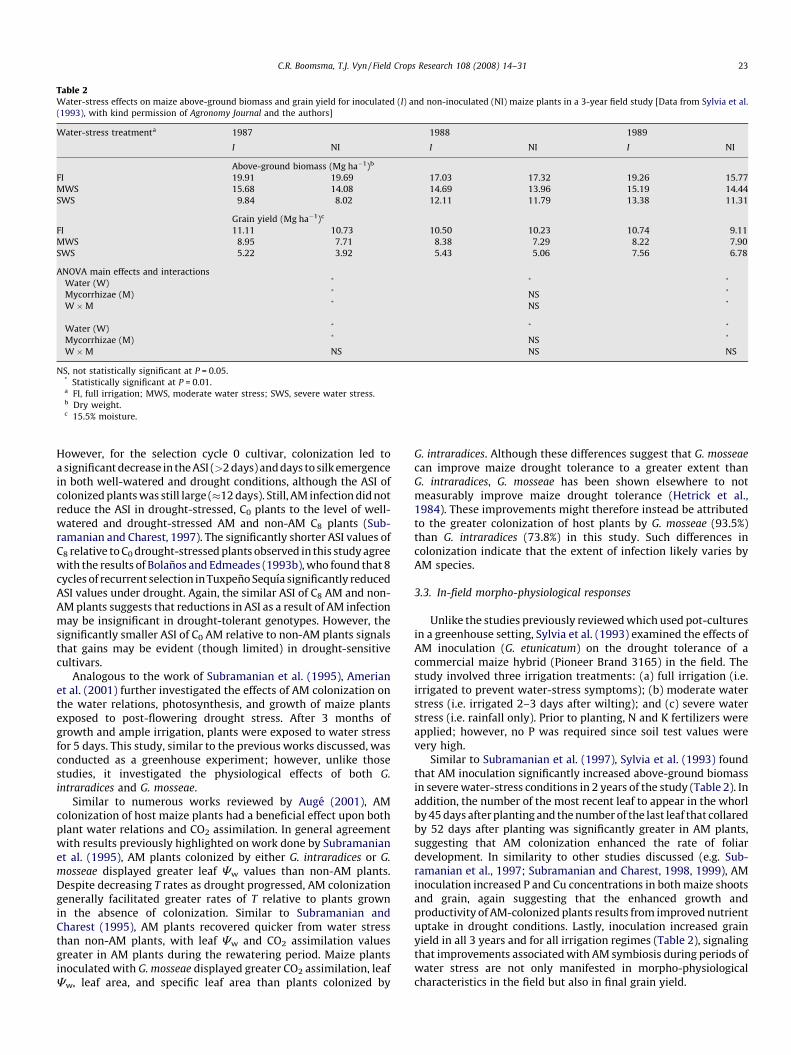
Fig. 4. Mean anthesis-silking interval (ASI) of selection cycles 0 (C0, drought-
sensitive) and 8 (C8, drought-tolerant) maize cultivars under drought-stressed (DS)
and well-watered (WW) conditions with and without arbuscular mycorrhizal (AM)
colonization. Means with different letters indicate statistically significant
differences at P � 0.05 using Tukey’s Studentized Range (HSD) test (Data from
Subramanian and Charest (1997), with kind permission of Springer Science and
Business Media and the authors).
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3122
colonized plants relative to their respective non-AM counterpartsafter 3 weeks of drought stress (Subramanian et al., 1995) istherefore significant. Such reductions in leaf senescence duringwater stress could be mediated by either enhanced chlorophyllproduction (Auge et al., 1987) or reduced degradation (i.e.photooxidation) (Subramanian and Charest, 1995), with eitherpotentially resulting from improved plant water status. Thisretention of photosynthetic capacity potentially explains thehigher foliar concentrations of soluble sugars observed bySubramanian and Charest (1995) in drought-stressed AM plants.
Similar to the observations of Subramanian and Charest (1998)during the pre-flowering period, Subramanian and Charest (1995)found that AM colonization resulted in less decline in the foliarprotein content of C0 plants under drought stress during the post-tasselling stage. Although overall foliar amino acid levels rose inboth AM and non-AM plants of both cultivars in response to waterstress, these increases were only 10.7% and 19.2% in C0 and C8 AMplants, respectively, as compared to 40.6% and 43.7% in C0 and C8
non-AM plants, respectively. This response may indicate a smallerneed among AM plants in this study to osmotically adjustsymplasm or osmoprotect enzymes due to successful droughtavoidance. However, it may also signify a reduced capabilityamong AM plants to adjust osmotically to water stress (Auge,2001). Given that amino acid levels have been observed to increasemore in AM than non-AM maize plants in response to water stressduring the pre-flowering period (Subramanian and Charest, 1998),the former rather than the latter explanation is more likely correct.
In another pot-culture, greenhouse study with the maizecultivar and selection cycles, fungal species, and imposed irrigationregimes identical to those presented by Subramanian et al. (1995),Subramanian and Charest (1997) examined the effects of AMinoculation on host-plant nutritional status, growth, and produc-tivity under drought conditions. After 3 weeks of drought followingtasselling, the shoots of both C0 and C8 AM-colonized plantsdisplayed greater N, Ca, and Cu concentrations relative to non-colonized plants of both selection cycles. After 5 weeks of irrigationfollowing drought stress, AM colonization significantly increasedshoot N, Cu, and Zn concentrations in formerly drought-stressed C0
plants and N, P, S, and Fe concentrations in previously water-stressed C8 plants. Relative to drought-stressed, non-AM plants,grain concentrations of N, Mg, and Mn were greater in water-stressed, C0 AM plants while N, P, K, Mn, and Zn concentrationswere higher in drought-stressed, C8 AM plants. Given these results,Subramanian and Charest (1997) contend that AM symbiosisimproved the drought tolerance of both tropical cultivars primarilythrough the enhanced uptake of slowly diffusing nutrients (e.g. P).Greater N levels in the shoots and grain of post-flowering AMplants (Subramanian and Charest, 1997) and shoots and roots ofpre-flowering hosts (Subramanian and Charest, 1999) are poten-tially indicative of greater N uptake in the presence of symbiosis.Since N stress increases leaf senescence and also lowers photo-synthetic rates (Banziger et al., 2000), the greater N concentrationsobserved by Subramanian and Charest (1997) in multiple tissues ofwater-stressed, AM plants potentially explain the greater GLAobserved in drought-stressed, colonized plants by Subramanianet al. (1995).
As previously discussed, selection for drought tolerance inTuxpeno Sequıa has been shown to improve grain yield in water-stress conditions primarily through the improved partitioning ofassimilates toward the developing ear at flowering (i.e. increasedHI) (Bolanos and Edmeades, 1993a; Bolanos et al., 1993; Edmeadeset al., 1999). If AM symbiosis does confer drought tolerance,colonization might be expected to increase HI in drought-sensitive,C0 and drought-tolerant, C8 plants relative to non-colonized plantsof both selection cycles in water-stress conditions. Subramanian
and Charest (1997) found that grain yield, HI, and shoot biomass(after 3 weeks of drought stress following tasselling and at harvest)were significantly greater in C0 AM relative to non-AM plantsexposed to water stress. As previously discussed, C0 AM plantsexposed to water stress during the pre-flowering period possessedgreater shoot and root biomass than their non-AM counterparts(Subramanian et al., 1997). Greater post-flowering shoot biomassmay therefore have been associated with (a) increased pre-flowering nutrient uptake due to hyphal nutrient acquisition orimproved root growth and/or (b) enhanced pre-flowering photo-assimilate accumulation in the shoot due to greater leaf area (andthus photosynthetic capacity). Improvements in HI and grain yieldin colonized C0 plants therefore potentially resulted from delays inleaf senescence combined with enhanced nutrient and assimilatetranslocation from source to sink tissues during later ear andkernel development (Subramanian et al., 1995; Rajcan andTollenaar, 1999). Yet AM symbiosis failed to significantly increasegrain yield, HI, and shoot biomass in the C8, drought-adaptedcultivar under water-stress conditions. Furthermore, under waterstress, differences in shoot mass (after 3 weeks of drought and atharvest), HI, and grain yield between C0 AM plants and AM andnon-AM plants of selection cycle 8 were not significant. Thecomparable grain yield and HI of C0 AM plants relative to C8 AMand non-AM plants suggests that AM symbiosis can potentiallyincrease the drought tolerance of a drought-sensitive cultivar.However, it also suggests that AM-mediated improvements maynot be as significant in drought-adapted varieties.
Maize is particularly susceptible to drought stress fromapproximately 10 days before till 2 weeks after 50% anthesis, withthe greatest detrimental effect occurring between the post-tasselling and pre-silking periods. Drought stress during earlyflowering often slows ear growth more than it does tassel growth,leading to a widening of the interval between anthesis and silking(Banziger et al., 2000; Campos et al., 2004). This asynchrony in floraldevelopment (i.e. large ASI values) is often associated with a delay insilk emergence, a loss of silk receptivity, a decrease in fertilization, anincrease in embryo abortion, a decrease in kernel number, and,consequently, a reduction in grain yield (Westgate, 1997; Banzigeret al., 2000; Bruce et al., 2002). In C8 plants, AM colonization did notsignificantly decrease ASI values or days to tassel or silk emergencein either drought-stressed or well-watered conditions (Fig. 4).
Table 2Water-stress effects on maize above-ground biomass and grain yield for inoculated (I) and non-inoculated (NI) maize plants in a 3-year field study [Data from Sylvia et al.
(1993), with kind permission of Agronomy Journal and the authors]
Water-stress treatmenta 1987 1988 1989
I NI I NI I NI
Above-ground biomass (Mg ha�1)b
FI 19.91 19.69 17.03 17.32 19.26 15.77
MWS 15.68 14.08 14.69 13.96 15.19 14.44
SWS 9.84 8.02 12.11 11.79 13.38 11.31
Grain yield (Mg ha�1)c
FI 11.11 10.73 10.50 10.23 10.74 9.11
MWS 8.95 7.71 8.38 7.29 8.22 7.90
SWS 5.22 3.92 5.43 5.06 7.56 6.78
ANOVA main effects and interactions
Water (W) * * *
Mycorrhizae (M) * NS *
W �M * NS *
Water (W) * * *
Mycorrhizae (M) * NS *
W �M NS NS NS
NS, not statistically significant at P = 0.05.* Statistically significant at P = 0.01.a FI, full irrigation; MWS, moderate water stress; SWS, severe water stress.b Dry weight.c 15.5% moisture.
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 23
However, for the selection cycle 0 cultivar, colonization led toa significant decrease in the ASI (>2 days) and days to silk emergencein both well-watered and drought conditions, although the ASI ofcolonized plants was still large (�12 days). Still, AM infection did notreduce the ASI in drought-stressed, C0 plants to the level of well-watered and drought-stressed AM and non-AM C8 plants (Sub-ramanian and Charest, 1997). The significantly shorter ASI values ofC8 relative to C0 drought-stressed plants observed in this study agreewith the results of Bolanos and Edmeades (1993b), who found that 8cycles of recurrent selection in Tuxpeno Sequıa significantly reducedASI values under drought. Again, the similar ASI of C8 AM and non-AM plants suggests that reductions in ASI as a result of AM infectionmay be insignificant in drought-tolerant genotypes. However, thesignificantly smaller ASI of C0 AM relative to non-AM plants signalsthat gains may be evident (though limited) in drought-sensitivecultivars.
Analogous to the work of Subramanian et al. (1995), Amerianet al. (2001) further investigated the effects of AM colonization onthe water relations, photosynthesis, and growth of maize plantsexposed to post-flowering drought stress. After 3 months ofgrowth and ample irrigation, plants were exposed to water stressfor 5 days. This study, similar to the previous works discussed, wasconducted as a greenhouse experiment; however, unlike thosestudies, it investigated the physiological effects of both G.
intraradices and G. mosseae.Similar to numerous works reviewed by Auge (2001), AM
colonization of host maize plants had a beneficial effect upon bothplant water relations and CO2 assimilation. In general agreementwith results previously highlighted on work done by Subramanianet al. (1995), AM plants colonized by either G. intraradices or G.
mosseae displayed greater leaf Cw values than non-AM plants.Despite decreasing T rates as drought progressed, AM colonizationgenerally facilitated greater rates of T relative to plants grownin the absence of colonization. Similar to Subramanian andCharest (1995), AM plants recovered quicker from water stressthan non-AM plants, with leaf Cw and CO2 assimilation valuesgreater in AM plants during the rewatering period. Maize plantsinoculated with G. mosseae displayed greater CO2 assimilation, leafCw, leaf area, and specific leaf area than plants colonized by
G. intraradices. Although these differences suggest that G. mosseae
can improve maize drought tolerance to a greater extent thanG. intraradices, G. mosseae has been shown elsewhere to notmeasurably improve maize drought tolerance (Hetrick et al.,1984). These improvements might therefore instead be attributedto the greater colonization of host plants by G. mosseae (93.5%)than G. intraradices (73.8%) in this study. Such differences incolonization indicate that the extent of infection likely varies byAM species.
3.3. In-field morpho-physiological responses
Unlike the studies previously reviewed which used pot-culturesin a greenhouse setting, Sylvia et al. (1993) examined the effects ofAM inoculation (G. etunicatum) on the drought tolerance of acommercial maize hybrid (Pioneer Brand 3165) in the field. Thestudy involved three irrigation treatments: (a) full irrigation (i.e.irrigated to prevent water-stress symptoms); (b) moderate waterstress (i.e. irrigated 2–3 days after wilting); and (c) severe waterstress (i.e. rainfall only). Prior to planting, N and K fertilizers wereapplied; however, no P was required since soil test values werevery high.
Similar to Subramanian et al. (1997), Sylvia et al. (1993) foundthat AM inoculation significantly increased above-ground biomassin severe water-stress conditions in 2 years of the study (Table 2). Inaddition, the number of the most recent leaf to appear in the whorlby 45 days after planting and the number of the last leaf that collaredby 52 days after planting was significantly greater in AM plants,suggesting that AM colonization enhanced the rate of foliardevelopment. In similarity to other studies discussed (e.g. Sub-ramanian et al., 1997; Subramanian and Charest, 1998, 1999), AMinoculation increased P and Cu concentrations in both maize shootsand grain, again suggesting that the enhanced growth andproductivity of AM-colonized plants results from improved nutrientuptake in drought conditions. Lastly, inoculation increased grainyield in all 3 years and for all irrigation regimes (Table 2), signalingthat improvements associated with AM symbiosis during periods ofwater stress are not only manifested in morpho-physiologicalcharacteristics in the field but also in final grain yield.
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3124
4. Future research
Research discussed in detail in this review (i.e. Sylvia et al., 1993;Subramanian et al., 1995, 1997; Subramanian and Charest, 1995,1997, 1998, 1999; Schellenbaum et al., 1998; and Amerian et al.,2001) illustrates that AM symbiosis can positively affect maizegrowth, development, and overall productivity under water-limit-ing conditions. However, it is important to note that the majority ofthese studies (i.e. Subramanian and Charest, 1995, 1997, 1998, 1999;Subramanian et al., 1995, 1997; Schellenbaum et al., 1998; Amerianet al., 2001) were conducted in a pot-culture, greenhouse environ-ment. Results would likely differ to some extent in field conditionsand at the canopy level of organization, particularly since thenumber of plants sampled in these greenhouse studies wasrelatively small for both the AM and non-AM treatments. Wheninterpreting the results of these studies, readers should also notethat the tropical cultivar Tuxpeno Sequıa is open-pollinated and nota hybrid per se, meaning plants sampled were genetically distinct.This genetic variation, combined with potentially small samplesizes, may have resulted in artificially large differences between AMand non-AM plants in some cases. Nevertheless, the studiesreviewed here provide a general overview of AM effects on maizedrought tolerance, and suggest that future research on this topic mayyield improvements in maize drought tolerance.
At present, research significantly expounding upon the resultsof the studies discussed in this review is currently lacking,particularly in a field setting. Further investigation into thealleviation of maize drought stress through AM symbiosis haspotential benefits including (a) an improved understanding of theeffects of AM colonization on drought-stress maize physiology; (b)the creation of commercial maize varieties with enhancedproductivity during drought stress in fertile and infertile soils;and (c) the identification and adoption of economically-viableproduction practices which facilitate AM colonization and survivalin water- and nutrient-limiting conditions. Potential avenues forfuture research are therefore numerous, but could potentiallyfocus on the following areas.
4.1. Treatments, field research, and cultural practices
Future physiological studies examining AM effects on maizedrought tolerance should employ variations of the treatmentsimposed in the studies previously reviewed. For example, theduration, timing, and/or number of drought episodes could bevaried to examine morpho-physiological responses to water stressat multiple stages of development (Amerian et al., 2001; Bruceet al., 2002). Such changes in occurrence length, timing, andnumber could simulate the severe drought conditions experiencedby maize hybrids in temperate and tropical dryland-productionenvironments throughout a growing season. Furthermore, studiesshould examine morpho-physiological responses in varieties otherthan those repeatedly used in the studies previously highlighted(i.e. C0 and C8 plants of Tuxpeno Sequıa).
Further field examination of many of the morpho-physiologicaleffects characterized in greenhouse studies of drought-stressed,AM plants would be highly beneficial. Such research couldelaborate upon the treatments and cultural practices used bySylvia et al. (1993). The effects of AM on a variety of crop plantshave typically been studied using pot-cultures in greenhouse orgrowth chamber environments in which specific AM effects, fungalproducts, and fungal–plant interactions were studied in acontrolled manner (Wright, 2005). This is also true in maize, asevident in nearly all of the studies examined in detail in thisreview. Yet in field settings, AM symbiosis is affected by factors notpresent in controlled greenhouse or laboratory conditions. For
example, light intensity and volume for root growth may belimited in greenhouse, pot-culture studies, potentially affectingphotosynthesis and root expansion (i.e. lateral root development)(Wright, 2005). In order to fully understand AM effects on maizedrought tolerance, future experiments should be conducted in fieldenvironments that mimic typical growing conditions in bothtemperate and tropical regions. Such field studies might involverain-out shelters and a combination of arid, irrigated, and rainfedfields. These studies could be conducted at varying levels of waterstress and nutrient availability and at multiple locations so that the‘drought � soil fertility’ interaction could be investigated across anumber of environments.
Cropping practices which mimic those used by many growersshould also be imposed in future field research, including the use ofvarious nutrient amendments, tillage systems, plant densities, croprotations, and pesticide applications appropriate for optimumgrain yield in a given environment. The findings of a number ofstudies could potentially serve as guides for examining the effectsof cropping practices on the AM-mediated alleviation of maizedrought stress. For example, recent work by Tanaka and Yano(2005) has already highlighted some of the effects of form of Napplied on hyphal N transport in maize, and a recent review by Liuet al. (2007) extensively examined AM effects on plant N nutrition.In another recent review, Kabir (2005) described the effects oftillage practices on AM, while in a potted study, Karasawa et al.(2000) investigated the effects of previous AM and non-AM cropson subsequent maize growth and AM colonization at various soilmoisture levels. In a field study, Johnson et al. (1991) examined theeffects of maize and soybean (Glycine max (L.) Merrill) croppinghistory on the species composition of AM communities. Similarly,Gavito and Miller (1998) investigated the effects of crop rotation,tillage, and P fertilization on early AM intraradical colonization inmaize. Although preliminary evidence suggests that glyphosatedoes not significantly affect AM colonization in maize (Daigh et al.,2006) and other crops (Daigh et al., 2006; Malty et al., 2006), themarkedly increasing use of this herbicide in glyphosate-resistantmaize production warrants future research into its effects on AMcolonization, prolificacy, and survival.
4.2. Secondary traits, conventional breeding, and QTL analysis
Improvements in AM-mediated drought tolerance throughconventional breeding will likely require the selection of secondarytraits and potentially the identification, analysis, and use of theirrespective quantitative trait loci (QTLs). A full examination of thesecondary traits, phenotypic screening methods, managed environ-ments, breeding techniques, and QTL analysis strategies common inmaize drought-stress research are (a) beyond the scope of this articleand (b) extensively reviewed by others (Edmeades et al., 1997b;Banziger et al., 2000; Bruce et al., 2002; Tuberosa et al., 2002;Campos et al., 2004). Readers are therefore encouraged to consultthese articles and works cited therein for further information inthese areas.
Traditionally, increasing maize grain yield under droughtthrough direct selection for yield has been restricted by its lowheritability and large ‘genotype� environment’ interaction. How-ever, given that selection for AM colonization and its beneficialeffects on maize drought tolerance has been minimal to nonexistent,breeding programs may benefit from initial selection for yield. Still,morpho-physiological traits correlated with yield and displayingincreased genetic variability in drought conditions have beencommonly identified and selected for in maize breeding programs(Bolanos et al., 1993; Bolanos and Edmeades, 1996; Tuberosa et al.,2002; Campos et al., 2004) and may similarly be helpful in AM-basedmaize breeding programs. Morpho-physiological traits such as
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 25
rooting intensity and depth, WUE, T rate, water extraction timing,leaf rolling and erectness, diurnal Cw, and leaf osmotic potentialhave facilitated the identification of hybrid differences in plantwater status, root exploration, and water extraction capacity underdrought conditions. Similarly, chlorophyll concentration has servedas a measure of a variety’s stay-green capability (i.e. delayed foliarsenescence) (Bolanos et al., 1993; Pena-Valdivia et al., 1997; Betranet al., 2003; Campos et al., 2004). Other useful secondary traits havebeen associated with various developmental stages (e.g. flowering,grain-filling). In particular, the selection for reduced ASI values hasprovided a rapid and effective mechanism for the creation of stable,high-yielding maize varieties (Bolanos and Edmeades, 1993b; Bruceet al., 2002). Further beneficial traits associated with flowering andgrain-filling include ears plant�1, kernels ear�1, kernels plant�1,tassel size, and HI (Bolanos and Edmeades, 1993a; Bolanos et al.,1993; Edmeades et al., 1997b; Chapman and Edmeades, 1999;Edmeades et al., 1999). A number of these morpho-physiologicaltraits along with those associated with enhanced drought tolerancein AM-colonized maize (e.g. greater leaf Cw, T, gs, GLA) in the studiespreviously discussed (i.e. Subramanian and Charest, 1995, 1997,1998, 1999; Subramanian et al., 1995, 1997; Amerian et al., 2001)could be further examined in drought-stress, physiological researchprograms utilizing AM and non-AM plants of multiple genotypes.Some of these morpho-physiological responses might also serve aseffective secondary traits (i.e. cheap and easy to score/measure, highgenetic correlation with yield, highly heritable, genetically variable,not associated with grain yield loss in ideal conditions) for indirectselection of yield under drought conditions.
When comparing the morpho-physiological responses of AMand non-AM maize to water stress, future studies may benefit fromusing the top-down approach proposed by Passioura (1977, 1996)for investigating crop drought tolerance. This approach is basedupon resource economics, and seeks to identify factors thatdetermine the effectiveness with which a crop can use a restrictedwater supply to produce a harvestable yield (Passioura, 1996). Asreviewed by Passioura (1996), the most effective use of waterinvolves three principle factors: (a) acquiring as much water aspossible; (b) using the acquired water as efficiently as possible toproduce photoassimilates (e.g. restricting water loss during CO2
uptake); and (c) converting a maximal amount of photoassimilatesinto harvestable yield (typically grain in maize). For maize, each ofthese factors can be expressed using the following identity:
GY ¼ T �WUE�HI (1)
where GY is the amount of grain produced, T is the volume of watercaptured by the plant and subsequently transpired, WUE is theamount of biomass produced per unit of transpired water, and HI isthe proportion of biomass allocated to the grain. An increase in anyof these factors is likely to increase yield, while any morpho-physiological phenomenon unrelated to any of these componentsis unlikely to improve water-limited grain production (Passioura,1996).
From the limited number of studies extensively examined inthis review, initial suppositions can be made concerning whichcomponent(s) of this identity may be affected most by AMcolonization of drought-stressed maize. For example, greater pre-flowering leaf Cw (Subramanian et al., 1997), pre- and post-flowering tissue N concentrations (Subramanian and Charest,1997, 1999), pre-flowering root biomass (Subramanian et al.,1997), and post-flowering gs and T (Subramanian et al., 1995;Amerian et al., 2001) in AM-colonized, water-stressed plantssuggest AM may increase the T component. Similarly, sincedrought-stress treatments were identical for both AM and non-AMplants, higher pre-flowering shoot and root biomass (Subramanian
et al., 1997), post-flowering GLA (Subramanian et al., 1995), andpre-flowering shoot amino acid concentrations (Subramanian andCharest, 1998) in colonized, drought-stressed plants may indicateAM-mediated improvements in the WUE component (i.e. AMplants exhibited greater productivity per unit available water).Some observed physiological phenomena are likely to beassociated with more than one component of the identity proposedby Passioura (1977). For example, enhancements in plant N statusvia AM symbiosis could result in increased T (e.g. higher soil Nuptake results in greater shoot and root biomass, GLA, and,subsequently, T). However, given that (a) WUE can be greater in C4
plants at higher leaf N concentrations (Sage and Pearcy, 1987) and(b) greater N availability can improve WUE in maize (e.g. Derconet al., 2006), increased N uptake via AM symbiosis might alsoenhance WUE. In the future, it would therefore be beneficial tosimultaneously analyze AM effects on water capture and T, WUE,and biomass partitioning. Further research based upon thisproposed top-down approach could be helpful in proving and/ordisproving past and present suppositions surrounding the effectsof AM on maize morpho-physiological responses to water stress.
The identification of drought-tolerant genotypes based solelyupon the phenotypic expression of morpho-physiological traits hasproven difficult. The use of molecular markers for the identificationof QTLs controlling particular secondary traits has eliminated someof these constraints. A number of studies have reported QTLsassociated with specific traits observed under drought stress, withmost of these efforts having targeted grain yield, ASI, root traits,plant water use (including gs), and leaf and xylem ABA content(Tuberosa et al., 2002; Campos et al., 2004). Yet studies employingQTL analysis to understand AM-mediated changes in maize droughttolerance are presently lacking. Nevertheless, Kaeppler et al. (2000)detected QTL for growth at low P and AM responsiveness in aB73�Mo17 population of 197 recombinant inbred lines (RILs),indicating that QTL analysis can be effectively used to examine theresponsiveness among maize lines to AM symbiosis. The QTLtechniques detailed by Tuberosa et al. (2002) and Campos et al.(2004) and articles cited within each review could be used toelucidate the genetic basis of traits conferring enhanced droughttolerance in AM-colonized plants. Given the large number ofmolecular markers available for maize, QTL analysis could alsoenhance the efficiency and ease of selection of these traits. In general,it would likely be beneficial to search for QTLs that improve grainyield in both water-limiting and well-watered conditions. Still, asdiscussed by Campos et al. (2004), a variety of challenges exist inidentifying QTLs associated with drought-tolerant phenotypes (e.g.most QTL for drought are only detected in water-stress or optimalconditions), and these challenges may be more pronounced whenseeking improvements through AM symbiosis.
4.3. AM colonization
Although AM-mediated improvements in maize droughttolerance will clearly involve the identification of and selectionfor secondary traits, further efforts must also incorporate theselection of maize genotypes exhibiting effective AM colonizationunder water-stress conditions. The proportion of modern geno-types capable of forming an effective symbiosis in water-limitingenvironments is generally unknown. Toth et al. (1990) contendthat past selection in maize for fungal and bacterial pathogenresistance has reduced the potential for AM colonization in somemaize inbreds, and thus the number of current genotypes capableof forming an effective symbiosis may be limited. However, maizecultivars are clearly capable of forming beneficial AM symbiosesin not only drought environments (e.g. Subramanian et al., 1995;Subramanian and Charest, 1997; Schellenbaum et al., 1998;
9 LEA proteins perform such functions during water stress as protein and
membrane structural maintenance, water binding, and ion sequestration (Close,
1996; Koag et al., 2003; Ruiz-Lozano, 2003; Porcel et al., 2005a), and are therefore
often associated with drought tolerance in plants (Close, 1996; Xu et al., 1996).10 For further details on the principles of DNA-based gene expression microarrays,
their potential applications and benefits in drought-stress research, and their
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3126
Amerian et al., 2001) but also low P conditions (e.g. Khalil et al.,1994; Kaeppler et al., 2000; Liu et al., 2000a, 2000b; Zhu et al.,2005). Furthermore, effective AM colonization under water-stressconditions can occur in both drought-susceptible and drought-tolerant genotypes (e.g. Subramanian and Charest, 1995; Schel-lenbaum et al., 1998). Investigating the percent colonization ofthree improved (i.e. cultivated) and three unimproved (i.e. wild-type) maize cultivars, Khalil et al. (1994) discovered that 49–68% ofmaize roots were colonized by AM, with the level of AMcolonization similar among cultivated and wild-type cultivars.Examining the variation in colonization and responsiveness ofMidwest-adapted, maize inbreds to AM symbiosis in low and highP conditions, Kaeppler et al. (2000) found that all inbred lines werecolonized by at least a single AM species. Although responsivenessto AM colonization was highly variable among the inbredsexamined, the authors concluded that modern, adapted genotypescan form beneficial symbioses despite the selection of thesegenotypes in adequate P conditions. In the study by Sylvia et al.(1993) previously discussed, inoculation of the maize hybridPioneer Brand 3165 resulted in the rapid and sustained coloniza-tion of roots, while an absence of inoculation resulted in poorcolonization. Also, in a single year of the 3-year study, water stressincreased AM colonization. Although the potential for effectivecolonization among today’s genotypes is relatively unknown, theselimited results suggest that the proportion of genotypes capable ofestablishing AM symbiosis in water- and nutrient-stress condi-tions may be significant. However, a thorough analysis of thepotential for effective intra-radical colonization throughoutmodern maize germplasm is clearly necessary, particularly inRoundup Ready1 hybrids receiving glyphosate application.
Although future research should focus on identifying maizevarieties capable of forming an effective symbiosis, it should alsodetermine fungal isolates, species, and communities that demon-strate the greatest enhancement of maize productivity in water-limiting environments in both fertile and infertile soils. Morpho-logically-identical isolates of a particular fungal species can havevarying effects on host growth, with the effects of a single isolatevarying by such environmental factors as soil temperature, pH,moisture, nutrient status, and salinity (Picone, 2003). Although anAM isolate or species can benefit a particular genotype, it may notdo so for another (Liu et al., 2000b). These conditional phenomenasuggest that beneficial AM species may need to be determined bothfor specific genotypes and local production environments. Suchspecies need to be capable of surviving and spreading amongnative AM communities (when introduced) and in typicalenvironmental and agronomic conditions8.
Although AM species must be chosen which effectively colonizemaize genotypes, some research suggests that the importance of AMstrain-maize genotype specificity may be negligible in somecircumstances. Reviewing assorted past studies which comparedthe effectiveness of multiple AM strains on multiple crops, Riveraet al. (2007) suggest that crop-AM strain specificity has relativelylittle importance, while ‘‘selecting strains for their effectiveness ingiven soil types is crucial’’. Effective AM strains may therefore besimilar across maize genotypes, but may differ based upon soil types.Although this presents challenges for future AM-based hybriddevelopment, these challenges may be overcome by multi-sitetesting at breeding and research stations situated on various soiltypes. Still, it is obvious that more research is needed on AM strain-maize genotype specificity before the importance of specific AM-genotype pairings across multiple soil types is fully understood.
8 For further information on the use of AM inoculation as an agricultural input,
including the selection of superior fungal species and the production and
application of inoculum, refer to Picone (2003).
4.4. Molecular and genomic techniques
Further understanding of maize metabolic and physiologicalresponses to AM infection during water stress could be enhancedthrough the use of molecular and genomic techniques commonlyavailable to the maize research community. A review by Ruiz-Lozano (2003) highlights aspects of AM symbiosis under waterstress that should be further investigated on a molecular level. Theauthor suggests focusing future research on the molecular aspectsof osmotic adjustment, aquaporins, antioxidant proteins, and lateembryogenesis abundant (LEA) proteins9. Although molecular-based studies examining AM effects on maize drought toleranceare lacking, such studies have occurred in other crops (e.g. Porcelet al., 2005a,b, 2006). Similar techniques could be applied in maize.Ruiz-Lozano (2003) suggests that future studies could involve non-targeted screenings of cDNA libraries from both AM fungi and AM-colonized plants. Such techniques could enable the identificationof water stress-regulated genes that permit enhanced droughttolerance in AM-colonized hosts.
With the advent of technologies such as expressed sequence tag(EST) sequencing and microarrays, analysis of the transcriptome (i.e.the mRNA pool of a cell at a particular moment) is now possible, thuspermitting the detection of significant changes in the expression of awide spectrum of genes. Gene expression during AM symbiosis hasbeen previously examined in a variety of plants (Harrison, 1999;Parniske, 2004; Balestrini and Lanfranco, 2006) including maize (e.g.Bonfante et al., 1996; Kaldorf et al., 1998; Ravnskov et al., 2003).However, to the best of the authors’ knowledge, no studies haveextensively detailed gene expression profiles in AM-colonized maizeduring drought stress. Gene expression profiling has enabled theidentification of a large number of genes expressed during theestablishment and latter stages of AM symbiosis (Balestrini andLanfranco, 2006) and could similarly be used to examine geneexpression patterns unique to AM-colonized plants during droughtstress. Using microarrays, drought-tolerance mechanisms (i.e.morpho-physiological and molecular processes) associated withAM symbiosis could be either identified or more fully understoodthrough the comparison of AM and non-AM plants (e.g. AM-minusmutants) of the same genotype. In both plant types, gene expressioncould be analyzed in multiple vegetative and reproductive tissuesand at several developmental stages.
For example, Zinselmeier et al. (2002) examined the impacts oftransient water limitations on maize ear and early kernelformation through the analysis of gene expression in femalereproductive tissues. The microarray used was designed to providea survey of 27 maize metabolic pathways, therefore permitting anon-targeted genomic approach for identifying gene function. Theuse of a similar technique could facilitate the discovery of sets ofgenes which are differentially expressed either temporally orspatially during drought stress and which co-vary with AM-colonized, drought-tolerant genotypes. Genes responsible for theregulation of these mechanisms could then potentially serve asmolecular markers for the more efficient selection of AM-colonized, drought-tolerant plants10.
limitations for gene discovery, refer to Zinselmeier et al. (2002) and Kathiresan et al.
(2006). For an extensive review of fungal and plant gene expression profiles during
the early stages of plant-AM interactions and later symbiotic phases, readers are
encouraged to consult Franken and Requena (2001) and Balestrini and Lanfranco
(2006).
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 27
5. Considerations
Whether in temperate or tropical maize production systems, thebenefits of AM symbiosis (e.g. enhanced P uptake, improved droughttolerance) will only be realized when management practices areemployed that sustain native and introduced AM populations. Suchpractices include (a) incorporating reduced tillage systems toprevent the destruction of AM hyphal networks; (b) maintainingcover crops, intercrops, or continuous cropping to providecarbohydrate sources for AM persistence; (c) reducing commercialfertilizer P application to help avoid selection pressure for ineffectiveAM and reductions in root colonization, soil hyphae, and sporepopulations; and (d) avoiding fungicides to maintain effective AMpopulations (Johnson and Pfleger, 1992; Picone, 2003; Plenchetteet al., 2005; Garcia et al., 2007). In existing conventional productionsystems based upon high fertilizer and fungicide inputs, simplisticcrop rotations or continuous maize monocultures, and intensivetillage practices, the growth of AM-symbiotic hybrids will likelyneed to be accompanied by the adoption of cropping practices whichfavor AM long-term survival and repeated colonization (Ryan andGraham, 2002; Plenchette et al., 2005). Still, as shown by Sylvia et al.(1993), AM symbiosis can improve maize growth and final grainyield under water-stress conditions even in conventional produc-tion systems involving intensive tillage practices (i.e. moldboardplow and disking) and synthetic fertilizer application. Furthermore,the management of AM native to agricultural soils seems achievableeven in soils previously exposed to intensive crop production(Hamel and Strullu, 2006).
Whether commercial maize seed companies in developednations will consider AM-derived benefits in their maize researchand breeding programs is debatable. As Hamel and Strullu (2006)point out, plant breeding is a business directed by the requests ofthe agricultural industry in general. In countries dominated byhigh-input maize production systems, breeding for effective AMsymbiosis potentially awaits the adoption of cropping practicesthat take the survival, health, and potential benefits of AM intoconsideration. With increases in synthetic fertilizer costs, reduc-tions in water supplies for irrigation, and heightening publicconcern surrounding the environmental impacts of modern cropproduction, the adoption rate of cropping systems which take AMinto account may grow in the future. Still, many practical issuesneed to be addressed before AM symbiosis becomes a viabletechnology that can (a) facilitate improvements in maize abioticstress tolerance and (b) enable enhanced commercial-scale maizeproduction with reduced resource inputs. Questions such as thefollowing therefore need to be addressed.
(i) C
an commercial maize producers depend upon indigenous AMpopulations for effective colonization of maize or do growersneed to routinely apply AM inoculum?
(ii) H
ow can large-scale, uniform inoculation be achieved inmodern maize production systems?
(iii) H
ow can AM inoculum be produced and applied in aneconomically-viable, commercial-scale manner when applica-tion is necessary?
(iv) W
hat tillage practices, crop rotations, and other crophusbandry practices need to be implemented to effectivelyproduce an AM-based maize crop?
(v) In
modern maize breeding and research programs, how canlarge-scale, uniform inoculation and extensive plant genotypescreening for effective AM colonization be achieved acrossnumerous maize genotypes colonized by a variety of AMspecies?
(vi) H
ow can optimal AM species-maize genotype combinationsbe effectively and economically identified?
Answering these and similar questions is challenging,particularly since research addressing these questions has beenrather limited. Nevertheless, as a result of applied researchfocused upon the practical use of AM in crop production inaddition to recent advances in molecular biology, answers tosome of these questions are already emerging. For example, incountries such as India and Cuba, successful cropping systemsbased upon AM inoculation have been developed and imple-mented to increase crop productivity and lower production costs(Hamel and Strullu, 2006). In large (land areas up to 1000 ha),high-input crop production systems in Cuba and Bolivia, AM(Glomus fasciculatum) inoculum applied to maize seeds improvedmaize yields from 21 to 78%, thus demonstrating the ability touse AM inoculum in large, input-intensive production systems(Rivera et al., 2007).
In some cases, improving maize productivity through the use ofAM inoculum instead of indigenous AM populations may bepreferable or even necessary (e.g. in areas that received extensiveuse of fungicides or on severely degraded land) (Douds et al., 2005).Unfortunately, due to the obligate biotrophic nature of AM, the costof inoculum production is presently quite high. Thus, withadvances in inoculum production technologies and the creationof inoculums for field use, the use of AM could be a viable practicein maize production (Saito and Marumoto, 2002; Hamel andStrullu, 2006). Given the high cost of commercial inoculum, on-farm production of AM may be a viable alternative, particularly indeveloping countries (Douds et al., 2005).
If research and breeding efforts aimed at improving maizedrought tolerance through AM symbiosis are to arise, they wouldlikely benefit from the use of modern DNA-molecular techniques.Polymerase chain reaction (PCR) techniques utilizing AM-specificprimers have recently been used in maize, soybean, alfalfa(Medicago sativa L. subsp. sativa), sudan grass (Sorghum sudanense
[Piper] Staph), and other species to rapidly, reliably, and relativelyinexpensively identify AM species (G. intraradices, G. mosseae) andestimate species composition after measuring AM colonizationrates via trypan blue staining (Alkan et al., 2004; Ishii andLoynachan, 2004; Sharrock et al., 2004). Multiple techniques forisolating AM spores from the soil (Gupta and Mukerji, 2002) anddetermining spore viability (Bansal and Mukerji, 2002) have alsobeen recently reviewed, and, combined with DNA-based techni-ques, would be beneficial to future research.
Presently, potential benefits from AM symbiosis in maize may bemost significant in drought-prone, infertile regions of the tropics (i.e.portions of Africa and Asia) in which maize production does notcommonly involve intensive irrigation and fertilizer application;practices common in the high-input systems of industrialized,developed nations. In the soils of many of these regions, nearly 80% ofapplied P is not immediately available, with much being convertedto unavailable forms (Sanchez, 2002; Picone, 2003). In semi-aridregions and areas of recurrent drought, the soil is often inherentlyinfertile or depleted of nutrients (principally N and P) and low inorganic matter as a result of grower practices (Kamara et al., 2005;Cardoso and Kuyper, 2006; Ncube et al., 2007). Maize is thereforeroutinely exposed to multiple abiotic stresses in these areas. Withinorganic fertilizers potentially costing at least twice as much inAfrica relative to Europe and North America (Sanchez, 2002) and theincomes of tropical farmers far below those in many temperate areas(Cardoso and Kuyper, 2006), the benefits of AM for improved maizedrought tolerance in adapted cultivars may be most substantialwhen both nutrient deficiencies and water limitations simulta-neously lower crop productivity and when economic constraintsand government policies prevent grower access to syntheticfertilizers and irrigation technologies. As reviewed by Rivera et al.(2007), inoculation with effective AM strains may be simpler and
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3128
more effective than manipulation of native AM populations.However, in portions of Africa and Asia, the management of ordependence upon indigenous fungi as opposed to the introduction ofselected strains may be recommended due to the relatively high costof AM inoculum (Plenchette et al., 2005) and limited grower accessto AM resources.
6. Conclusions
Long-term trends in global climate change and the expansion ofmaize production into marginal areas are generating a greaternumber of drought-prone maize production environments acrossthe globe. Increases in global temperatures as a result of risinggreenhouse gas concentrations could potentially accelerate maizegrowth and development, hasten maturity, and reduce soilmoisture. Improvements in maize drought tolerance are thereforevital for maintaining local and global food security, particularly inAfrica where climate change is predicted to significantly increasethe intensity and frequency of drought stress (IPCC, 2007). Fromthe limited number of studies extensively examined in this review(i.e. Sylvia et al., 1993; Subramanian and Charest, 1995, 1997,1998, 1999; Subramanian et al., 1995, 1997; Schellenbaum et al.,1998; Amerian et al., 2001), it is evident that AM symbiosis canpotentially improve maize drought tolerance. The alleviation ofdrought stress through AM symbiosis has potential benefitsincluding the creation of commercial maize hybrids with enhancedproductivity during drought stress in fertile and infertile soils andthe identification and adoption of economically-viable productionpractices that facilitate AM colonization and survival in water- andnutrient-limiting conditions. Yet clearly more extensive research isnecessary, particularly in field settings across multiple environ-ments. Future endeavors seeking to elucidate, validate, andimplement improvements in drought tolerance through AMsymbiosis will likely require extensive collaboration betweencrop physiologists, plant breeders, genomicists (Edmeades et al.,2004), and molecular and soil biologists.
Acknowledgements
This work was supported by the Pioneer Hi-Bred Intl., Inc.Fellowship in Plant Sciences and the Purdue University AndrewsFoundation Fellowship. The authors would like to thank M. Canepa,M.T. Humphreys, and B.A. Ochs for helpful comments andsuggestions. The authors would also like to express gratitude toan anonymous reviewer who provided numerous helpful com-ments that markedly improved this review. We apologize tocolleagues whose work was not cited in this article due to spacelimitations.
References
Acosta-Avalos, D., Alvarado-Gil, J.J., Vargas, H., Frıas-Hernandez, J., Olalde-Portugal,V., Miranda, L.C.M., 1996. Photoacoustic monitoring of the influence of arbus-cular mycorrhizal infection on the photosynthesis of corn (Zea mays L.). PlantSci. 119, 183–190.
Alkan, N., Gadkar, V., Coburn, J., Yarden, O., Kapulnik, Y., 2004. Quantification of thearbuscular mycorrhizal fungus Glomus intraradices in host tissue using real-time polymerase chain reaction. New Phytol. 161, 877–885.
Amerian, M.R., Stewart, W.S., Griffiths, H., 2001. Effect of two species of arbuscularmycorrhizal fungi on growth, assimilation and leaf water relations in maize (Zeamays). Asp. Appl. Biol. 63, 73–76.
Aparicio-Tejo, P.M., Boyer, J.S., 1983. Significance of accelerated leaf senescence atlow water potentials for water loss and grain yield in maize. Crop Sci. 23, 1198–1202.
Atkinson, D., Berta, G., Hooker, J.E., 1994. Impact of mycorrhizal colonization on rootarchitecture, root longevity and the formation of growth regulators. In: Giani-nazzi, S., Schuepp, H. (Eds.), Impact of Arbuscular Mycorrhizas on SustainableAgriculture and Natural Ecosystems. Birkhauser Verlag, Basel, Switzerland, pp.89–99.
Auge, R.M., 2001. Water relations, drought and vesicular-arbuscular mycorrhizalsymbiosis. Mycorrhiza 11, 3–42.
Auge, R.M., Schekel, K.A., Wample, R.L., 1987. Leaf water and carbohydrate status ofVA mycorrhizal rose exposed to drought stress. Plant Soil 99, 291–302.
Auge, R.M., Duan, X., Ebel, R.C., Stodola, A.J.W., 1994. Nonhydraulic signaling of soildrying in mycorrhizal maize. Planta 193, 74–82.
Azcon, R., Tobar, R.M., 1998. Activity of nitrate reductase and glutamine synthetasein shoot and root of mycorrhizal Allium cepa. Effects of drought stress. Plant Sci.133, 1–8.
Azcon, R., Gomez, M., Tobar, R., 1992. Effects of nitrogen source on growth, nutrition,photosynthetic rate and nitrogen metabolism of mycorrhizal and phosphorus-fertilized plants of Lactuca sativa L. New Phytol. 121, 227–234.
Azcon, R., Gomez, M., Tobar, R., 1996. Physiological and nutritional responses byLactuca sativa L. to nitrogen sources and mycorrhizal fungi under droughtconditions. Biol. Fertil. Soils 22, 156–161.
Azcon-Aguilar, C., Bago, B., 1994. Physiological characteristics of the host plantpromoting an undisturbed functioning of the mycorrhizal symbiosis. In: Gia-ninazzi, S., Schuepp, H. (Eds.), Impact of Arbuscular Mycorrhizas on SustainableAgriculture and Natural Ecosystems. Birkhauser Verlag, Basel, Switzerland, pp.47–60.
Balestrini, R., Lanfranco, L., 2006. Fungal and plant gene expression in arbuscularmycorrhizal symbiosis. Mycorrhiza 16, 509–524.
Bansal, M., Mukerji, K.G., 2002. Methods in study of viability of VAM fungal spores.In: Mukerji, K.G., Manoharachary, C., Chamola, B.P. (Eds.), Techniques in Mycor-rhizal Studies. Kluwer Academic Publishers, Dordrecht, Netherlands, pp. 217–229.
Banziger, M., Edmeades, G.O., Beck, D., Bellon, M., 2000. Breeding for Drought andNitrogen Stress Tolerance in Maize: From Theory to Practice. CIMMYT,Mexico D.F. , http://www.cimmyt.org/Research/Maize/map/guides_tools/Bred-Droug Maize/BredDroug.htm.
Bennett, J.M., Jones, J.W., Zur, B., Hammond, L.C., 1986. Interactive effects ofnitrogen and water stresses on water relations of field-grown corn leaves.Agron. J. 78, 273–280.
Berta, G., Fusconi, A., Trotta, A., 1993. VA mycorrhizal infection and the morphologyand function of root systems. Environ. Exp. Bot. 33, 159–173.
Bethlenfalvay, G.J., Schuepp, H., 1994. Arbuscular mycorrhizas and agroecosystemstability. In: Gianinazzi, S., Schuepp, H. (Eds.), Impact of Arbuscular Mycorrhizason Sustainable Agriculture and Natural Ecosystems. Birkhauser Verlag, Basel,Switzerland, pp. 117–131.
Betran, F.J., Beck, D., Banziger, M., Edmeades, G.O., 2003. Secondary traits in parentalinbreds and hybrids under stress and non-stress environments in tropicalmaize. Field Crops Res. 83, 51–65.
Bolanos, J., Edmeades, G.O., 1991. Value of selection for osmotic potential in tropicalmaize. Agron. J. 83, 948–956.
Bolanos, J., Edmeades, G.O., 1993a. Eight cycles of selection for drought tolerance inlowland tropical maize. I. Responses in grain yield, biomass, and radiationutilization. Field Crops Res. 31, 233–252.
Bolanos, J., Edmeades, G.O., 1993b. Eight cycles of selection for drought tolerance inlowland tropical maize. II. Responses in reproductive behaviour. Field CropsRes. 31, 253–268.
Bolanos, J., Edmeades, G.O., 1996. The importance of the anthesis-silking interval inbreeding for drought tolerance in tropical maize. Field Crops Res. 48, 65–80.
Bolanos, J., Edmeades, G.O., Martinez, L., 1993. Eight cycles of selection of droughttolerance in lowland tropical maize. III. Responses in drought adaptive phy-siological and morphological traits. Field Crops Res. 31, 269–286.
Bonfante, P., Bergero, R., Uribe, X., Romera, C., Rigau, J., Puigdomenech, P., 1996.Transcriptional activation of a maize alpha-tubulin gene in mycorrhizal maizeand transgenic tobacco plants. Plant J. 9, 737–743.
Bray, E.A., Bailey-Serres, J., Weretilnyk, E., 2000. Responses to abiotic stresses. In:Buchanan, B., Gruissem, W., Jones, R. (Eds.), Biochemistry & Molecular Biologyof Plants. American Society of Plant Physiologists, Rockville, MD, pp. 1158–1203.
Bressan, W., Vasconcellos, C.A., 2002. Alteracoes morfologicas no sistema radiculardo milho induzidas por fungos micorrızicos e fosforo. Pesq. Agropec. Bras. 37,509–517.
Bruce, W.B., Edmeades, G.O., Barker, T.C., 2002. Molecular and physiologicalapproaches to maize improvement for drought tolerance. J. Exp. Bot. 53, 13–25.
Bryla, D.R., Eissenstat, D.M., 2005. Respiratory costs of mycorrhizal associations. In:Lambers, H., Ribas-Carbo, M. (Eds.), Plant Respiration: From Cell to Ecosystem.Advances in Photosynthesis and Respiration, vol. 18. Springer, Dordrecht, TheNetherlands, pp. 207–224.
Campos, H., Cooper, M., Habben, J.E., Edmeades, G.O., Schussler, J.R., 2004. Improv-ing drought tolerance in maize: a view from industry. Field Crops Res. 90, 19–34.
Close, T.J., 1996. Dehydrins: emergence of a biochemical role of a family of plantdehydration proteins. Physiol. Plant. 97, 795–803.
Daigh, A., Savin, M., Purcell, L., 2006. Mycorrhizal infection in Roundup Ready rowcrops. Abstract. 2006 American Society of Agronomy-Crop Science Society of
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 29
America-Soil Science Society of America International Annual Meetings, India-napolis.
Dekankova, K., Luxova, M., Gasparıkova, O., Kolarovic, L., 2004. Response of maizeplants to water stress, Biologia. Bratislava 59, 151–155.
Dercon, G., Clymans, E., Diels, J., Merckx, R., Deckers, J., 2006. Differential 13Cisotopic discrimination in maize at varying water stress and at low to highnitrogen availability. Plant Soil 282, 313–326.
Edmeades, G.O., Banziger, M., Elings, A., Chapman, S.C., Ribaut, J.M., 1997a. Recentadvances in breeding for drought tolerance in maize. In: Kropff, M.J., Teng,P.S., Aggarwal, P.K., Bouman, B.A.M., Jones, J.W., vanLaar, H.H. (Eds.), Ap-plications of Systems Approaches at the Field Level—Proceedings of theSecond International Symposium on Systems Approaches for AgriculturalDevelopment. International Rice Research Institute, Los Banos, Philippines,pp. 63–78.
Edmeades, G.O., Bolanos, J., Chapman, S.C., 1997b. Value of secondary traits inselecting for drought tolerance in tropical maize. In: Edmeades, G.O., Banziger,M., Mickelson, H.R., Pena-Valdivia, C.B. (Eds.), Developing Drought and Low-NTolerant Maize. CIMMYT, El Batan, Mexico, pp. 222–234.
Edmeades, G.O., Bolanos, J., Chapman, S.C., Lafitte, H.R., Banziger, M., 1999. Selectionimproves drought tolerance in tropical maize populations. I. Gains in biomass,grain yield, and harvest index. Crop Sci. 39, 1306–1315.
Edmeades, G.O., McMaster, G.S., White, J.W., Campos, H., 2004. Genomics and thephysiologist: bridging the gap between genes and crop response. Field CropsRes. 90, 5–18.
Foyer, C.H., Valadier, M.H., Migge, A., Becker, T.W., 1998. Drought-induced effects onnitrate reductase activity and mRNA and on the coordination of nitrogen andcarbon metabolism in maize leaves. Plant Physiol. 117, 283–292.
Franken, P., Requena, N., 2001. Analysis of gene expression in arbuscular mycor-rhizas: new approaches and challenges. New Phytol. 150, 517–523.
Garcia, J.P., Wortmann, C.S., Mamo, M., Drijber, R., Tarkalson, D., 2007. One-timetillage of no-till: effects on nutrients, mycorrhizae, and phosphorus uptake.Agron. J. 99, 1093–1103.
Gavito, M.E., Miller, M.H., 1998. Changes in mycorrhiza development in maizeinduced by crop management practices. Plant Soil 198, 185–192.
George, E., Marschner, H., Jakobsen, I., 1995. Role of arbuscular mycorrhizal fungi inuptake of phosphorus and nitrogen from soil. Crit. Rev. Biotechnol. 15, 257–270.
Giuliani, S., Sanguineti, M.C., Tuberosa, R., Bellotti, M., Salvi, S., Landi, P., 2005. Root-ABA-1, a major constitutive QTL, affects maize root architecture and leaf ABAconcentration at different water regimes. J. Exp. Bot. 56, 3061–3070.
Graham, J.H., 2000. Assessing costs of arbuscular mycorrhizal symbiosis in agroe-cosystems. In: Podila, G.K., Douds, Jr., D.D. (Eds.), Current Advances in Mycor-rhizal Research. APS Press, St. Paul, MN, pp. 127–140.
Gupta, R., Mukerji, K.G., 2002. Techniques for the isolation of VAM/AM fungi in soil.In: Mukerji, K.G., Manoharachary, C., Chamola, B.P. (Eds.), Techniques inMycorrhizal Studies. Kluwer Academic Publishers, Dordrecht, Netherlands,pp. 201–216.
Gutierrez, R., San Miguel, M.Ch., Larque-Saavedra, R.A., 1997. Stomatal conductancein successive selection cycles of the drought tolerant maize population ‘Tux-peno sequıa’. In: Edmeades, G.O., Banziger, M., Mickelson, H.R., Pena-Valdivia,C.B. (Eds.), Developing Drought and Low-N Tolerant Maize. CIMMYT, El Batan,Mexico, pp. 212–215.
Hamel, C., Strullu, D.-G., 2006. Arbuscular mycorrhizal fungi in field crop produc-tion: potential and new direction. Can. J. Plant Sci. 86, 941–950.
Harrison, M.J., 1999. Molecular and cellular aspects of the arbuscular mycorrhizalsymbiosis. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50, 361–389.
Hetrick, B.A.D., Hetrick, J.A., Bloom, J., 1984. Interaction of mycorrhizal infection,phosphorus level, and moisture stress in growth of field corn. Can. J. Bot. 62,2267–2271.
Hetrick, B.A.D., Kitt, D.G., Wilson, G.T., 1987. Effects of drought stress on growthresponse in corn, sudan grass, and big bluestem to Glomus etunicatum. NewPhytol. 105, 403–410.
Hillel, D., Rosenzweig, C., 2002. Desertification in relation to climate variability andchange. Adv. Agron. 77, 1–38.
IPCC, 2001. Summary for Policymakers. In: Watson, R.T. and the Core Writing Team(Eds.), Climate Change 2001: Synthesis Report. Intergovernmental Panel onClimate Change, Geneva, Switzerland, pp. 1–34.
IPCC, 2007. Summary for Policymakers. In: Parry, M.L., Canziani, O.F., Palutikof, J.P.,van der Linden, P.J., Hanson, C.E. (Eds.), Climate Change 2007: Impacts, Adapta-tion and Vulnerability. Contribution of Working Group II to the Fourth Assess-ment Report of the Intergovernmental Panel on Climate Change. CambridgeUniversity Press, Cambridge, UK, pp. 7–22.
Ishii, S., Loynachan, T.E., 2004. Rapid and reliable DNA extraction techniques fromtrypan-blue-stained mycorrhizal roots: comparison of two methods. Mycor-rhiza 14, 271–275.
Jeffries, P., 1987. Use of mycorrhizae in agriculture. Crit. Rev. Biotechnol. 5,319–357.
Jensen, S.D., 1995. Genetic improvement of maize for drought tolerance. In: Jewell,D.C., Waddington, S.R., Ransom, J.K., Pixley, K.V. (Eds.), Proceedings of theFourth Eastern and Southern African Regional Maize Conference, Harare, Zim-babwe. CIMMYT, El Batan, Mexico, pp. 67–75.
Johnson, N.C., Pfleger, F.L., 1992. Vesicular-arbuscular mycorrhizae and culturalstresses. In: Bethlenfalvay, J., Linderman, R.G. (Eds.), Mycorrhizae in Sustainable
Agriculture. American Society of Agronomy, Crop Science Society of America,and Soil Science Society of America, Madison, pp. 71–99.
Johnson, N.C., Pfleger, F.L., Crookston, R.K., Simmons, S.R., Copeland, P.J., 1991.Vesicular-arbuscular mycorrhizas respond to corn and soybean cropping his-tory. New Phytol. 117, 657–663.
Kabir, Z., 2005. Tillage or no-tillage: impact on mycorrhizae. Can. J. Plant Sci. 85,23–29.
Kabir, Z., O’Halloran, I.P., Widden, P., Hamel, C., 1998. Vertical distribution ofarbuscular mycorrhizal fungi under corn (Zea mays L.) in no-till and conven-tional tillage systems. Mycorrhiza 8, 53–55.
Kaeppler, S.M., Parke, J.L., Mueller, S.M., Senior, L., Stuber, C., Tracy, W.F., 2000.Variation among maize inbred lines and detection of quantitative trait loci forgrowth at low phosphorus and responsiveness to arbuscular mycorrhizal fungi.Crop Sci. 40, 358–364.
Kaldorf, M., Ludwig-Muller, J., 2000. AM fungi might affect the root morphology ofmaize by increasing indole-3-butyric acid biosynthesis. Physiol. Plant. 109,58–67.
Kaldorf, M., Schmelzer, E., Bothe, H., 1998. Expression of maize and fungal nitratereductase genes in arbuscular mycorrhiza. Mol. Plant-Microb. Interact. 11, 439–448.
Kamara, A.Y., Menkir, A., Ajala, S.O., Kureh, I., 2005. Performance of diverse maizegenotypes under nitrogen deficiency in the northern Guinea savanna of Nigeria.Exp. Agric. 41, 199–212.
Karasawa, T., Arihara, J., Kasahara, Y., 2000. Effects of previous crops on arbuscularmycorrhizal formation and growth of maize under various soil moisture con-ditions. Soil Sci. Plant Nutr. 46, 53–60.
Kathiresan, A., Lafitte, H.R., Chen, J., Mansueto, L., Bruskiewich, R., Bennett, J., 2006.Gene expression microarrays and their application in drought stress research.Field Crops Res. 97, 101–110.
Khalil, S., Loynachan, T.E., Tabatabai, M.A., 1994. Mycorrhizal dependency andnutrient uptake by improved and unimproved corn and soybean cultivars.Agron. J. 86, 949–958.
Koag, M.C., Fenton, R.D., Wilkens, S., Close, T.J., 2003. The binding of maize DHN1 tolipid vesicles. Gain of structure and lipid specificity. Plant Physiol. 131,309–316.
Kondo, M., Murty, M.V.R., Aragones, D.V., 2000. Characteristics of root growth andwater uptake from soil in upland rice and maize under water stress. Soil Sci.Plant Nutr. 46, 721–732.
Kothari, S.K., Marschner, H., George, E., 1990. Effect of VA mycorrhizal fungi andrhizosphere microorganisms on root and shoot morphology, growth and waterrelations in maize. New Phytol. 116, 303–311.
Kuchenbuch, R.O., Ingram, K.T., Buczko, U., 2006. Effects of decreasing soil watercontent on seminal and lateral roots of young maize plants. J. Plant Nutr. Soil Sci.169, 841–848.
Liu, A., Hamel, C., Hamilton, R.I., Ma, B.L., Smith, D.L., 2000a. Acquisition of Cu, Zn,Mn and Fe by mycorrhizal maize (Zea mays L.) grown in soil at different P andmicronutrient levels. Mycorrhiza 9, 331–336.
Liu, A., Hamel, C., Hamilton, R.I., Smith, D.L., 2000b. Mycorrhizae formation andnutrient uptake of new corn (Zea mays L.) hybrids with extreme canopy and leafarchitecture as influenced by soil N and P levels. Plant Soil 221, 157–166.
Liu, A., Hamel, C., Elmi, A.A., Zhang, T., Smith, D.L., 2003. Reduction of the availablephosphorus pool in field soils growing maize genotypes with extensive mycor-rhizal development. Can. J. Plant Sci. 83, 737–744.
Liu, A., Plenchette, C., Hamel, C., 2007. Soil nutrient and water providers: howarbuscular mycorrhizal mycelia support plant performance in a resource-limited world. In: Hamel, C., Plenchette, C. (Eds.), Mycorrhizae in Crop Produc-tion. Haworth Food & Agricultural Products Press, Binghamton, NY, pp. 37–66.
Logrono, M.L., Lothrop, J.E., 1997. Impact of drought and low nitrogen on maizeproduction in South Asia. In: Edmeades, G.O.,Banziger, M., Mickelson, H.R.,Pena-Valdivia, C.B. (Eds.),Developing Drought and Low-N Tolerant Maize. CIMMYT, ElBatan, Mexico, pp. 39–43.
Lorens, G.F., Bennett, J.M., Loggale, L.B., 1987. Differences in drought resistancebetween two corn hybrids. I. Water relations and root length density. Agron. J.79, 802–807.
Ludlow, M.M., Muchow, R.C., 1990. A critical evaluation of traits for improving cropyields in water-limited environments. Adv. Agron. 43, 107–153.
Malty, J.D., Siqueira, J.O., Moreira, F.M.D., 2006. Effects of glyphosate on soybeansymbiotic microorganisms, in culture media and in greenhouse. Pesq. Agropec.Bras. 41, 285–291.
Marschner, H., 1995. Mineral Nutrition of Higher Plants. Academic Press, San Diego.Marschner, H., Dell, B., 1994. Nutrient uptake in mycorrhizal symbiosis. Plant Soil
159, 89–102.Mickelson, S.M., Kaeppler, S.M., 2005. Evaluation of six mycorrhizal isolates for their
ability to promote growth of maize genotypes under phosphorus deficiency.Maydica 50, 137–146.
Miller, M.H., 2000. Arbuscular mycorrhizae and the phosphorus nutrition of maize:a review of Guelph studies. Can. J. Plant Sci. 80, 47–52.
Morgan, J.M., 1984. Osmoregulation and water stress in higher plants. Ann. Rev.Plant Physiol. 35, 299–319.
Muller, I., Hofner, W., 1991. Influence of the VA-mycorrhiza on P-uptake andrecovery potential of corn (Zea mays L.) under water stress conditions. Z.Pflanzenernaehr Bodenkd. 154, 321–323.
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–3130
Ncube, B., Dimes, J.P., Twomlow, S.J., Mupangwa, W., Giller, K.E., 2007. Raising theproductivity of smallholder farms under semi-arid conditions by use of smalldoses of manure and nitrogen: a case of participatory research. Nutr. Cycl.Agroecosyst. 77, 53–67.
Nelsen, C.E., Safir, G.R., 1982. Increased drought tolerance of mycorrhizal onionplants caused by improved phosphorus nutrition. Planta 154, 407–413.
Oehl, F., Sieverding, E., Ineichen, K., Riss, E., Boller, T., Wiemken, A., 2005. Commu-nity structure of arbuscular mycorrhizal fungi at different soil depths inextensively and intensively managed agroecosystems. New Phytol. 165,273–283.
Ogawa, A., Yamauchi, A., 2006a. Root osmotic adjustment under osmotic stress inmaize seedlings. 1. Transient change of growth and water relations in roots inresponse to osmotic stress. Plant Prod. Sci. 9, 27–38.
Ogawa, A., Yamauchi, A., 2006b. Root osmotic adjustment under osmotic stress inmaize seedlings. 2. Mode of accumulation of several solutes for osmoticadjustment in the root. Plant Prod. Sci. 9, 39–46.
Oikeh, S.O., Kling, J.G., Horst, W.J., Chude, V.O., Carsky, R.J., 1999. Growth anddistribution of maize roots under nitrogen fertilization in plinthite soil. FieldCrops Res. 62, 1–13.
Parniske, M., 2004. Molecular genetics of the arbuscular mycorrhizal symbiosis.Curr. Opin. Plant Biol. 7, 414–421.
Passioura, J.B., 1977. Grain yield, harvest index, and water use of wheat. J. Aust. Inst.Agric. Sci. 43, 117–120.
Passioura, J.B., 1996. Drought and drought tolerance. Plant Growth Regul. 20, 79–83.Paszkowski, U., Boller, T., 2002. The growth defect of lrt1, a maize mutant lacking
lateral roots, can be complemented by symbiotic fungi or high phosphatenutrition. Planta 214, 584–590.
Pena-Valdivia, C.B., Raya, J.C., Perales, H.R., Lotina, R.B., Merchant, H.R., 1997. Bio-chemical characterization of drought tolerant and susceptible young maizeplants under well-watered conditions. In: Edmeades, G.O., Banziger, M., Mick-elson, H.R., Pena-Valdivia, C.B. (Eds.), Developing Drought and Low-N TolerantMaize. CIMMYT, El Batan, Mexico, pp. 163–168.
Picone, C., 2003. Managing mycorrhizae for sustainable agriculture in the tropics.In: Vandermeer, J.H. (Ed.), Tropical Agroecosystems. CRC Press, Boca Raton, FL,pp. 95–132.
Pinter, L., Kalman, L., Palfi, G., 1978. Determination of drought resistance indifferent hybrids of maize by field trials and biochemical tests. Maydica 23,121–127.
Porcel, R., Azcon, R., Ruiz-Lozano, J.M., 2005a. Evaluation of the role of genesencoding for dehydrin proteins (LEA D-11) during drought stress in arbuscularmycorrhizal Glycine max and Lactuca sativa plants. J. Exp. Bot. 56, 1933–1942.
Porcel, R., Gomez, M., Kaldenhoff, R., Ruiz-Lozano, J.M., 2005b. Impairment ofNtAQP1 gene expression in tobacco plants does not affect root colonisationpattern by arbuscular mycorrhizal fungi but decreases their symbiotic effi-ciency under drought. Mycorrhiza 15, 417–423.
Porcel, R., Aroca, R., Azcon, R., Ruiz-Lozano, J.M., 2006. PIP aquaporin gene expres-sion in arbuscular mycorrhizal Glycine max and Lactuca sativa plants in relationto drought stress tolerance. Plant Mol. Biol. 60, 389–404.
Prystupa, P., Lemcoff, J.H., 1998. Increased root density of maize crops underdrought in a Typic Argiudoll. Maydica 43, 271–276.
Rajcan, I., Tollenaar, M., 1999. Source:sink ratio and leaf senescence in maize: I Drymatter accumulation and partitioning during grain filling. Field Crops Res. 60,245–253.
Ramakrishnan, B., Johri, B.N., Gupta, R.K., 1988a. Effect of vesicular arbuscularmycorrhizal fungus on photosynthesis and photorespiration in water stressedmaize. Photosynthetica 22, 443–447.
Ramakrishnan, B., Johri, B.N., Gupta, R.K., 1988b. Influence of the VAM fungusGlomus caledonius on free proline accumulation in water-stressed maize. Curr.Sci. 57, 1082–1084.
Ravnskov, S., Wu, Y., Graham, J.H., 2003. Arbuscular mycorrhizal fungi differentiallyaffect expression of genes coding for sucrose synthases in maize roots. NewPhytol. 157, 539–545.
Ryan, M.H., Graham, J.H., 2002. Is there a role for arbuscular mycorrhizal fungi inproduction agriculture? Plant Soil 244, 263–271.
Sage, R.F., Pearcy, R.W., 1987. The nitrogen use efficiency of C3 and C4 plants. II. Leafnitrogen effects on the gas exchange characteristics of Chenopodium album (L.)and Amaranthus retroflexus (L.). Plant Physiol. 84, 959–963.
Saito, M., Marumoto, T., 2002. Inoculation with arbuscular mycorrhizal fungi: thestatus quo in Japan and future prospects. Plant Soil 244, 273–279.
Sanchez, P.A., 2002. Ecology–soil fertility and hunger in Africa. Science 295, 2019–2020.
Sanchez-Dıaz, M., Honrubia, M., 1994. Water relations and alleviation of droughtstress in mycorrhizal plants. In: Gianinazzi, S., Schuepp, H. (Eds.), Impact ofArbuscular Mycorrhizas on Sustainable Agriculture and Natural Ecosystems.Birkhauser Verlag, Basel, Switzerland, pp. 167–178.
Schellenbaum, L., Muller, J., Boller, T., Wiemken, A., Schuepp, H., 1998. Effects ofdrought on non-mycorrhizal and mycorrhizal maize: changes in the pools ofnon-structural carbohydrates, in the activities of invertase and trehalase, and inthe pools of amino acids and imino acids. New Phytol. 138, 59–66.
Schussler, J.R., Westgate, M.E., 1995. Assimilate flux determines kernel set at lowwater potential in maize. Crop Sci. 35, 1074–1080.
Setter, T.L., 1997. Role of the phytohormone ABA in drought tolerance: potentialutility as a selection tool. In: Edmeades, G.O., Banziger, M., Mickelson, H.R.,Pena-Valdivia, C.B. (Eds.),Developing Drought and Low-N Tolerant Maize. CIMMYT, ElBatan, Mexico, pp. 142–150.
Sharp, R.E., Davies, W.J., 1985. Root growth and water uptake by maize plants indrying soil. J. Exp. Bot. 36, 1441–1456.
Sharrock, R.A., Sinclair, F.L., Gliddon, C., Rao, I.M., Barrios, E., Mustonen, P.J.,Smithson, P., Jones, D.L., Godbold, D.L., 2004. A global assessment using PCRtechniques of mycorrhizal fungal populations colonising Tithonia diversifolia.Mycorrhiza 14, 103–109.
Simpson, D., Daft, M.J., 1990. Interactions between water-stress and differentmycorrhizal inocula on plant growth and mycorrhizal development in maizeand sorghum. Plant Soil 121, 179–186.
Smith, S.E., Smith, F.A., Jakobsen, I., 2003. Mycorrhizal fungi can dominatephosphate supply to plant irrespective of growth responses. Plant Physiol.133, 16–20.
St-Arnaud, M., Vujanovic, V., 2007. Effects of the arbuscular mycorrhizal symbiosison plant diseases and pests. In: Hamel, C., Plenchette, C. (Eds.), Mycorrhizae inCrop Production. Haworth Food & Agricultural Products Press, Binghamton, NY,pp. 67–122.
Studer, C., Hu, Y., Schmidhalter, U., 2007. Evaluation of the differential osmoticadjustments between roots and leaves of maize seedlings with single orcombined NPK-nutrient supply. Funct. Plant Biol. 34, 228–236.
Subramanian, K.S., Charest, C., 1995. Influence of arbuscular mycorrhizae on themetabolism of maize under drought stress. Mycorrhiza 5, 273–278.
Subramanian, K.S., Charest, C., 1997. Nutritional, growth, and reproductiveresponses of maize (Zea mays L.) to arbuscular mycorrhizal inoculation duringand after drought stress at tasselling. Mycorrhiza 7, 25–32.
Subramanian, K.S., Charest, C., 1998. Arbuscular mycorrhizae and nitrogen assim-ilation in maize after drought and recovery. Physiologia Plant. 102, 285–296.
Subramanian, K.S., Charest, C., 1999. Acquisition of N by external hyphae of anarbuscular mycorrhizal fungus and its impact on physiological responsesin maize under drought-stressed and well-watered conditions. Mycorrhiza 9,69–75.
Subramanian, K.S., Charest, C., Dwyer, L.M., Hamilton, R.I., 1995. Arbuscular mycor-rhizas and water relations in maize under drought stress at tasselling. NewPhytol. 129, 643–650.
Subramanian, K.S., Charest, C., Dwyer, L.M., Hamilton, R.I., 1997. Effects of arbus-cular mycorrhizae on leaf water potential, sugar content, and P content duringdrought and recovery of maize. Can. J. Bot. 75, 1582–1591.
Sylvia, D.E., Hammond, L.C., Bennet, J.M., Hass, J.H., Linda, S.B., 1993. Fieldresponse of maize to a VAM fungus and water management. Agron. J. 85,193–198.
Tanaka, T., Yano, K., 2005. Nitrogen delivery to maize via mycorrhizal hyphaedepends on the form of N supplied. Plant Cell Env. 28, 1247–1254.
Tobar, R.M., Azcon, R., Barea, J.M., 1994a. Improved nitrogen uptake and transportfrom 15N-labeled nitrate by external hyphae of arbuscular mycorrhizae underwater-stressed conditions. New Phytol. 126, 119–122.
Tobar, R.M., Azcon, R., Barea, J.M., 1994b. The improvement of plant N acquisitionfrom an ammonium-treated, drought-stressed soil by the fungal symbiont inarbuscular mycorrhizae. Mycorrhiza 4, 105–108.
Toth, R., Toth, D., Starke, D., Smith, D.R., 1990. Vesicular-arbuscular mycorrhizalcolonization in Zea mays affected by breeding for resistance to fungal patho-gens. Can. J. Bot. 68, 1039–1044.
Tuberosa, R., Salvi, S., Sanguineti, M.C., Landi, P., Maccaferri, M., Conti, S., 2002.Mapping QTLs regulating morpho-physiological traits and yield: case studies,shortcomings and perspectives in drought-stressed maize. Ann. Bot. 89,941–963.
Wassom, J.J., Reese, R.N., Schumacher, T.E., Wicks III, Z.W., 2000. Proline accumula-tion in response to dehydration and diurnal hydration cycles varies amongmaize genotypes. Maydica 45, 335–343.
Westgate, M.E., 1997. Physiology of flowering in maize: identifying avenues toimprove kernel set during drought. In: Edmeades, G.O., Banziger, M., Mickel-son, H.R., Pena-Valdivia, C.B. (Eds.), Developing Drought and Low-N TolerantMaize. CIMMYT, El Batan, Mexico, pp. 136–141.
Wright, S.F., 2005. Management of arbuscular mycorrhizal fungi. In: Zobel, R.W.,Wright, S.F. (Eds.), Roots and Soil Management: Interactions between Roots andthe Soil. American Society of Agronomy, Crop Science Society of America, SoilScience Society of America, Madison, WI, pp. 183–197.
C.R. Boomsma, T.J. Vyn / Field Crops Research 108 (2008) 14–31 31
Xu, D., Duan, X., Wang, B., Hong, B., Ho, T.H.D., Wu, R., 1996. Expression of a lateembryogenesis abundant protein gene, HVA1, from barley confers tolerance towater deficit and salt stress in transgenic rice. Plant Physiol. 110, 249–257.
Yano, K., Yamauchi, A., Kono, T., 1996. Localized alteration in lateral root develop-ment in roots colonized by an arbuscular mycorrhizal fungus. Mycorrhiza 6,409–415.
Zhang, J., Jia, W., Yang, J., Ismail, A.M., 2006. Role of ABA in integrating plantresponses to drought and salt stresses. Field Crops Res. 97, 111–119.
Zinselmeier, C., Westgate, M.E., Schussler, J.R., Jones, R.J., 1995. Low water potentialdisrupts carbohydrate metabolism in maize (Zea mays L.) ovaries. Plant Physiol.107, 385–391.
Zinselmeier, C., Sun, Y., Helentjaris, T., Beatty, M., Yang, S., Smith, H., Habben, J.,2002. The use of gene expression profiling to dissect the stress sensitivity ofreproductive development in maize. Field Crops Res. 75, 111–121.