INFECTION AND IMMUNITY, Jan. 1993, p. 124-134 0019-9567/93/010124-11$02.00/0 Copyright X) 1993, American Society for Microbiology Effects of Cytokines on Intracellular Growth of Brucella abortus XIAOSUI JIANG AND CYNTHIA L. BALDWIN* Department of Microbiology, The Ohio State University, Columbus, Ohio 43210-1292 Received 2 July 1992/Accepted 28 October 1992 Interleukin la (IL-la), IL-2, IL-4, IL-6, gamma interferon (IFN-y), tumor necrosis factor alpha (TNF-c), and granulocyte macrophage colony-stimulating factor (GM-CSF) were tested for their abilities to alter the growth of Brucella abortus in BALB/c J774A.1 murine macrophages. IL-la, IL-4, IL-6, tumor necrosis factor alpha, and granulocyte macrophage-colony-stimulating factor had no consistent or significant effect on the growth of the avirulent B. abortus strain 19. In contrast, the addition of either IFN-y or IL-2 at 100 U/ml to the macrophage cultures resulted in a significant reduction in the number of intracellular bacteria that was not attributable to decreased infection rates. With IL-2, the reduction was most often apparent only during the first 24 h after infection, while inhibition with IFN-y was apparent throughout the culture period of 48 h. The addition of either IL-2 or IFN-y to macrophage cultures also resulted in reduced intracellular CFU of the virulent B. abortus strain 2308 and the attenuated rough mutant B. abortus strain RB51. Inhibition of intracellular growth was not augmented by combinations of cytokines. Additional studies with IFN-'y and IL-2 indicated that they could mediate the inhibition of intracellular growth of B. abortus in resident and thioglycolate broth-induced BALB/c peritoneal macrophages and in splenic macrophages. IFN-y also inhibited bacterial growth when added after infection of the macrophages, although the magnitude of the antibrucellae effects was less than that when it was added before infection. Furthermore, the maximal inhibitory effect was sustained only when IFN-y remained in the cultures after infection of the macrophages. Brucella are gram-negative, facultative intracellular bac- teria which replicate in host mononuclear phagocytes, caus- ing chronic infections in humans as well as in agricultural animals, particularly cattle, sheep, goats, and pigs (32, 47). Infections in humans are almost exclusively due to zoonosis, either through direct contact with infected animals or from contaminated dairy products (33). Thus, the control of br.;cellosis in animals is necessary for its control in humans. Inbred strains of mice which differ in their capacities to prevent chronic infections with Brucella abortus have been previously identified and employed to evaluate the nature of innate resistali2e as well as that of acquired immunity (21, 28). When B. abortus 19, an attenuated vaccine strain, is used to infect BALB/c mice, the mice clear the infection (21, 28), whereas when the mice are infected with B. abortus 2308, a virulent field isolate, a chronic infection is estab- lished (28). This pattern of resistance and susceptibility to virulent and avirulent strains of B. abortus in BALB/c mice is similar to that observed following infection of cattle, making them a good model for bovine brucellosis (28). By using the murine model, it has been previously estab- lished that acquired immunity to B. abortus is mediated both by antibodies, predominantly to lipopolysaccharide (LPS) molecules (2, 28), and by CD4 and CD8 T cells (1, 35). An adoptive transfer of either population of T cells to naive mice results in similar levels of protection (1). While the mecha- nisms of T-cell-mediated immunity have not been eluci- dated, it has been previously demonstrated with both bovine (19) and guinea pig (7) macrophages that virulent strains of B. abortus have increased survival and growth in macro- phages compared with those of avirulent strains or rough mutants. Cell-mediated immunity to brucellosis, therefore, is likely to include the production of cytokines which acti- * Corresponding author. vate macrophages for antibrucella activities. To elucidate the role of cytokines in protective, cell-mediated immunity to B. abortus, we investigated the abilities of a panel of seven cytokines to either inhibit or facilitate the intracellular survival and growth of B. abortus in macrophages. The literature concerning macrophage activation and in- duction of antimicrobial mechanisms by cytokines is com- plex, partially as a result of the multiple pathways used for microbial killing. Gamma interferon (IFN-y) is capable of upregulating three major pathways described for microbial killing, i.e., the production of reactive oxygen intermediates (31), the production of nitrogen intermediates (44), and the induction of indolamine 2,3-dioxygenase to catabolize tryp- tophan (see reference 8), and it has been shown to induce antimicrobial activities against a variety of intracellular parasites, including Mycobacterium bovis (16), Mycobacte- rium tuberculosis (16, 41), Leishmania major (15, 17, 34), and Listeria monocytogenes (38). The abilities of other cytokines to activate macrophage antimicrobial activities are more variable. Tumor necrosis factor (TNF) can elevate H202 production and nitrite release from IFN-y-phorbol myristate acetate-stimulated macrophages (14), and TNF- treated macrophages inhibit the survival or growth of Myco- bacterium lepraemurium (10) and Mycobacterium avium (5, 12). TNF also synergizes with IFN--y to activate macro- phages to kill L. major (6, 26). Granulocyte macrophage colony-stimulating factor (GM-CSF) increases the produc- tion of H202 (14) by macrophages and activates macro- phages to inhibit the growth of Trypanosoma cruzi (39) and to kill Leishmania donovani (46). GM-CSF also synergizes with IFN-y to activate macrophages for resistance to infec- tion by L. major (3). Interleukin 2 (IL-2) synergizes with TNF or LPS to stimulate production of reactive oxygen intermediates by macrophages (5, 45) and, in conjunction with IFN-,y, activates macrophages to inhibit infection by L. major (3). IL-4 synergizes with IFN-,y to activate macro- 124 Vol. 61, No. 1 on February 25, 2020 by guest http://iai.asm.org/ Downloaded from
Transcript
INFECTION AND IMMUNITY, Jan. 1993, p. 124-1340019-9567/93/010124-11$02.00/0Copyright X) 1993, American Society for Microbiology
Effects of Cytokines on Intracellular Growth ofBrucella abortus
XIAOSUI JIANG AND CYNTHIA L. BALDWIN*Department of Microbiology, The Ohio State University, Columbus, Ohio 43210-1292
Received 2 July 1992/Accepted 28 October 1992
Interleukin la (IL-la), IL-2, IL-4, IL-6, gamma interferon (IFN-y), tumor necrosis factor alpha (TNF-c),and granulocyte macrophage colony-stimulating factor (GM-CSF) were tested for their abilities to alter thegrowth of Brucella abortus in BALB/c J774A.1 murine macrophages. IL-la, IL-4, IL-6, tumor necrosis factoralpha, and granulocyte macrophage-colony-stimulating factor had no consistent or significant effect on thegrowth of the avirulent B. abortus strain 19. In contrast, the addition of either IFN-y or IL-2 at 100 U/ml tothe macrophage cultures resulted in a significant reduction in the number of intracellular bacteria that was notattributable to decreased infection rates. With IL-2, the reduction was most often apparent only during the first24 h after infection, while inhibition with IFN-y was apparent throughout the culture period of 48 h. Theaddition of either IL-2 or IFN-y to macrophage cultures also resulted in reduced intracellular CFU of thevirulent B. abortus strain 2308 and the attenuated rough mutant B. abortus strain RB51. Inhibition ofintracellular growth was not augmented by combinations of cytokines. Additional studies with IFN-'y and IL-2indicated that they could mediate the inhibition of intracellular growth of B. abortus in resident andthioglycolate broth-induced BALB/c peritoneal macrophages and in splenic macrophages. IFN-y also inhibitedbacterial growth when added after infection of the macrophages, although the magnitude of the antibrucellaeeffects was less than that when it was added before infection. Furthermore, the maximal inhibitory effect wassustained only when IFN-y remained in the cultures after infection of the macrophages.
Brucella are gram-negative, facultative intracellular bac-teria which replicate in host mononuclear phagocytes, caus-ing chronic infections in humans as well as in agriculturalanimals, particularly cattle, sheep, goats, and pigs (32, 47).Infections in humans are almost exclusively due to zoonosis,either through direct contact with infected animals or fromcontaminated dairy products (33). Thus, the control ofbr.;cellosis in animals is necessary for its control in humans.Inbred strains of mice which differ in their capacities toprevent chronic infections with Brucella abortus have beenpreviously identified and employed to evaluate the nature ofinnate resistali2e as well as that of acquired immunity (21,28). When B. abortus 19, an attenuated vaccine strain, isused to infect BALB/c mice, the mice clear the infection (21,28), whereas when the mice are infected with B. abortus2308, a virulent field isolate, a chronic infection is estab-lished (28). This pattern of resistance and susceptibility tovirulent and avirulent strains of B. abortus in BALB/c miceis similar to that observed following infection of cattle,making them a good model for bovine brucellosis (28).By using the murine model, it has been previously estab-
lished that acquired immunity to B. abortus is mediated bothby antibodies, predominantly to lipopolysaccharide (LPS)molecules (2, 28), and by CD4 and CD8 T cells (1, 35). Anadoptive transfer of either population ofT cells to naive miceresults in similar levels of protection (1). While the mecha-nisms of T-cell-mediated immunity have not been eluci-dated, it has been previously demonstrated with both bovine(19) and guinea pig (7) macrophages that virulent strains ofB. abortus have increased survival and growth in macro-
phages compared with those of avirulent strains or roughmutants. Cell-mediated immunity to brucellosis, therefore,is likely to include the production of cytokines which acti-
* Corresponding author.
vate macrophages for antibrucella activities. To elucidatethe role of cytokines in protective, cell-mediated immunityto B. abortus, we investigated the abilities of a panel ofseven cytokines to either inhibit or facilitate the intracellularsurvival and growth of B. abortus in macrophages.The literature concerning macrophage activation and in-
duction of antimicrobial mechanisms by cytokines is com-plex, partially as a result of the multiple pathways used formicrobial killing. Gamma interferon (IFN-y) is capable ofupregulating three major pathways described for microbialkilling, i.e., the production of reactive oxygen intermediates(31), the production of nitrogen intermediates (44), and theinduction of indolamine 2,3-dioxygenase to catabolize tryp-tophan (see reference 8), and it has been shown to induceantimicrobial activities against a variety of intracellularparasites, including Mycobacterium bovis (16), Mycobacte-rium tuberculosis (16, 41), Leishmania major (15, 17, 34),and Listeria monocytogenes (38). The abilities of othercytokines to activate macrophage antimicrobial activities aremore variable. Tumor necrosis factor (TNF) can elevateH202 production and nitrite release from IFN-y-phorbolmyristate acetate-stimulated macrophages (14), and TNF-treated macrophages inhibit the survival or growth ofMyco-bacterium lepraemurium (10) and Mycobacterium avium (5,12). TNF also synergizes with IFN--y to activate macro-phages to kill L. major (6, 26). Granulocyte macrophagecolony-stimulating factor (GM-CSF) increases the produc-tion of H202 (14) by macrophages and activates macro-phages to inhibit the growth of Trypanosoma cruzi (39) andto kill Leishmania donovani (46). GM-CSF also synergizeswith IFN-y to activate macrophages for resistance to infec-tion by L. major (3). Interleukin 2 (IL-2) synergizes withTNF or LPS to stimulate production of reactive oxygenintermediates by macrophages (5, 45) and, in conjunctionwith IFN-,y, activates macrophages to inhibit infection by L.major (3). IL-4 synergizes with IFN-,y to activate macro-
phages to resist infection by L. major (3). IL-1 plays a role inmacrophage activation (9, 37) and, when administered tomice, prevents chronic infection by B. abornus (48). Incontrast, some cytokines, namely, IL-1, IL-3, IL-6, andM-CSF, have been shown previously to facilitate the growthof M. avium in human macrophages (11, 12, 43). The studiesreported in this article indicate that of the seven cytokinestested, only IFN--y and IL-2 altered the intracellular growthof B. abortus. Both mediated brucellastatic effects.
MATERIALS AND METHODS
Macrophages. The J774A.1 macrophage cell line, origi-nally obtained from the American Type Culture Collection,was kindly provided by F. Zavala. The cells were cultured inflasks in complete tissue culture medium (c-RPMI) consist-ing of RPMI 1640 (Whittaker M. A. Bioproducts, Walkers-ville, Md.) with 10% heat-inactivated fetal bovine serum(Hyclone Laboratories, Logan, Utah), 5 x 10'5 M 2-mer-captoethanol (Sigma, St. Louis, Mo.), and 50,ug of L-glu-tamine per ml (GIBCO Life Technologies, Grand Island,N.Y.).
Eight- to 12-week-old female virus-free BALB/c mice(Harlan Sprague-Dawley, Indianapolis, Ind.) were used asthe source of peritoneal and splenic macrophages. Micewere maintained in barrier housing with filtered inflow air ina restricted-access room. Employees who handled micewore gloves, masks, coats, and shoe covers to maintain theintegrity of the facility. To obtain peritoneal macrophages,mice were injected intraperitoneally with 3 ml of 4% thio-glycolate broth (BBL Microbiology Systems, Cockeysville,Md.). Three to 5 days later, mice were sacrificed by spinalcord dislocation, and their peritoneal cavities were openedon the midline. Cold RPMI was added to the cavity, and cellswere withdrawn with a 10-ml-volume syringe. To obtainsplenic macrophages, spleens were perfused with phos-phate-buffered saline (PBS) (Whittaker) and disrupted with asyringe plunger. The cell suspension was layered overFicoll-Paque (Pharmacia, Uppsala, Sweden) and centrifugedat 800 x g for 15 min. The leukocyte layer was collected, andthe cells were washed twice before being suspended inc-RPMI and plated onto petri dishes (150 by 15 mm) (Amer-ican Scientific Products, McGaw Park, Ill.). After beingincubated at 37°C for 2 h, nonadherent cells were removedfrom the plate and splenic macrophages were collected byscraping with a rubber policeman.
Bacteria. B. abortus 19 is a semirough mutant, strain 2308is a virulent field strain, and strain RB51 is a rough mutant ofstrain 2308 developed by G. Schurig et al. (42). Bacterialstrains were provided by A. J. Winter. All bacterial strainswere cultured in 2.8% brucella broth (Difco, Detroit, Mich.)at 37°C for 3 days and harvested by centrifugation at 5,800 xg for 20 min. Aliquots were stored in brucella broth with 10%glycerol in liquid nitrogen. To infect macrophages, B. abor-tus stocks were thawed and 1.5 x 109 cells were inoculatedinto 5 ml of broth and cultured at 37°C for 17 h so that cellswould be in the exponential growth phase when used.Preliminary experiments indicated that this was importantfor optimal intracellular growth. The concentration of bac-teria was determined by measuring culture turbidity at 550nm with a spectrophotometer and reading from a standardcurve. Serial dilutions of a sample were plated on 3.75%blood agar (Schaedler; Difco) plates, and bacterial CFUwere determined after 3 days to confirm the CFU estimatedby determining optical densities. B. abortus cells werewashed, suspended to 108/ml in RPMI 1640, and opsonized
with antibodies by being incubated with a subagglutinatingconcentration of inactivated B. abotus-immune bovine se-rum at room temperature for 30 min. After being incubated,the bacterial cells were washed and resuspended in c-RPMI.
Reagents. Human recombinant IL-la (HrIL-la), HrIL-6,HrTNF-a, and mouse recombinant IFN--y (MrIFN-y) werepurchased from Boehringer Mannheim (Indianapolis, Ind.).MrIL-2, MrIL-4, and MrGM-CSF were purchased fromGenzyme (Cambridge, Mass.). The cytokines were stored inaliquots at -70°C as per the manufacturer's instructions anddiluted with c-RPMI for use in the assays. All humanrecombinant cytokines used are known to have activity onmurine cells.
Infection of macrophages with B. abortus and addition ofcytokines to cell cultures. Macrophages were infected in amanner similar to that described by Halling et al. (18).Except in the experiment described in the section "Incuba-tion times and inhibition by IFN--y" below, murine macro-phages in c-RPMI were cultured in 96-well flat-bottom plates(Costar, Cambridge, Mass.) at 1.67 x 104 cells per well.Triplicate macrophage cultures were established for all treat-ments. Unless otherwise indicated, macrophages seeded atthis concentration were incubated with cytokines for 24 hbefore being infected. Following this, 20 ,ul of antibody-opsonized bacteria, suspended to the appropriate concentra-tion in c-RPMI, was added to each well such that the ratio ofbacteria to macrophages was 100:1. Preliminary experimentswhich evaluated ratios of bacteria to macrophages from300:1 to 3:1 showed that this ratio resulted in a satisfactoryinfection rate after a 2-h infection period when the protocoldescribed below was used. The microwell plates containingmacrophages and bacteria were centrifuged at 2,800 x g atroom temperature for 10 min to increase the attachment of B.abortus to macrophages and then incubated at 37°C for 1 hand 50 min. To ensure that macrophages which became lessadherent after infection or cytokine treatments were not lost,macrophage cultures were centrifuged at 2,800 x g betweentreatments which involved the removal of supematant.
Following the 2-h infection period, the majority of extra-cellular bacteria were removed by washing the macrophagecultures twice with medium containing antibiotics. To dothis, 0.15 ml of RPMI 1640 containing 120 ,ug of gentamicinper ml (GIBCO) and 71.5 ,ug of streptomycin sulfate per ml(GIBCO) was added to each well by allowing the medium torun down the side of the well. Since B. abortus is afacultative intracellular bacteria and since our initial exper-iments indicated that it proliferated in c-RPMI, it wasnecessary to add antibiotics to the macrophage cultures. Anumber of trials were performed to determine the concen-tration of antibiotics and the time required to kill extracel-lular brucellae. We found a combination of gentamicin andstreptomycin sulfate which killed extracellular brucellae in 1h. To ensure that extracellular brucellae were killed follow-ing this treatment, at 1, 3, 24, and 48 h after the addition ofantibiotics, 150 tl of supernatant was collected from eachinfected macrophage culture following centrifugation of themicrotiter plates at 200 x g to pellet macrophages but notbacteria (determined in trials ascertaining the differentialpelleting forces required for macrophages and bacteria). Theabove supernatants were then centrifuged at 12,000 x g in aminifuge to pellet bacteria. The pelleted bacteria werewashed with antibiotic-free medium and plated on agarplates to determine CFU. The supernatants of macrophagecultures contained 0 to 3 CFU (less than 0.01% of theintracellular CFU measured at that time). Microscopic ob-servations indicated that macrophages remain adherent dur-
ing this gentle washing procedure. Extracellular killed bac-teria were removed from the cultures to minimize theirpotentially confounding effect on the macrophage activationwith cytokines. Our concern for the effect of extracellularbacteria arises from the fact that B. abortus contains LPS,and Enterobacteriaceae LPS, at least, is a potent trigger foractivated macrophages. To kill any residual bacteria whichwere not removed during the wash procedure and anyadditional extracellular bacteria which were released as aresult of macrophage deaths, macrophage cultures wereincubated overnight in c-RPMI with the same concentrationof antibiotics. Fresh cytokines were also added to themacrophage cultures following the wash procedure. Afterthe overnight incubation, the macrophage cultures to beassessed after 24 h (1 day) were treated as described below,whereas the macrophage cultures which were to be contin-ued for 2 or 4 days were washed at this time to remove thehigh concentration of antibiotics and were provided withc-RPMI containing fresh cytokines and 60 ,ug of gentamicinper ml. Microscopic evaluations of the macrophage cultureswere made to ensure that cells were not lost during the washprocedure. Initial experiments also evaluated the viabilitiesof J774A.1 and peritoneal macrophages, which were main-tained in these cultures, at various times after infection bytrypan blue exclusion of cells.
Microscopic evaluation of infected macrophages. To deter-mine intracellular infection of macrophages, 105 macro-phages per well were plated on round coverslips in 24-wellplates for light-microscopic evaluation. Following infectionof macrophages with B. abortus as described for the micro-well cultures and after incubation for various periods oftime, macrophages were washed twice with PBS, fixed withmethanol, and stained with Giemsa stain.Macrophages were plated in 24-well plates at 105 per well,
infected, and evaluated by transmission electron microscopyafter a 48-h incubation period. To prepare them for electronmicroscopy, macrophages were washed twice with RPMIand removed by gentle scraping with a rubber policeman.The cells were fixed by standard techniques with 1% glutar-aldehyde (Sigma) in 0.1 M phosphate buffer for 1 h at 4°C andthen embedded in 1% agarose. The cell-containing agarosewas cut into 1-mm3 cubes, postfixed with 1% osmiumtetroxide in phosphate buffer on ice for 1 h, and embedded inSpurr resin (Sigma). Thin sections were cut and viewed witha Philips electron microscope.
Incubation times and inhibition by IFN-'y. To determine theeffect of the addition of IFN-y to macrophage cultures fordiffering lengths of time prior to infection, 5 x 103 macro-phages per well were added to 96-well flat-bottom plates. Itwas necessary to reduce the number of macrophages perwell for this experiment because of the length of culturing (6days total), since J774A.1 is a transformed macrophage cellline which continues to grow in these cultures. Thus, lowerseeding densities at the time of infection were necessary toaccommodate the number of cells per well achieved by theend of the culture period. Macrophage cultures to be pre-treated with IFN--y for different lengths of time were allestablished 4 days before infection. At the appropriate timebefore infection, i.e., 96, 72, 48, or 24 h, IFN-y was added tothe cultures. Control cultures which never received IFN--ywere also established in parallel. A second experimentevaluated the effect of adding IFN--y to macrophages 2 or 24h after infection with B. abortus. In that experiment, mac-rophages were plated at the usual concentration, i.e., 1.67 x104 cells per well, 24 h before infection. IFN-y was added tocontrol cultures 24 h before, as well as after, infection and to
the principal cultures either 2 or 24 h after infection but notbefore. Control cultures which did not receive IFN--y at anytime were also established in parallel. In a third experiment,the effect of removing IFN-y from macrophage cultures wasevaluated. Macrophages were preincubated with IFN--y for24 h before infection, as described in the preceding sections.In one set of replicate cultures, IFN--y was added back afterinfection, while in another set it was not. Control cultureswhich never received IFN-y were established in parallel. Allcultures were infected simultaneously, and all other mani-pulations were the same as described above.
Combinations of cytokines. The J774A.1 macrophage cellline or thioglycolate-induced peritoneal macrophages wereincubated for 96 h with IFN-y at either 3 U/ml (suboptimalconcentration) or 100 U/ml (optimal concentration). Othercytokines, which were evaluated for their abilities to en-hance or diminish the inhibition induced by IFN--y, werethen added to the macrophage cultures, and the cultureswere incubated for a further 24 h before infection. Furtherprocedures were as described above, except that the com-binations of cytokines were added back after infection. Inexperiments which evaluated the effect of combining IL-2and TNF, the cytokines were added together 24 h beforeinfection as well as after infection.
Determination of the number of intracellular bacteria. Afterinfection for 1, 2, or 4 days, the adherent macrophages werewashed three times by adding 0.15 ml of PBS to the culturewells, removing it, and then adding 100 ,ul of 0.1% deoxy-cholate (Sigma) in PBS to each well in a manner similar tothat of a method described previously (18). After beingincubated at room temperature for 5 min, the cell lysatesfrom triplicate wells were pooled. Following lysis of themacrophages, the contents of the wells were resuspended,the replicate cultures pooled and immediately centrifuged at12,000 x g for 5 min in microcentrifuge tubes to pelletbrucellae, the supematant was discarded, and the bacteriawere resuspended in 0.5 ml of PBS. Preliminary observa-tions indicated that the number of brucellae recovered afterincubation of the bacteria in c-RPMI with 0.1% deoxy-cholate for 10 min was the same as that in the control media(CFU per culture were log1o 8.15 versus 8.13, respectively).Also, when macrophages were lysed by three rounds offreeze-thaw lysis and compared with those lysed by deoxy-cholate lysis, CFU per culture were log1o 5.54 versus 6.17,respectively. To quantitate the number of intracellular bac-teria, serial dilutions were made of the bacterial suspension,and aliquots were plated onto duplicate blood agar plates.Bacterial CFU were counted after 3 days of incubation at37°C with 5% CO2 in air, and the CFU were transformed tolog1o. The geometric mean and SD of CFU for each poolwere determined from the counts made on replicate platesover a range of dilutions.
Neutralization with antibody to IFN-y. To neutralize theeffect of IFN-y in the recombinant cytokine preparation,MrIFN-,y was preincubated with hamster anti-murine IFN--yantibodies, purchased from Genzyme (catalog no. 1222-00),at a ratio of 7.14 ng of antibody to 1 U of IFN-y at roomtemperature for 1 h (as per the manufacturer's instructions)before being added to macrophage cultures. The controlconsisted of MrIFN-,y, which had been similarly preincu-bated but without anti-IFN--y antibody. Both neutralized andcontrol IFN--y were used at 100 U/ml in this assay.
Statistical evaluations. Statistical analyses for experimentswhich were repeated more than five times were performedby the two-tailed Student t test (see IFN--y and IL-2 in Fig.3). For experiments which were repeated fewer than six
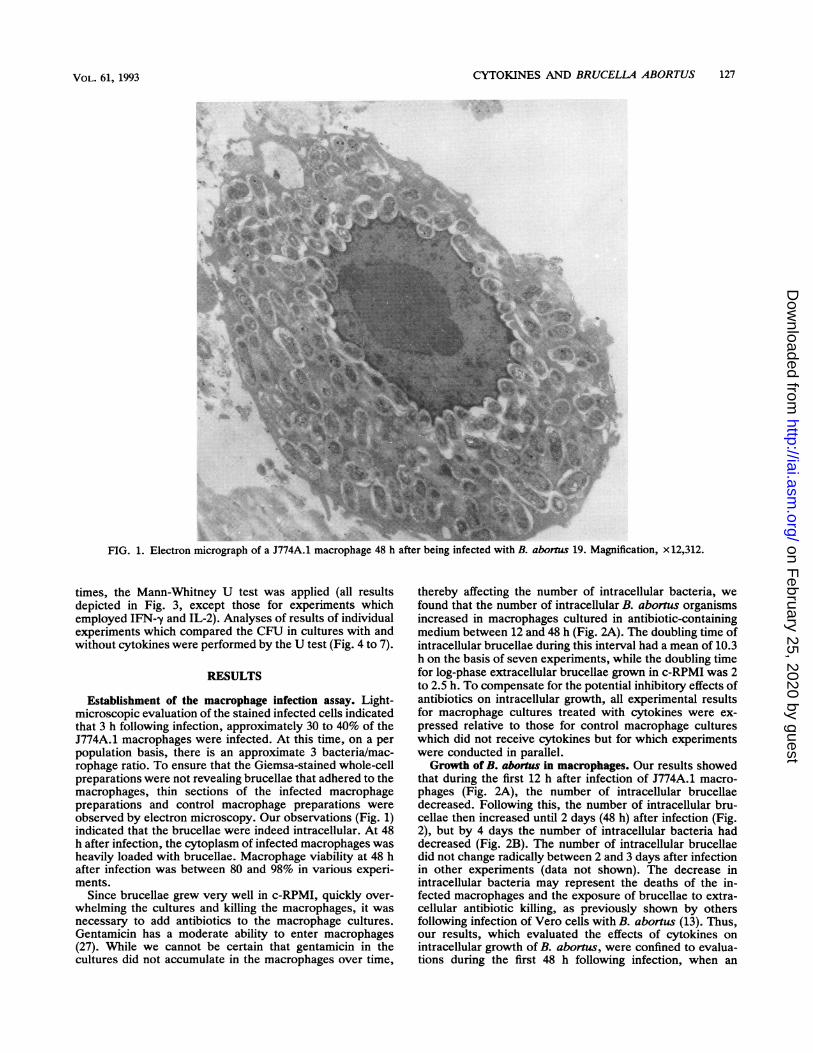
FIG. 1. Electron micrograph of a J774A.1 macrophage 48 h after being infected with B. abornus 19. Magnification, x 12,312.
times, the Mann-Whitney U test was applied (all resultsdepicted in Fig. 3, except those for experiments whichemployed IFN--y and IL-2). Analyses of results of individualexperiments which compared the CFU in cultures with andwithout cytokines were performed by the U test (Fig. 4 to 7).
RESULTS
Establishment of the macrophage infection assay. Light-microscopic evaluation of the stained infected cells indicatedthat 3 h following infection, approximately 30 to 40% of theJ774A.1 macrophages were infected. At this time, on a perpopulation basis, there is an approximate 3 bacteria/mac-rophage ratio. To ensure that the Giemsa-stained whole-cellpreparations were not revealing brucellae that adhered to themacrophages, thin sections of the infected macrophagepreparations and control macrophage preparations wereobserved by electron microscopy. Our observations (Fig. 1)indicated that the brucellae were indeed intracellular. At 48h after infection, the cytoplasm of infected macrophages washeavily loaded with brucellae. Macrophage viability at 48 hafter infection was between 80 and 98% in various experi-ments.
Since brucellae grew very well in c-RPMI, quickly over-whelming the cultures and killing the macrophages, it wasnecessary to add antibiotics to the macrophage cultures.Gentamicin has a moderate ability to enter macrophages(27). While we cannot be certain that gentamicin in thecultures did not accumulate in the macrophages over time,
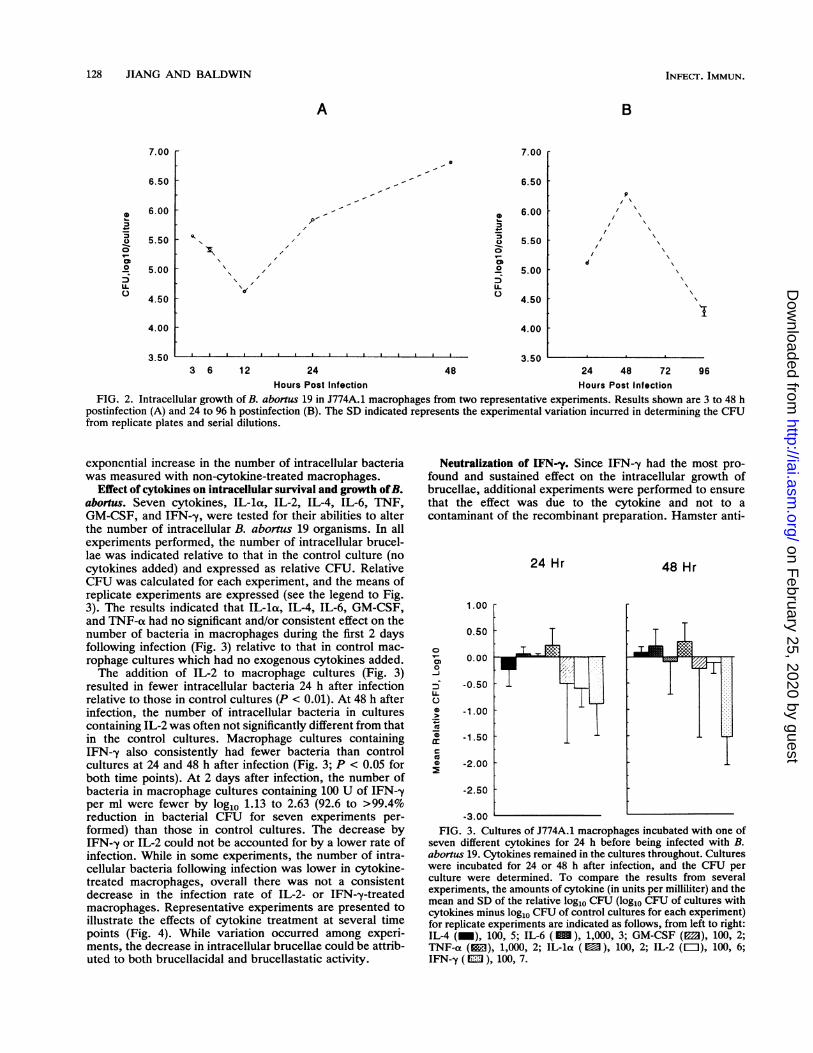
thereby affecting the number of intracellular bacteria, wefound that the number of intracellular B. abortus organismsincreased in macrophages cultured in antibiotic-containingmedium between 12 and 48 h (Fig. 2A). The doubling time ofintracellular brucellae during this interval had a mean of 10.3h on the basis of seven experiments, while the doubling timefor log-phase extracellular brucellae grown in c-RPMI was 2to 2.5 h. To compensate for the potential inhibitory effects ofantibiotics on intracellular growth, all experimental resultsfor macrophage cultures treated with cytokines were ex-pressed relative to those for control macrophage cultureswhich did not receive cytokines but for which experimentswere conducted in parallel.Growth of B. abortus in macrophages. Our results showed
that during the first 12 h after infection of J774A.1 macro-phages (Fig. 2A), the number of intracellular brucellaedecreased. Following this, the number of intracellular bru-cellae then increased until 2 days (48 h) after infection (Fig.2), but by 4 days the number of intracellular bacteria haddecreased (Fig. 2B). The number of intracellular brucellaedid not change radically between 2 and 3 days after infectionin other experiments (data not shown). The decrease inintracellular bacteria may represent the deaths of the in-fected macrophages and the exposure of brucellae to extra-cellular antibiotic killing, as previously shown by othersfollowing infection of Vero cells with B. abortus (13). Thus,our results, which evaluated the effects of cytokines onintracellular growth of B. abortus, were confined to evalua-tions during the first 48 h following infection, when an
Hours Post Infection Hours Post InfectionFIG. 2. Intracellular growth of B. abortus 19 in J774A.1 macrophages from two representative experiments. Results shown are 3 to 48 h
postinfection (A) and 24 to 96 h postinfection (B). The SD indicated represents the experimental variation incurred in determining the CFUfrom replicate plates and serial dilutions.
exponential increase in the number of intracellular bacteriawas measured with non-cytokine-treated macrophages.
Effect of cytokines on intracellular survival and growth ofB.abortus. Seven cytokines, IL-la, IL-2, IL-4, IL-6, TNF,GM-CSF, and IFN--y, were tested for their abilities to alterthe number of intracellular B. abortus 19 organisms. In allexperiments performed, the number of intracellular brucel-lae was indicated relative to that in the control culture (nocytokines added) and expressed as relative CFU. RelativeCFU was calculated for each experiment, and the means ofreplicate experiments are expressed (see the legend to Fig.3). The results indicated that IL-la, IL-4, IL-6, GM-CSF,and TNF-ot had no significant and/or consistent effect on thenumber of bacteria in macrophages during the first 2 daysfollowing infection (Fig. 3) relative to that in control mac-rophage cultures which had no exogenous cytokines added.The addition of IL-2 to macrophage cultures (Fig. 3)
resulted in fewer intracellular bacteria 24 h after infectionrelative to those in control cultures (P < 0.01). At 48 h afterinfection, the number of intracellular bacteria in culturescontaining IL-2 was often not significantly different from thatin the control cultures. Macrophage cultures containingIFN-y also consistently had fewer bacteria than controlcultures at 24 and 48 h after infection (Fig. 3; P < 0.05 forboth time points). At 2 days after infection, the number ofbacteria in macrophage cultures containing 100 U of IFN--yper ml were fewer by log1o 1.13 to 2.63 (92.6 to >99.4%reduction in bacterial CFU for seven experiments per-formed) than those in control cultures. The decrease byIFN-,y or IL-2 could not be accounted for by a lower rate ofinfection. While in some experiments, the number of intra-cellular bacteria following infection was lower in cytokine-treated macrophages, overall there was not a consistentdecrease in the infection rate of IL-2- or IFN--y-treatedmacrophages. Representative experiments are presented toillustrate the effects of cytokine treatment at several timepoints (Fig. 4). While variation occurred among experi-ments, the decrease in intracellular brucellae could be attrib-uted to both brucellacidal and brucellastatic activity.
Neutralization of IFN-y. Since IFN-y had the most pro-found and sustained effect on the intracellular growth ofbrucellae, additional experiments were performed to ensurethat the effect was due to the cytokine and not to acontaminant of the recombinant preparation. Hamster anti-
24 Hr
1.00
0.500
0-J
0
-
I0
0S
0.00
-0.50
-1.00
-1.50
-2.00
-2.50
48 Hr
-3.00FIG. 3. Cultures of J774A.1 macrophages incubated with one of
seven different cytokines for 24 h before being infected with B.abornus 19. Cytokines remained in the cultures throughout. Cultureswere incubated for 24 or 48 h after infection, and the CFU perculture were determined. To compare the results from severalexperiments, the amounts of cytokine (in units per milliliter) and themean and SD of the relative log1o CFU (log1o CFU of cultures withcytokines minus log1o CFU of control cultures for each experiment)for replicate experiments are indicated as follows, from left to right:IL-4 (_), 100, 5; IL-6 (), 1,000, 3; GM-CSF (E3), 100, 2;TNF-a (l), 1,000, 2; IL-la (E), 100, 2; IL-2 (=), 100, 6;IFN--y ( ml ), 100, 7.
FIG. 4. Macrophages were activated with cytokines (---) (A) (J774A.1 cell line with IFN--y) or IL-2 (0) (B) (induced peritonealmacrophages with IL-2) for 24 h before being infected with B. abortus. CFU in cytokine-activated ---- -) and control cultures ( ) wereevaluated at several times as indicated. Some SDs of CFU were too low to indicate.
bodies against MrIFN--y completely neutralized the ability ofthe recombinant IFN-y preparation to alter the number of B.abortus isolates (data not shown).
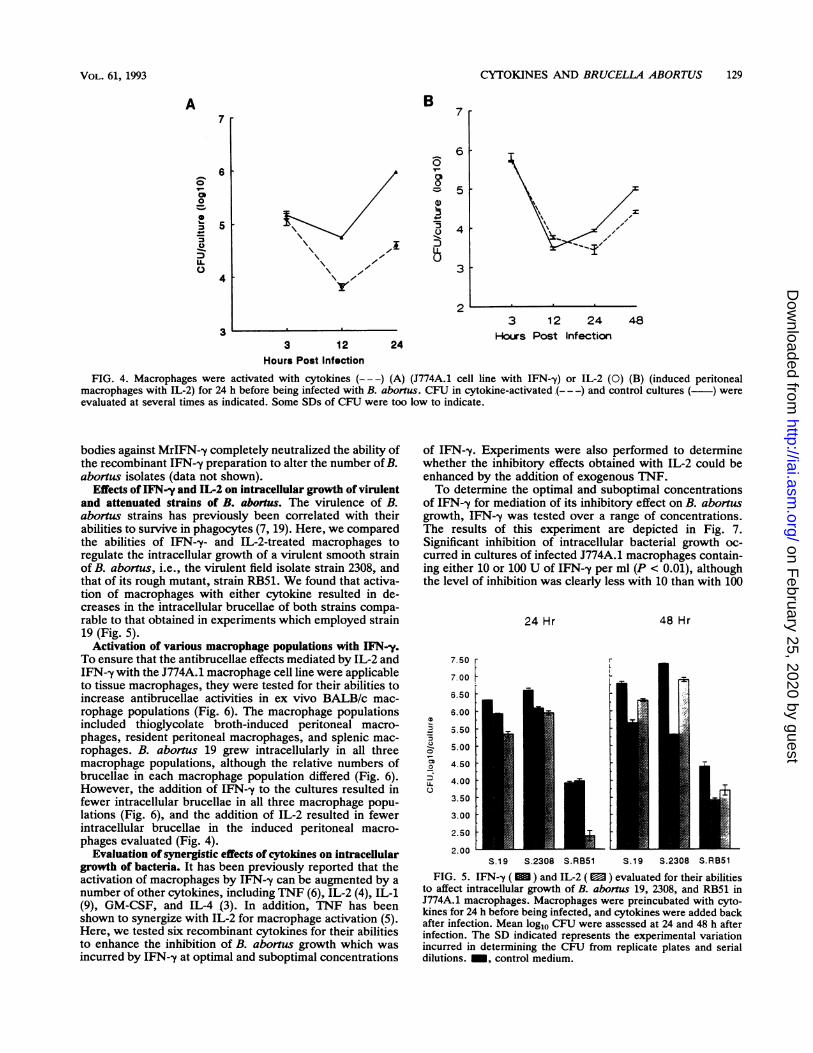
Effects of IFN-y and IL-2 on intracellular growth of virulentand attenuated strains of B. abortus. The virulence of B.abortus strains has previously been correlated with theirabilities to survive in phagocytes (7, 19). Here, we comparedthe abilities of IFN--y- and IL-2-treated macrophages toregulate the intracellular growth of a virulent smooth strainof B. abortus, i.e., the virulent field isolate strain 2308, andthat of its rough mutant, strain RB51. We found that activa-tion of macrophages with either cytokine resulted in de-creases in the intracellular brucellae of both strains compa-rable to that obtained in experiments which employed strain19 (Fig. 5).
Activation of various macrophage populations with IFN-y.To ensure that the antibrucellae effects mediated by IL-2 andIFN-y with the J774A.1 macrophage cell line were applicableto tissue macrophages, they were tested for their abilities toincrease antibrucellae activities in ex vivo BALB/c mac-rophage populations (Fig. 6). The macrophage populationsincluded thioglycolate broth-induced peritoneal macro-phages, resident peritoneal macrophages, and splenic mac-rophages. B. abortus 19 grew intracellularly in all threemacrophage populations, although the relative numbers ofbrucellae in each macrophage population differed (Fig. 6).However, the addition of IFN--y to the cultures resulted infewer intracellular brucellae in all three macrophage popu-lations (Fig. 6), and the addition of IL-2 resulted in fewerintracellular brucellae in the induced peritoneal macro-phages evaluated (Fig. 4).
Evaluation of synergistic effects of cytokines on intracellulargrowth of bacteria. It has been previously reported that theactivation of macrophages by IFN--y can be augmented by anumber of other cytokines, including TNF (6), IL-2 (4), IL-1(9), GM-CSF, and IL-4 (3). In addition, TNF has beenshown to synergize with IL-2 for macrophage activation (5).Here, we tested six recombinant cytokines for their abilitiesto enhance the inhibition of B. abortus growth which wasincurred by IFN-y at optimal and suboptimal concentrations
of IFN--y. Experiments were also performed to determinewhether the inhibitory effects obtained with IL-2 could beenhanced by the addition of exogenous TNF.To determine the optimal and suboptimal concentrations
of IFN--y for mediation of its inhibitory effect on B. abornusgrowth, IFN--y was tested over a range of concentrations.The results of this experiment are depicted in Fig. 7.Significant inhibition of intracellular bacterial growth oc-curred in cultures of infected J774A.1 macrophages contain-ing either 10 or 100 U of IFN--y per ml (P < 0.01), althoughthe level of inhibition was clearly less with 10 than with 100
24 Hr
7.50
7.00
6.50
6.00
_ 5.50u 5.000
48 Hr
Lo 'o1kU4.00
4003.50
3.00
2.50
2.00
S.19 S.2308 S.RB51 S.19 S.2308 S.RB51
FIG. 5. IFN-y ( ) and IL-2 ( ) evaluated for their abilitiesto affect intracellular growth of B. abortus 19, 2308, and RB51 inJ774A.1 macrophages. Macrophages were preincubated with cyto-kines for 24 h before being infected, and cytokines were added backafter infection. Mean log10 CFU were assessed at 24 and 48 h afterinfection. The SD indicated represents the experimental variationincurred in determining the CFU from replicate plates and serialdilutions. _, control medium.
FIG. 6. IFN-y ( ) evaluated for its ability to affect growth ofB. abortus 19 in four different BALB/c macrophage populations.The J774A.1 macrophage cell line (J7), splenic macrophages (SP),and resident (RP) or thioglycolate broth-induced peritoneal macro-
phages (IP) were infected with B. abortus 19. Macrophage popula-tions were incubated for 24 h with IFN-y before being infected, andIFN--y was present throughout the culture period. Mean log10 CFUwere assessed at 24 and 48 h after infection. The SD indicatedrepresents the experimental variation incurred in determining theCFU from replicate plates and serial dilutions. EJ, control medium.
U/ml (Fig. 7). IFN-y at 1 U/ml resulted in slight inhibition.Concentrations below 1 U/ml never significantly decreasedthe growth of B. aborts. Additional experiments showedthat the level of inhibition was not increased when IFN-ywas added at 1,000 U/ml compared with inhibition at 100U/ml (data not shown).
6.50r
6.00
0.5
0)
0.0U-
0.5
4.00
FIG. 7. J774A.1 macrophages cultured with IFN-y at a variety ofconcentrations ranging, from right to left, from 100 (M) to 0.1
( u ) U/ml, both before and after infection with B. abortus 19.Bacterial CFU in the cultures were assessed at 24 and 48 h afterinfection and are represented as mean log10 CFU. The SD indicatedrepresents the experimental variation incurred in determining theCFU from replicate plates and serial dilutions. 1 , IFN--y at 1U/ml; l, IFN at 10 U/ml; FOi, control medium.
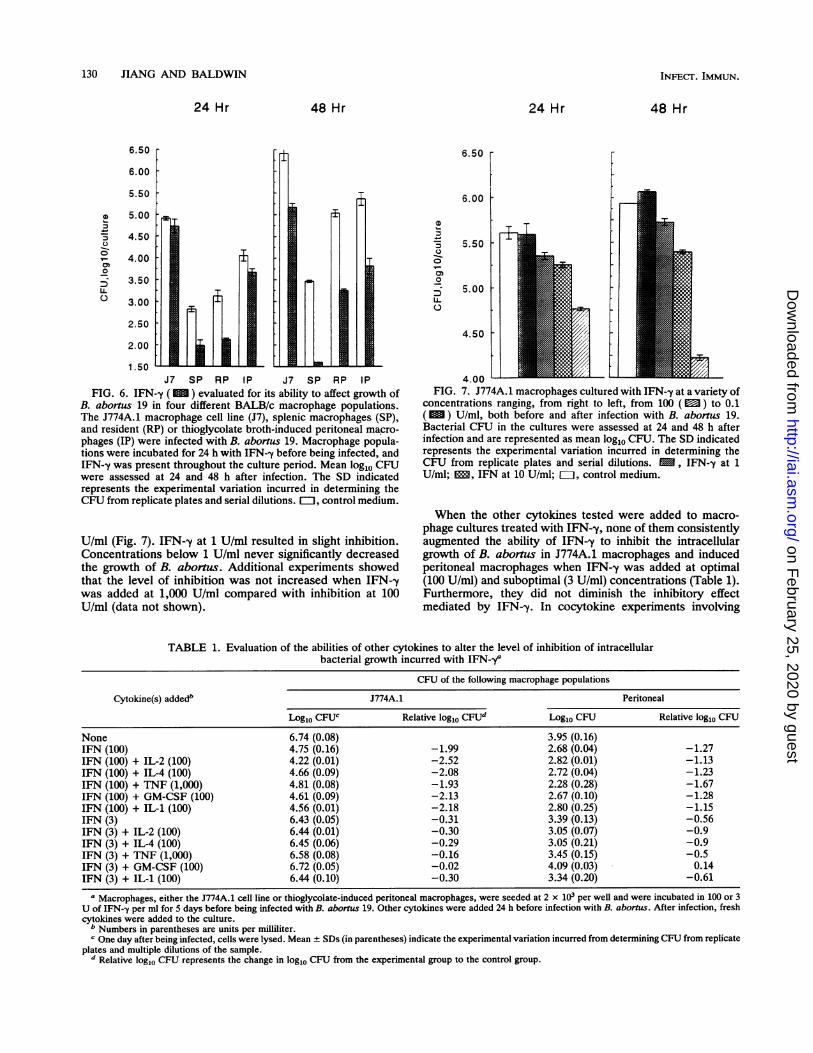
When the other cytokines tested were added to macro-phage cultures treated with IFN--y, none of them consistentlyaugmented the ability of IFN-y to inhibit the intracellulargrowth of B. aborus in J774A.1 macrophages and inducedperitoneal macrophages when IFN-y was added at optimal(100 U/ml) and suboptimal (3 U/ml) concentrations (Table 1).Furthermore, they did not diminish the inhibitory effectmediated by IFN-y. In cocytokine experiments involving
TABLE 1. Evaluation of the abilities of other cytokines to alter the level of inhibition of intracellularbacterial growth incurred with IFN-ya
a Macrophages, either the J774A.1 cell line or thioglycolate-induced peritoneal macrophages, were seeded at 2 x 103 per well and were incubated in 100 or 3U of IFN-,y per ml for 5 days before being infected with B. abortus 19. Other cytokines were added 24 h before infection with B. abortus. After infection, freshcytokines were added to the culture.
b Numbers in parentheses are units per milliliter.c One day after being infected, cells were lysed. Mean + SDs (in parentheses) indicate the experimental variation incurred from determining CFU from replicate
plates and multiple dilutions of the sample.d Relative log10 CFU represents the change in log10 CFU from the experimental group to the control group.
4.00Med -96 -72 -48 -24 0Hr Pro-incubation with IFN
B
D
-9
0tco
IL
7.00 [
6.50
6.00
5.50
5.00Med -24 +2 +24
Time (Hr) IFN first added
C
7.00
00
U-0
6.50
6.00
5.50
S.00 LI.I
+
24 Hr 48 Hr
Hours Post Infection
FIG. 8. J774A.1 macrophages incubated for various amounts oftime with IFN-y. The mean loglo CFU is indicated for eachexperiment. The SD indicated for each experiment represents the
IL-2, the IL-2 was used at 100 U/ml, since the inhibition ofB. abortus growth mediated by it alone was most often lessthan that mediated by optimal concentrations of IFN--y,suggesting that maximal activation of the macrophages hadnot occurred with IL-2 alone. The addition of TNF alongwith IL-2 to macrophage cultures did not result in inhibitionin the number of intracellular bacteria greater than thatobtained when IL-2 alone was added (data not shown).
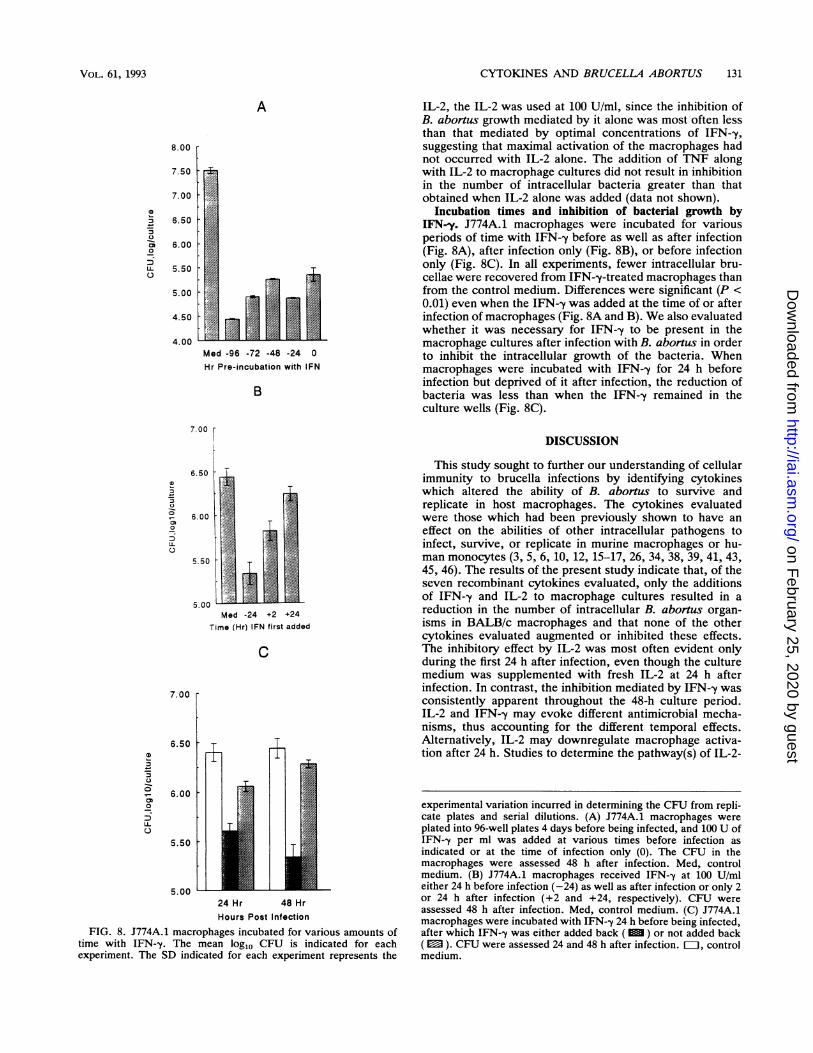
Incubation times and inhibition of bacterial growth byIFN-y. J774A.1 macrophages were incubated for variousperiods of time with IFN-y before as well as after infection(Fig. 8A), after infection only (Fig. 8B), or before infectiononly (Fig. 8C). In all experiments, fewer intracellular bru-cellae were recovered from IFN--y-treated macrophages thanfrom the control medium. Differences were significant (P <0.01) even when the IFN--y was added at the time of or afterinfection of macrophages (Fig. 8A and B). We also evaluatedwhether it was necessary for IFN-y to be present in themacrophage cultures after infection with B. abortus in orderto inhibit the intracellular growth of the bacteria. Whenmacrophages were incubated with IFN-y for 24 h beforeinfection but deprived of it after infection, the reduction ofbacteria was less than when the IFN--y remained in theculture wells (Fig. 8C).
DISCUSSION
This study sought to further our understanding of cellularimmunity to brucella infections by identifying cytokineswhich altered the ability of B. abortus to survive andreplicate in host macrophages. The cytokines evaluatedwere those which had been previously shown to have aneffect on the abilities of other intracellular pathogens toinfect, survive, or replicate in murine macrophages or hu-man monocytes (3, 5, 6, 10, 12, 15-17, 26, 34, 38, 39, 41, 43,45, 46). The results of the present study indicate that, of theseven recombinant cytokines evaluated, only the additionsof IFN--y and IL-2 to macrophage cultures resulted in areduction in the number of intracellular B. abortus organ-isms in BALB/c macrophages and that none of the othercytokines evaluated augmented or inhibited these effects.The inhibitory effect by IL-2 was most often evident onlyduring the first 24 h after infection, even though the culturemedium was supplemented with fresh IL-2 at 24 h afterinfection. In contrast, the inhibition mediated by IFN--y wasconsistently apparent throughout the 48-h culture period.IL-2 and IFN--y may evoke different antimicrobial mecha-nisms, thus accounting for the different temporal effects.Alternatively, IL-2 may downregulate macrophage activa-tion after 24 h. Studies to determine the pathway(s) of IL-2-
experimental variation incurred in determining the CFU from repli-cate plates and serial dilutions. (A) J774A.1 macrophages wereplated into 96-well plates 4 days before being infected, and 100 U ofIFN--y per ml was added at various times before infection asindicated or at the time of infection only (0). The CFU in themacrophages were assessed 48 h after infection. Med, controlmedium. (B) J774A.1 macrophages received IFN--y at 100 U/mleither 24 h before infection (-24) as well as after infection or only 2or 24 h after infection (+2 and +24, respectively). CFU wereassessed 48 h after infection. Med, control medium. (C) J774A.1macrophages were incubated with IFN-y 24 h before being infected,after which IFN--y was either added back ( M ) or not added back( M ). CFU were assessed 24 and 48 h after infection. EL1, controlmedium.
and IFN-y-mediated antibrucella effects are under way.Although it could be argued that the effects which we haveobserved with IFN-y and IL-2 occur as a result of thecytokines increasing uptakes of antibiotics by macrophages,we feel that this is an unlikely explanation. Recent studieshave shown that the antibrucella effects by IFN--y-activatedmacrophages can be blocked by the addition of superoxidedismutase and catalase (22). Since more brucellae wererecovered from IFN--y-activated macrophage cultures whichhad also received inhibitors of reactive oxygen intermedi-ates, it is unlikely that the IFN--y-mediated inhibition ofbrucella growth is a spurious impression resulting fromeither increased accumulation of antibiotics following IFN-ytreatments or from enhanced growth of brucellae in thetreated macrophages which ultimately resulted in lysis of"overloaded" macrophages. Furthermore, while evalua-tions performed with the J774A.1 cell line are complicatedby the fact that the cells can continue to growth afterinfection, the fact that we observed similar effects withIFN--y and IL-2 on tissue macrophages indicates that theresults are not due to an erroneous impression resulting fromthe inhibition of growth of J774A.1 cells by the cytokines,thereby providing fewer target cells for infection.While IFN--y alone has been previously shown to be
sufficient for activation of macrophages for the antimicrobialactivities of a number of intracellular parasites (15-17, 34,41), there is less precedent for IL-2 to mediate this role. IL-2has been reported, however, to act as a cofactor for mac-rophage activation (3-5, 45). In our study, no other factorswere required for the induction of brucellastatic activity.Similarly, IFN--y-mediated inhibition of B. abortus growthwas not augmented by cytokines previously shown else-where to cooperate with IFN--y for induction of macrophageantimicrobial or tumoricidal activity (3, 6, 9, 14, 20, 26). Theunique nature of our results may be due to the source ofmacrophages (i.e., a different mouse strain), or they mayindicate that IL-2 and IFN--y limit intracellular growth of B.abortus by mechanisms which differ from those operative inthe studies reported. In addition, brucella has LPS in its cellenvelope and, therefore, may alter the requirements ofcytokine-induced macrophage activation. Enterobacten-aceae LPS is a potent cofactor for activation of macro-phages. While B. abortus LPS differs significantly fromEnterobacteriaceae LPS in terms of biological activity (29,30), we cannot discount its contribution to macrophageactivation in our system.
In this study, we employed antibody-opsonized brucellaefor two reasons. First, opsonization facilitated the infectionof macrophages, and second, in vivo infection with brucellastimulates antibody production. Thus, it is likely that mac-rophages are phagocytosing opsonized brucellae in naturalinfections following the expression of acquired immunity.While IFN-y has been previously reported to inhibit immu-noglobulin G-mediated phagocytosis (40), the mechanism ofantimicrobial activity measured here is not inhibition ofinfection. Macrophages either cultured in tissue culturemedium alone or precultured for 24 h in medium whichcontained 100 U of IFN--y per ml had equal numbers ofintracellular brucellae following the infection period. Fur-thermore, we found that IFN--y was effective at inhibiting theintracellular growth of B. abortus even when it was added tomacrophage cultures only after infection and that IFN--yactivates macrophages to inhibit replication of brucellaefollowing infection of macrophages with nonopsonized bru-cellae as well (22a).The results of our study indicate that IFN--y is likely to be
an important cytokine in controlling B. abortus infections,since it was effective at inducing antimicrobial activity in allthree ex vivo macrophage populations tested and againstthree strains of B. abortus which differ substantially invirulence (see also reference 23). Similar results have beenobtained with macrophages from a number of other strains ofmice (22a). However, it was necessary to add fresh IFN--y tothe culture system every 24 h in order to maintain macro-phage brucellastatic activity. Thus, B. abortus infectionalone cannot sustain the IFN--y-induced activation. It hasbeen shown elsewhere that while extracellular bacteriamaintain the tumoricidal activity of macrophages, intracel-lular bacteria do not (25).There is evidence from other systems that IFN--y can
protect nonphagocytic cells from microbial parasitism, asshown for infection of fibroblasts by Toxoplasma gondiithrough induction of indolamine 2,3-dioxygenase activity(36). Since B. abortus is capable of infecting nonphagocyticcells (13), IFN--y, as well as other cytokines which werefound here to be inactive with regard to macrophages, mayplay a role in immunity to B. abortus by limiting parasitismof nonphagocytic cells. IL-1, for instance, may play such arole. While IL-la- has previously been shown to be involvedin macrophage activation (9, 37) and to have an importantrole in protection against B. abortus infection in vivo (48),our results indicate that its role is not likely to be throughdirect activation, or through augmentation of IFN--y-medi-ated activation, of macrophages for inhibition of intracellulargrowth of B. abortus. However, it must be recognized thatdifferences may occur among macrophages from variousstrains of mice. In our current studies, we are evaluating therole of cytokines in the various stages of B. abortus infec-tions in vivo and the mechanisms by which macrophagesmediate antibrucella activities.
ACKNOWLEDGMENTS
This work was supported in part by a grant from the Office ofResearch and Graduate Studies of The Ohio State University.We thank Kelly Taggart, Roger Buzatu, and Rita Benson for
technical assistance.
REFERENCES
1. Araya, L. N., P. H. Elzer, G. E. Rowe, F. M. Enright, and A. J.Winter. 1989. Temporal development of protective cell-medi-ated and humoral immunity in BALB/c mice infected withBrucella abortus. J. Immunol. 143:3330-3337.
2. Araya, L. N., and A. J. Winter. 1990. Comparative protection ofmice against virulent and attenuated strains of Brucella abortusby passive transfer of immune T cells or serum. Infect. Immun.58:254-256.
3. Belosevic, M., C. E. Davis, M. S. Meltzer, and C. A. Nacy. 1988.Regulation of activated macrophage antimicrobial activities.Identification of lymphokines that cooperate with IFN--yfor induction of resistance to infection. J. Immunol. 141:890-896.
4. Belosevic, M., D. S. Finbloom, M. S. Meltzer, and C. A. Nacy.1990. IL-2: a cofactor for induction of activated macrophageresistance to infection. J. Immunol. 145:831-839.
5. Bermudez, L. E. M., and L. S. Young. 1988. Tumor necrosisfactor, alone or in combination with IL-2, but not IFN--y, isassociated with macrophage killing of Mycobacterium aviumcomplex. J. Immunol. 140:3006-3013.
6. Bogdan, C., H. Moll, W. Solbach, and M. Rollinghoff. 1990.Tumor necrosis factor-a in combination with interferon--y, butnot with interleukin 4, activates murine macrophages for elim-ination of Leishmania major amastigotes. Eur. J. Immunol.
20:1131-1135.7. Braun, W., A. Pomales-Lebron, and W. R. Stinebring. 1958.
Interactions between mononuclear phagocytes and Brucellaabortus strains of different virulence. Proc. Exp. Soc. Biol.Med. 97:393-397.
8. Carlin, J. M., Y. Ozaki, G. I. Bryne, R. R. Brown, and E. C.Borden. 1989. Interferons and indoleamine 2,3-dioxygenase:role in antimicrobial and antitumor effects. Experientia 45:535-541.
9. Chen, L., Y. Suzuki, and E. F. Wheelock. 1987. Interferon-ysynergizes with tumor necrosis factor and with interleukin-1 andrequires the presence of both monokines to induce antitumorcytotoxic activity in macrophages. J. Immunol. 12:4096-4101.
10. Denis, M. 1991. Modulation of Mycobacterium lepraemuriumgrowth in murine macrophages: beneficial effect of tumor ne-crosis factor alpha and granulocyte-macrophage colony-stimu-lating factor. Infect. Immun. 59:705-707.
11. Denis, M. 1991. Growth of Mycobacterium avium in humanmonocytes: identification of cytokines which reduce and en-hance intracellular microbial growth. Eur. J. Immunol. 21:391-395.
12. Denis, M., and E. 0. Gregg. 1990. Recombinant tumour necrosisfactor-a decreases whereas recombinant interleukin-6 increasesgrowth of a virulent strain of Mycobactenum avium in humanmacrophages. Immunology 71:139-141.
13. Detilleux, P. G., B. L. Deyoe, and N. F. Cheville. 1990. Penetra-tion and intracellular growth of Brucella abortus in nonphago-cytic cells in vitro. Infect. Immun. 58:2320-2328.
14. Ding, A. H., C. F. Nathan, and D. J. Stueher. 1988. Release ofreactive nitrogen intermediates and reactive oxygen intermedi-ates from mouse peritoneal macrophages. J. Immunol. 141:2407-2412.
15. Douvas, G. S., D. L. Looker, A. E. Vatter, and A. J. Crowle.1985. -y-Interferon activates human macrophages to becometumoricidal and leishmanicidal but enhances replication of mac-rophage-associated mycobacteria. Infect. Immun. 50:1-8.
16. Flesch, I. E. A., and S. H. E. Kaufmann. 1988. Attempts tocharacterize the mechanisms involved in mycobacterial growthinhibition by gamma-interferon-activated bone marrow macro-phages. Infect. Immun. 56:1464-1469.
17. Green, S. J., R. M. Crawford, J. T. Hockmeyer, M. S. Meltzer,and C. A. Nacy. 1990. Leishmania major amastigotes initiate theL-arginine-dependent killing mechanism in IFN--y-stimulatedmacrophages by induction of tumor necrosis factor-a. J. Immu-nol. 145:4290-4297.
18. Hailing, S. M., P. G. Detilleux, F. M. Tatum, B. A. Judge, andJ. E. Mayfield. 1991. Deletion of the BCSP31 gene of Brucellaabortus by replacement. Infect. Immun. 59:3863-3868.
19. Harmon, B. G., L. G. Adams, and M. Frey. 1988. Survival ofrough and smooth strains of Brucella abortus in bovine mam-mary gland macrophages. Am. J. Vet. Res. 49:1092-1097.
20. Heidenreich, S., M. Weyers, J.-H. Gong, H. Sprenger, M. Nain,and D. Gemsa. 1988. Potentiation of lymphokine-induced mac-rophage activation by tumor necrosis factor-a. J. Immunol.140:1511-1518.
21. Ho, M., and C. Cheers. 1982. Resistance and susceptibility ofmice to bacterial infection. IV. Genetic and cellular basis ofresistance to chronic infection with Brucella abortus. J. Infect.Dis. 146:381-387.
22. Jiang, X., and C. L. Baldwin. Inhibition of growth of Brucellaabortus by noncytokine-activated and IFN-y-activated macro-phages is mediated by reactive oxygen intermediates but not bynitric oxide, submitted for publication.
22a.Jiang, X., and C. L. Baldwin. Unpublished data.23. Jones, S. M., and A. J. Winter. 1992. Survival of virulent and
attenuated strains of Brucella abortus in normal and gammainterferon-activated murine peritoneal macrophages. Infect. Im-mun. 60:3011-3017.
24. Keller, R., R. Keist, and K. Frei. 1990. Lymphokines andbacteria that induce tumoricidal activity, trigger a differentsecretory response in macrophages. Eur. J. Immunol. 20:695-698.
25. Keller, R., R. Keist, P. Van der Meide, P. Groseurth, M. Aguet,
and T. P. Leist. 1987. Induction, maintenance, and reinductionof tumoricidal activity in bone marrow-derived mononuclearphagocytes by Corynebacterium parvum. J. Immunol. 138:2366-2371.
26. Liew, F. Y., L. Yun, and S. Millott. 1990. Tumor necrosisfactor-a synergizes with IFN--y in mediating killing of Leishma-nia major through the induction of nitric oxide. J. Immunol.145:4306-4310.
27. McGee, Z. A., G. L. Gorby, and W. S. Updike. 1989. The use ofneutrophils, macrophages and organ cultures to assess thepenetration of human cells by antimicrobials. Prog. Drug. Res.33:83-92.
28. Montaraz, J. A., and A. J. Winter. 1986. Comparison of livingand nonliving vaccines for Brucella abortus in BALB/c mice.Infect. Immun. 53:245-251.
29. Moreno, E., D. Borowiak, and H. Mayer. 1987. Brucella lipo-polysaccharides and polysaccharides. Ann. Inst. Pasteur Micro-biol. 38:102-105.
30. Moriyon, I., C. Gamazo, and R. Diaz. 1987. Properties of theouter membrane of Brucella. Ann. Inst. Pasteur Microbiol.138:89-91.
31. Nathan, C. F., H. W. Murray, M. E. Wiebe, and B. Y. Rubin.1983. Identification of interferon--y as the lymphokine thatactivates human macrophage oxidative metabolism and antimi-crobial activity. J. Exp. Med. 158:670-689.
32. Nicoletti, P. 1980. The epidemiology of bovine brucellosis. Adv.Vet. Sci. Comp. Med. 24:69-98.
33. Nicoletti, P. 1992. The control of brucellosis-a veterinaryresponsibility. Saudi Med. J. 13:10-13.
34. Passwell, J. H., R. Shor, and J. Shoham. 1986. The enhancingeffect of interferon ,B and -y on the killing of Leishmania tropicamajor in human mononuclear phagocytes in vitro. J. Immunol.136:3062-3066.
35. Pavlov, H., M. Hogarth, I. F. C. McKenzie, and C. Cheers. 1982.In vivo and in vitro effects of monoclonal antibody to Lyantigens on immunity to infection. Cell. Immunol. 71:127-138.
36. Pfefferkorn, E. R. 1984. Interferon-y blocks the growth ofToxoplasma gondii in human fibroblasts by inducing the hostcells to degrade tryptophan. Proc. Natl. Acad. Sci. USA 81:908-912.
37. Philip, R., and L. B. Epstein. 1986. Tumor necrosis factor asimmunomodulator and mediator of monocyte cytotoxicity in-duced by itself, -y-interferon and interleukin-1. Nature (London)323:86-89.
38. Portoy, D. A., R. D. Schreiber, P. Cannelly, and L. G. Tilney.1989. -y-Interferon limits access of Listeria monocytogenes tothe macrophage cytoplasm. J. Exp. Med. 170:2141-2146.
39. Reed, S. G., C. F. Nathan, D. L. Pihi, P. Rodricks, K. Shane-beck, P. J. Conlon, and K. H. Grabstein. 1987. Recombinantgranulocyte macrophage-colony stimulating factor activatesmacrophages to inhibit Trypanosoma cruzi and release hydro-gen peroxide. Comparison with interferon-y. J. Exp. Med.166:1734-1746.
40. Rollag, H., M. Degre, and G. Sonnenfeld. 1984. Effects ofinterferon-al/ and interferon--y preparations on phagocytosisby mouse peritoneal macrophages. Scand. J. Immunol. 20:149-155.
41. Rook, G. A. W., J. Steele, M. Ainsworth, and B. R. Champion.1986. Activation of macrophages to inhibit proliferation ofMycobacterium tuberculosis: comparison of the effects of re-combinant y-interferon on human monocytes and murine peri-toneal macrophages. Immunology 59:333-338.
42. Schurig, G. G., R. M. Roop, D. Buhrman, S. Boyle, T. Bagchi,and N. Sriranganathan. 1991. Biological properties of RB51, astable, 0-chain deficient mutant of Brucella abortus. Vet.Microbiol. 2:171-188.
43. Shiratsuchi, H., J. L. Johnson, and J. J. Ellner. 1991. Bidirec-tional effects of cytokines on the growth of Mycobacteriumavium within human monocytes. J. Immunol. 146:3165-3170.
44. Stuehr, D. J., and M. A. Marietta. 1987. Induction of nitrite/nitrate synthesis in murine macrophages by BCG infection,lymphokines, or interferon-g. J. Immunol. 139:518-525.
45. Wahl, S. M., N. McCartney-Francis, D. A. Hunt, P. D. Smith,
L. M. Wahl, and I. M. Katona. 1988. Monocyte interleukin 2receptor gene expression and interleukin 2 augmentation ofmicrobicidal activity. J. Immunol. 139:1342-1347.
46. Weiser, W. Y., A. Van Niel, S. C. Clark, J. R. David, and H. G.Renold. 1987. Recombinant human granulocyte macrophage-col-ony stimulating factor activates killing of L. donovani by human
INFECT. IMMUN.
monocyte-derived macrophages. J. Exp. Med. 166:1436-1446.47. Young, E. J. 1983. Human brucellosis. Rev. Infect. Dis. 5:821-
842.48. Zhan, Y. F., E. R. Stanley, and C. Cheers. 1991. Prophylaxis or
treatment of experimental brucellosis with interleukin-1. Infect.Immun. 59:1790-1794.